Genetic Diversity of Field Pennycress (Thlaspi arvense) Reveals Untapped Variability and Paths Toward Selection for Domestication

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Isolation
2.2. Genotyping and SNP Identification
2.3. Analysis of Genetic Structure
3. Results
3.1. Germplasm and Genotypic Data
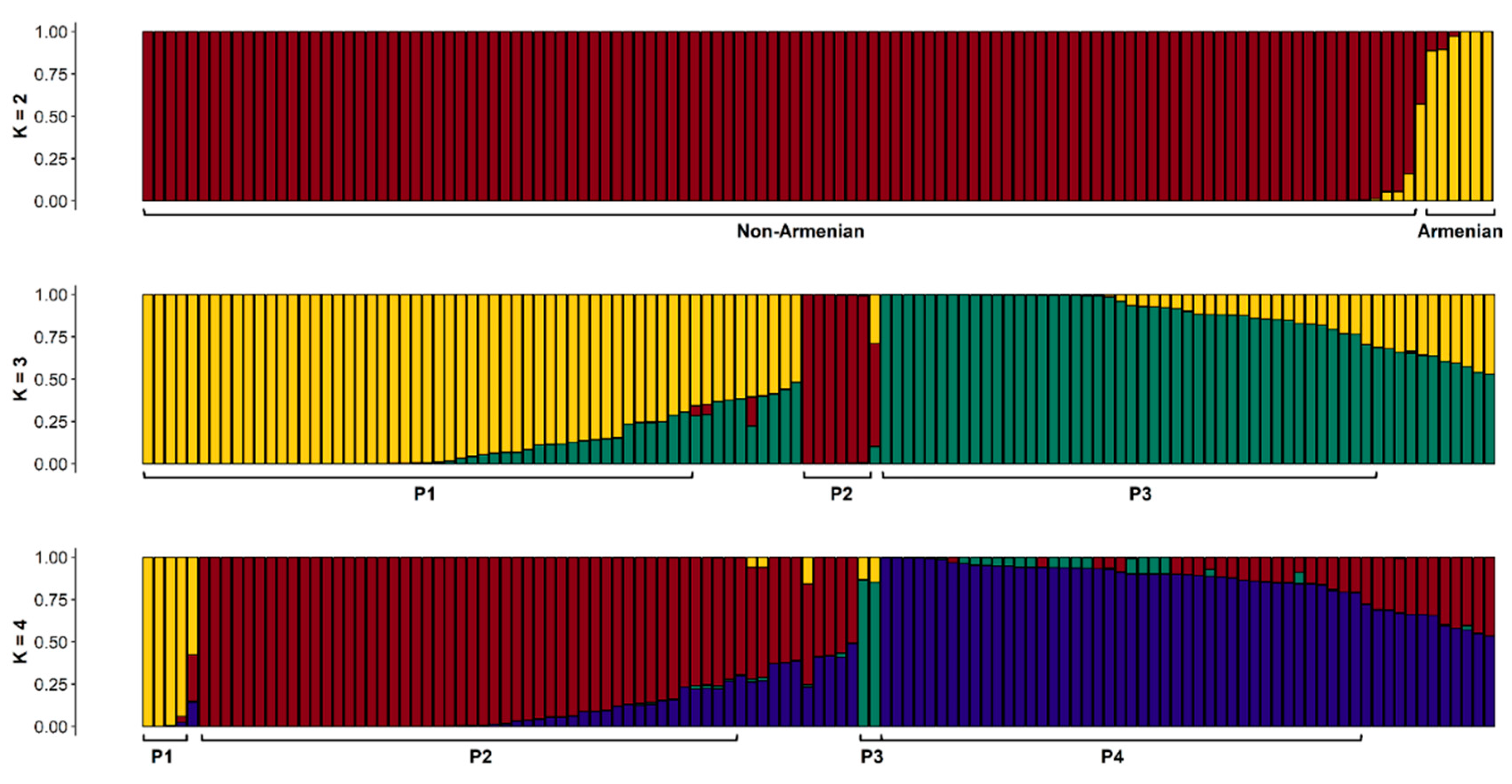
3.2. Bayesian Clustering (STRUCTURE) Analysis
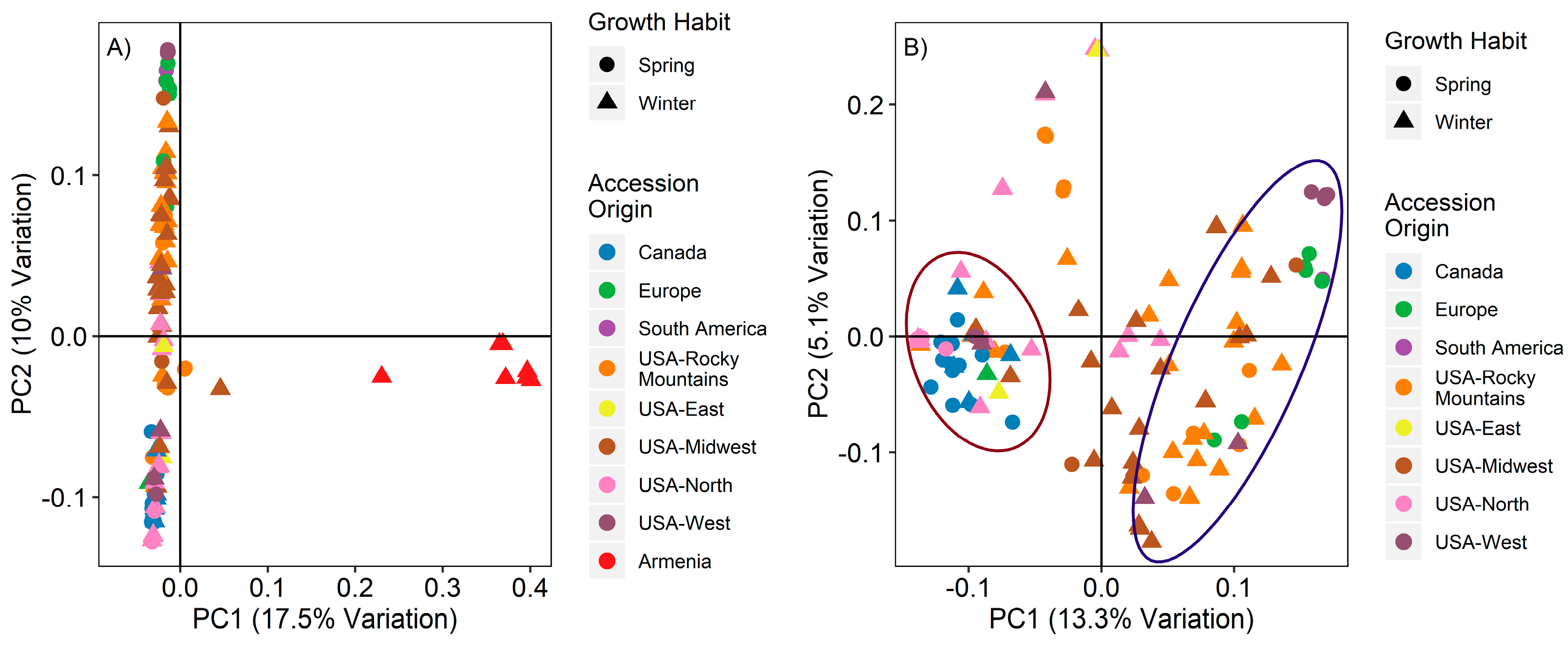
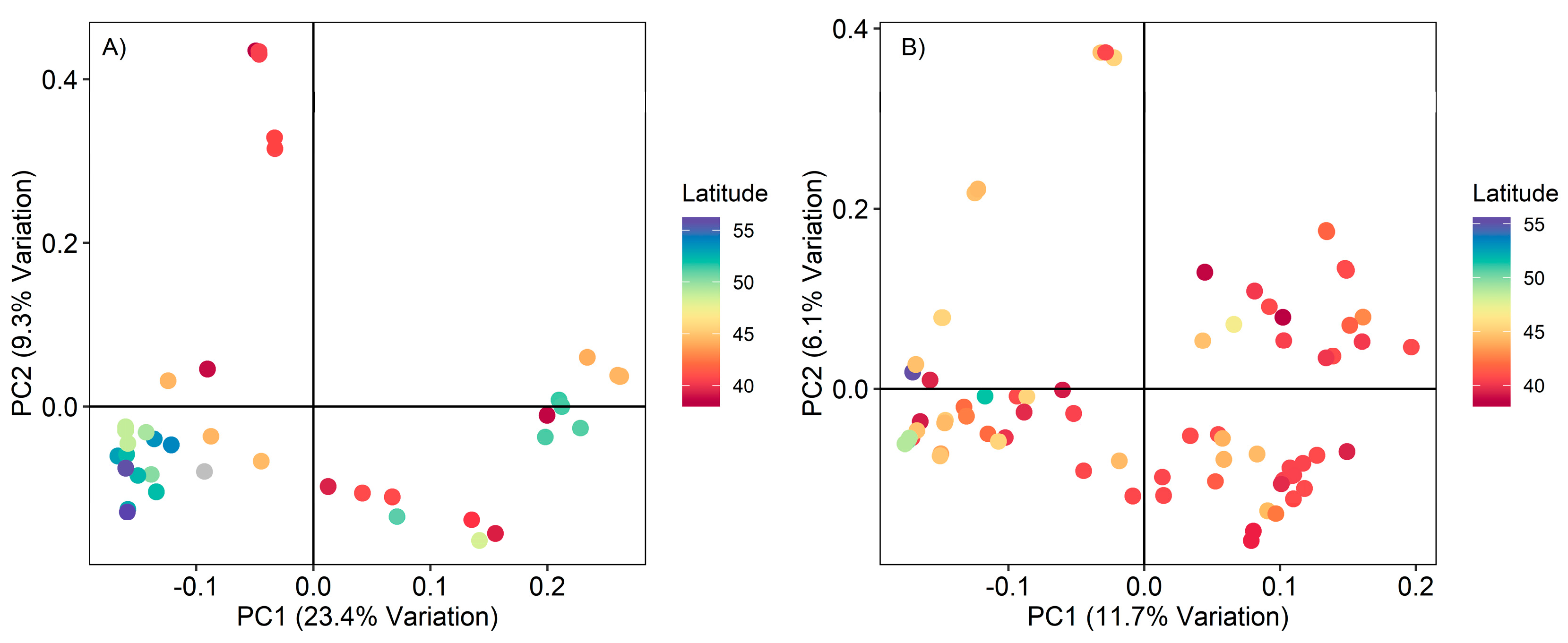
3.3. Principal Component Analysis
3.4. Population Genetic Diversity and Pairwise Comparison of Genetic Distances
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sedbrook, J.C.; Phippen, W.B.; Marks, M.D. New approaches to facilitate rapid domestication of a wild plant to an oilseed crop: Example pennycress (Thlaspi arvense L.). Plant Sci. 2014, 227, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Chopra, R.; Johnson, E.B.; Daniels, E.; McGinn, M.; Dorn, K.M.; Esfahanian, M.; Folstad, N.; Amundson, K.; Altendorf, K.; Betts, K.; et al. Translational genomics using Arabidopsis as a model enables the characterization of pennycress genes through forward and reverse genetics. Plant J. 2018, 96, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Boateng, A.A.; Mullen, C.A.; Goldberg, N.M. Producing stable pyrolysis liquids from the oil-seed presscakes of mustard family plants: Pennycress (Thlaspi arvense L.) and camelina (Camelina sativa). Energ. Fuel. 2010, 24, 6624–6632. [Google Scholar] [CrossRef]
- Johnson, G.A.; Wells, M.S.; Anderson, K.; Gesch, R.W.; Forcella, F.; Wyse, D.L. Yield tradeoffs and nitrogen between pennycress, camelina, and soybean in relay- and double-crop systems. Agron. J. 2017, 109, 2128–2135. [Google Scholar] [CrossRef]
- Johnson, G.A.; Kantar, M.B.; Betts, K.J.; Wyse, D.L. Field pennycress production and weed control in a double crop system with soybean in minnesota. Agron. J. 2015, 107, 532–540. [Google Scholar] [CrossRef]
- Warwick, S.I.; Francis, A.; Susko, D.J. The biology of Canadian weeds. Thlaspi arvense L. (updated). Can. J. Plant Sci. 2002, 82, 803–823. [Google Scholar] [CrossRef]
- Weyers, S.; Thom, M.; Forcella, F.; Eberle, C.; Matthees, H.; Gesch, R.; Ott, M.; Feyereisen, G.; Strock, J.; Wyse, D. Reduced Potential for Nitrogen Loss in Cover Crop–Soybean Relay Systems in a Cold Climate. J. Environ. Qual. 2019, 48, 660. [Google Scholar] [CrossRef]
- Eberle, C.A.; Thom, M.D.; Nemec, K.T.; Forcella, F.; Lundgren, J.G.; Gesch, R.W.; Riedell, W.E.; Papiernik, S.K.; Wagner, A.; Peterson, D.H.; et al. Using pennycress, camelina, and canola cash cover crops to provision pollinators. Ind. Crops Prod. 2015, 75, 20–25. [Google Scholar] [CrossRef]
- Phippen, W.B.; Phippen, M.E. Soybean seed yield and quality as a response to field pennycress residue. Crop Sci. 2012, 52, 2767–2773. [Google Scholar] [CrossRef]
- Fan, J.; Shonnard, D.R.; Kalnes, T.N.; Johnsen, P.B.; Rao, S. A life cycle assessment of pennycress (Thlaspi arvense L.)—Derived jet fuel and diesel. Biomass Bioenergy 2013, 55, 87–100. [Google Scholar] [CrossRef]
- Moser, B.R.; Shah, S.N.; Winkler-Moser, J.K.; Vaughn, S.F.; Evangelista, R.L. Composition and physical properties of cress (Lepidium sativum L.) and field pennycress (Thlaspi arvense L.) oils. Ind. Crops Prod. 2009, 30, 199–205. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel from alternative oilseed feedstocks: Camelina and field pennycress. Biofuels 2012, 3, 193–209. [Google Scholar] [CrossRef]
- McGinn, M.; Phippen, W.B.; Chopra, R.; Bansal, S.; Jarvis, B.A.; Phippen, M.E.; Dorn, K.M.; Esfahanian, M.; Nazarenus, T.J.; Cahoon, E.B.; et al. Molecular tools enabling pennycress (Thlaspi arvense) as a model plant and oilseed cash cover crop. Plant Biotechnol. J. 2019, 17, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Chopra, R.; Johnson, E.B.; Emenecker, R.; Cahoon, E.B.; Lyons, J.; Kliebenstein, D.J.; Daniels, E.; Dorn, K.M.; Esfahanian, M.; Folstad, N.; et al. Progress toward the identification and stacking of crucial domestication traits in pennycress. bioRxiv 2019, 609990. [Google Scholar]
- Claver, A.; Rey, R.; López, M.V.; Picorel, R.; Alfonso, M. Identification of target genes and processes involved in erucic acid accumulation during seed development in the biodiesel feedstock Pennycress (Thlaspi arvense L.). J. Plant Physiol. 2017, 208, 7–16. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dorn, K.M.; Fankhauser, J.D.; Wyse, D.L.; Marks, M.D. A draft genome of field pennycress (Thlaspi arvense) provides tools for the domestication of a new winter biofuel crop. DNA Res. 2015, 22, 121–131. [Google Scholar] [CrossRef]
- Dorn, K.M.; Fankhauser, J.D.; Wyse, D.L.; Marks, M.D. De novo assembly of the pennycress (Thlaspi arvense) transcriptome provides tools for the development of a winter cover crop and biodiesel feedstock. Plant J. 2013, 75, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Vollmann, J.; Grausgruber, H.; Stift, G.; Dryzhyruk, V.; Lelley, T. Genetic diversity in camelina germplasm as revealed by seed quality characteristics and RAPD polymorphism. Plant Breed. 2005, 124, 446–453. [Google Scholar] [CrossRef]
- Luo, Z.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.; Fahlgren, N.; Abdel-Haleem, H. Genetic Diversity and Population Structure of a Camelina sativa Spring Panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef]
- Altendorf, K.; Isbell, T.; Wyse, D.L.; Anderson, J.A. Significant variation for seed oil content, fatty acid profile, and seed weight in natural populations of field pennycress (Thlaspi arvense L.). Ind. Crops Prod. 2019, 129, 261–268. [Google Scholar] [CrossRef]
- Dorn, K.M.; Johnson, E.B.; Daniels, E.C.; Wyse, D.L.; Marks, M.D. Spring flowering habit in field pennycress (Thlaspi arvense) has arisen multiple independent times. Plant Direct 2018, 2, e00097. [Google Scholar] [CrossRef]
- An, M.; Zeng, L.; Zhang, T.; Zhong, Y. Phylogeography of Thlaspi arvense (Brassicaceae) in China Inferred from Chloroplast and Nuclear DNA Sequences and Ecological Niche Modeling. Int. J. Mol. Sci. 2015, 16, 13339–13355. [Google Scholar] [CrossRef] [PubMed]
- Chopra, R.; Folstad, N.; Lyons, J.; Ulmasov, T.; Gallaher, C.; Sullivan, L.; McGovern, A.; Mitacek, R.; Frels, K.; Altendorf, K.; et al. The adaptable use of Brassica NIRS calibration equations to identify pennycress variants to facilitate the rapid domestication of a new winter oilseed crop. Ind. Crops Prod. 2019, 128, 55–61. [Google Scholar] [CrossRef]
- Hyten, D.L.; Song, Q.; Zhu, Y.; Choi, I.-Y.; Nelson, R.L.; Costa, J.M.; Specht, J.E.; Shoemaker, R.C.; Cregan, P.B. Impacts of genetic bottlenecks on soybean genome diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 16666–16671. [Google Scholar] [CrossRef] [PubMed]
- Kilian, B.; Martin, W.; Salamini, F. Genetic Diversity, Evolution and Domestication of Wheat and Barley in the Fertile Crescent. In Evolution in Action; Springer: Berlin/Heidelberg, Germany, 2010; pp. 137–166. [Google Scholar]
- Haudry, A.; Cenci, A.; Ravel, C.; Bataillon, T.; Brunel, D.; Poncet, C.; Hochu, I.; Poirier, S.; Santoni, S.; Glémin, S.; et al. Grinding up Wheat: A Massive Loss of Nucleotide Diversity Since Domestication. Mol. Biol. Evol. 2007, 24, 1506–1517. [Google Scholar] [CrossRef]
- Best, K.F.; Intyre, G.I.M. Studies on the Flowering of Thlaspi arvense L. III. The Influence of Vernalization Under Natural and Controlled Conditions. Bot. Gaz. 1976, 137, 121–127. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Ren, S.; Bertels, K.; Al-Ars, Z. Efficient Acceleration of the Pair-HMMs Forward Algorithm for GATK HaplotypeCaller on Graphics Processing Units. Evol. Bioinform. 2018, 14, 117693431876054. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Poets, A.M.; Mohammadi, M.; Seth, K.; Wang, H.; Kono, T.J.Y.; Fang, Z.; Muehlbauer, G.J.; Smith, K.P.; Morrell, P.L. The Effects of Both Recent and Long-Term Selection and Genetic Drift Are Readily Evident in North American Barley Breeding Populations. G3 (Bethesda) 2015, 6, 609–622. [Google Scholar] [CrossRef]
- Wilkinson, S.P.; Davy, S.K. Phylogram: An R package for phylogenetic analysis with nested lists. J. Open Source Softw. 2018, 3, 790. [Google Scholar] [CrossRef]
- Filippi, C.V.; Aguirre, N.; Rivas, J.G.; Zubrzycki, J.; Puebla, A.; Cordes, D.; Moreno, M.V.; Fusari, C.M.; Alvarez, D.; Heinz, R.A.; et al. Population structure and genetic diversity characterization of a sunflower association mapping population using SSR and SNP markers. BMC Plant Biol. 2015, 15, 52. [Google Scholar] [CrossRef]
- Altendorf, K. Characterization of Field Pennycress (Thlaspi arvense L.) Germplasm for Use as a Cover Crop and Biofuel Feedstock. Master’s Thesis, University of Minnesota, St. Paul, MN, USA, 2017. [Google Scholar]
- Alonso-Blanco, C.; Andrade, J.; Becker, C.; Bemm, F.; Bergelson, J.; Borgwardt, K.M.; Cao, J.; Chae, E.; Dezwaan, T.M.; Ding, W.; et al. 1,135 Genomes Reveal the Global Pattern of Polymorphism in Arabidopsis thaliana. Cell 2016, 166, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.D.; Olsen, M.T.; Samaniego, J.A.; Zimmer, E.A.; Gilbert, M.T.P. The population genomic basis of geographic differentiation in North American common ragweed (Ambrosia artemisiifolia L.). Ecol. Evol. 2016, 6, 3760–3771. [Google Scholar] [CrossRef] [PubMed]
- Durka, W.; Bossdorf, O.; Prati, D.; Auge, H. Molecular evidence for multiple introductions of garlic mustard (Alliaria petiolata, Brassicaceae) to North America. Mol. Ecol. 2005, 14, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Kang, J.; He, F.; Qu, L.-J.; Gu, H. The origin of populations of Arabidopsis thaliana in China, based on the chloroplast DNA sequences. BMC Plant Biol. 2010, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.A.; Kiefer, C.; Ehrich, D.; Vogel, J.; Brochmann, C.; Mummenhoff, K. Three times out of Asia Minor: The phylogeography of Arabis alpina L. (Brassicaceae). Mol. Ecol. 2006, 15, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.B.; Schmuths, H.; Schaal, B.A. Native range genetic variation in Arabidopsis thaliana is strongly geographically structured and reflects Pleistocene glacial dynamics. Mol. Ecol. 2007, 17, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.-T.; Lee, J.; Moon, J.-K.; Yoon, M.-S.; Baek, H.-J.; Kang, J.-H.; Kim, T.-S.; Paek, N.-C. Genetic diversity and population structure of Korean soybean landrace [Glycine max (L.) Merr.]. J. Crop Sci. Biotech. 2008, 11, 83–90. [Google Scholar]
- Kiang, Y.T.; Chiang, Y.C.; Kaizuma, N. Genetic diversity in natural populations of wild soybean in Iwate Prefecture, Japan. J. Hered. 1992, 83, 325–329. [Google Scholar] [CrossRef]
- Baker, H.G. The Evolution of Weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Parker, I.M. Founding events in species invasions: Genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 2008, 17, 431–449. [Google Scholar] [CrossRef]
- Bossdorf, O.; Auge, H.; Lafuma, L.; Rogers, W.E.; Siemann, E.; Prati, D. Phenotypic and genetic differentiation between native and introduced plant populations. Oecologia 2005, 144, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rasmusson, D.C.; Phillips, R.L. Plant breeding progress and genetic diversity from de novo variation and elevated epistasis. Crop Sci. 1997, 37, 303–310. [Google Scholar] [CrossRef]
- Glaszmann, J.; Kilian, B.; Upadhyaya, H.; Varshney, R. Accessing genetic diversity for crop improvement. Curr. Opin. Plant Biol. 2010, 13, 167–173. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| . | P1 | P2 | P3 | P4 | Admix |
|---|---|---|---|---|---|
| P1 | 0.043 | ||||
| P2 | 0.32 | 0.072 | |||
| P3 | 0.31 | 0.4 | 0.032 | ||
| P4 | 0.33 | 0.11 | 0.41 | 0.094 | |
| Admix | 0.31 | 0.098 | 0.4 | 0.11 | 0.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frels, K.; Chopra, R.; Dorn, K.M.; Wyse, D.L.; Marks, M.D.; Anderson, J.A. Genetic Diversity of Field Pennycress (Thlaspi arvense) Reveals Untapped Variability and Paths Toward Selection for Domestication. Agronomy 2019, 9, 302. https://doi.org/10.3390/agronomy9060302
Frels K, Chopra R, Dorn KM, Wyse DL, Marks MD, Anderson JA. Genetic Diversity of Field Pennycress (Thlaspi arvense) Reveals Untapped Variability and Paths Toward Selection for Domestication. Agronomy. 2019; 9(6):302. https://doi.org/10.3390/agronomy9060302
Chicago/Turabian StyleFrels, Katherine, Ratan Chopra, Kevin M. Dorn, Donald L. Wyse, M. David Marks, and James A. Anderson. 2019. "Genetic Diversity of Field Pennycress (Thlaspi arvense) Reveals Untapped Variability and Paths Toward Selection for Domestication" Agronomy 9, no. 6: 302. https://doi.org/10.3390/agronomy9060302
APA StyleFrels, K., Chopra, R., Dorn, K. M., Wyse, D. L., Marks, M. D., & Anderson, J. A. (2019). Genetic Diversity of Field Pennycress (Thlaspi arvense) Reveals Untapped Variability and Paths Toward Selection for Domestication. Agronomy, 9(6), 302. https://doi.org/10.3390/agronomy9060302