Quinoa: In Perspective of Global Challenges

Abstract
:1. Introduction
2. Historical and Cultural Perspective
3. Climate Resilience
3.1. Salinity Tolerance
3.2. Drought Tolerance
3.3. Response to Temperature
4. Botanical and Agronomic Features
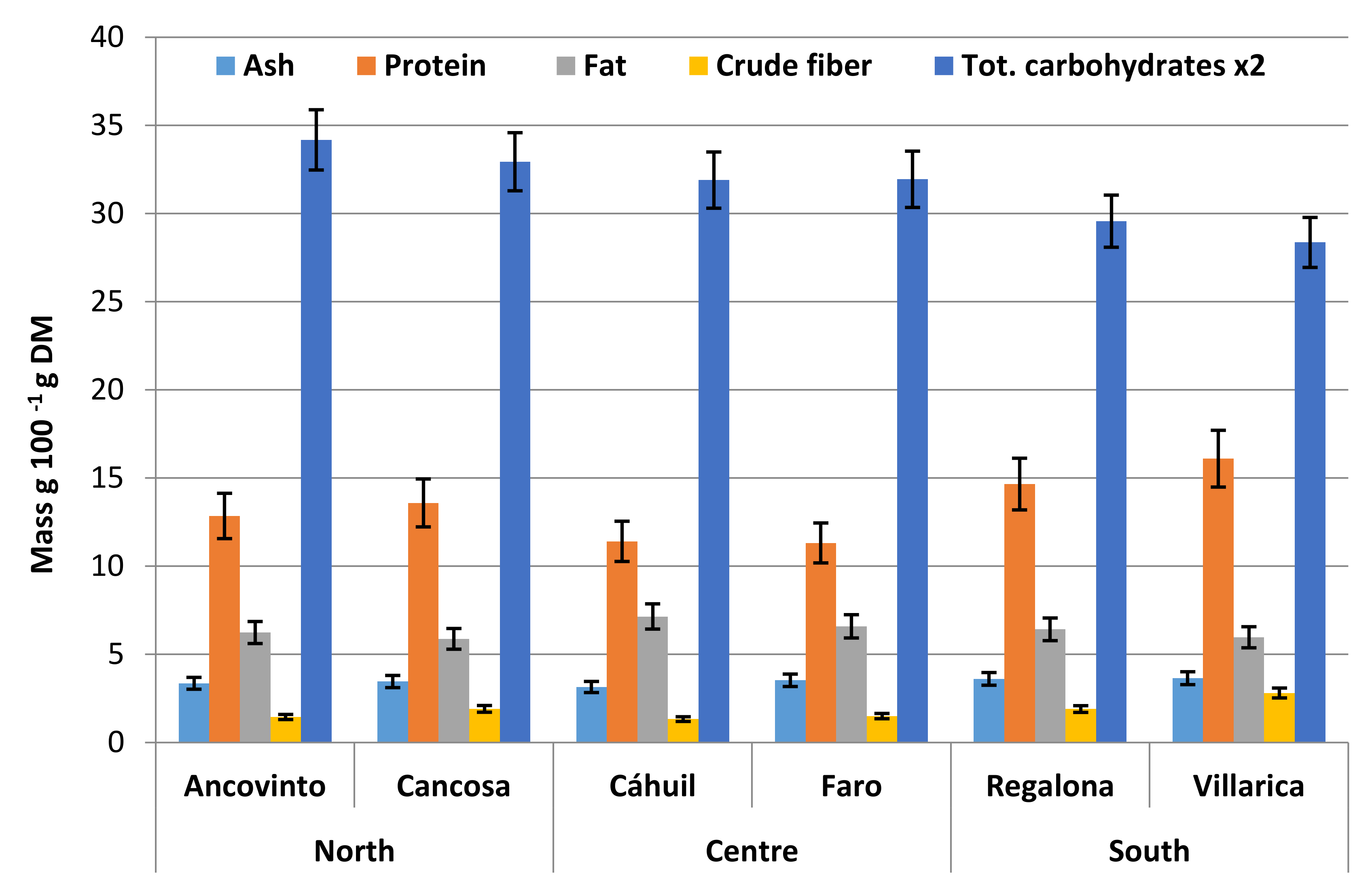
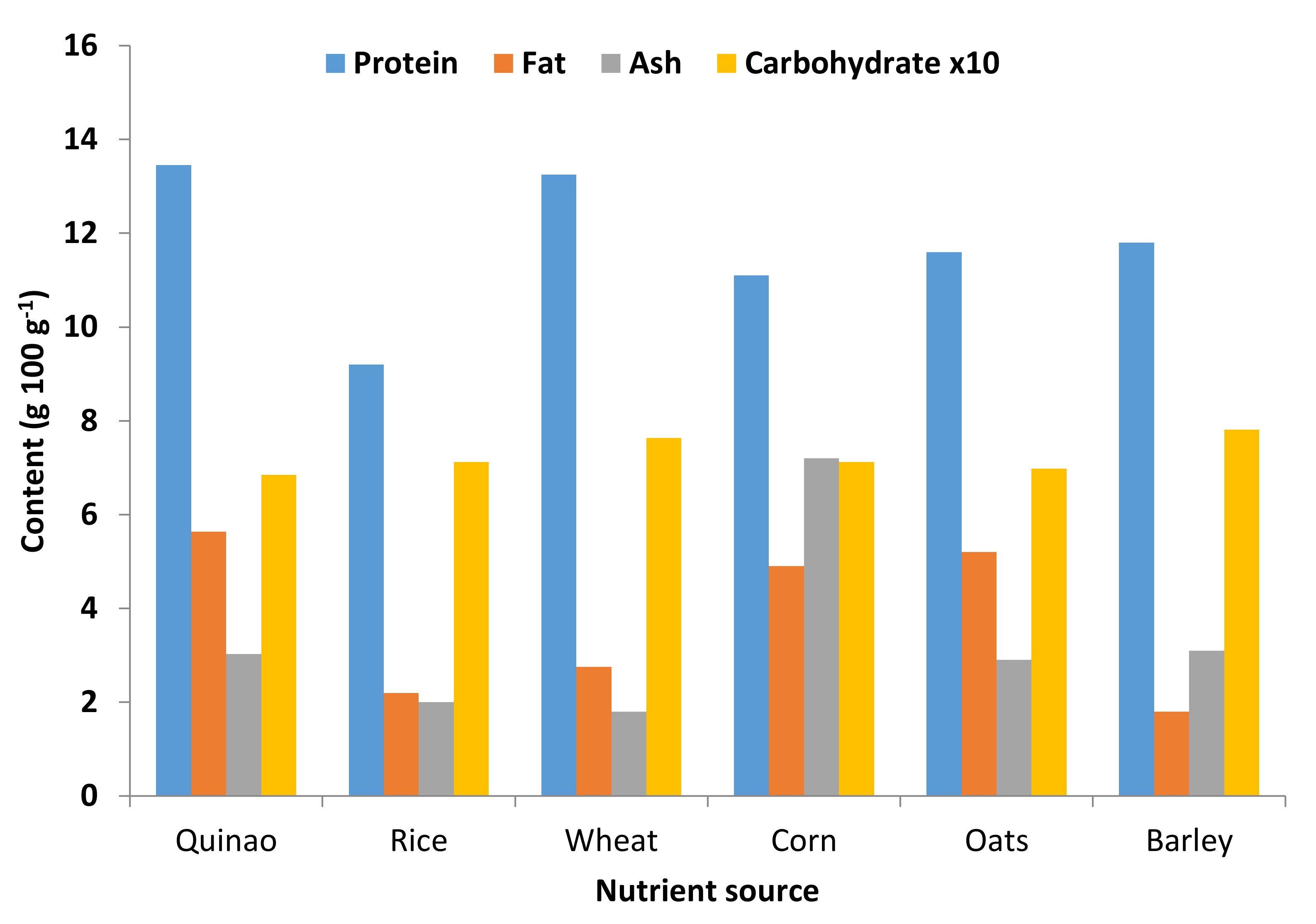
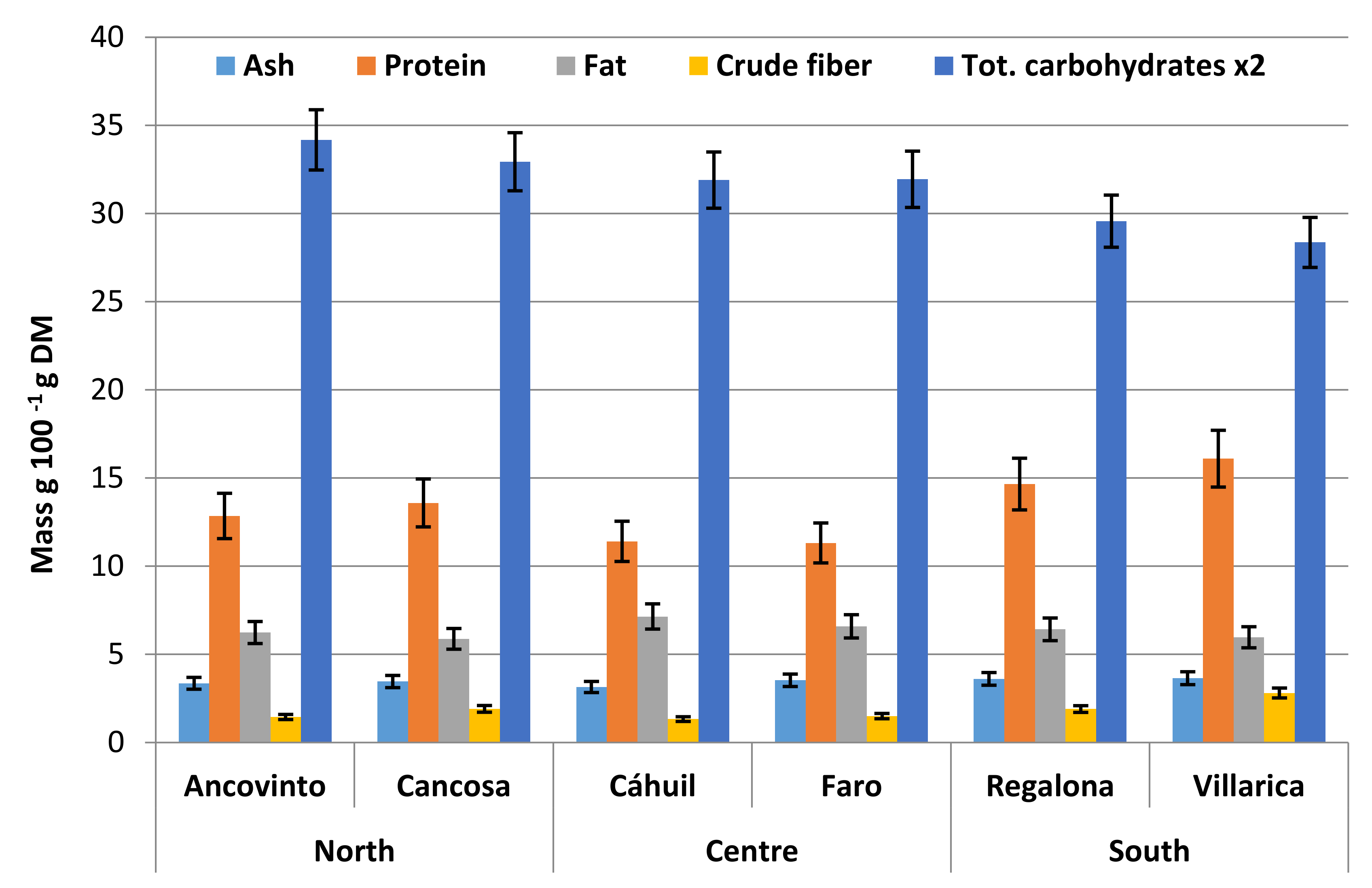
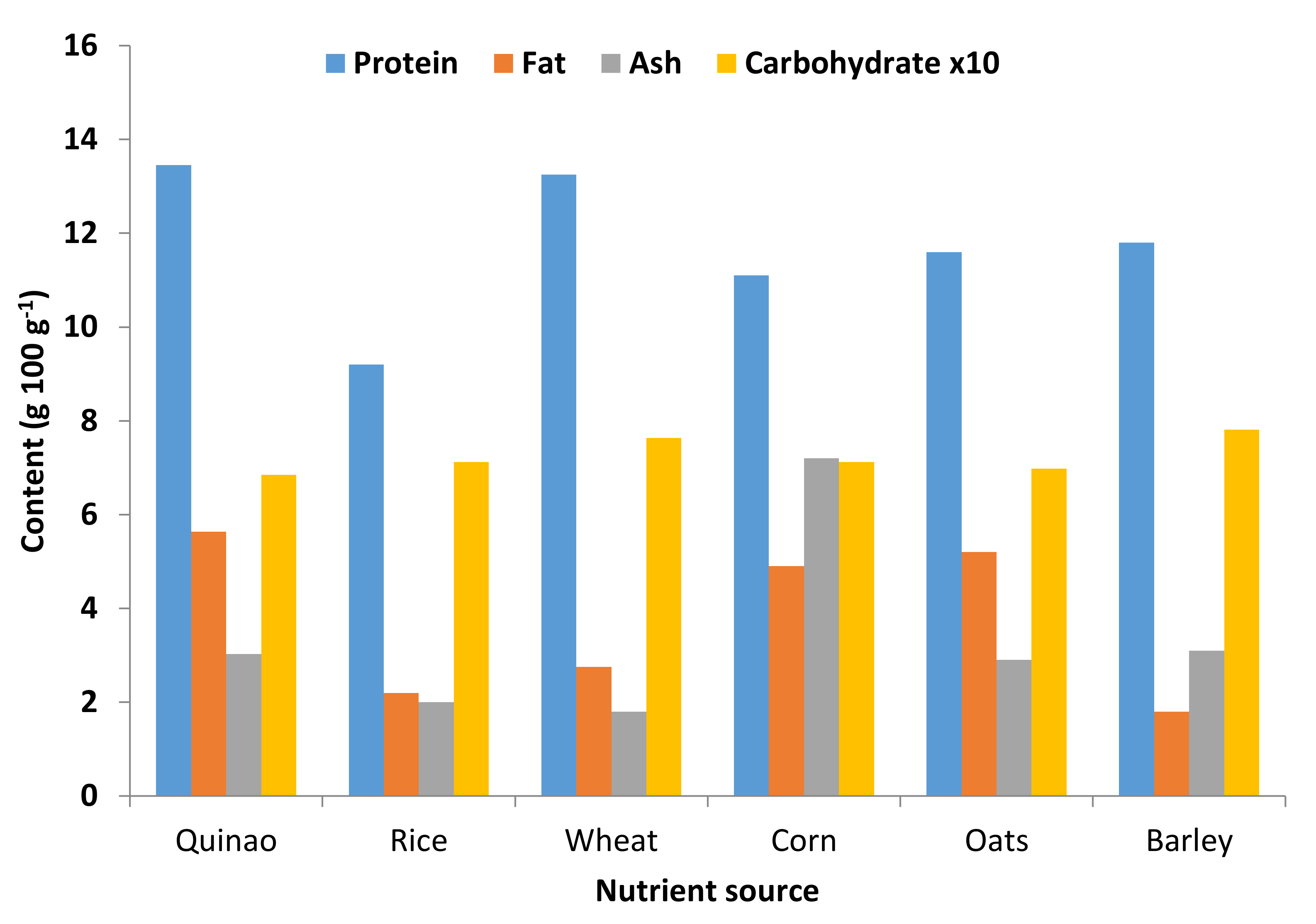
5. Nutritional Importance
6. Ecosystem Functions
6.1. Phytodesalinization
6.2. Phytoremediation
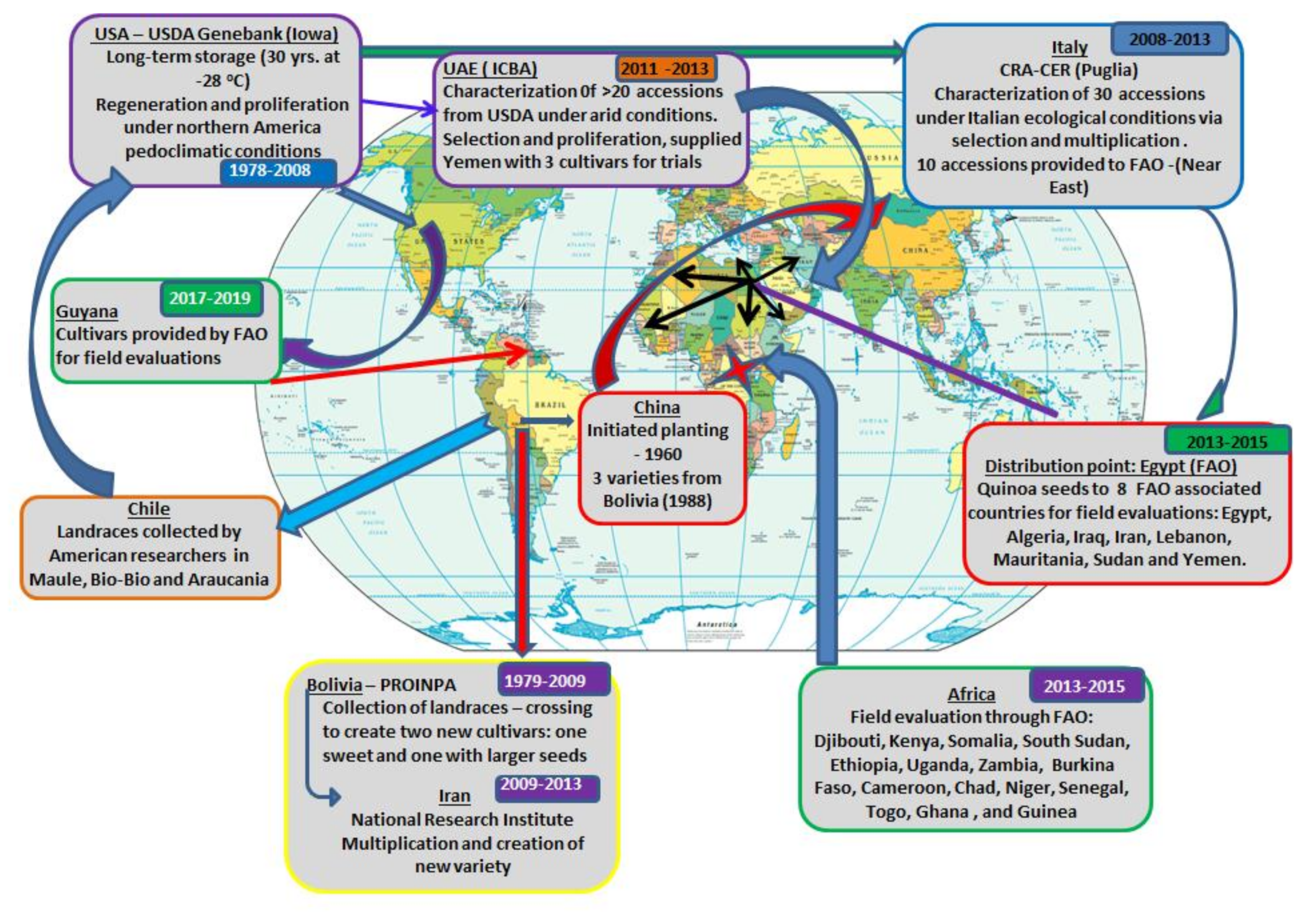
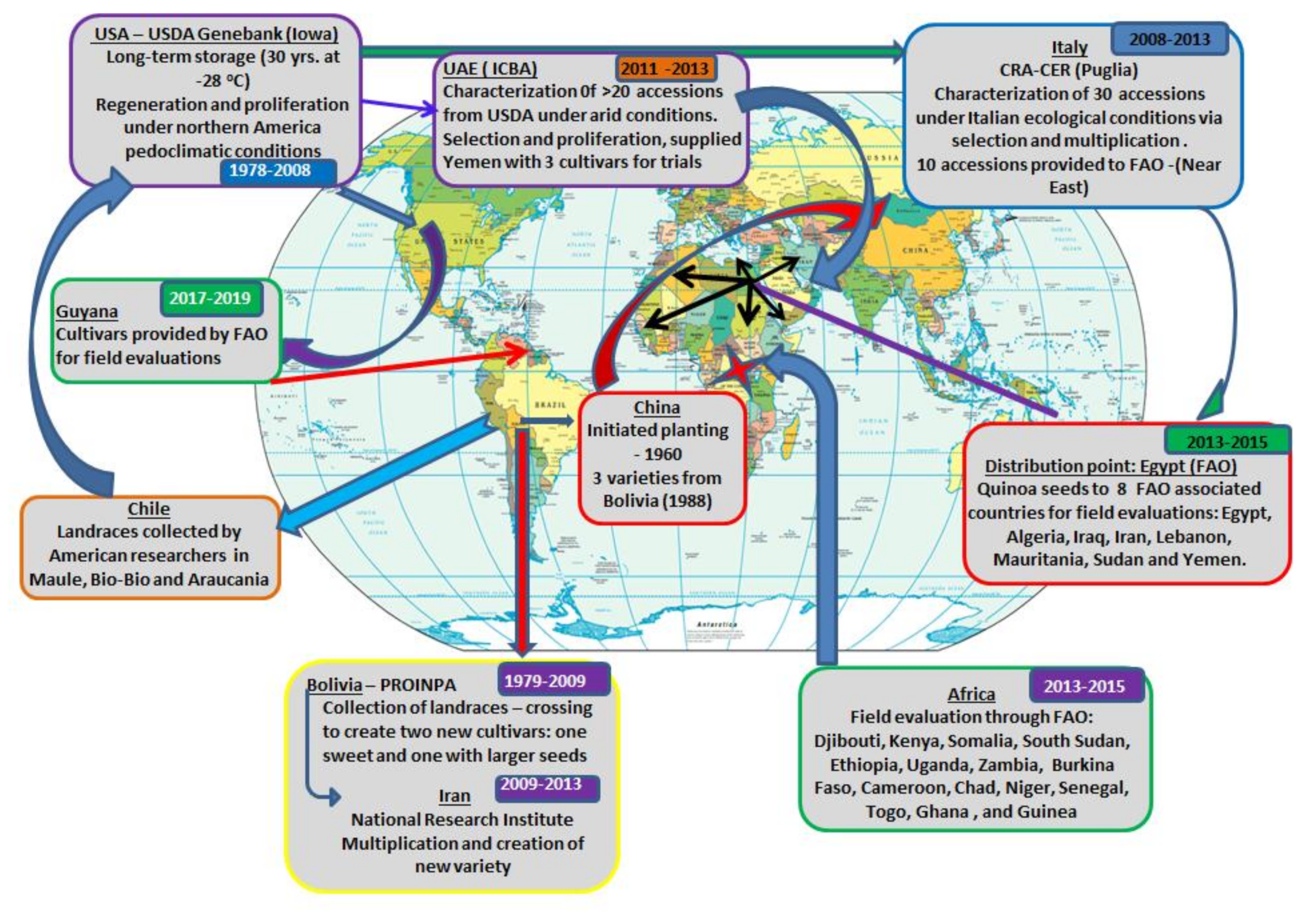
7. Global Expansion
8. Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- United Nations Convention to Combat Desertification. Policy Brief No. 3: Sustainable Land Management Solutions. Available online: http://catalogue.unccd.int/SER_SPIPB_3_ENG_WEB.pdf (accessed on 2 January 2019).
- González, J.A.; Eisa, S.S.S.; Hussin, S.A.E.S.; Prado, F.E. Quinoa: An Incan crop to face global changes in agriculture. Quinoa Improv. Sustain. Prod. 2015, 1–18. [Google Scholar] [CrossRef]
- Food and Agricultural Orgainisation of the United Nations (FAO); International Fund for Agricultural Development (IFAD); United Nations Children’s Fund (UNICEF); World Food Programme (WFP); World Health Organization (WHO). The State of Food Security and Nutrition in the World 2018. Building Climate Resilience for Food Security and Nutrition; FAO: Rome, Italy, 2018; Available online: http://www.fao.org/3/i9553en/i9553en.pdf (accessed on 1 January 2019).
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The global expansion of quinoa: Trends and limits. Front. Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Muscolo, A. Introduction to the Special Issue: Halophytes in a changing world. AoB Plants 2015, 7. [Google Scholar] [CrossRef] [PubMed]
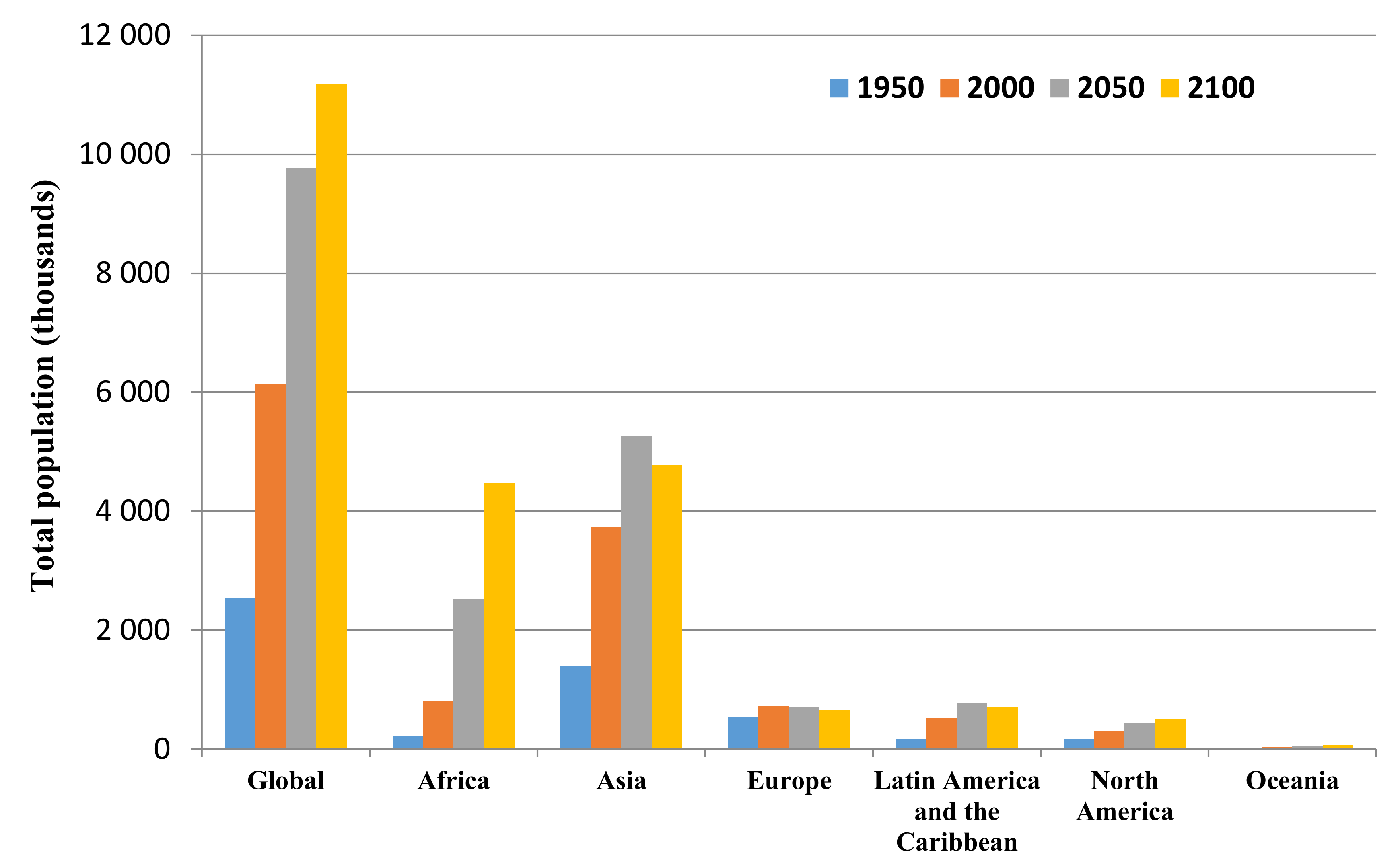
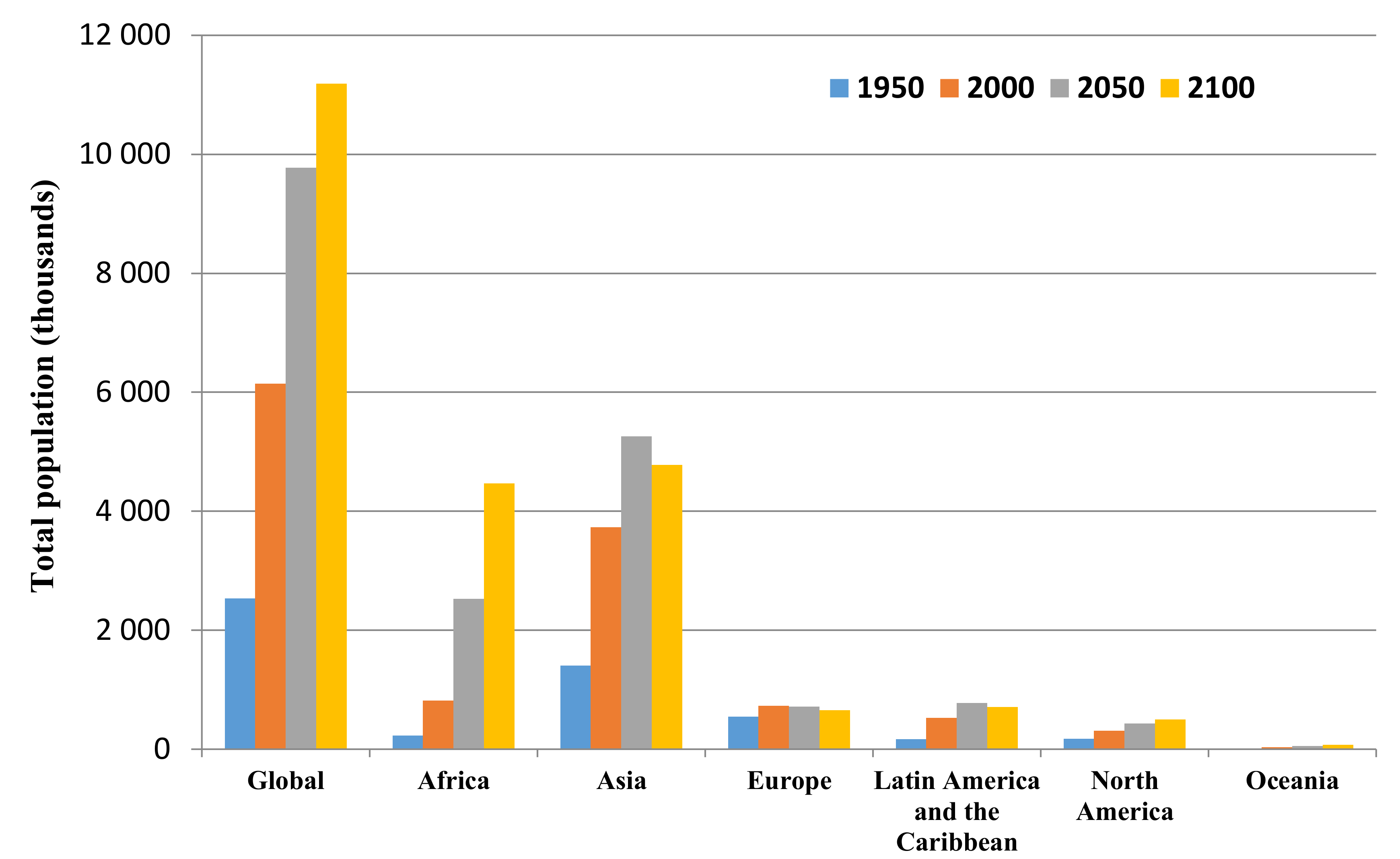
- UN. Department of Economic and Social Affairs, Population Division. World Population Prospects: The 2017 Revision, Key Findings and Advance Tables. Working Paper No. ESA/P/WP/248. Available online: https://population.un.org/wpp/Publications/ (accessed on 2 January 2019).
- Ruiz, K.B.; Biondi, S.; Martinez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.-E. Quinoa—A model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosyst. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Oses, R.; Acuña-Rodríguez, I.S.; Antognoni, F.; Martinez-Mosqueira, E.A.; Coulibaly, A.; Canahua-Murillo, A.; Pinto, M.; Zurita-Silva, A.; et al. Quinoa biodiversity and sustainability for food security under climate change. A review. Agron. Sustain. Dev. 2014, 34, 349–359. [Google Scholar] [CrossRef]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.-E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2010, 62, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.; Mujica, A. The genetic resources of Andean grain amaranths (Amaranthus caudatus L., A. cruentus L. and A. hipochondriacus L.) in America. Plant Genet. Resour. Newsl. 2003, 133, 41–44. [Google Scholar]
- Jacobsen, S.-E.; Liu, F.; Jensen, C.R. Does root-sourced ABA play a role for regulation of stomata under drought in quinoa (Chenopodium quinoa Willd.). Sci. Hortic. 2009, 122, 281–287. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Maughan, P.J.; Turner, T.B.; Coleman, C.E.; Elzinga, D.B.; Jellen, E.N.; Morales, J.A.; Udall, J.A.; Fairbanks, D.J.; Bonifacio, A. Characterization of Salt Overly Sensitive 1 (SOS1) gene homoeologs in quinoa (Chenopodium quinoa Willd.). Genome 2009, 52, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Konishi, Y. Nutritional characteristics of pseudocereal amaranth and quinoa: Alternative foodstuff for patients with food allergy. J. Jpn. Soc. Nutr. Food Sci. 2002, 55, 299–302. [Google Scholar] [CrossRef]
- Schlick, G.; Bubenheim, D.L. Quinoa: An Emerging New Crop with Potential for CELSS. Available online: https://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/19940015664.pdf (accessed on 21 September 2019).
- Lescano, R.J.L. Genetics and Improvement of High Andean Crops: Quinoa, Kañihua, Tarwi, Kiwicha, Bitter Potato, Olluco, Mashua and Oca Programa Interinstitucional de Waru Waru; CIMA: Cieneguilla, Peru, 1994; pp. 459–561. [Google Scholar]
- Gonzalez, J.A.; Konishi, Y.; Bruno, M.; Valoy, M.; Prado, F.E. Interrelationships among seed yield, total protein and amino acid composition of ten quinoa (Chenopodium quinoa) cultivars from two different agroecological regions. J. Sci. Food Agric. 2012, 92, 1222–1229. [Google Scholar] [CrossRef]
- Abugoch, L.; Castro, E.; Tapia, C.; Anon, M.C.; Gajardo, P.; Villarroel, A. Stability of quinoa flour proteins (Chenopodium quinoa Willd.) during storage. Int. J. Food Sci. Technol. 2009, 44, 2013–2020. [Google Scholar] [CrossRef]
- Tapia, M. La Quinua: Historia, Distribución Geográfica, Actual Producción y Usos. Ambienta: La Revista del Ministerio de Medio Ambiente 2012, 99, 104–119. Available online: http://www.revistaambienta.es/WebAmbienta/marm/Dinamicas/secciones (accessed on 21 January 2019).
- Heiser, C.B.; Nelson, D.C. On the origin of the cultivated chenopods (Chenopodium). Genetics 1974, 78, 503–505. [Google Scholar]
- Wilson, H.D.; Heiser, C.B., Jr. The origin and evolutionary relationships of ‘huauzontle’ (Chenopodium nuttalliae Safford), domesticated chenopod of Mexico. Am. J. Bot. 1979, 66, 198–206. [Google Scholar] [CrossRef]
- Kolano, B.; McCann, J.; Orzechowska, M.; Siwinska, D.; Temsch, E.; Weiss-Schneeweiss, H. Molecular and cytogenetic evidence for an allotetraploid origin of Chenopodium quinoa and C. berlandieri (Amaranthaceae). Mol. Phylogenet. Evol. 2016, 100, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Galwey, N.; Leakey, C.L.A.; Price, K.R.; Fenwick, G.R. Chemical composition and nutritional characteristics of quinoa (Chenopodium quinoa Willd.). Food Sci. Nutr. 1989, 42, 245–261. [Google Scholar] [CrossRef]
- Jensen, C.; Jacobsen, S.-E.; Andersen, M.; Andersen, S.; Mogensen, V.; Núñez, N.; Rasmussen, L. Leaf gas exchange and water relation characteristics of field quinoa (Chenopodium quinoa Willd.) during soil drying. Eur. J. Agron. 2000, 13, 11–25. [Google Scholar] [CrossRef]
- Field, C.B.; Barros, V.R.; Dokken, D.; Mach, K.; Mastrandrea, M.; Bilir, T.; Chatterjee, M.; Ebi, K.L.; Estrada, Y.O.; Genova, R.C.; et al. IPCC 2014: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/WGIIAR5 (accessed on 12 September 2018).
- Metz, B.; Davidson, O.; De Coninck, H.; Loos, M.; Meyer, L. IPCC, 2005: IPCC Special Report on Carbon Dioxide Capture and Storage. Prepared by Working Group III of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/report/srccs (accessed on 1 January 2018).
- IPCC. Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change 2014. Available online: https://www.ipcc.ch/report/ar5/wg2 (accessed on 22 March 2019).
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Bazile, D.; Bertero, H.D.; Nieto, C. State of the Art Report on Quinoa Around the World in 2013; 2015. Available online: http://www.fao.org/3/i4042e/I4042E.pdf (accessed on 21 November 2018).
- Valencia-Chamorro, S. Quinoa. In Encyclopedia of Food Science and Nutrition; Caballero, B., Ed.; Academic Press: Amsterdam, The Nethelands, 2003; Volume 8, pp. 4895–4902. [Google Scholar]
- Miranda, M.; Vega-Gálvez, A.; Quispe-Fuentes, I.; Rodríguez, M.J.; Maureira, H.; Martínez, E.A. Nutritional aspects of six quinoa (Chenopodium quinoa Willd.) ecotypes from three geographical areas of Chile. Chil. J. Agric. Res. 2012, 72, 175. [Google Scholar] [CrossRef]
- Anjum, N.A.; Ahmad, I.; Válega, M.; Mohmood, I.; Gill, S.S.; Tuteja, N.; Duarte, A.C.; Pereira, E. Salt marsh halophyte services to metal-metalloid remediation: Assessment of the processes and underlying mechanisms. Crit. Rev. Environ. Sci. Technol. 2014, 44, 2038–2106. [Google Scholar] [CrossRef]
- Flowers, T.; Yeo, A. Breeding for salinity resistance in crop plants: Where next? Funct. Plant Biol. 1995, 22, 875–884. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Glenn, E.P.; Anday, T.; Chaturvedi, R.; Martinez-Garcia, R.; Pearlstein, S.; Soliz, D.; Nelson, S.G.; Felger, R.S. Three halophytes for saline-water agriculture: An oilseed, a forage and a grain crop. Environ. Exp. Bot. 2013, 92, 110–121. [Google Scholar] [CrossRef]
- Adolf, V.I.; Shabala, S.; Andersen, M.N.; Razzaghi, F.; Jacobsen, S.-E. Varietal differences of quinoa’s tolerance to saline conditions. Plant Soil. 2012, 357, 117–129. [Google Scholar] [CrossRef]
- Mäser, P.; Eckelman, B.; Vaidyanathan, R.; Horie, T.; Fairbairn, D.J.; Kubo, M.; Yamagami, M.; Yamaguchi, K.; Nishimura, M.; Uozumi, N.; et al. Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1. FEBS Lett. 2002, 531, 157–161. [Google Scholar] [CrossRef]
- Blumwald, E. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol. 2000, 12, 431–434. [Google Scholar] [CrossRef]
- Brownlee, C. Plant physiology: One way to dump salt. Curr. Biol. 2018, 28, R1145–R1147. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Christiansen, J.L.; Ruiz-Tapia, E.; Jørnsgård, B.; Jacobsen, S.-E. Fast Seed Germination of Quinoa (Chenopodium quinoa) at Low Temperature. Alternative Crops for Sustainable Agriculture: Research Progress: COST 814, Workshop, Turku, 1999; 29/01/2019. Available online: https://research.ku.dk/search/?.../fast-seed-germination-of-quinoa (accessed on 29 January 2019).
- Karyotis, T.; Iliadis, C.; Noulas, C.; Mitsibonas, T. Preliminary research on seed production and nutrient content for certain quinoa varieties in a saline-sodic soil. J. Agron. Crop Sci. 2003, 189, 402–408. [Google Scholar] [CrossRef]
- Wilson, C.; Read, J.J.; Abo-Kassem, E. Effect of mixed-salt salinity on growth and ion relations of a quinoa and a wheat variety. J. Plant Nutr. 2002, 25, 2689–2704. [Google Scholar] [CrossRef]
- Gomez-Pando, L.R.; Alvarez-Castro, R.; Eguiluz-de La Barra, A. Effect of salt stress on Peruvian Germplasm of Chenopodium quinoa Willd.: A promising crop. J. Agron. Crop Sci. 2010, 196, 391–396. [Google Scholar] [CrossRef]
- Razzaghi, F.; Jacobsen, S.-E.; Jensen, C.R.; Andersen, M.N. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought—Mechanisms of tolerance. Funct. Plant Biol. 2015, 42, 136–148. [Google Scholar] [CrossRef]
- Morales, A.J.; Bajgain, P.; Garver, Z.; Maughan, P.J.; Udall, J.A. Physiological responses of Chenopodium quinoa to salt stress. Int. J. Plant Physiol. Biochem. 2011, 3, 219–232. [Google Scholar] [CrossRef]
- Orsini, F.; Accorsi, M.; Gianquinto, G.; Dinelli, G.; Antognoni, F.; Carrasco, K.B.R.; Martinez, E.A.; Alnayef, M.; Marotti, I.; Bosi, S.; et al. Beyond the ionic and osmotic response to salinity in Chenopodium quinoa: Functional elements of successful halophytism. Funct. Plant Biol. 2011, 38, 818–831. [Google Scholar] [CrossRef]
- Metternicht, G.I.; Zinck, J.A. Remote sensing of soil salinity: Potentials and constraints. Remote Sens. Environ. 2003, 85, 1–20. [Google Scholar] [CrossRef]
- Qadir, M.; Quillerou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.; Drechsel, P.; Noble, A. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Barrera-Figueroa, B.; Gao, L.; Diop, N.N.; Wu, Z.; Ehlers, J.D.; A Roberts, P.; Close, T.J.; Zhu, J.-K.; Liu, R. Identification and comparative analysis of drought-associated microRNAs in two cowpea genotypes. BMC Plant Biol. 2011, 11, 127. [Google Scholar] [CrossRef]
- Bhargava, A.; Srivastava, S. Quinoa: Botany, Production and Uses; CABI: Wallingford, UK, 2013. [Google Scholar]
- Raney, J.A. Transcriptome analysis of drought induced stress in Chenopodium quinoa. Am. J. Plant Sci. 2012, 5, 338–357. [Google Scholar] [CrossRef]
- UNCCD. Global Land Outlook. UN Convention to Combat Desertification Bonn. Available online: https://www.unccd.int/actions/global-land-outlook-glo (accessed on 2 January 2019).
- Bhargava, A.; Carmona, F.F.; Bhargava, M.; Srivastava, S. Approaches for enhanced phytoextraction of heavy metals. J. Environ. Manag. 2012, 105, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Andrés, A.-S.; Sven-Erik, J.; Fatemeh, R.; Ricardo, A.-F. Quinoa Drought Responses and Adaptation. Available online: http://www.fao.org/3/i4042e/I4042E.pdf (accessed on 21 November 2018).
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.; Raes, D.; Jacobsen, S.E.; Michel, T. Agroclimatic constraints for rainfed agriculture in the Bolivian Altiplano. J. Arid Environ. 2007, 71, 109–121. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Quispe, H.; Mujica, A. Quinoa: An Alternative Crop for Saline Soils in the Andes, Scientists and Farmer—Partners in Research for the 21st Century; CIP: Miraflores, Peru, 1999. [Google Scholar]
- Canahua, A. Observaciones del comportamiento de la quinua a la sequía. In Primer Congreso Internacional Sobre Cultivos Andinos; Universidad Nacional San Cristobal de Huamanga, Instituto Interamericano de Ciencias Agricolas: Ayacucho, Peru, 1977; pp. 390–392. [Google Scholar]
- Garcia, M. Agroclimatic Study and Drought Resistance Analysis of Quinoa for An Irrigation Strategy in the Bolivian Altiplano. Dissertationes de Agricultura 556; KU Leuven: Leuven, Belgium, 2003. [Google Scholar]
- Naik, G.; Joshi, G. Ineffectual role of proline metabolism in salt-stressed sugarcane leaves. Proc. Plant Sci. 1983, 92, 265–269. [Google Scholar] [CrossRef]
- Siddiqu, S.; Krishnamoorthy, H. Effect of B-nine on the proline content of gram (Cicer arietinum) under saline conditions. Indian J. Plant Physiol. 1987, 1, 107–110. [Google Scholar]
- Vera-Hernández, P.; Ramírez, M.; Núñez, M.; Ruiz-Rivas, M.; Rosas-Cárdenas, F. Proline as a probable biomarker of cold stress tolerance in Sorghum (Sorghum bicolor). Mex. J. Biotechnol. 2018, 3, 77–86. [Google Scholar] [CrossRef]
- Le Tacon, P.; Vacher, J.; Eldin, M.; Imaña, E. Los Riesgos de Helada en el Altiplano Boliviano. ORSTOM/IBTA La Paz. Available online: http://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers09-11/38556.pdf (accessed on 21 September 2018).
- Bois, J.E.; Winkel, T.; Lhomme, J.P.; Raffaillac, J.P.; Rocheteau, A. Response of some Andean cultivars of quinoa (Chenopodium quinoa Willd.) to temperature: Effects on germination, phenology, growth and freezing. Eur. J. Agron. 2006, 25, 299–308. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Amjad, M.; Iqbal, S.; Jacobsen, S.E. Growth and physiological responses of quinoa to drought and temperature stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Aloisi, I.; Parrotta, L.; Ruiz, K.B.; Landi, C.; Bini, L.; Cai, G.; Biondi, S.; Del Duca, S. New insight into quinoa seed quality under salinity: Changes in proteomic and amino acid profiles, phenolic content, and antioxidant activity of protein extracts. Front. Plant Sci. 2016, 7, 656. [Google Scholar] [CrossRef] [PubMed]
- Oelke, E.; Putnam, D.; Teynor, T.; Oplinger, E. Quinoa. Alternative Field Crops Manual. University of Wisconsin Cooperative Extension Service, University of Minnesota Extension Service. Available online: https://hort.purdue.edu/newcrop/afcm/quinoa.html (accessed on 24 March 2019).
- Jacobsen, S.-E.; Stølen, O. Quinoa-morphology, phenology and prospects for its production as a new crop in Europe. Eur. J. Agron. 1993, 2, 19–29. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Rajan, S.; Ohri, D. Genetic diversity for morphological and quality traits in quinoa (Chenopodium quinoa Willd.) germplasm. Genet. Resour. Crop Evol. 2007, 54, 167–173. [Google Scholar] [CrossRef]
- Sosa-Zuniga, V.; Brito, V.; Fuentes, F.; Steinfort, U. Phenological growth stages of quinoa (Chenopodium quinoa) based on the BBCH scale. Ann. Appl. Biol. 2017, 171, 117–124. [Google Scholar] [CrossRef]
- Maliro, M.F.A.; Guwela, V.F.; Nyaika, J.; Murphy, K.M. Preliminary studies of the performance of quinoa (Chenopodium quinoa Willd.) genotypes under irrigated and rainfed conditions of Central Malawi. Front. Plant Sci. 2017, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Präger, A.; Munz, S.; Nkebiwe, P.M.; Mast, B.; Graeff-Hönninger, S. Yield and quality characteristics of different quinoa (Chenopodium quinoa Willd.) cultivars grown under field conditions in southwestern Germany. Agronomy 2018, 8, 197. [Google Scholar] [CrossRef]
- Escuredo, O.; Martin, M.I.G.; Moncada, G.W.; Fischer, S.; Hierro, J.M.H. Amino acid profile of the quinoa (Chenopodium quinoa Willd.) using near infrared spectroscopy and chemometric techniques. J. Cereal Sci. 2014, 60, 67–74. [Google Scholar] [CrossRef]
- Filho, A.M.; Pirozi, M.R.; Borges, J.T.; Pinheiro Sant’Ana, H.M.; Chaves, J.B.; Coimbra, J.S. Quinoa: Nutritional, functional, and antinutritional aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco, R.; Espinoza, C.; Jacobsen, S.-E. Nutritional value and use of the Andean crops quinoa (Chenopodium quinoa) and kañiwa (Chenopodium pallidicaule). Food Rev. Int. 2003, 19, 179–189. [Google Scholar] [CrossRef]
- USDA. National Nutrient Database for Standard Reference, Release 28. Available online: https://ndb.nal.usda.gov/ndb/ (accessed on 2 January 2019).
- Aluko, R.E.; Monu, E. Functional and bioactive properties of quinoa seed protein hydrolysates. J. Food Sci. 2003, 68, 1254–1258. [Google Scholar] [CrossRef]
- Caperuto, L.C.; Amaya-Farfan, J.; Camargo, C.R.O. Performance of quinoa (Chenopodium quinoa Willd) flour in the manufacture of gluten-free spaghetti. J. Sci. Food Agric. 2001, 81, 95–101. [Google Scholar] [CrossRef]
- Mastebroek, H.D.; Limburg, H.; Gilles, T.; Marvin, H.J.P. Occurrence of sapogenins in leaves and seeds of quinoa (Chenopodium quinoa Willd). J. Sci. Food Agric. 2000, 80, 152–156. [Google Scholar] [CrossRef]
- Ruales, J.; Nair, B.M. Nutritional quality of the protein in quinoa (Chenopodium quinoa, Willd) seeds. Plant Foods Hum. Nutr. 1992, 42, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Lawson, L.; Fairbanks, D.J.; Robison, L.; Andersen, W. Seed lipid content and fatty acid composition of three quinoa cultivars. J. Food Compos. Anal. 1993, 6, 41–44. [Google Scholar] [CrossRef]
- Wright, K.H.; Huber, K.C.; Fairbanks, D.J.; Huber, C.S. Isolation and characterization of Atriplex hortensis and sweet Chenopodium quinoa starches. Cereal Chem. 2002, 79, 715–719. [Google Scholar] [CrossRef]
- Wright, K.H.; Pike, O.A.; Fairbanks, D.J.; Huber, C.S. Composition of Atriplex hortensis, sweet and bitter Chenopodium quinoa seeds. J. Food Sci. 2002, 67, 1383–1385. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- FAO/WHO/UNU. Energy and Protein Requirements: Report of a Joint FAO/WHO/UNU Expert Consultation. WHO. Report No.: 9241207248. Available online: https://www.ncbi.nlm.nih.gov/pubmed/3937340 (accessed on 19 December 2017).
- Ahamed, N.T.; Singhal, R.S.; Kulkarni, P.R.; Pal, M. A lesser-known grain, Chenopodium quinoa: Review of the chemical composition of its edible parts. Food Nutr. Bull. 1998, 19, 61–70. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, Y.J.; Kim, Y.H.; Yoon, K.S. Antioxidant and antimicrobial activities of quinoa (Chenopodium quinoa Willd.) seeds cultivated in Korea. Prev. Nutr. Food Sci. 2017, 22, 195. [Google Scholar] [CrossRef]
- Kozioł, M. Chemical composition and nutritional evaluation of quinoa (Chenopodium quinoa Willd.). J. Food Compos. Anal. 1992, 5, 35–68. [Google Scholar] [CrossRef]
- Halford, N.G.; Curtis, T.Y.; Chen, Z.; Huang, J. Effects of abiotic stress and crop management on cereal grain composition: Implications for food quality and safety. J. Exp. Bot. 2015, 66, 1145–1156. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2014, 115, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Ado, M.N.; Guero, Y.; Michot, D.; Soubeiga, B.; Kiesse, T.S.; Walter, C. Phytodesalinization of irrigated saline Vertisols in the Niger Valley by Echinochloa stagnina. Agric. Water Manag. 2016, 177, 229–240. [Google Scholar] [CrossRef]
- Zorrig, W.; Rabhi, M.; Ferchichi, S.; Smaoui, A.; Abdelly, C. Phytodesalination: A solution for salt-affected soils in arid and semi-arid regions. J. Arid Land Stud. 2012, 22, 299–302. [Google Scholar]
- Bhargava, A.; Shukla, S.; Srivastava, J.; Singh, N.; Ohri, D. Chenopodium: A prospective plant for phytoextraction. Acta Physiol. Plant. 2008, 30, 111–120. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Taamalli, M.; Ghabriche, R.; Amari, T.; Mnasri, M.; Zolla, L.; Lutts, S.; Abdely, C.; Ghnaya, T. Comparative study of Cd tolerance and accumulation potential between Cakile maritima L. (halophyte) and Brassica juncea L. Ecol. Eng. 2014, 71, 623–627. [Google Scholar] [CrossRef]
- Wang, H.-L.; Tian, C.-Y.; Jiang, L.; Wang, L. Remediation of heavy metals contaminated saline soils: A halophyte choice? Environ. Sci. Technol. 2013, 48, 21–22. [Google Scholar] [CrossRef]
- Zulfqar, S.; Wahid, A.; Farooq, M.; Maqbool, N.; Arfan, M. Phytoremediation of soil cadmium using Chenopodium species. Pak. J. Agric. Sci. 2012, 49, 435–445. [Google Scholar]
- Haseeb, M.; Basra, S.M.A.; Afzal, I.; Wahid, A. Quinoa response to lead: Growth and lead partitioning. Int. J. Agric. Biol. 2018, 20, 338–344. [Google Scholar] [CrossRef]
- FAO/WHO. Codex Alimentarius Commission. Food Additives and Contaminants. Joint FAO. Available online: www.fao.org/input/download/report/17/al9112ae.pdf (accessed on 11 February 2016).
- Bhargava, A.; Shukla, S.; Ohri, D. Chenopodium quinoa—An Indian perspective. Ind. Crops Prod. 2006, 23, 73–87. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristics of quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Food and Agricultural Organisation of the United Nations Statitics (FAOSTAT). Food and Agricultural Data. 2017. Available online: http://www.fao.org/faostat/en (accessed on 30 March 2019).
- CIA. The World Factbook 2012–13. Central Intelligence Agency. Report No.: 0160911427. Available online: https://geology.com/world/cia-world-map.gif (accessed on 29 March 2019).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Areas | ||||||
|---|---|---|---|---|---|---|
| North | Centre | South | ||||
| Ancovinto | Cancosa | Cáhuil | Faro | Regalona | Villarrica | |
| Ca | 77.10 ± 2.25 | 105.01 ± 4.56 | 120.33 ± 1.55 | 121.39 ± 3.74 | 118.08 ± 5.93 | 211.29 ± 0.35 |
| Mg | 152.31 ± 0.44 | 150.91 ± 0.54 | 160.55 ± 0.72 | 166.74 ± 0.22 | 155.77 ± 1.36 | 160.44 ± 0.19 |
| Na | <0.01 ± 0 | <0.01 ± 0 | 18.46 ± 11.68 | 17.14 ± 0.70 | 19.58 ± 3.19 | 23.41 ± 1.08 |
| K | 2188.54 ± 0.67 | 2325.56 ± 12.22 | 1672.79 ± 0.56 | 1774.43 ± 11.89 | 2038.42 ± 10.88 | 2243.99 ± 37.49 |
| Fe | 6.91 ± 0.31 | 7.19 ± 0.15 | 5.06 ± 0.03 | 5.84 ± 0.08 | 7.03 ± 0.26 | 4.82 ± 0.28 |
| Cu | 1.52 ± 0.20 | 0.75 ± 0 | 0.86 ± 0.04 | 0.89 ± 0.02 | 0.95 ± 0.02 | 0.89 ± 0.02 |
| Mn | 2.39 ± 0.02 | 2.36 ± 0.02 | 6.47 ± 0.17 | 5.73 ± 0.08 | 4.67 ± 0.15 | 2.76 ± 0.19 |
| Zn | 3.75 ± 0.22 | 3.10 ± 0.03 | 4.02 ± 0.17 | 2.73 ± 0.01 | 4.65 ± 0.06 | 5.01 ± 0.38 |
| P | 526.36 ± 3.79 | 434.65 ± 14.27 | 409.92 ± 8.42 | 480.96 ± 2.62 | 353.53 ± 10.88 | 285.06 ± 65.81 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaikishun, S.; Li, W.; Yang, Z.; Song, S. Quinoa: In Perspective of Global Challenges. Agronomy 2019, 9, 176. https://doi.org/10.3390/agronomy9040176
Jaikishun S, Li W, Yang Z, Song S. Quinoa: In Perspective of Global Challenges. Agronomy. 2019; 9(4):176. https://doi.org/10.3390/agronomy9040176
Chicago/Turabian StyleJaikishun, Sirpaul, Wenqiang Li, Zhenbiao Yang, and Shikui Song. 2019. "Quinoa: In Perspective of Global Challenges" Agronomy 9, no. 4: 176. https://doi.org/10.3390/agronomy9040176
APA StyleJaikishun, S., Li, W., Yang, Z., & Song, S. (2019). Quinoa: In Perspective of Global Challenges. Agronomy, 9(4), 176. https://doi.org/10.3390/agronomy9040176