Biomass Production and Potential Fixed Nitrogen Inputs from Leguminous Cover Crops in Subtropical Avocado Plantations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites and Trial Design
2.1.1. Trial Site 1—Tuckombil NSW
2.1.2. Trial Site 2—Alstonville New South Wales
2.2. Biomass Production, N Accumulation and N Fixation in Legume Cover Crops
3. Results
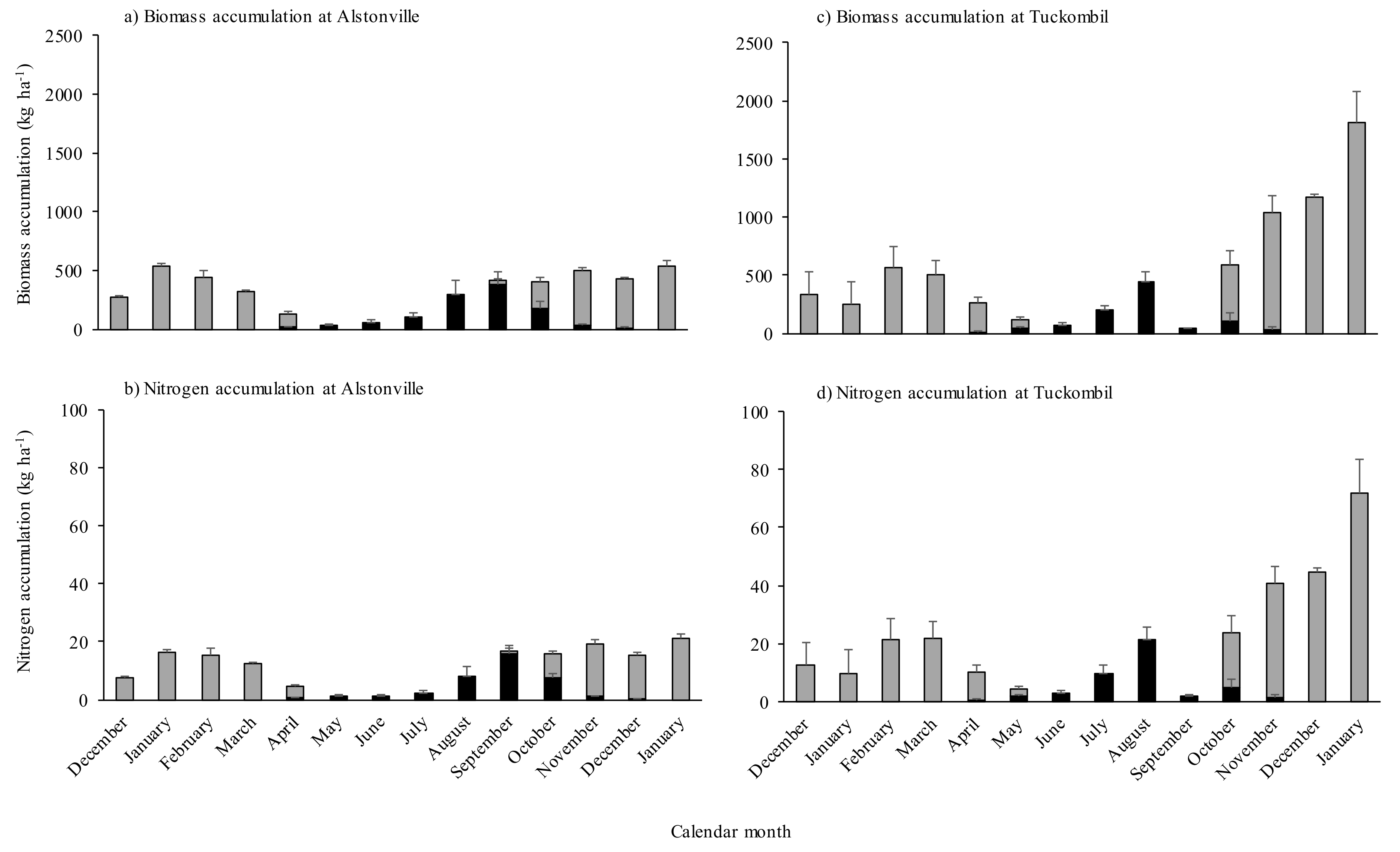
3.1. Dry Matter Production
3.2. Nitrogen Fixation and N Accumulation in Shoots
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Huett, D.O.; Dirou, J.F. An evaluation of the rationale for fertiliser management of tropical fruit crops. Aust. J. Exp. Agric. 2000, 40, 1137–1143. [Google Scholar] [CrossRef]
- Lovatt, C.J. Properly timed soil-applied nitrogen fertilizer increases yield and fruit size of ‘Hass’ avocado. J. Am. Soc. Hort. Sci. 2001, 126, 555–559. [Google Scholar] [CrossRef]
- Masso, C.; Baijukya, F.; Ebanyat, P.; Bouaziz, S.; Wendt, J.; Bekunda, M.; Vanlauwe, B. Dilemma of nitrogen management for future food security in sub-Saharan Africa—A review. Soil Res. 2017, 55, 425–434. [Google Scholar] [CrossRef]
- Rose, T.J.; Morris, S.J.; Quin, P.; Kearney, L.J.; Kimber, S.; Van Zwieten, L. The nitrification inhibitor DMPP applied to subtropical rice has an inconsistent effect on nitrous oxide emissions. Soil Res. 2017, 55, 547–552. [Google Scholar] [CrossRef]
- Rempelos, L.; Almuayrifi, A.M.; Baranski, M.; Tetard-Jones, C.; Eyre, M.; Shotton, P.; Cakmak, I.; Ozturk, L.; Cooper, J.; Volakakis, N.; et al. Effects of agronomic management and climate on leaf phenolic profiles, disease severity and grain yield in organic and conventional wheat production systems. J. Agric. Food Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.S.; Peoples, M.B.; Boddey, R.M.; Gresshoff, P.M.; Hauggaard-Nielsen, H.; Alves, B.J.R.; Morrison, M.J. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron. Sustain. Dev. 2012, 32, 329–364. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover crops and living mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Santos, J.C.F.; da Cunha, A.J.; Ferreira, F.A.; Santos, R.H.S.; Sakiyama, N.S.; de Lima, P.C. Herbaceous legumes intercropping in weed management of the coffee crop. J. Agric. Environ. Sci. 2016, 5, 91–100. [Google Scholar]
- Snoeck, D.; Zapata, F.; Domenach, A.-M. Isotopic evidence of the transfer of nitrogen fixed by legumes to coffee trees. Biotechnol. Agron. Soc. Environ. 2000, 4, 95–100. [Google Scholar]
- Agamuthu, P.; Broughton, W.J. Nutrient cycling within the developing oil palm-legume ecosystem. Agric. Ecosys. Environ. 1985, 13, 111–123. [Google Scholar] [CrossRef]
- Rose, T.J.; Kearney, L.J.; Morris, S.G.; Van Zwieten, L.; Erler, D.V. Pinto peanut cover crop nitrogen contributions and potential to mitigate nitrous oxide emissions from subtropical coffee plantations. Sci. Total Environ. 2019, 656, 108–117. [Google Scholar] [CrossRef]
- Fulkerson, W.J.; Slack, K. Management of Lolium perenne/Trifolium repens pastures in the subtropics. II. Effect of summer defoliation, irrigation duration, seedbed preparation and pasture type. Aust. J. Agric. Res. 1994, 45, 721–728. [Google Scholar] [CrossRef]
- Unkovich, M.; Herridge, D.; Peoples, M.; Cadisch, G.; Boddey, R.; Giller, K.; Alves, B.; Chalk, P. Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; ACIAR Monograph No. 136: Canberra, Australia, 2008. [Google Scholar]
- FAO–UNESCO. Soil Map of the World. Food and Agriculture Organization of the United Nations: Rome, Italy, 1974. [Google Scholar]
- Rayment, G.E.; Lyons, D.J. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press: Port Melbourne, Australia, 2011. [Google Scholar]
- Shearer, G.; Kohl, D.H. N2 fixation in field settings: Estimations based on natural 15N abundance. Aust. J. Plant Physiol. 1986, 13, 699–756. [Google Scholar]
- Rose, T.J.; Rengel, Z.; Ma, Q.; Bowden, J.W. Post-flowering supply of P, but not K, is required for maximum canola seed yields. Eur. J. Agron. 2008, 28, 371–379. [Google Scholar] [CrossRef]
- Ferreira, I.Q.; Rodrigues, M.A.; Claro, A.M.; Arrobas, M. Management of nitrogen-rich legume cover crops as mulch in traditional olive orchards. Commun. Soil Sci. Plant Anal. 2015, 46, 1881–1894. [Google Scholar] [CrossRef]
- Streeter, J. Inhibition of legume nodule formation and N2 fixation by nitrate. Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Tang, C.; Hinsinger, P.; Drevon, J.J.; Jaillard, B. Phosphorus deficiency impairs early nodule functioning and enhances proton release in roots of Medicago truncatula L. Ann. Bot. 2001, 88, 131–138. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Gridley, K.L.; Brady, J.N.; Phillips, T.; Tyerman, S.D. The role of molybdenum in agricultural plant production. Ann. Bot. 2005, 96, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Van Zwieten, L.; Rose, T.J.; Herridge, D.; Kimber, S.; Rust, J.; Cowie, A.; Morris, S. Enhanced biological N2 fixation and yield of faba bean (Vicia faba L.) in an acid soil following biochar addition: Dissection of causal mechanisms. Plant Soil 2015, 395, 7–20. [Google Scholar] [CrossRef]
- Reuter, D.J.; Robinson, J.B. Plant Analysis: An Interpretation Manual, 2nd ed.; CSIRO Publishing: Collingwood, Australia, 1997. [Google Scholar]
- Peoples, M.B.; Brockwell, J.; Hunt, J.R.; Swan, A.D.; Watson, L.; Hayes, R.C.; Li, G.D.; Hackney, B.; Nuttall, J.G.; Davies, S.L.; et al. Factors affecting the potential contributions of N2 fixation by legumes in Australian pasture systems. Crop Pasture Sci. 2012, 63, 759–786. [Google Scholar] [CrossRef]
- Høgh-Jensen, H.; Schjoerring, J.K. Rhizodeposition of nitrogen by red clover, white clover and ryegrass leys. Soil Biol. Biochem. 2001, 33, 439–448. [Google Scholar]
- Rose, T.J.; Kearney, L.J.; Erler, D.V.; Rose, M.T.; Van Zwieten, L.; Raymond, C.A. Influence of growth stage and seed nitrogen on B values and potential contributions to error in estimating biological N2 fixation using the 15N natural abundance method. Plant Soil 2018, 425, 389–399. [Google Scholar] [CrossRef]

{kind=link}
| Soil Property (0–100mm) | Site 1 Tuckombil | Site 2 Alstonville |
|---|---|---|
| Texture | Clay loam | Clay loam |
| pH (1:5 Water) | 5.93 | 6.46 |
| Total N (%) | 0.35 | 0.41 |
| Total C (%) | 4.09 | 5.85 |
| EC (dS m−1) | 0.13 | 0.09 |
| Bray 2 P (mg kg−1) | 23 | 7.6 |
| Colwell P (mg kg−1) | 114 | 58 |
| Cation exchange capacity (cmol+ kg−1) | 7.7 | 10.6 |
| Base cations (cmol+ kg−1) | ||
| Calcium | 5.98 | 5.86 |
| Magnesium | 1.13 | 3.17 |
| Potassium | 0.44 | 1.15 |
| Sodium | 0.09 | 0.20 |
| DTPA-extractable micronutrients (mg kg−1) | ||
| Zinc | 2.4 | 4.6 |
| Manganese | 9.0 | 7.0 |
| Iron | 102 | 206 |
| Copper | 5.7 | 1.6 |
| CaCl2-extratable B (mg kg−1) | 0.95 | 1.1 |
| KCl extractable ammonium (mg kg−1) | ||
| 0–100mm | 4.5 | 7.8 |
| 100–300mm | 5.0 | 4.1 |
| 300–600m | 6.4 | 3.3 |
| 600–900mm | 8.0 | 8.2 |
| KCl extractable nitrate (mg kg−1) | ||
| 0–100mm | 35 | 4.0 |
| 100–300mm | 109 | 4.7 |
| 300–600m | 79 | 4.2 |
| 600–900mm | 26 | 3.4 |
| Month | %Ndfa Pinto Peanut | Pinto Peanut Plot Fixed N (kg ha−1) * | %Ndfa White Clover | White Clover Plot Fixed N (kg ha−1) * | Plantation Fixed-N (kg ha−1) ** |
|---|---|---|---|---|---|
| December (2014) | 62 ± 9 | 3.2 ± 1.1 | NA | 2.4 | |
| January (2015) | 61 ± 7 | 9.9 ± 0.9 | NA | 7.5 | |
| February | 50 ± 7 | 7.2 ± 0.6 | NA | 5.4 | |
| March | 31 ± 6 | 3.9 ± 0.6 | NA | 2.9 | |
| April | 34 ± 3 | 1.3 ± 0.3 | 13 ± 0.7 | 0.3 ± 0.1 | 1.3 |
| May | NA | 0 | 28 ± 1.6 | 0.7 ± 0.3 | 0.5 |
| June | NA | 0 | 32 ± 1.0 | 1.4 ± 0.1 | 1.1 |
| July | NA | 0 | 41 ± 0.5 | 2.4 ± 0.0 | 1.8 |
| August | NA | 0 | 51 ± 3.2 | 7.3 ± 1.6 | 5.5 |
| September | NA | 0 | 69 ± 3.7 | 10.9 ± 1.6 | 8.1 |
| October | 32 ± 6 | 1.6 ± 0.9 | 47 ± 1.9 | 2.7 ± 1.7 | 3.2 |
| November | 50 ± 5 | 8.8 ± 1.0 | 39 ± 4.1 | 0.3 ± 1.1 | 6.8 |
| December | 50 ± 9 | 7.4 ± 0.9 | NA | 5.6 | |
| January (2016) | 52 ± 8 | 10.7 ± 1.5 | NA | 8.0 | |
| Cumulative (2015 year) | 40.3 | 26.1 | 49.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rose, T.J.; Kearney, L.J. Biomass Production and Potential Fixed Nitrogen Inputs from Leguminous Cover Crops in Subtropical Avocado Plantations. Agronomy 2019, 9, 70. https://doi.org/10.3390/agronomy9020070
Rose TJ, Kearney LJ. Biomass Production and Potential Fixed Nitrogen Inputs from Leguminous Cover Crops in Subtropical Avocado Plantations. Agronomy. 2019; 9(2):70. https://doi.org/10.3390/agronomy9020070
Chicago/Turabian StyleRose, Terry J., and Lee J. Kearney. 2019. "Biomass Production and Potential Fixed Nitrogen Inputs from Leguminous Cover Crops in Subtropical Avocado Plantations" Agronomy 9, no. 2: 70. https://doi.org/10.3390/agronomy9020070
APA StyleRose, T. J., & Kearney, L. J. (2019). Biomass Production and Potential Fixed Nitrogen Inputs from Leguminous Cover Crops in Subtropical Avocado Plantations. Agronomy, 9(2), 70. https://doi.org/10.3390/agronomy9020070