Phosphate Uptake is Correlated with the Root Length of Celery Plants Following the Association between Arbuscular Mycorrhizal Fungi, Pseudomonas sp. and Biochar with Different Phosphate Fertilization Levels


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Set Up
2.2. Biomass, Nutrient, Root Growth and Colonization Analysis
2.3. Statistical Analysis
3. Results
3.1. Root Growth and Morphology of Plants
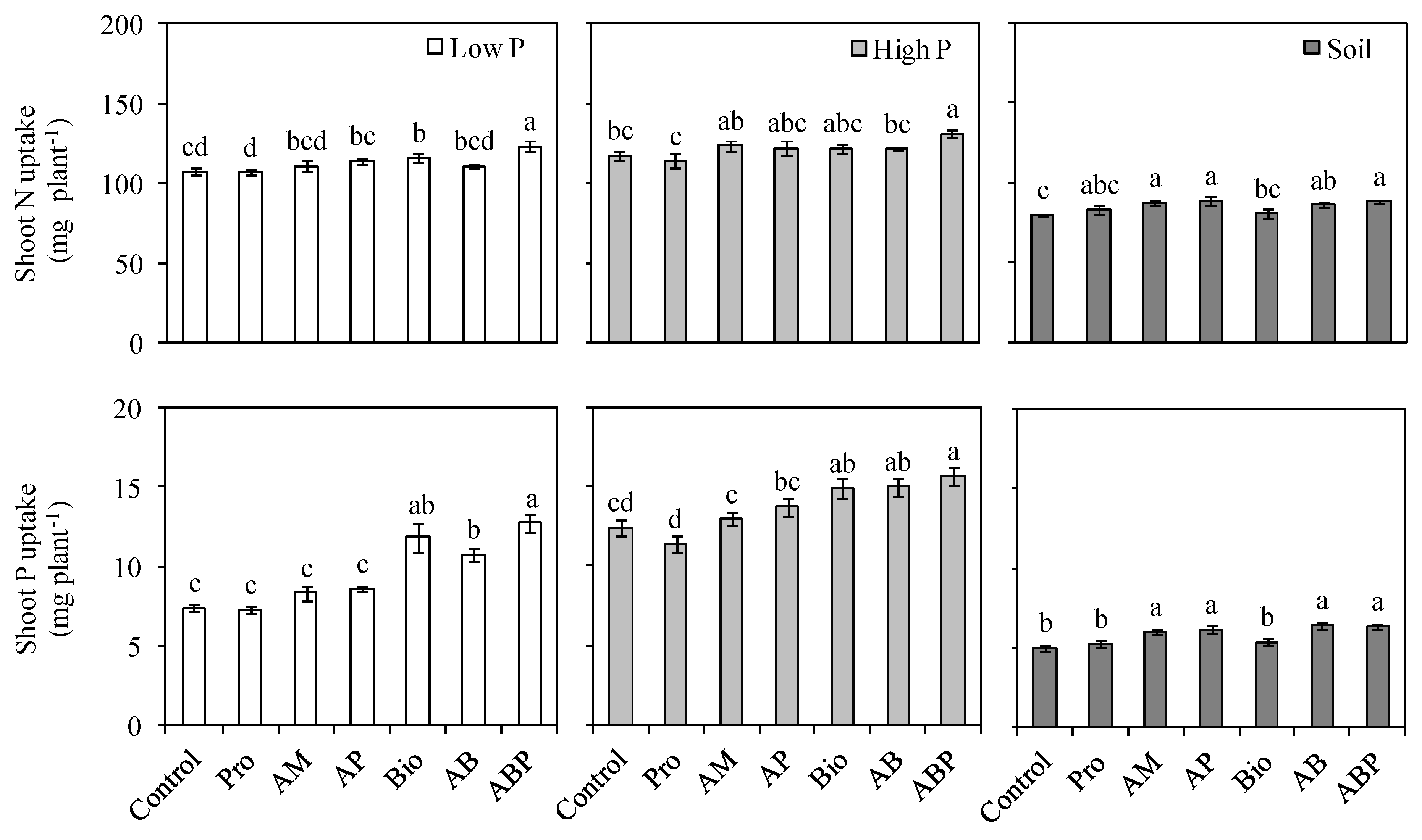
3.2. Phosphorus and Nitrogen Uptake
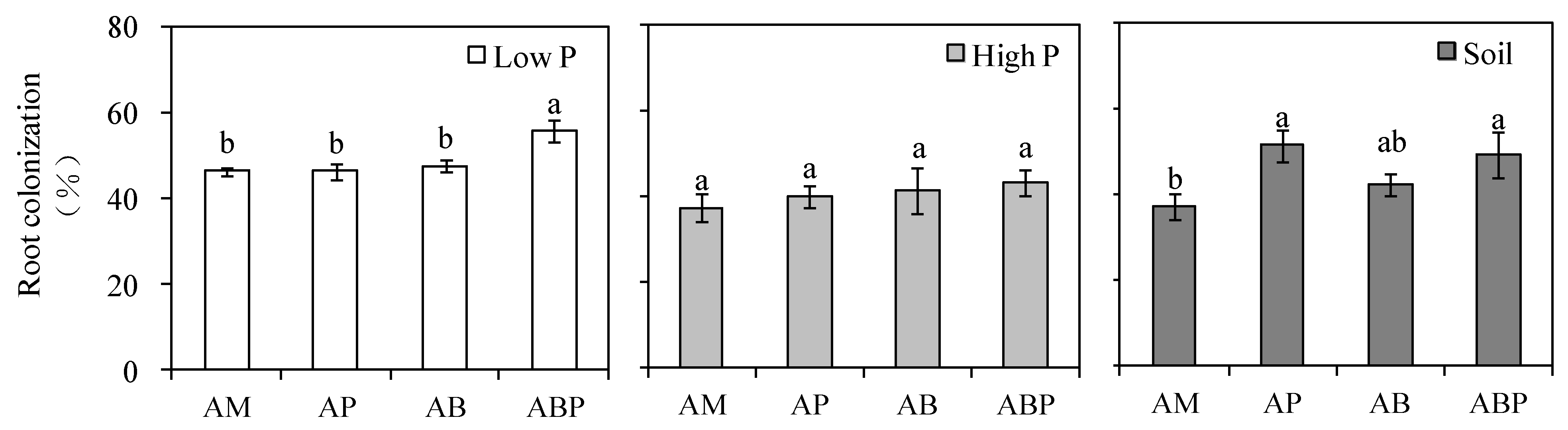
3.3. Plant Growth and Root Colonization Rates of AM Fungi
4. Discussion
4.1. Root Morphology and Phosphorus Uptake
4.2. Nutrient Uptake and Plant Growth Responses
4.3. Colonization of AM Fungi Rate
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malhotra, S.K. Handbook of Herbs and Spices; Woodhead Publishing: Cambridge, UK, 2006; Volume 3, pp. 317–336. [Google Scholar]
- Tanwar, A.; Yadav, K.; Prasad, K.; Aggarwal, A. Biological Amendments on Growth, Nutritional Quality, and Yield of Celery. Int. J. Veg. Sci. 2013, 19, 228–239. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.R.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bucking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Estrada, B.; Aroca, R.; Barea, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 2013, 2013, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, K.; Basheer, C.; Bajic, V.B.; Swarup, S. Enhancement of plant-microbe interactions using a rhizosphere metabolomics-driven approach and its application in the removal of polychlorinated biphenyls. Plant Physiol. 2003, 132, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Chithrashree, A.C.; Udayashankar, S.; Chandra Nayaka, M.S.; Srinivas, R.C. Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. Oryzae. Biol. Control 2011, 59, 114–122. [Google Scholar] [CrossRef]
- Dey, R.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R. Isolation and characterization of PGPR and their effect on growth, yield and nutrient content in wheat (Triticum aestivum L.). Biocatal. Agric. Biotechnol. 2014, 3, 121–128. [Google Scholar] [CrossRef]
- Barea, J.M.; Azcón-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Horii, S.; Ishii, T. Effect of arbuscular mycorrhizal fungi and their partner bacteria on the growth of Sesame plants and the concentration of Sesamin in the seeds. Am. J. Plant Sci. 2014, 5, 3066–3072. [Google Scholar] [CrossRef]
- Froehlich, A.; Buddrus-Schiemann, K.; Durner, J.; Hartmann, A.; von Rad, U. Response of barley to root colonization by Pseudomonas sp. DSMZ 13134 under laboratory, greenhouse, and field conditions. J. Plant Interact. 2012, 7, 1–9. [Google Scholar] [CrossRef]
- Kolton, M.; Meller, H.Y.; Pasternak, Z.; Graber, E.R.; Elad, Y.; Cytryn, E. Impact of Biochar Application to Soil on the Root-Associated Bacterial Community Structure of Fully Developed Greenhouse Pepper Plants. Appl. Environ. Microbiol. 2011, 77, 4924–4930. [Google Scholar] [CrossRef] [PubMed]
- Douds Jr, D.D.; Lee, J.; Uknalis, J.; Boateng, A.A.; Zieglerulsh, C. Pelletized Biochar as a Carrier for AM Fungi in the On-Farm System of Inoculum Production in Compost and Vermiculite Mixtures. Compost Sci. Util. 2014, 22, 253–262. [Google Scholar] [CrossRef]
- Li, Z.F.; Wang, Q.; Zhang, W.J.; Du, Z.L.; He, X.H.; Zhang, Q.Z. Contributions of nutrients in biochar to increase spinach production: A pot experiment. Commun. Soil Sci. Plant Anal. 2016, 47, 2003–2007. [Google Scholar] [CrossRef]
- Ishii, T.; Kadoya, K. Effects of Charcoal as a Soil Conditioner on Citrus Growth and Vesicular-Arbuscular Mycorrhizal Development. J. Jpn. Soc. Hortic. Sci. 1994, 63, 529–535. [Google Scholar] [CrossRef]
- Ijaz, M.; Sher, A.; Sattar, A.; Naeem, W.H.M. Cumulative effect of biochar, microbes and herbicide on the growth and yield of wheat (Triticum aestivum L.). Pak. J. Life Soc. Sci. 2015, 13, 73–78. [Google Scholar]
- Ogawa, M.; Okimori, Y.; Krull, E. Pioneering works in biochar research, Japan. Soil Res. 2010, 48, 489–500. [Google Scholar] [CrossRef]
- Solaiman, Z.M.; Blackwell, P.; Abbott, L.K.; Storer, P.; Krull, E. Direct and residual effect of biochar application on mycorrhizal root colonisation, growth and nutrition of wheat. Aust. J. Soil Res. 2010, 48, 546–554. [Google Scholar] [CrossRef]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil—Concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Conversa, G.; Bonasia, A.; Lazzizera, C.; Elia, A. Influence of biochar, mycorrhizal inoculation, and fertilizer rate on growth and flowering of Pelargonium (Pelargonium zonale L.) plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Hammer, E.C.; Balogh-Brunstad, Z.; Jakobsen, I.; Olsson, P.A.; Stipp, S.L.S.; Rillig, M.C. A mycorrhizal fungus grows on biochar and captures phosphorus from its surfaces. Soil Biol. Biochem. 2014, 77, 252–260. [Google Scholar] [CrossRef]
- Abiven, S.; Hund, A.; Martinsen, V.; Cornelissen, G. Biochar amendment increases maize root surface areas and branching: A shovelomics study in zambia. Plant Soil 2015, 395, 1–11. [Google Scholar] [CrossRef]
- Bruun, E.W.; Petersen, C.T.; Hansen, E.; Holm, J.K.; Hauggaardnielsen, H. Biochar amendment to coarse sandy subsoil improves root growth and increases water retention. Soil Use Manag. 2014, 30, 109–118. [Google Scholar] [CrossRef]
- Singh, A.P.; Singh, S.K.; Singh, C.M. Effect of Biochar Along with Plant Growth Promoting Rhizobacteria (PGPR) on Growth and Total Dry Matter Yield of Rice. J. Pure Appl. Microbio. 2015, 9, 1627–1632. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Ngwene, B.; Gabriel, E.; George, E. Influence of different mineral nitrogen sources (NO3−-N vs. NH4+-N) on arbuscular mycorrhiza development and N transfer in a Glomus intraradices–cowpea symbiosis. Mycorrhiza 2013, 23, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–505. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Berta, G.; Fusconi, A.; Trotta, A.; Scannerini, S. Morphogenetic modifications induced by the mycorrhizal fungus Glomus strain E3 in the root system of Allium porrum L. New Phytol. 1990, 114, 207–215. [Google Scholar] [CrossRef]
- Schellenbaum, L.; Berta, G.; Ravolanirina, F.; Tisserant, B.; Gianinazzi, S.; Fitter, A.H. Influence of endomycorrhizal infection on root morphology in a micropropagated woody plant species (Vitis vinifera L.). Ann. Bot. 1991, 68, 135–141. [Google Scholar] [CrossRef]
- Cruz, C.; Green, J.J.; Watson, C.A.; Wilson, F.; Martins-Loucao, M.A. Functional aspects of root architecture and mycorrhizal inoculation with respect to nutrient uptake capacity. Mycorrhiza 2004, 14, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pan, Q.; Chen, F.; Yan, X.; Liao, H. Effects of co-inoculation with arbuscular mycorrhizal fungi and rhizobia on soybean growth as related to root architecture and availability of N and P. Mycorrhiza 2011, 21, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Olmo, M.; Villar, R.; Salazar, P.; Alburquerque, J.A. Changes in soil nutrient availability explain biochar’s impact on wheat root development. Plant Soil 2016, 399, 333–343. [Google Scholar] [CrossRef]
- Alburquerque, J.A.; Salazar, P.; Barrón, V.; Torrent, J.; Campillo, M.C.; Gallardo, A.; Villar, R. Enhanced wheat yield by biochar addition under different mineral fertilization levels. Agron. Sustain. Dev. 2013, 33, 475–484. [Google Scholar] [CrossRef]
- Hammer, E.C.; Forstreuter, M.; Rillig, M.C.; Kohle, J. Biochar increases arbuscular mycorrhizal plant growth enhancement and ameliorates salinity stress. Appl. Soil Ecol. 2015, 96, 114–121. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Richardson, A.E.; Barea, J.; Mcneill, A.M.; Prigentcombaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Meyer, J.R.; Linderman, R.G. Response of Subterranean Clover to Dual Inoculation with Vesicular-Arbuscular Mycorrhizal Fungi and a Plant Growth-Promoting Bacterium, Pseudomonas putida. Soil Biol. Biochem. 1986, 18, 185–190. [Google Scholar] [CrossRef]
- Bianciotto, V.; Minerdi, D.; Perotto, S.; Bonfante, P. Cellular Interactions between Arbuscular Mycorrhizal Fungi and Rhizosphere Bacteria. Protoplasma 1996, 193, 123–131. [Google Scholar] [CrossRef]
- Sun, D.Q.; Hale, L.; Crowley, D. Nutrient supplementation of pinewood biochar for use as a bacterial inoculum carrier. Biol. Fertil. Soils 2016, 52, 515–522. [Google Scholar] [CrossRef]
- Mickan, B.S.; Abbott, L.K.; Stefanova, K.; Solaiman, Z.M. Interactions between biochar and mycorrhizal fungi in a water-stressed agricultural soil. Mycorrhiza 2016, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic C | Total N | Total P | Total K | Available N | Olsen-P | Available K | |
|---|---|---|---|---|---|---|---|
| * Biochar (g kg−1) | 52.09 | 4.88 | 0.83 | 15.98 | 0.005 | 0.162 | 9.60 |
| # Added to Substrates as Biochar (mg L−1) | 71 | 67 | 11 | 218 | 0.1 | 2.2 | 131 |
| Fertilizers Added to Substrates (mg L−1) | 0 | 150 | 30 or 60 | 200 | 150 | 30 or 60 | 200 |
| Commercial Substrates (mg L−1) | 30495 | - | - | - | 15.3 | 1.82 | 16.8 |
| Soil (mg kg−1) | 560 | - | - | - | 20.1 | 12.1 | 131 |
| Nutrition Additive (mg L−1) | - | 160 | 30/60 | 200 | 160 | 30/60 | 200 |
| Treatment | Shoot N Concentration (%) | Shoot P Concentration (mg g−1) | ||||
|---|---|---|---|---|---|---|
| Low P | High P | Soil | Low P | High P | Soil | |
| Control | 1.61 ± 0.05 a | 1.19 ± 0.03 | 3.06 ± 0.12 | 1.11 ± 0.06 cd | 1.26 ± 0.05 cd | 1.92 ± 0.07 b |
| Pro | 1.56 ± 0.08 a | 1.19 ± 0.03 | 3.00 ± 0.09 | 1.00 ± 0.03 d | 1.19 ± 0.04 d | 1.91 ± 0.09 b |
| AM | 1.57 ± 0.04 a | 1.20 ± 0.03 | 2.93 ± 0.08 | 1.18 ± 0.03 bc | 1.26 ± 0.04 cd | 2.01 ± 0.04 b |
| AP | 1.48 ± 0.04 ab | 1.20 ± 0.05 | 2.94 ± 0.07 | 1.12 ± 0.05 cd | 1.35 ± 0.07 bc | 2.05 ± 0.06 b |
| Bio | 1.36 ± 0.07 bc | 1.22 ± 0.03 | 3.04 ± 0.07 | 1.36 ± 0.03 a | 1.49 ± 0.07 ab | 2.04 ± 0.07 b |
| AB | 1.27 ± 0.04 c | 1.19 ± 0.02 | 3.01 ± 0.05 | 1.22 ± 0.02 b | 1.47 ± 0.04 ab | 2.26 ± 0.06 a |
| ABP | 1.34 ± 0.06 bc | 1.26 ± 0.02 | 2.88 ± 0.06 | 1.38 ± 0.03 a | 1.51 ± 0.06 a | 2.08 ± 0.06 ab |
| Treatments | Shoot Biomass (g plant−1) | Root Biomass (g plant−1) | Total Biomass (g plant−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Low P | High P | Soil | Low P | High P | Soil | Low P | High P | Soil | |
| Control | 6.7 ± 0.3 d | 9.9 ± 0.3 ab | 2.6 ± 0.1 c | 2.9 ± 0.1 cd | 3.6 ± 0.1 ab | 1.4 ± 0.1 | 9.6 ± 0.3 b | 13.5 ± 0.3 a | 4.0 ± 0.2 |
| Pro | 6.9 ± 0.3 cd | 9.6 ± 0.3 b | 2.8 ± 0.2 abc | 2.6 ± 0.1 d | 3.3 ± 0.1b | 1.5 ± 0.1 | 9.6 ± 0.2 b | 12.9 ± 0.3 b | 4.3 ± 0.1 |
| AM | 7.1 ± 0.3 cd | 10.3 ± 0.2 a | 3.0 ± 0.1 ab | 2.8 ± 0.1 d | 3.9 ± 0.2 a | 1.4 ± 0.1 | 9.9 ± 0.3 b | 14.2 ± 0.3 a | 4.4 ± 0.2 |
| AP | 7.7 ± 0.3 bc | 10.2 ± 0.2 ab | 3.0 ± 0.1 ab | 2.8 ± 0.2 d | 3.9 ± 0.1 a | 1.4 ± 0.1 | 10.5 ± 0.4 b | 14.1 ± 0.3 a | 4.5 ± 0.2 |
| Bio | 8.7 ± 0.5 ab | 10.0 ± 0.3 ab | 2.7 ± 0.2 bc | 3.4 ± 0.1 ab | 3.9 ± 0.2 a | 1.5 ± 0.2 | 12.0 ± 0.6 a | 13.9 ± 0.4 a | 4.2 ± 0.3 |
| AB | 8.8 ± 0.2 a | 10.2 ± 0.2 ab | 2.9 ± 0.1 abc | 3.2 ± 0.1 bc | 4.0 ± 0.2 a | 1.3 ± 0.1 | 12.0 ± 0.3 a | 14.2 ± 0.3 a | 4.2 ± 0.2 |
| ABP | 9.3 ± 0.4 a | 10.4 ± 0.3 a | 3.1 ± 0.1 a | 3.6 ± 0.2 a | 3.6 ± 0.2 ab | 1.4 ± 0.1 | 12.9 ± 0.6 a | 14.0 ± 0.3 a | 4.4 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, Y.; Xiao, Z.; Weinmann, M.; Li, Z. Phosphate Uptake is Correlated with the Root Length of Celery Plants Following the Association between Arbuscular Mycorrhizal Fungi, Pseudomonas sp. and Biochar with Different Phosphate Fertilization Levels. Agronomy 2019, 9, 824. https://doi.org/10.3390/agronomy9120824
Ning Y, Xiao Z, Weinmann M, Li Z. Phosphate Uptake is Correlated with the Root Length of Celery Plants Following the Association between Arbuscular Mycorrhizal Fungi, Pseudomonas sp. and Biochar with Different Phosphate Fertilization Levels. Agronomy. 2019; 9(12):824. https://doi.org/10.3390/agronomy9120824
Chicago/Turabian StyleNing, Yani, Zhiyong Xiao, Markus Weinmann, and Zhifang Li. 2019. "Phosphate Uptake is Correlated with the Root Length of Celery Plants Following the Association between Arbuscular Mycorrhizal Fungi, Pseudomonas sp. and Biochar with Different Phosphate Fertilization Levels" Agronomy 9, no. 12: 824. https://doi.org/10.3390/agronomy9120824
APA StyleNing, Y., Xiao, Z., Weinmann, M., & Li, Z. (2019). Phosphate Uptake is Correlated with the Root Length of Celery Plants Following the Association between Arbuscular Mycorrhizal Fungi, Pseudomonas sp. and Biochar with Different Phosphate Fertilization Levels. Agronomy, 9(12), 824. https://doi.org/10.3390/agronomy9120824