1. Introduction
Iodine (I) is an essential micronutrient, which plays a critical role in the human metabolism. It is a component of thyroid hormones thyroxine (T3) and tri-iodothyrosine (T4) [
1]. These hormones are involved in physiological body processes such as reproductive functions, growth, development, and metabolism [
2]. Inadequate uptake of I in the diet, sometimes colloquially referred to as “hidden hunger”, results in physiological disorders and biological function abnormalities called iodine deficiency disorders (IDD) such as enlarged thyroid gland (goiter) and cretinism [
3,
4]. Globally, 2 billion people are estimated to be deficient of I. In Africa, 332 million people are reported to have inadequate I intake [
5]. The food and nutrition technical assistance (FANTA II) [
6] study reported the total goiter rate (TGR) among school going children of age 6–12 years in Uganda at 31% with recent stakeholder study revealing that goiter cases are frequently seen in the community [
7]. This is substantially higher than the goiter rate of 5% threshold reported by Bimenya et al. [
8] to consider I deficiency as a public health problem.
Generally, the high prevalence of IDD has been attributed to the limited access to I-rich foods such as sea foods as well as inadequate amount of I in edible plant parts [
9]. In an effort to increase the intake of I, universal salt iodization (USI) has been widely implemented. However, despite its implementation for decades now, I deficiency still remains a major threat to the health and development of populations in Sub-Saharan Africa [
10]. Additionally, salt iodization has been associated with an increase in the occurrence of cardiovascular diseases as a result of excessive salt consumption [
11].
Following these limitations, the World Health Organization [
12] recommended reduction in salt content in the diet as well as searching for alternative approaches to increase the I content in the diet. One such novel strategy is agronomic biofortification, a technique of supplying micronutrients through soils or plant leaves to increase their contents in edible plant parts [
13]. Agronomic biofortification of staple crops is considered to be a cost-effective and sustainable approach to address IDD in resource constrained settings [
14].
A number of agronomic biofortification studies that focused on other micronutrients such as zinc and iron have yielded promising results using horticultural crops, such as tomato, spinach, and fruit [
15,
16]. However, several issues concerning agronomic I biofortification remain to be addressed. For instance, limited studies exist on the efficiency of I retention in crops, the effect of application rate, and the effect of form of application are yet to be conclusively studied. Additionally, most of the available studies have been done in pots and hydroponic culture systems in laboratory settings [
17,
18]. While past studies give initial suggestions of the effectiveness of agronomic biofortification, the use of controlled environments does not provide reliable estimates of how such applications would behave under open field conditions [
18], hence limiting the applicability of laboratory findings. Consequently, there is a need to move beyond the controlled environment to field trials. To this effect, few field trials have been conducted to assess the efficiency of different plants to absorb and accumulate exogenous I [
19,
20,
21]. If applied through the soil, the low mobility as well as physicochemical properties of soil can limit plant uptake efficiency of I. In addition, soil application can also affect translocation efficiency. For instance, Weng [
21] observed a decreasing trend of retention of soil applied I from root, leaf, to fruit. On the other hand, biofortification of I through foliar application is known to be more effective than soil application. Lawson et al. [
20] reported higher efficiency of foliar applied I compared to soil application in butterhead lettuce. However, the authors noted that the effectiveness of foliar spray can be increased by use of surfactant in the spray solution. This study, therefore, used farmer field-based trials for evaluating the efficiency of cabbage (
Brassica oleracea var. capitata) and cowpea (
Vigna unguiculata L.) crops, both commonly consumed as leafy vegetables in Northern Uganda, to take up and accumulate foliar applied I. Specifically, this study aimed to (1) evaluate the effect of I form on absorption of I in edible parts of cabbage and cowpea leafy vegetable crops; and (2) determine the effect of I application rate on I content in edible parts of cabbage and cowpea vegetable crops.
2. Materials and Methods
2.1. Study Area and Experimental Set Up
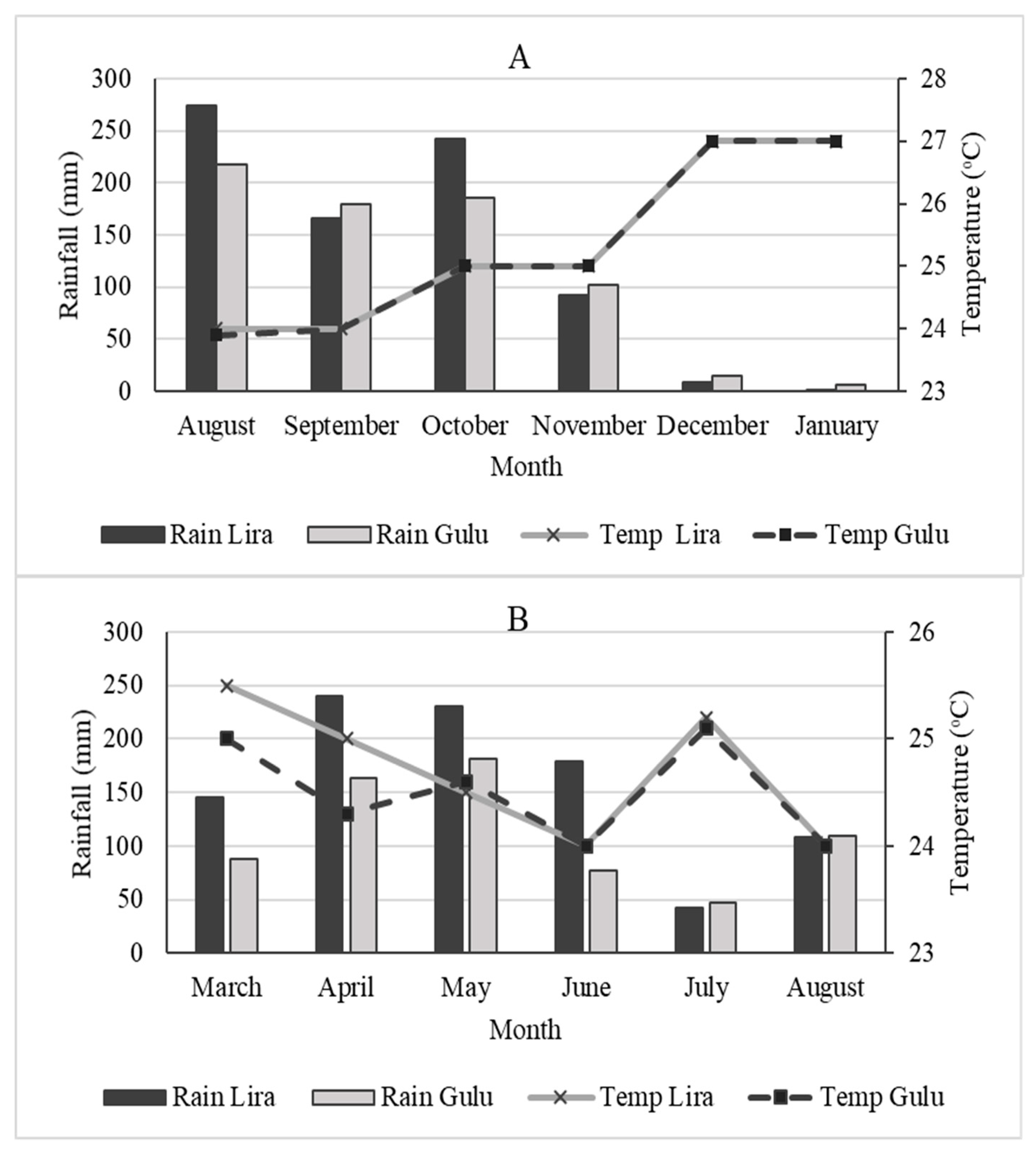
Field trials were conducted at Gulu (02°46.742ʹ N, 032°20.097ʹ E) and Lira (02°18.075ʹ N, 032° 56.690ʹ E) in Northern Uganda, between September 2017 and January 2018, and from March to August 2018. Total monthly rainfall and average air temperature for both locations and planting seasons were computed (
Figure 1). A randomized complete block design was used with 8 treatments and 3 replications for each of the Cabbage and Cowpea crops. A factorial arrangement consisting of two I forms i.e., potassium iodide (KI) and potassium iodate (KIO
3), and four application rates (0, 5, 10, and 15 kg I ha
−1) was used in the experiments.
Cabbage and Cowpea seeds were disinfected by soaking in 1% potassium permanganate (KMnO4) solution for 20 min and thoroughly rinsed with clean water before planting. Baraka F1 hybrid Cabbage seeds were sown into a nursery bed at a row spacing of 15 cm by continuous and later thinned to 15 × 5 cm. After four weeks, seedlings with 5–6 leaves were transplanted at 45 × 60 cm into 7 m2 main experimental plots. Cowpea seeds were planted directly in the experimental plots at 30 × 20 cm on 7 m2 plots.
2.2. Initial Soil Conditions
Prior to implementing the treatment structure, soil samples were taken at 0–15 cm depth and analyzed for organic matter (OM), soil pH, total nitrogen (TN), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), I, and soil texture at the National Agricultural Research Laboratories (NARL), Kawanda (Uganda) (
Table 1). The air-dried soil samples were pounded, sieved through 2 mm to remove any debris, then subjected to physical and chemical analysis following standard methods described by Okalebo et al. [
22] Soil pH was measured in a soil water solution ratio of 1:2.5, organic matter was measured using potassium dichromate wet acid oxidation method, N was determined by Kjeldhal digestion, extractable P was estimated following Bray P1 method, while exchangeable bases were determined from an ammonium acetate extract using flame photometry (K
+) and atomic absorption spectrophotometry (Ca
2+, Mg
2+). Soil texture was determined using the Bouyoucos (hydrometer) method. The soil analysis results (
Table 1) generally indicated that the soils in the two sites were of average to low soil fertility with low pH of 4.5 on average. Agricultural lime was applied at a rate of 2.5 t ha
−1 to raise the soil pH from 4.5 to above the required critical value of 5.2, while NPK (20:5:5) was applied at a rate of 150 kg ha
−1 according to NARL recommendations. Iodine content of the soil was generally lower in Gulu site (1.3 mg kg
−1) compared to Lira experimental site (5.6 mg kg
−1). Iodine content in the water used in foliar application was not measured. However, the same water source was used for all the experimental plots (treatments) and therefore the effect of the background iodine in water is expected to have a negligible effect on the results.
2.3. Foliar Application of Iodine
A fixed amount of KI and KIO
3 required for each treatment was weighed using a digital pocket scale BL-100 (American Weigh Scales, Cumming, GA, USA) and dissolved in 500 ml of water to reach a concentration in the spray solution that will provide a dose of I per ha equivalent to a practical application of this amount at a spray volume of 600 L ha
−1 for each treatment [
23]. Application was done using automated hand sprayers. The fertilizers (KI and KIO
3) were mixed with dimethoate pesticide (Rogo, 30% EC) at a rate of 1.5 ml/L of water. Iodine fertilizer applications were split into four doses for each crop, each at ¼ of the reported total KI or KIO
3 dose per ha. Foliar application of cowpeas was started at 21 days after planting (DAP) and the subsequent three applications were carried out at a five-day interval. For cabbages, initial foliar application was carried out at 30 days after transplanting (DAT) and the subsequent three applications were done at a 14-day interval. Sampling of cowpea plants was done five days after the last spraying, while cabbage plants were sampled on average 14 days after the last quarter of foliar application. Spraying was only done on the days when no rainfall was anticipated. Generally, little rainfall was received in November and December of 2017 and January of 2018 at both locations (
Figure 1). As a result, experimental plants were irrigated using a watering can.
2.4. Sampling, Pre-Processing, and Analysis
Cowpea and cabbage sample plants were collected using 0.5 m2 quadrants. For each plot, a quadrant was thrown into the middle of the plot and plants within the quadrant were uprooted, packed in polythene bags, labeled, and transported to Bioscience analytical laboratory at the National Crop Resources Research Institute (NACRRI), Namulonge for freeze drying and milling. At the laboratory, cowpea and cabbage leaves were plucked and washed thoroughly with running tap water to imitate a common domestic cleaning and spread on the table for excess water to evaporate. Composite leaf samples for each plot were then packed in polythene bags and transferred into freezers maintained between minus 40 °C to –80 °C for preservation prior to lyophilization.
Lyophilization was performed using Labconco (Labconco Corporation, Kansas City, MO, USA) following manufacturers’ instructions. Lyophilized samples were pulverized using a motorized seed grinder fitted with a sieve and a collector (Osaw Industrial Products pvt. Ltd, Ambala, India). Pulverized samples were weighed and packed in airtight polythene bags for storage prior to I analysis. Laboratory analysis was conducted using Inductively Coupled Plasma-Mass Spectrometer, ICP-MS (PerkinElmer Inc., Waltham, MA, USA) after I extraction using tetra methyl-ammonium hydroxide (TMAH). Chemicals used included: milliQ water, liquid argon (high purity, 99.99%), 25% (m/m) tetra methyl-ammonium hydroxide (TMAH, analytical grade, 99.9999% purity). A commercial stock iodine standard (1000 mg L−1), internal standard stock solution containing 1000 ± 3 mg L−1 tellurium (Te) in 2% HNO3 and 0.2% HF were procured from Innochem (Ghent, Belgium). The certified reference material ERM-BD151 (skimmed milk powder) and ERM-BB422 (fish muscle) were analyzed for quality control.
Analysis was done at the laboratory of Analytical Chemistry and Applied Eco chemistry (Ghent University, Belgium). Standard procedure ‘NBN EN 15111’ for determination of I in foodstuffs by ICP-MS was followed. This consisted of TMAH extraction followed by I determination by ICP-MS. Approximately 0.5 g of the sample was weighed into a 50 mL centrifuge tube followed by adding 10 mL of 5% TMAH, capped and vortexed for one minute. The mixture was incubated for three hours in a water bath set at 90 °C. The mixture was cooled to room temperature, vortexed for one minute, diluted to a total volume of 50 mL using milliQ water, weighed, and the final mass was recorded. The solution was centrifuged at 8000 rpm for 10 min and the supernatant was filtered through a 0.45 μm syringe filter to remove large particles that could clog the nebulizer or sample probe during ICP-MS analysis.
A PerkinElmer NexION 350D ICP-MS system (PerkinElmer Inc., Waltham, MA, USA) was used for analysis. The concentration of I in the samples was determined after external calibration using 0, 1, 2.5, 10, 25, 50, 100, and 100 µgL−1 I standards prepared in the same TMAH matrix as the sample extracts. As part of quality control, two reference materials (skimmed milk powder ERM-BD151 and fish muscle ERM-BB422) were analyzed and the percentage recovery of I was determined. Moreover, various levels of iodate (at 5 and 10 µg L−1) were spiked to the extracts of the plant samples and reference materials and the spike recovery was determined. All results were expressed as iodine concentration (mg kg−1) per dry weight of the sample.
2.5. Statistical Analysis
The data obtained were analyzed using the SAS statistical software package version 9.4 [
24]. The GLM procedure was used to conduct the analysis of variance (ANOVA) to compare mean concentration of I in cowpea and cabbage samples at various application rates and forms. Means were compared using Tukey’s HSD test. In all cases, the level of statistical significance was set at 5% (
p ≤ 0.05).
4. Discussion
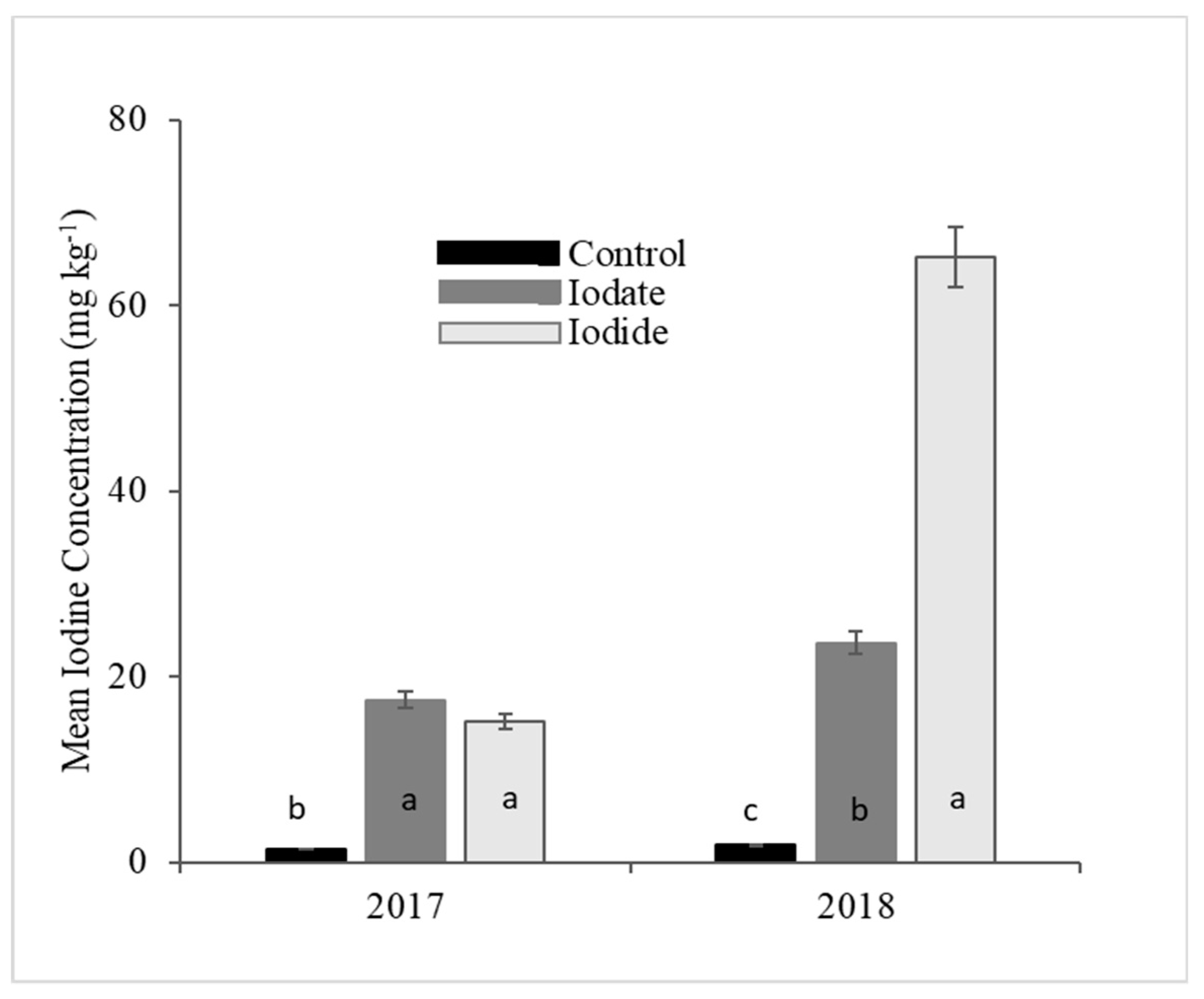
This study demonstrates significant effects of two I forms on the level of I absorption and retention in cowpea and cabbage leaves. However, the influence of the I form was not consistent for cabbage across the experimental period (2017 and 2018). Overall, the concentration of I in cabbage and cowpea was significantly higher when KI was applied. These results are in line with those obtained in the I biofortification of several other studies [
23,
25,
26]. While explaining the difference in the efficacy between KI and KIO3, Lawson et al. [
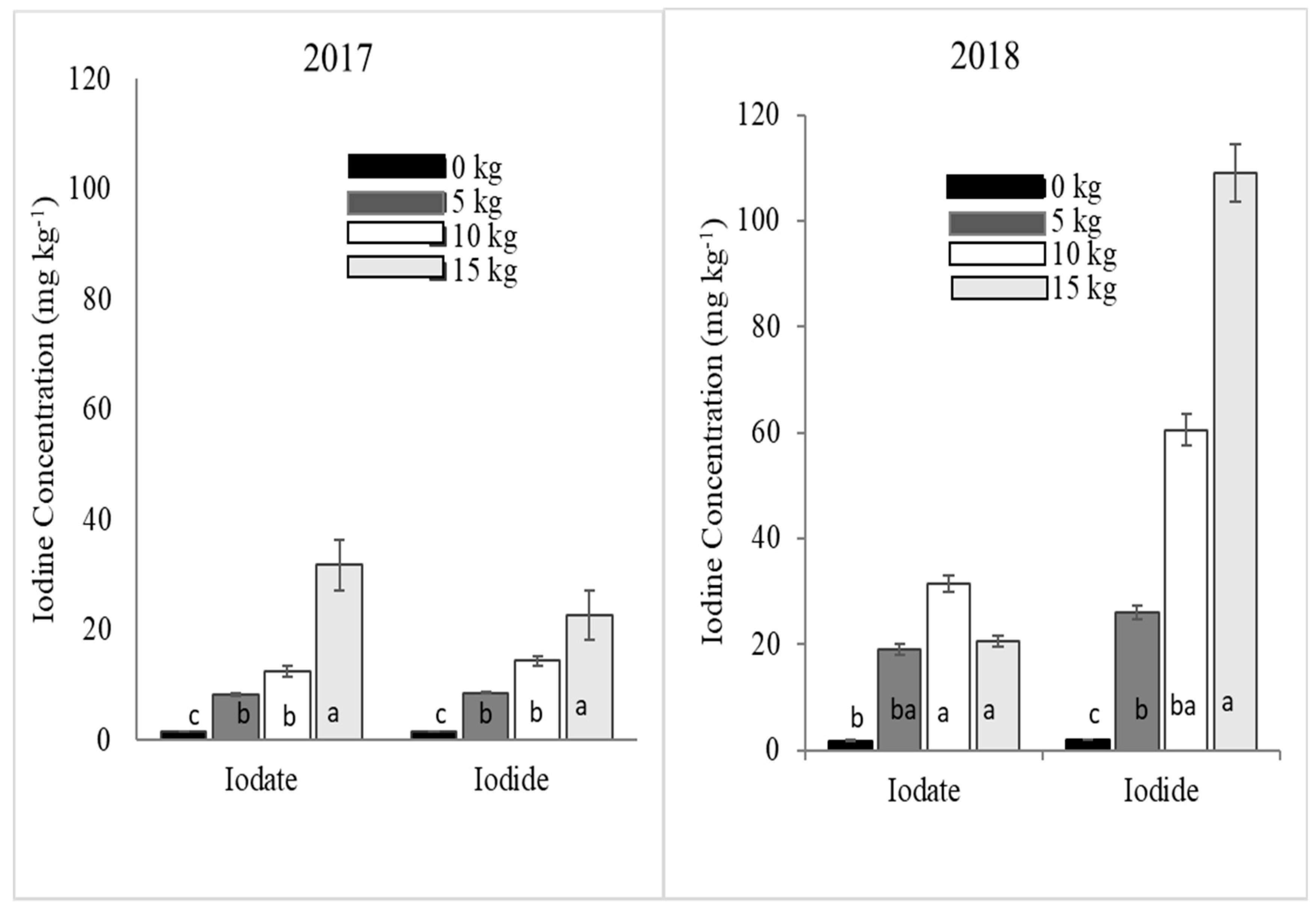
23] stated that the higher efficacy of KI could comparatively be due to its lower point of deliquescence and smaller anion size.
The increase in I concentrations in cabbage leaf tissue when both iodide and iodate were applied was generally low as compared to cowpea. This could be because cabbage has a more closed canopy structure, making complete wetting of the crop difficult compared to cowpea with relatively open canopy structure. Furthermore, the thick waxy layer in cabbage makes foliar application in general challenging due to lower absorption of water by the leaves. As a result, the level of I absorption in cabbage was lower compared to cowpea. The other possible reason for relatively low absorption of I by cabbage could be attributed to the fact that cabbage plants produce enzymes, halogen methyltransferases, which are responsible for the volatilization of the absorbed I from their leaves. Saini et al. [
27] reported that I is phytotoxic and its volatilization by aerial plants is seen as a detoxification mechanism. It is also noted that the activity of these enzymes to dissipate I increases with temperature, changes in developmental phases, and under stressful conditions [
26]. In the present study, cabbage I concentration was lower in 2017 compared to 2018. Therefore, the high temperature in the last three months of 2017 could have increased the dissipation rate of I in cabbage trials compared to I concentrations of cabbage planted in 2018.
The concentration of I in cowpea samples was generally low in the control treatments. This is an indication of the low availability of plant available soil I in the study area, 1.3 mg kg
−1 for Gulu and 5.6 mg kg
−1 for Lira experimental location. Similar observations were made by Lawson et al. [
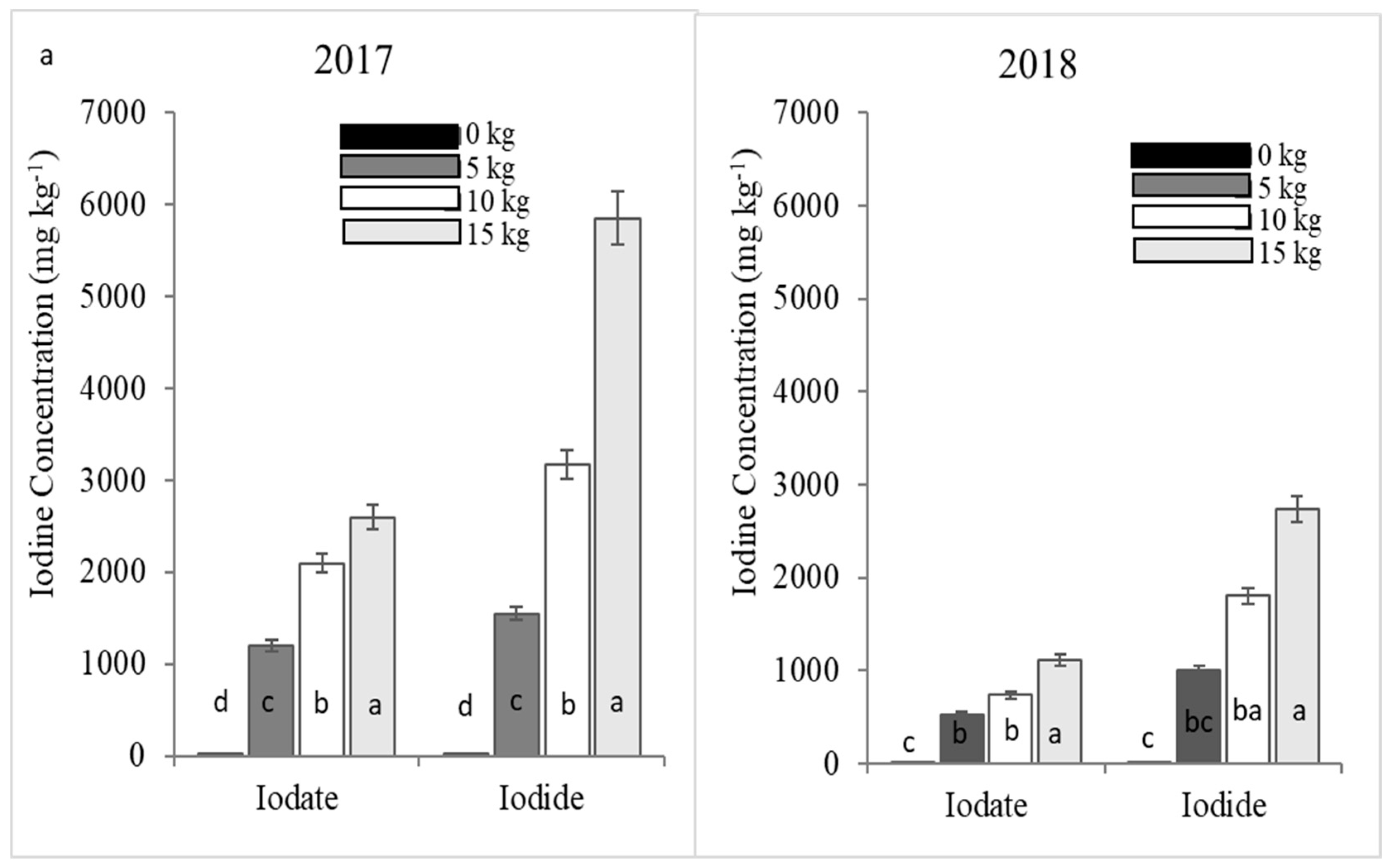
23] in Germany where lettuce I concentration in the check plots was generally low and far below the recommended daily I intake. In the treated plots, there were significant differences in the mean concentrations of I, which demonstrated the efficiency of cowpea in uptake of foliar applied I from both carriers. This finding is consistent with the study of Caffagni et al. [
25] on tomato fruits and potato tubers. Medrano et al. [
28] also reported a positive result when I was applied to soil as KIO
3 at doses of 7.5 kg ha
−1, 10 mg kg
−1 of soil in pots, and leaf spray with KI at 0.5 kg ha
−1. In the current study, potassium iodide exhibited a higher efficiency than potassium iodate in increasing I concentration in cowpea. This finding corresponds with studies of Zhu et al. [
29] and Voogt et al. [
30] where I concentrations in plant tissues (spinach and lettuce) were up to five times higher with iodide than with iodate form. Lawson et al. [
23] also reported that foliar application of potassium iodide led to a higher concentration of I in lettuce when compared to potassium iodate. Krzepiłko [
31], in a similar study, demonstrated that radish and lettuce seedlings grown in the presence of potassium iodide had a higher I content than those grown with potassium iodate. Findings from the current study, however, contradict that of Zhu et al. [
29] where at a lower I concentration (1 µM) in nutrient solution, I concentration in the leaves of spinach plants treated with iodate exceeded that in leaves treated with iodide. This could be because I was supplied in soil and taken through the roots instead of the leaves. In addition to the difference in the uptake mechanisms between roots and leaves, there is a potential for sorption of I to the soil hence reducing root efficiency.
The efficiency with which plants absorb and retain I seem to be highly dependent on plant species. This is evident from the current study where large differences in the concentration of I were observed between cabbage and cowpea. This confirms earlier assertions made by Weng et al. [
32] where the absorption of I in various vegetables was determined by plant type and physiological action of the plant. Similarly, Weng et al. [
21] observed that the I uptake was controlled by the I fertilizer application rate as well as the individual plant characteristics. Fernández and Brown [
33] summarized that physiological status of the plant and species characteristics such as leaf shape, leaf chemistry, and leaf surface characteristics differ from plant to plant and significantly influence absorption of foliar applied fertilizers.
Generally, I concentration in both cowpea and cabbage plants increased with the rate of I fertilizer applied similar to observations by Zhu et al. [
29] and Lawson et al. [
23] Unlike for cowpea, no detrimental effect of high I application rates was observed for cabbage growth. Many studies reported that once absorbed and accumulated in different plant organs, I is not stable. Medrano et al. [
28] reported that some plant species dissipate I as methyl iodide (CH
3I) using the enzymes halide ion methyltransferase. This corresponds with earlier findings of Itoh et al. [
34] that dissipation of CH
3I from the sprouting leaves of
R. sativus, T. aestivum, and
O. sativa grown hydroponically increased with increasing concentrations of iodide. Therefore, the concentration of I in the tissues of such plants reduces with time by the activity of the halide enzyme, methyltransferase [
28,
35]. The methyl halide emission potential for plant tissues changes significantly from planting to harvest time [
36]. Most importantly is the low uptake efficiency of I by cabbage compounded by thick layering of cabbage leaves can possibly reduce absorption of I initially applied compared to cowpea.
Phytotoxic symptoms observed in cowpea few days after spraying with both potassium iodide and iodate could be due to smaller unit of leaf mass compared to cabbage since similar dosages were applied. Because of relatively smaller unit of leaf mass, the dosage for cowpea seemed high and this was accompanied by the short time interval between the split dose applications. Although the plants could regain after a few days before the next spray, slight defoliation, chlorosis, and necrotic spots on the older leaves remained visible after foliar spray of both I fertilizers at 10 kg I ha
−1. Immense defoliation and eventual plant death were observed when potassium iodide was applied at 15 kg I ha
-1. Toxicity of iodide has been attributed to its photo-oxidation to free I in the presence of light, causing chlorophyll destruction [
37]. This finding is consistent with observations on butterhead lettuce [
23] and spinach [
29,
38].
5. Conclusions
The present study showed that cowpea had higher I concentration than cabbage. Form of I had a significant influence in the concentration of I in the edible tissues of both cowpea and cabbage. In cowpea, iodide was more efficient than iodate. For cabbage, the form of I carrier was inconsistent across experimental years. In 2018, I content in cabbage tissue was 72% higher when fertilizer was applied as iodide than iodate. However, for the experiment conducted in 2017, there was no significant difference between the two forms. Further investigations using foliar method with more site-years could provide conclusive results on the potential of cabbage to absorb and retain I. Additionally, biofortification experiments that embed biomass measurement in the calculation of total I in crop samples could provide accurate information on the efficiency of I uptake of different crop species. Application rate had a significant influence on the concentration of I in all the crops studied. However, phytotoxic effects were exhibited in cowpea at application rates ≥ 5 kg I ha
−1. Generally, I concentrations obtained at all application rates in the two crops are more than sufficient to meet the daily adult human required intake of 150 μg. Specifically, our results indicate that consumption of 100g of KIO
3 biofortified cabbage at 5 kg ha
−1 would be an equivalent of 820 μg while I content of KI biofortified cabbage and both KI and KIO
3 biofortified cowpea at rates ≥ 5 kg I ha
−1 would be higher than the upper limit of 1100 μg [
39] per 100g of food consumed. Future studies could determine the optimal application rate less than 5 kg ha
−1 and calculate the contribution to the recommended daily intake of I at each application rate. In addition, future studies could assess the stability of I absorbed when cabbage and cowpea are subjected to various heat treatment processes.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}