Abstract
The objective of this study was to estimate the direct and indirect effects of agricultural, morphological, physiological, and root traits of popcorn lines, under two conditions of water supply: full irrigation (FI) and drought stress (DS). A complete randomized block design with three replications was used to evaluate the following traits: expanded popcorn volume per hectare (EPV), 100-grain weight (100GW), mean ear length, mean ear diameter, number of grains per row (NGR), prolificacy (PRO), anthesis to silking interval, tassel length, number of tassel branches, plant height, shoot dry matter, green index (SPAD), and canopy temperature depression (CTD), root angle (RA), tap root volume (TRV), and crown root volume (CRV). Analyses of variance and path analysis, and subsequent partitioning of direct and indirect effects were performed. For the traits EPV, PRO, CTD, RA, CRV and TRV, the genotype x environment interaction was significant. Drought stress had significant effects on EPV (−7.15%), 100GW (−23.52%), SPAD (−29.31%), CTD (87.15%), RA (24.54%), and CRV (44.89%). The traits NGR and SPAD were found to be decisive for the expression of EPV. The exploitation of these traits by indirect selection is expected to induce increments in EPV in environments under DS and FI.
1. Introduction
The seed market offers cultivars with high potential for grain production, but to ensure high yields, optimized management conditions are necessary, with appropriate nutrient supply and pest, disease and weed control, as well as adequate water supply. In intensive and family agriculture, abiotic stresses are obstacles to the growth and full development of maize [1,2,3,4].
Drought, considered the most limiting abiotic factor for plant growth and development, compared to other stress types, has a strong effect on grain yield, limiting maize production in both tropical and subtropical regions [3,5,6,7,8,9,10]. In the South-Center and Southeast region of Brazil, corn production is concentrated in the second growing season, from February to June [11]. During this season, water limitations caused by heavy rainfall variations can drastically reduce grain yield, implying the importance of using drought-tolerant maize genotypes [11,12]. These effects are influenced by sensitivity levels of the genotypes, stress intensity and duration and the plant development stage during drought stress [9].
Despite the harmful effects of drought stress and the recurrent occurrence of irregular rainfall distribution, little effort has been made by research groups and breeding companies to develop genotypes with increased drought tolerance. In the case of popcorn, a crop that turns over about 2 billion dollars annually in the USA, to date there has been no research related to the selection of drought-adapted genotypes. Consequently, the exploitation of variability in popcorn becomes fundamental to access so far unexplored sources of drought tolerance in plant breeding. For this, the North Fluminense State University “Darcy Ribeiro” (UENF) has a germplasm collection containing accessions from temperate and tropical climatic adaptated regions of Latin America, as well as inbred lines from improved elite materials in Brazil [13]. In this regard, a better understanding of the morphoagricultural and physiological responses of plant genetic resources is required and some strategies to optimize plant selection in water-deficit environments are proposed.
Stress caused by drought at the pre-anthesis phenological stage causes harmful effects on the meiotic stage of reproductive cells, affecting the subsequent development of the male gametophyte, inducing pollen sterility [14], reflecting in fewer grains [9]. For the evaluation of germplasm with a view to drought tolerance, we consider the traits associated with grain yield under stress, and at reproductive structures, i.e., shorter interval between male and female anthesis [11,15,16], late leaf senescence [15,17], increased prolificacy [11,15,18], and lower number of tassel branches [6,15]. More recently, physiological traits were highlighted, namely: (i) canopy temperature [9,10]; (ii) estimate green biomass by remote sensing tools based on the use of irradiation [3,9]; (iii) green leaf color index (SPAD) [9]; and, (iv) root morphology and depth [19]. The phenological stage of male pre-anthesis is the most recommended period for the evaluation of more productive genotypes under dry conditions [8,9,10,11], and the pre-anthesis period coincides with the moment of shortage of rainfall in corn crops in Brazil [11]. Meiosis-related processes, which include pollen viability, in addition to zygote formation and initiation of grain filling, are biological processes highly sensitive to water stress [20].
Thus, the following scientific questions arise: (i) Which of the trait(s) associated with drought stress is(are) most appropriate for indirect selection of drought-tolerant germplasm? (ii) Under well-irrigated conditions, the direct and indirect effects of trait selection associated with greater tolerance to drought stress are the same as in a stressed environment?
In this scenario of questions, it is known that a high correlation between two traits may be the result of the effect of a third or even a group of traits on the former [21]. It is not possible to observe whether the correlations are estimated based on real and direct cause and effect relationships [22]. In this respect, Wright′s [23] path analysis allows conclusions about direct and indirect effects by partitioning the correlations of a group of (explanatory) traits in relation to a given trait considered to be the most important (main).
Therefore, the objective of this study was to estimate the effects of direct and indirect influence of agricultural, morphological, physiological and root architecture traits of popcorn genotypes under two water regimes, on the main variable: expanded popcorn volume per hectare (EPV), considered a supertrait, for representing the product of the two main economically important traits of the crop: popping expansion and grain yield [24]. In practice, this research concludes with the indication of secondary characteristics that can be used for the selection of more productive genotypes in dry conditions, that is, more agronomically efficient genotypes in water use [25], favoring the practice of a more sustainable agriculture in the use of water resources. More productive and drought-adapted plants imply a lower ratio of grain yield per unit of available or evapotranspirated water [26].
2. Materials and Methods
2.1. Plant Material, Experimental Design and Cultural Treatment
Twenty popcorn lines (S7) of the Active Germplasm Bank of the UENF were evaluated, derived from germplasm adapted to tropical climatic (L61, L63, L65, L69, L70, and L71—derived from population ‘BRS-Angela’) and temperate/tropical germplasm (P1, P5, P6 and P7—derived from the commercial hybrid ‘Zélia’; P2 and P3—from compound CMS-42; P4—from South American races; P8 and P9—L54, L55 and L59—from population ‘Beija-Flor’; and L75 and L76—derived from population ‘Viçosa’).
These popcorn lines were evaluated under two water regimes, i.e., under drought stress (DS) under full irrigation (FI), by irrigation management. In the drought-stressed environment, irrigation was interrupted at the phenological stage of pre-anthesis, 15 days before male flowering (10 June 2016), while in the environment without water restriction irrigation was maintained until physiological grain maturation, to ensure complete replacement of crop evapotranspiration.
To synchronize the onset of stress with anthesis, the lines were separated in two maturation groups, based on previously studied male flowering data: (i) early (L54, L59, P2, P3, P4, P5, P6, P7, P8, and P9) and late (L55, L61, L63, L65, L69, L70, L71, L75, L76, and P1). Thus, staggered sowing was carried out, with the first sowing on 19 April for the late-maturing group and the second on 25 April for the early-maturing group in 2016. The male flowering date occurred precisely on 24 June 2016, representing 67 and 61 days after sowing, for both late and early genotypes, respectively.
The experiments, with and without water stress, were carried out at the UENF Experimental Station, Rio de Janeiro, Brazil (Latitude 21°42′48” S, Longitude 41°20′38”) during the dry season (April to August) to allow the effective imposition of stress. The experimental design was a randomized complete block with three replications per water condition. Each plot consisted of four rows of 4.40 m, spaced 0.80 m between rows and 0.20 m between plants (23 plants per row). The evaluated area of each plot consisted of 8.00 m2 of the central rows. Fertilization at sowing consisted of 30 kg N ha−1, 60 kg P2O5 ha−1 and 60 kg K2O ha−1, and sidedressing was applied 30 days after sowing, consisting of 100 kg N ha−1.
Drip irrigation was applied (flow 2.3 mm h−1), with one Katif dripper per plant at the recommended spacing (0.20 × 0.80 m). During the cultivation period, irrigation levels were applied (mm) in the well-irrigated condition (FI) as well as in the drought-stressed (DS) condition (Table S1). Rainfall events were recorded by the weather station installed in the experimental area. The total amount of water applied in early and late maturation genotypes was 60 mm for the WS condition, while WW condition plants received a total amount of 138 mm. There was rainfall, totaling 133 mm (Table S1). The temperature and relative humidity during the crop cycle ranged from 12 to 37 °C and 23% to 97%, respectively, and mean solar radiation was 20.35 MJ m−2 day−1 (≅1300 µmol m−2 s−1) (Figure S1).
The soil of the experimental station was classified as Yellow Dystrophic Latosolic Fragipan Argisol and contains high clay and silt levels. The soil water contents at field capacity and permanent wilting point were 38.21% and 28.41%, respectively. The soil water potential was monitored with Decagon MPS-6 tensiometers (Decagon, Pullman, Washington, 99163, USA) (Figure 1). The soil of the FI water condition was maintained at field capacity (−10 kPa), while under DS, the soil reached the permanent wilting point (−1500 kPa) 12 days after male anthesis (Figure 1).
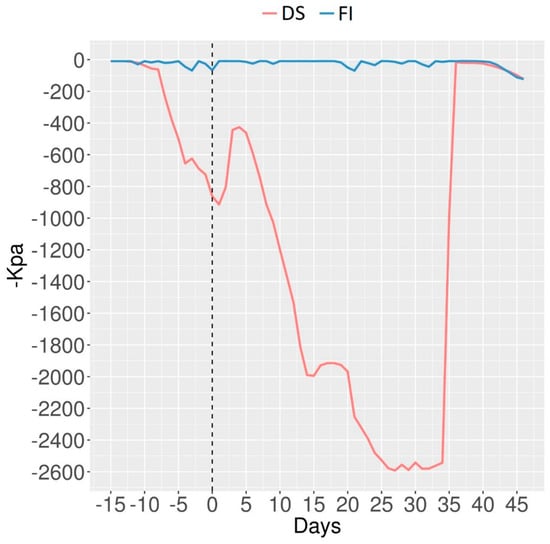
Figure 1.
Soil water potential (−kPa) of full irrigation (FI) and drought stress (DS) during irrigation suspension. The male flowering date occurred precisely on 24 June 2016, representing 67 and 61 days after sowing, for both late and early genotypes, respectively, and is represented on day zero.
2.2. Traits Evaluated
Harvesting was undertaken 120 days after sowing. The main trait expanded popcorn volume per hectare (m3 ha−1) (EPV), was estimated by multiplying the mean plot yield with popping expansion. To this end, grain yield per hectare (GY) was estimated (Kg ha−1), and corrected to a moisture content of 13%, and the popping expansion (PE) was measured for the mass of 30 g grain, microwave-irradiated in a paper bag for popping, at 1000 W, for 2.25 min. The popcorn volume was quantified in a beaker (2000 mL). Popping expansion was determined as the quotient of the obtained popcorn volume by grain weight (mL g−1).
For path analysis, the traits of the first order were 100-grain weight (100GW), ear length (EL), ear diameter (ED), number of grains per row (NGR) and prolificacy (PRO). Based on all plants per plot, 100GW was measured by weighing (g) three sub-samples of 100 grains, and PRO determined as the quotient of the number of harvested ears by the total number of plants per plot. A random sample of six healthy plants per plot was taken to estimate EL and ED, quantified with a pachymeter (cm) and NGR, determined by counting.
The traits representing the secondary-order factors consisted of: anthesis to silking interval (ASI), tassel length (TL), number of tassel branches (NTB), plant height (PH), shoot dry matter (SDM), green index (SPAD), canopy temperature depression (CTD), root angle (RA), crown root volume (CRV) and tap root volume (TRV).
The trait PH was measured from the ground to the flag leaf (cm); SDM was determined by weighing (g) after oven drying at 70 °C for 72 h; ASI was calculated by the difference between the mean values of the male and female flowering dates (days); TL was measured from the lowest tassel rachis node to its extremity (cm); and NTB was determined by counting. The trait associated with leaf green intensity (SPAD) consisted of the average of three readings in the mean third of the third leaf, counted from the apex and below the flag leaf, with a portable chlorophyll meter Minolta SPAD-502 (soil plant analysis development) (Minolta, Osaka, 590-8551, Japan). The trait SPAD was evaluated 35 days after the onset of DS (soil water potential around −1500 kPa—Figure 1). These traits were measured in a random sample of six useful plants.
The trait CTD (°C) was calculated as the difference between the air temperature around the leaf and that of the canopy (Tair −Tcanopy). The canopy temperature was estimated by means of thermographic images, obtained in a FLIR i50® thermograph (FLIR Systems AB, Täby, 187 66 Sweden—19,600 pixels) and processed by software FLIR Tools/Tools + ® program. Images were taken of the two middle rows of each plot between 12:00 and 14:00 (Soil water potential: −1500 kPa—July 08—Figure 1), at 0.60 m above the canopy.
The root architecture traits were determined according to the methodology proposed by Trachsel et al. (2011) [27]. After the harvest, the soil of the water condition under FI and DS was watered with 50 mm irrigation, facilitating the mechanical removal of the plants with shovels. The intact root system of two plants per plot was removed in a soil cylinder (diameter 40 cm, depth 25 cm). The cylinders were washed until complete removal of the soil. The following traits were measured: root angle (RA), measured with a protractor and expressed in relation to the soil (°); tap root volume (TRV), calculated as the product of the number of support roots and the mean lateral root density; and crown root volume (CRV), calculated as the product of the number of crown roots and the mean lateral root density.
2.3. Analysis of Variance and Estimates of Genetic Parameters
Analyses of individual variance were performed (, where Yij is the observation of the ith genotype of the jth block; μ is the general constant, gi the genotype effect; bj the block effect; and εij the experimental error) and of combined variance for different water condition (, where: observation of the ith genotype in the the jth environment in the kth block; is the general constant; gi: the random effect of the ith genotype; : effect of the kth block within environment j; : fixed effect of the jth environment; : random effect of the interaction between the ith genotype with the jth environment; and : random experimental error associated with observation ).
Estimates of the genetic components were calculated by the mean squares of the individual analysis of variance. Heritability based on the means of the genotypes was estimated by the expression: , where: . = genotypic variance and . = residual variance. The standard deviation was estimated by: . The coefficient of genetic variation was calculated by: . The coefficient of experimental variation was calculated as: .
2.4. Path Analysis
The values of each trait per plot were standardized and then subjected to analysis of variance to generate the genotype correlation matrix (rg). The rg matrix was subjected to the collinearity test of Montgomery et al. (1981) [28]. Multicollinearity occurs when the sample observations of the explanatory variables, or their linear combinations, are correlated. According to the Montgomery and Peck Table (1981) [28], we have that for condition number less than 100 there is low data collinearity; For condition number greater than 100, moderate to strong collinearity is required, and path analysis is not recommended. The significance of rg was tested by the t-test, at 5% and 1% probability, with n-2 degrees of freedom, between all pairs of combinations.
The rg were used for the estimation of the path coefficients, partitioning them in direct (pjy) and indirect (rijpjy) effects, by estimating the regression equations, under both water regimes (FI and DS), according to the model of primary and secondary traits, which explain the variation of Y (EPV). Thus, rij is the correlation coefficient of trait i with j, and pjy is the direct effect (or path coefficient) of trait j in the final product Y. The direct and indirect effects of the secondary traits on each primary trait were calculated in the same way, by applying the appropriate model and the system of equations derived therefrom.
The effect of the five primary traits on the main variable (EPV) was considered: 100GW (P01), EL (P02), ED (P03), NGR (P04), and PRO (P05). However, the mode of action of the 10 secondary traits on the primary traits and the main variable EPV was considered as: ASI (S06), TL (S07), NTB (S08), PH (S09), SDM (S10), SPAD (S11), CTD (S12), RA (S13), CRV (S14), and TRV (S15). These assumptions show the interrelationships between the variables, taking into account the logical and additive relationship between the traits (Figure 2). The resolution was determined by the system of normal equations X′Xβ = X′Y, where X′X is a non-singular matrix of correlations between explanatory variables; β is a column vector of path coefficients and; XY is a column vector of correlations between the main and explanatory variables.
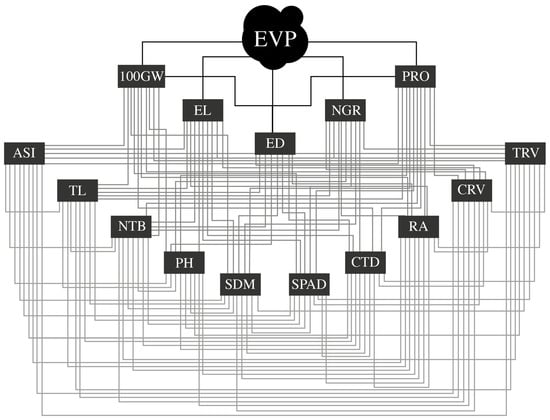
Figure 2.
Causal chain graph showing the interrelationship of the main variable expanded popcorn volume per hectare (EPV) with the primary variables—100-grain weight (100GW), ear length (EL), ear diameter (ED), number of grains per row (NGR), and prolificacy (PRO)—and secondary—range between male and female flowering (ASI), tassel length (TL), tassel branch number (NTB), plant height (PH), shoot dry matter (SDM), green index (SPAD), canopy temperature depression (CTD), root angle (RA), crown root volume (CRV) and tap root volume (TRV).
The coefficient of determination for the analysis of explanatory variables on the main variable and between the secondary traits on the primary is given by: . The residual effect is expressed by: . The analyses were performed with the software Genes [29].
3. Results
3.1. Genetic and Environmental Effects of Traits in Full Irrigation (FI) and Drought Stress (DS)
In the FI and DS environments, genetic variability was detected among the evaluated genotypes, for all traits (Table 1).

Table 1.
Summary of analyses of variances in popcorn lines under full irrigation (FI) and drought stress (DS), genotype–environment (GE) interactions, general means, standard deviations (SD), coefficients of environmental variation (CVe), coefficients of genetic variation (CVg) and heritability (H2), for agricultural, morphological, physiological and root architecture traits.
The overall mean of expanded popcorn volume per ha (EPV) in the fully irrigated water condition (FI) was 74.79 m3 ha−1 (grain yield: 2548.08 Kg ha−1 × popping expansion: 29.35 g mL−1), while the mean of this trait in the drought-stressed condition (DS) was 23.82 m3 ha−1 (grain yield—1139.11 Kg ha−1 × popping expansion—20.91 g mL−1). These mean values show a reduction of 68.15% (Figure 3) in EPV caused by drought stress (DS), compared to the condition with full water supply (FI). For this trait, CVe had a similar pattern in both water conditions. Under DS, the CVg was higher i.e., 39.83%, compared to 25.07% under FI. The heritability (H2) of EPV was higher under DS, with an estimated order of 89.50%, against 72.21% under FI (Table 1).
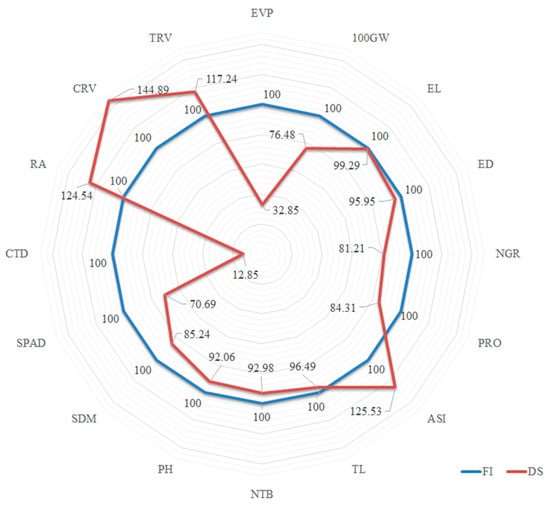
Figure 3.
Relative percentages of the means evaluated in the water condition with drought stress (DS) in relation to the means without water restriction (FI), for the main variable EPV and for the agricultural, morphoagricultural, physiological and root traits, evaluated in popcorn.
Under DS, the agricultural traits of the first order 100GW, NGR and PRO were considerably decreased, with lower mean estimates (23.52%, 18.79% and 15.69%, respectively) (Figure 3) than under FI. The marked reductions caused by water deficiency in the DS condition were also observed in the physiological traits (second chain), in particular for SDM (14.75%) and SPAD (29.31%) (Figure 3). Under DS, the mean CTD value increased in the order of 87.15%, in relation to the FI condition (Figure 3). This shows that under DS, the atmospheric temperature is lower than or equal to that of the canopy. Compared to the control (FI), the means of the root traits RA, CRV and TRV were 24.53%, 44.89% and 17.24% higher, respectively, under DS (Figure 3).
Under FI, CVe ranged from 5.21% (PH) to 43.57% (ASI), and under DS, from 5.03% (PH) to 88.36% (CTD). The trait CTD is influenced by atmospheric variations (wind, cloudiness and humidity) and by the time of image acquisition. On the other hand, the CVg ranged from 7.76% (ED) to 81.77% (ASI) under FI and from 8.60% (ED) to 144.13% (CTD) under DS (Table 1).
High H2 values were observed under both FI and DS (Table 1). Under FI, the H2 estimates ranged from 68.76% (100GW) to 95.96% (PH), and under DS, from 53.78% (PRO) to 95.48% (CRV), respectively (Table 1). Non-significant GE interactions were detected for the characteristics 100GW, EL, ED, NGR, ASI, TL, NTB, PH, SDM, and SPAD. Significant GE interactions occurred for EPV, PRO, CTD, RA, CRV, and TRV (Table 1).
3.2. Direct and Indirect Effects of the First-Order Explanatory Variables on the Variable Expanded Popcorn Volume Per Hectare in FI and DS
Under FI, the traits with significant total effects on the variable EPV were: EL (0.53), ED (0.47) and NGR (0.88) (Table 2). Among these, only NGR had a high direct effect (1.06) on EPV. The traits EL and ED, with zero direct effect on EPV, have marked effect of NGR, in the partitioning of indirect effects (Table 2).
Table 2.
Path analysis of the genotype correlations in components of direct and indirect effects, collinearity test, determination coefficient, and residual effect between the trait EPV and first-order agricultural traits evaluated in popcorn lines under FI and DS.
Under DS, the traits EL (0.71), ED (0.45), NGR (0.92), and PRO (0.65) had significant overall effects on EPV (Table 2). In the partitioning of effects, the traits EL, ED and PRO had direct negative effects on EPV (−1.16, −1.00 and −0.32, respectively). These traits revealed the marked action of trait NGR on all its effects on EPV. Only trait NGR had a high positive direct effect (2.65) on the main variable (Table 2).
The comparison of the total effects of traits between FI and DS indicated a similar behavior. The traits 100GW, EL, ED, and NGR expressed similar values with the same direction of total effects on EPV (Table 2). Only the variable PRO had a total effect in the opposite direction in the different water conditions, i.e., a value of −0.42 under FI and 0.65 under DS. In both water conditions, the total effect of trait 100GW on EPV was not significant.
The path coefficient under DS explained 88% (R2 = 0.88) of the variation in EPV, with a residual effect of 34% (Table 2). Under FI, 100% of the variation in EPV was explained, expressed by the high coefficient of determination in the model (R2 = 1.21) and a zero residual effect. Values above 1.00 and/or below −1.00 may occur when regression analysis estimates are obtained for standardized data [30]. Under both water regimes, the setting of the model proved reliable to explain the genetic effects, reflecting the high contribution of the explanatory traits used in the model related to EPV. The data quality to estimate the path coefficients is also ensured by the collinearity values (Table 2), in which DS (73.37) and FI (50.36) were considered low (<100) [28].
3.3. Direct and Indirect Effects of Second-Order Variables on the First-Order Variable Number of Grains per Row (NGR) in FI and DS Environments
Since the direct and indirect effects of the first-order variables on the main variable (EPV) were only promising by path analysis via NGR, the other partitionings with the traits 100GW, EL, ED and PRO were discarded. The complete table of the partitioning of the genotype correlations in components of direct and indirect effects (path analysis) obtained between the first- and second-order agricultural traits is found in the supplementary material (Table S2). The main results in the path analysis in both water conditions were observed by the traits TL, PH and SPAD. In Table 3, we highlight these traits in italics for easier observation of the main results.
Table 3.
Pathway analysis of the partitioning of the genotype correlations in components of direct and indirect effects obtained between first-order agricultural traits and second- order morphological, physiological and root traits evaluated in popcorn lines under FI and DS.
Under DS, the total effect of NGR was significant on TL (0.67), PH (0.48) and SPAD (0.44) (Table 3). These three traits, under the aforementioned water regime, had direct effects on NGR (in the order of 0.55, 0.69 and 0.70, respectively).
Under FI, trait NGR stood out with significant associations with TL (0.79), PH (0.61), CTD (0.44), and CRV (−0.46) (Table 3). Among these four traits, the effect of TL on NGR occurred due to direct effects (1.17) of the first on the second trait. The other three (PH, CTD and CRV) had an effect on NGR, mainly due to indirect effects via TL, (0.48, 0.31 and −0.40, respectively). The indirect and less intense effects via SPAD, on the total effects of PH on NGR, are also worth mentioning.
In both water conditions, the path coefficients (R2) were significant (p-value of 1%) (Table 3), indicating a reliable setting of the model to explain the genetic effects, reflecting the high contribution of the second-order traits used in the model related to each first-order trait.
3.4. Direct and Indirect Effects of the Second-Order Explanatory Variables on the Main Variable Expanded Popcorn Volume per Hectare (EPV), via NGR, in FI and DS Environments
This section describes only the direct and indirect effects of the traits associated with NGR, i.e., under DS, on TL (0.67), PH (0.48) and SPAD (0.44), and under FI, on TL (0.79), PH (0.61), CTD (0.44), and CRV (−0.46) (Table 3). In both water conditions, the total effect of CRV on EPV was not significant, making further partitioning superfluous. The other traits had significant total effects and high direct effects. The complete table of the partitioning of genotype correlations in components of direct and indirect effects (path analysis) between the second-order explanatory variables and the main variable EPV is found in the supplementary material (Table S3). The main results in the path analysis in both water conditions were observed by the traits SPAD. In Table 4, we highlight these traits in italics for easier observation of the main results.
Table 4.
Path analysis of the partitioning of genotypic correlations in components of direct and indirect effects obtained between the main trait and the first-order agricultural traits and morphological, physiological, and second-order root traits evaluated in popcorn lines under FI and DS.
Under DS, the total effect of TL on EPV was significant (0.57), although it had a low direct effect on the latter (0.18) (Table 4). These effects were pronounced by the indirect action via ASI (0.99) and SDM (0.41). The magnitude of these effects was directed by NGR (1.80). Under FI, trait TL had a significant total effect on EPV (0.56), and a high direct effect (0.86) on the same, with a lower residual effect (−0.59). Trait NGR had the highest effects (0.85) on EPV of all first-order traits (Table 4).
Under DS, trait PH had a significant total effect on EPV (0.51), with a strong direct effect (1.85) on this variable (Table 4). The residual effect (1.35) was lower than the total effect on EPV; thus, the association between these two variables may be relevant. These effects were intensified by the effect determined by NGR (1.29). Under FI, the trait PH had a high total significant effect on EPV (0.59), in spite of having a low direct effect on EPV (−0.22), and negative residual effect (−0.65). Thus, the association between these two variables is not relevant. Trait NGR was responsible for the direction of the strongest effects (0.66) on EPV (Table 4).
Under DS, trait SPAD had a significant total effect on EPV (0.45) and a strong direct effect (1.00) on this variable, associated to a low residual effect (0.40) (Table 4). The association between these two variables may be relevant. Trait NGR had the strongest action by indirect effect (1.13) via SPAD on EPV. Under FI, SPAD had a significant total effect on EPV (0.53), a clear direct effect on EPV (1.88) and a low residual effect (0.18). Trait NGR (0.20) accounted for the highest magnitude of effects on EPV.
Under DS, the total effect of trait CTD on EPV was not significant (−0.02) (Table 4), making further partitioning superfluous. Under FI, CTD had a significant total effect on EPV (0.60) and a direct effect on EPV (1.06). The residual effect found among the above traits was 0.61. Trait NGR (0.48) accounted for the strongest effects on EPV (Table 4).
4. Discussion
Several studies deal with correlations and GE interaction between maize traits evaluated with and without drought stress [11,31,32,33,34], however, no scientific papers were found reporting similar results for popcorn. In general, the values of CVg were higher than the estimates of CVe in both water conditions (Table 1). These values also demonstrate that the experimental procedures were adequate [30].
The estimates of the heritability (H2) were higher under DS than FI, for EPV (89.50% versus 72.21%) and its components 100GW (90.44% versus 68.76%) and NGR (91.05% versus 90.1%) (Table 1), indicating greater reliability in the selection based on these traits in the water condition with drought, and because the DS condition promotes greater expression of the genetic variance. The proportion of the phenotypic variance of genetic origin is explained by H2 [21,35]. Thus, the selection of superior genotypes under DS can be maximized, providing greater genetic gains, mainly for EPV.
The GE interaction occurs when the responses of the genotypes evaluated in the different water conditions are differentiated, as observed for the traits EPV, PRO, CTD, RA, CRV, and TRV (Table 1). The GE interaction interferes with selection gains and the recommendation of cultivars [35]. The non-significance of the GE interaction observed for 100GW, EL, ED, NGR, ASI, TL, NTB, PH, SDM, and SPAD (Table 1) allows the conclusion that selection these traits in one of the environments can be effective for simultaneous genetic gains in both.
The reduction of 68.15% in trait EPV (Figure 3) can be attributed to the significant decreases in 100GW (23.53%), NGR (18.79%) and PRO (15.69%). The minor reduction in ear size (EL) (0.71%) confirms that there was no decrease in the mean of this trait under DS which was, however, the case for number of ears, as well as grain weight and number of grains produced. In an experiment conducted under similar conditions to this study, mainly in terms of the moment of irrigation stop (male pre-anthesis), Cairns et al. (2012) [9] reported a mean grain yield reduction of 80% in maize lines. For the authors, the reduction in grain yield was mainly due to the decrease in the number of grains produced.
As additional information, it was observed that grain yield was reduced by 55.29% in the comparison between FI and DS, based on the estimates of 2548.08 Kg ha−1 and 1139.11 Kg ha−1, respectively (data not shown). In relation to popping expansion, the proportional decrease was 28.75%, in the comparison of the estimates of 29.35 g mL−1 and 20.91 g mL−1, respectively, under FI and DS (data not shown).
When analyzing the harmful effects of water deficiency on the second-order traits, it was observed that TL (−3.51%), NTB (−7.02%), PH (−7.94%) and SDM (−14.76%) were least affected. Irrigation was interrupted in the pre-anthesis period (Figure 1) and, according to Durães et al. (2004) [6], plants at this stage are near the end of vegetative development, and the traits mentioned were affected little by DS. Greater alterations or effects on the means were observed for ASI (25.53%), SPAD (29.31%) and CTD (87.51%) (Figure 3). The ASI values tend to increase under DS in the pre-flowering phase, since the number of days until anthesis remains relatively unchanged (compared to stress-free environments), but there is a delay in the style–stigma release of the ears, increasing the interval between male and female flowering. The means for SPAD decrease more rapidly under DS [9], since severe water deficiency can cause chlorophyll degradation in leaves [36]. Water stress induces leaf stomatal closure, reduces transpiration, and consequently decreases water loss [37]. As a result of stomata closure (reduction of transpiration), the leaf temperature rises [9,10,38], resulting in higher estimates of trait CTD (Table 1).
It is important to describe the biological impact on the root system of popcorn lines under water stress. There was an increase in the root angle (24.54%) in relation to the soil, indicating that the roots grew towards more vertical layers to capture water in distant profiles. According to Trachsel et al. (2011) [27], genotypes that have larger angles (about 90% in relation to the soil), in drought conditions, direct the root growth to the deeper layers of the soil and have advantages for finding water in depth. Morphological adaptation was also observed in CRV and TRV, which increased by 44.89% and 17.24%, respectively (Figure 3). A higher root/shoot ratio is considered a mechanism to increase drought tolerance [19,39], confirming our observations, in that CRV and TRV increased significantly and SDM remained unchanged. Molecular studies show that the accumulation of terpenoid phytoalexins in maize roots is associated with drought tolerance [19]. Promising traits for the selection of plants with higher water status in dry conditions are root weight density and root length density [40].
The success of path analysis depends on two main factors: composition of the causal diagrams and diagnosis of multicollinearity of the data [21,30]. Thus, it was verified that the diagram used, divided in two orders, allowing us to explain most of the 100% (R2) variation in EPV (Table 2). The use of standardized data to obtain the path coefficients can result in correlation components with values greater than unity and, therefore, R2 values can also reach estimates higher than one [21,30]. R2 values above 1.00 in path analysis can be observed, as highlighted by Silva et al. (2012) [30]. The collinearity test for the main and the primary variables resulted in 73.37 under DS and 50.36 under FI, and in the second order, which includes the main and secondary variables, in 90.77 under DS and 63.36 under FI (data not shown), with weak multicollinearity (<100) in all cases. This condition indicates no problems for path analysis, making the presented results reliable.
Under DS, the traits EL (0.71), ED (0.45), NGR (0.92), and PRO (0.65) (first order) had a marked total effect on EPV, suggesting them as promising for indirect selection. When considering only the direct effects of these traits on EPV, a more marked effect via NGR (2.65) is observed, indicating a promising path. The traits EL, ED and PRO had a negative direct effect on EPV, indicating that they are non-essential variables to increase EPV, and consequently, of no avail for selection. The positive effect of EL, ED and PRO was due to indirect effects via NGR (Table 2). The traits with high correlation with the main variable, however with unfavorable direct effects, indicate the absence of cause and effect. These traits cause no alterations in the main variable, and others can have a greater impact in terms of selection gain.
Along the path from the first order of variables to the second, considering only NGR, the traits TL (0.67), PH (0.48) and SPAD (0.44) are highlighted, for having a significant total effect on NGR (Table 2), which is favorable for indirect selection. These three traits were characterized by action through a direct effect on NGR. The results of this research pinpointed these three as potential traits for the selection of more drought-tolerant plants, via NGR, to increase EPV, based on their total positive effects, direct effects and important contributions of indirect effects by other traits.
Analyzing the differential expression of TL, PH and SPAD under DS, as well as knowledge about the crop and of the main traits associated with DS, one should carefully examine the results of TL and PH (little affected). Reports in the literature suggest that a larger tassel size and greater number of tassel branches tend to reduce grain yield, due to the reduction in sunlight interception by the flag leaves (self-shading) and competition for photoassimilates by different plant structures [31]. Plant height is frequently positively correlated with grain yield in popcorn [24,41] and maize [17,42]. On the other hand, higher SPAD values reflect an intensified activity of the photosynthetic machinery, particularly under DS [9,36]. Therefore, the trait SPAD can be considered as relevant for the selection of genotypes with higher EPV under DS.
Under FI, the first-order traits EL (0.53), ED (0.47) and NGR (0.88) had a positive overall positive effect on EPV, suggesting that indirect selection with these traits can be efficient. When considering only the direct effects on EPV, a marked effect is observed via NGR (1.06), reinforcing the importance of this trait. The traits EL and ED had no direct effect on EPV (Table 2), eliminating them from selection. The positive effect of EL and ED is provided by indirect effects via NGR (Table 2), indicating the absence of cause and effect in EPV.
Moving along the path from the first-order variables to the second group, under FI, the traits TL (0.79), PH (0.61), CTD (0.44), and CRV (−0.46) had significant total effects for NGR (Table 3). The effect of TL on NGR was mainly due to the direct effects (1.17) of the first trait on the second. The other traits (PH, CTD and CRV) expressed an effect on NGR, mainly due to indirect effects via TL. Lower indirect effects via SPAD on the total effect of PH on NGR were also observed. Trait TL is highly relevant in the selective process to increase EPV under FI. Interestingly, more robust plants, i.e., with tall stature and longer tassel length, were associated with higher EPV values. Under DS however, the trait TL must be treated with caution, since genotypes with shorter tassel length are recommended for selection [15].
Under FI, the association of SPAD with NGR was not very high, since leaf senescence in this water condition is slower (Table 1). According to Cairns et al. (2012) [9], the same genotypes have the highest SPAD values under FI and DS and under both water regimes, SPAD is positively correlated with grain yield. If the total and direct effects of SPAD on EPV are considered (Table 4), the former becomes promising for the discrimination of drought-tolerant plants. This information, along with the fact that SPAD is free of GE interaction (Table 1), supports the now established hypothesis that SPAD can be used for discrimination under FI as well. The easy measurement of SPAD (non-destructive and rapid—2 to 3 s per measurement) and the possibility of an indirect measurement of chlorophyll content by equipment of precision agriculture (satellites and aircraft equipped with remote sensors) will favor a rapid phenotyping of genotypes with greater drought tolerance.
The trait SPAD was evaluated 35 days after irrigation suspension; in absolute values, with general means of 45.99 under FI and 32.51 under DS (Table 1). This trait is an important tool in the diagnosis of plant stress [9,36]. In maize, SPAD values are correlated with chlorophyll content, photosynthesis and maximum quantum efficiency of photosystem II (Fv/Fm); high values of this trait indicate the non-degradation of chlorophyll and can be used to estimate the photosynthetic capacity under FI and DS [9,36]. A longer photosynthetically active canopy optimizes photoassimilate production, which is reflected in higher final grain yield [9,43,44]. Genotypes with late or late leaf senescence may increase carbon fixation [44]. A secondary role of the canopy with higher SPAD values is the storage of N (as a protein) and the subsequent effective remobilization of this N to the grain during physiological maturation [44].
The trait canopy temperature has been widely investigated to study the genotype response to DS [45], and it was shown that the genotypes selected as DS-tolerant in terms of absolute yield had the lowest canopy temperature in the time around midday, with high solar radiation [10,38]. This allowed the assumption that in more DS-tolerant genotypes (higher EPV), the leaf temperature would be lower in the time around midday, under a clear sky without clouds, as shown in thermal images, and reflected in CTD values. In this study, only one negative and non-significant estimate was found, between EPV and CTD under DS. Variation in corn leaf temperature in response to drought is closely related to biomass accumulation [37]. Under water stress conditions, higher leaf temperatures provide reduced transpiration as a water-saving strategy, which results in low relative leaf water content [46] and low cell membrane stability, supported by a significantly higher injury rate [47]. According to Zia et al. (2013) [10], the lower canopy temperature along with a satisfactory grain yield is probably associated with a more efficient or more developed, i.e., deeper root system. Similarly, Kaman et al. (2011) [38] demonstrate that higher grain production is associated with lower leaf temperature, combined with a more developed root system.
Therefore, the identification and understanding of root phenotypes associated with greater drought tolerance was studied to provide additional information for breeding programs. A root ideotype for maize plants was proposed by Gao et al. (2016) [48], based on three traits: steep, cheap, and deep. The authors appointed “cheap” as representing a low metabolic cost of soil exploration; “deep” referring to the capacity to reach deep soil regions and “steep” indicating angles of root growth of around 90º, in relation to the soil. These three traits are associated with a lower number of crown roots, which have a greater capacity of length development [48]. In this study, the results for the traits RA, CRV and TRV were not correlated with EPV. Trait CRV was negatively associated with EL, ED and NGR, denoting that a lower root volume (product of root number and density) promotes increases in EPV components, confirming the assumption of Gao et al. (2016) [48]. It is presumed that there is an association between EPV and the “deep” root component, a trait not evaluated in this study, due to the difficulty of field measurement.
As contributions to popcorn breeding, with a view to increase EPV under DS, it is recommended to use the trait SPAD, via NGR, to facilitate selection, since both are decisive for the expression of EPV. Based on simple correlations, SPAD was considered a reliable trait for selection for DS tolerance in maize [9]. In addition, NGR is a trait frequently associated with higher yields [24]. The use of easily measurable and highly responsive traits can increase the efficiency of selection under stress conditions [35]. According to Edmeades et al. (1999) [31], under DS, a suitable trait is one with high heritability, measured easily and at low cost, and genetically associated with the main variable. Brestic et al. (2018) [49], evaluating wheat genotypes, maintain that selection for higher grain yield, including tolerance to non-ideal conditions, must be accompanied by increased photosynthetic productivity, which is accompanied by higher CO2 assimilation rate, electron transport rate and maximum carboxylation rate, as well as more tolerant photosynthetic thermostability.
Thus, the traits NGR and SPAD were identified as a feasible option to obtain correlated responses by indirect selection. Considering that these traits are also free of GE interaction, it is believed that under FI and DS, indirect selection via SPAD and via NGR will lead to increments in EPV. Therefore, it is proposed to measure these traits in popcorn breeding programs systematically, with the goal of breeding higher-yielding and more DS-tolerant genotypes. To ensure adequate food supply for the global population, three challenges should be considered, namely increasing production potential, protecting production potential and increasing resource efficiency to ensure sustainability [44]. This research provides results in order to increase water-use efficiency.
5. Conclusions
The agricultural, morphological, physiological and root architecture traits used in path analysis explained sufficiently the variation observed in the trait expanded popcorn volume per hectare, in water conditions with and without water stress. Indirect selection using SPAD and number of grais per row will promote increases in expanded popcorn volume per hectare under both water regimes, and is therefore indicated as an option to be exploited in popcorn breeding programs.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/9/12/792/s1: Figure S1: Mean weekly values, in days after sowing, of the temperature (°C), relative humidity (%) and solar radiation (MJ m−2 day−1) throughout the growth period of the popcorn lines (April to August 2016). Male flowering began 67 days after sowing (June 24). Table S1: Precipitation and irrigation (mm) applied, in days after sowing (DAS), to early and late popcorn genotypes under full irrigation (FI) and drought stress (DS). Table S2: Path analysis of the partitioning of genotypic correlations in components of direct and indirect effects obtained from first-order agricultural traits and morphoagricultural, physiological, and second-order root traits evaluated in popcorn lines under FI and DS. Table S3: Path analysis of the partitioning of genotypic correlations in components of direct and indirect effects obtained between the main trait and first-order agricultural traits and morphoagricultural, physiological, and second-order root traits evaluated in popcorn lines under FI and DS.
Author Contributions
Conceptualization, S.H.K., A.T.d.A.J., L.J.M.G. and C.D.C.; methodology, S.H.K., V.J.d.L., L.J.M.G., K.F.M.S., J.T.L., C.D.C. and E.C.; software, V.J.d.L.; validation, S.H.K. and V.J.d.L.; formal analysis, V.J.d.L.; investigation, S.H.K., A.T.d.A.J., V.J.d.L., L.J.M.G., K.F.M.S., J.T.L., P.H.A.D.S., M.M.C., G.S.M., D.R.d.S.J., C.D.C. and E.C.; resources, S.H.K.; data curation, S.H.K. and A.T.d.A.J.; writing - original draft preparation, S.H.K., A.T.d.A.J., V.J.d.L., L.J.M.G. and E.C.; writing: review and editing, S.H.K., V.J.d.L., L.J.M.G. and E.C.; supervision, S.H.K. and A.T.d.A.J.; project administration, S.H.K. and A.T.d.A.J.; funding acquisition, A.T.d.A.J.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Soares, M.O.; Miranda, G.V.; Guimarães, L.J.M.; Marriel, I.E.; Guimarães, C.T. Parâmetros genéticos de uma população de milho em níveis contrastantes de nitrogênio. Rev. Cienc. Agron. 2011, 42, 168–174. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Menkir, A.; Blay, E.; Gracen, V.; Danquah, E.; Hearne, S. Genetic analysis of drought tolerance in adapted × exotic crosses of maize inbred lines under managed stress conditions. Euphytica 2014, 196, 261–270. [Google Scholar] [CrossRef]
- Mendes, F.F.; Guimarães, L.J.M.; Souza, J.C.; Guimarães, P.E.O.; Magalhaes, J.V.; Garcia, A.A.F.; Parentoni, S.N.; Guimarães, C.T. Genetic Architecture of Phosphorus Use Efficiency in Tropical Maize Cultivated in a Low-P Soil. Crop Sci. 2014, 54, 1530–1538. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Durães, F.O.M.; dos Santos, M.X.; Gama, E.; Magalhães, P.C.; Albuquerque, P.E.P.; Guimarães, C.T. Fenotipagem Associada a Tolerância a Seca em Milho para Uso em Melhoramento, Estudos Genômicos e Seleção Assistida por Marcadores. Embrapa Milho e Sorgo-Circular Técnica (INFOTECA-E). 2004, Volume 39, p. 18. Available online: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/487708/1/Circ39.pdf (accessed on 22 November 2019).
- Ghannoum, O. C4 photosynthesis and water stress. Ann. Bot. 2008, 103, 635–644. [Google Scholar] [CrossRef]
- Romano, G.; Zia, S.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Use of thermography for high throughput phenotyping of tropical maize adaptation in water stress. Comput. Electron. Agric. 2011, 79, 67–74. [Google Scholar] [CrossRef]
- Sanchez, C.; Vargas, M.; Ordóñez, R.A.; Cairns, J.E.; Araus, J.L. Dissecting Maize Productivity: Ideotypes Associated with Grain Yield under Drought Stress and Well-watered Conditions. J. Integr. Plant Biol. 2012, 54, 1007–1020. [Google Scholar]
- Zia, S.; Romano, G.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Infrared Thermal Imaging as a Rapid Tool for Identifying Water-Stress Tolerant Maize Genotypes of Different Phenology. J. Agron. Crop Sci. 2013, 199, 75–84. [Google Scholar] [CrossRef]
- Das Gracas Dias, K.O.; Gezan, S.A.; Guimaraes, C.T.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Carneiro, N.P.; Portugal, A.F.; Bastos, E.A.; Cardoso, M.J.; de Oliveira Anoni, C.; et al. Estimating Genotype × Environment Interaction for and Genetic Correlations among Drought Tolerance Traits in Maize via Factor Analytic Multiplicative Mixed Models. Crop Sci. 2018, 58, 72–83. [Google Scholar] [CrossRef]
- Mendes, W.D.S.; Drews, T.A.; Medeiros, J.C.; Rosa, J.D.; Gualberto, A.V.S.; Mielezrski, F. Development and productivity of maize in response to spatial arrangement under semiarid condition of Northeastern Brazil. Aust. J. Crop Sci. 2017, 11, 313–321. [Google Scholar] [CrossRef]
- Vittorazzi, C.; Amaral Júnior, A.T.; Guimarães, A.G.; Silva, F.H.L.; Pena, G.F.; Daher, R.F.; Gerhardt, I.F.S.; Oliveira, G.H.F.; Santos, P.H.A.D.; Souza, Y.P.; et al. Evaluation of genetic variability to form heterotic groups in popcorn. Genet. Mol. Res. 2018, 17. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ren, G.; Yue, G.; Li, Z.; Qu, X.; Hou, G.; Zhu, Y.; Zhang, J. Effects of water-deficit stress on the transcriptomes of developing immature ear and tassel in maize. Plant Cell Rep. 2007, 26, 2137–2147. [Google Scholar] [CrossRef] [PubMed]
- Câmara, T.M.M.; Bento, D.A.V.; Alves, G.F.; Santos, M.F.; Moreira, J.U.V.; de Souza, C.L. Parâmetros genéticos de caracteres relacionados à tolerância à deficiência hídrica em milho tropical. Bragantia 2007, 66, 595–603. [Google Scholar] [CrossRef]
- Teixeira, F.F.; Gomide, R.L.; De Albuquerque, P.E.P.; De Andrade, C.L.T.; Leite, C.E.P.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Silva, A.R.; Bastos, E.A.; Cardoso, M.J. Evaluation of maize core collection for drought tolerance. Crop Breed. Appl. Biotechnol. 2010, 10, 312–320. [Google Scholar] [CrossRef]
- Costa, E.F.N.; Santos, M.F.; Moro, G.V.; Alves, G.F.; de Souza Júnior, C.L. Herança da senescência retardada em milho. Pesqui. Agropecu. Bras. 2008, 43, 207–213. [Google Scholar] [CrossRef]
- Li, X.H.; Liu, X.D.; Li, M.S.; Zhang, S.H. Identification of quantitative trait loci for anthesis-silking interval and yield components under drought stress in maize. Acta Bot. Sin. 2003, 45, 852–857. [Google Scholar]
- Vaughan, M.M.; Christensen, S.; Schmelz, E.A.; Huffaker, A.; Mcauslane, H.J.; Alborn, H.T.; Romero, M.; Allen, L.H.; Teal, P.E.A. Accumulation of terpenoid phytoalexins in maize roots is associated with drought tolerance. Plant Cell Environ. 2015, 38, 2195–2207. [Google Scholar] [CrossRef]
- Zinselmeier, S.A.; Lauer, M.J.; Boyer, J.S. Reversing Drought-Induced Losses in Grain Yield: Sucrose Maintains Embryo Growth in Maize. Crop Sci. 1995, 35, 1390–1400. [Google Scholar] [CrossRef]
- Cruz, C.D.; Carneiro, P.C.S.; Regazzi, A.J. Modelos Biométricos Aplicados ao Melhoramento Genético—Vol. II; UFV: Vicosa, Brazil, 2014; ISBN 8572691510. [Google Scholar]
- De Carvalho, C.G.P.; Oliveira, V.R.; Cruz, C.D.; Casali, V.W.D. Análise de trilha sob multicolinearidade em pimentão. Pesqui. Agropecu. Bras. 1999, 34, 603–613. [Google Scholar] [CrossRef]
- Wright, S. Correlation and causation. J. Agric. Res. 1921, 20, 557–580. [Google Scholar]
- Do Amaral Júnior, A.T.; Dos Santos, A.; Gerhardt, I.F.S.; Kurosawa, R.N.F.; Moreira, N.F.; Pereira, M.G.; Gravina, G.; Silva, F. Proposal of a super trait for the optimum selection of popcorn progenies based on path analysis. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Kamphorst, S.H.; de Lima, V.J.; do Amaral Júnior, A.T.; Schmitt, K.F.M.; Leite, J.T.; Carvalho, C.M.; Silva, R.M.R.; Xavier, K.B.; Fereira, F.R.A.; Santos, P.H.A.D.; et al. Popcorn breeding for water-stress tolerance or for agronomic water-use efficiency? Genet. Mol. Res. 2018, 17. [Google Scholar] [CrossRef]
- Jákli, B.; Hauer-Jákli, M.; Böttcher, F.; Meyer zur Müdehorst, J.; Senbayram, M.; Dittert, K. Leaf, canopy and agronomic water-use efficiency of field-grown sugar beet in response to potassium fertilization. J. Agron. Crop Sci. 2018, 204, 99–110. [Google Scholar] [CrossRef]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Shovelomics: High throughput phenotyping of maize (Zea mays L.) root architecture in the field. Plant Soil 2011, 341, 75–87. [Google Scholar] [CrossRef]
- Christensen, H.K.; Montgomery, C.A. Corporate economic performance: Diversification strategy versus market structure. Strat. Manag. J. 1981, 2, 327–343. [Google Scholar] [CrossRef]
- Cruz, C.D. Genes Software—Extended and integrated with the R, Matlab and Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Da Silva, P.P.; Soares, L.; Da Costa, J.G.; Viana, L.D.S.; De Andrade, J.C.F.; Gonçalves, E.R.; Dos Santos, J.M.; de Souza Barbosa, G.V.; Nascimento, V.X.; Todaro, A.R.; et al. Path analysis for selection of drought tolerant sugarcane genotypes through physiological components. Ind. Crop Prod. 2012, 37, 11–19. [Google Scholar] [CrossRef]
- Edmeades, G.O.; Bolanos, J.; Chapman, S.C.; Lafitte, H.R.; Banziger, M. Selection Improves Drought Tolerance in Tropical Maize Populations. Crop Sci. 1999, 39, 1306–1315. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- Maazou, A.R.S.; Tu, J.; Qiu, J.; Liu, Z. Breeding for Drought Tolerance in Maize (Zea mays L.). Am. J. Plant Sci. 2016, 7, 1858–1870. [Google Scholar] [CrossRef]
- Lu, Y.; Hao, Z.; Xie, C.; Crossa, J.; Araus, J.; Gao, S.; Vivek, B.S.; Magorokosho, C.; Mugo, S.; Makumbi, D.; et al. Field Crops Research Large-scale screening for maize drought resistance using multiple selection criteria evaluated under water-stressed and well-watered environments. Field Crop Res. 2011, 124, 37–45. [Google Scholar] [CrossRef]
- Carena, M.J.; Hallauer, A.R.; Filho, J.M. Quantitative Genetics in Maize Breeding; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- De Castro, F.A.; Campostrini, E.; Netto, A.T.; De Menezes De Assis Gomes, M.; Ferraz, T.M.; Glenn, D.M. Portable chlorophyll meter (PCM-502) values are related to total chlorophyll concentration and photosynthetic capacity in papaya (Carica papaya L.). Theor. Exp. Plant Physiol. 2014, 26, 201–210. [Google Scholar] [CrossRef]
- Liu, Y.; Subhash, C.; Yan, J.; Song, C.; Zhao, J.; Li, J. Maize leaf temperature responses to drought: Thermal imaging and quantitative trait loci (QTL) mapping. Environ. Exp. Bot. 2011, 71, 158–165. [Google Scholar] [CrossRef]
- Kaman, H.; Kirda, C.; Sesveren, S. Genotypic differences of maize in grain yield response to deficit irrigation. Agric. Water Manag. 2011, 98, 801–807. [Google Scholar] [CrossRef]
- Li, M.; Liu, S.; Zhang, S.; Hao, Z.; Li, X.; Xie, C.; Weng, J.; Zhang, D.; Liang, X.; Liu, L. Identification of Functional Genetic Variations Underlying Drought Tolerance in Maize Using SNP Markers. J. Integr. Plant Biol. 2011, 53, 641–652. [Google Scholar]
- Chairi, F.; ElAzab, A.; Sanchez-Bragado, R.; Araus, J.L.; Serret, M.D. Heterosis for water status in maize seedlings. Agric. Water Manag. 2016, 164, 100–109. [Google Scholar] [CrossRef]
- Cabral, P.D.S.; do Amaral Júnior, A.T.; de Freitas, I.L.; Ribeiro, R.M.; da Silva, T.R. Cause and effect of quantitative characteristics on grain expansion capacity in popcorn. Rev. Cienc. Agron. 2016, 47, 108–117. [Google Scholar] [CrossRef]
- Pacheco, C.A.P.; dos Santos, M.X.; Cruz, C.D.; Parentoni, S.N.; de Oliveira Guimarães, P.E.; Gama, E.E.G.E.; de Carvalho, H.W.L.; Vieira Júnior, P.A. Inbreeding depression of 28 maize elite open pollinated varieties. Genet. Mol. Biol. 2002, 25, 441–448. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A.J. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- Chen, D.; Neumann, K.; Friedel, S.; Kilian, B.; Chen, M.; Altmann, T.; Klukas, C. Dissecting the phenotypic components of crop plant growth and drought responses based on high-throughput image analysis. Plant Cell 2014, 26, 4636–4655. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, S.; Elhoweity, M.; Ibrahim, H.H.; Dewir, Y.H.; Migdadi, H.M.; Schmidhalter, U. Thermal imaging and passive reflectance sensing to estimate the water status and grain yield of wheat under different irrigation regimes. Agric. Water Manag. 2017, 189, 98–110. [Google Scholar] [CrossRef]
- Petrov, P.; Petrova, A.; Dimitrov, I.; Tashev, T.; Olsovska, K.; Brestic, M.; Misheva, S. Relationships between leaf morpho-anatomy, water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought. J. Agron. Crop Sci. 2018, 204, 219–227. [Google Scholar] [CrossRef]
- Gao, Y.; Lynch, J.P. Reduced crown root number improves water acquisition under water deficit stress in maize (Zea mays L.). J. Exp. Bot. 2016, 67, 4545–4557. [Google Scholar] [CrossRef]
- Brestic, M.; Živčák, M.; Hauptvogel, P.; Misheva, S.; Kocheva, K.; Yang, X.; Li, X.; Allakhverdiev, S.I. Wheat plant selection for high yields entailed improvement of leaf anatomical and biochemical traits including tolerance to non-optimal temperature conditions. Photosynth. Res. 2018, 136, 245–255. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).