Functional Analysis of Three miRNAs in Agropyron mongolicum Keng under Drought Stress

Abstract
1. Introduction
2. Methods
2.1. Ethics Statement
2.2. Plant Materials and Drought Treatment
2.3. Total RNA Extraction, Small RNA Library Construction, and Sequencing
2.4. Identification of miRNAs and Prediction of Target Genes
2.5. Differential Expression Analysis of miRNAs
2.6. qRT-PCR Analysis
2.7. Construction of Expression Vector of Drought-Responsive miRNAs
2.8. Agrobacterium-Mediated Genetic Transformation of Arabidopsis thaliana
2.9. Drought Treatment of Transgenic Arabidopsis Plants
3. Results
3.1. Sequencing and Identification of A. mongolicum Small RNAs
3.2. Identification of miRNAs in A. mongolicum
3.3. Differential Expression of miRNAs under Drought Stress
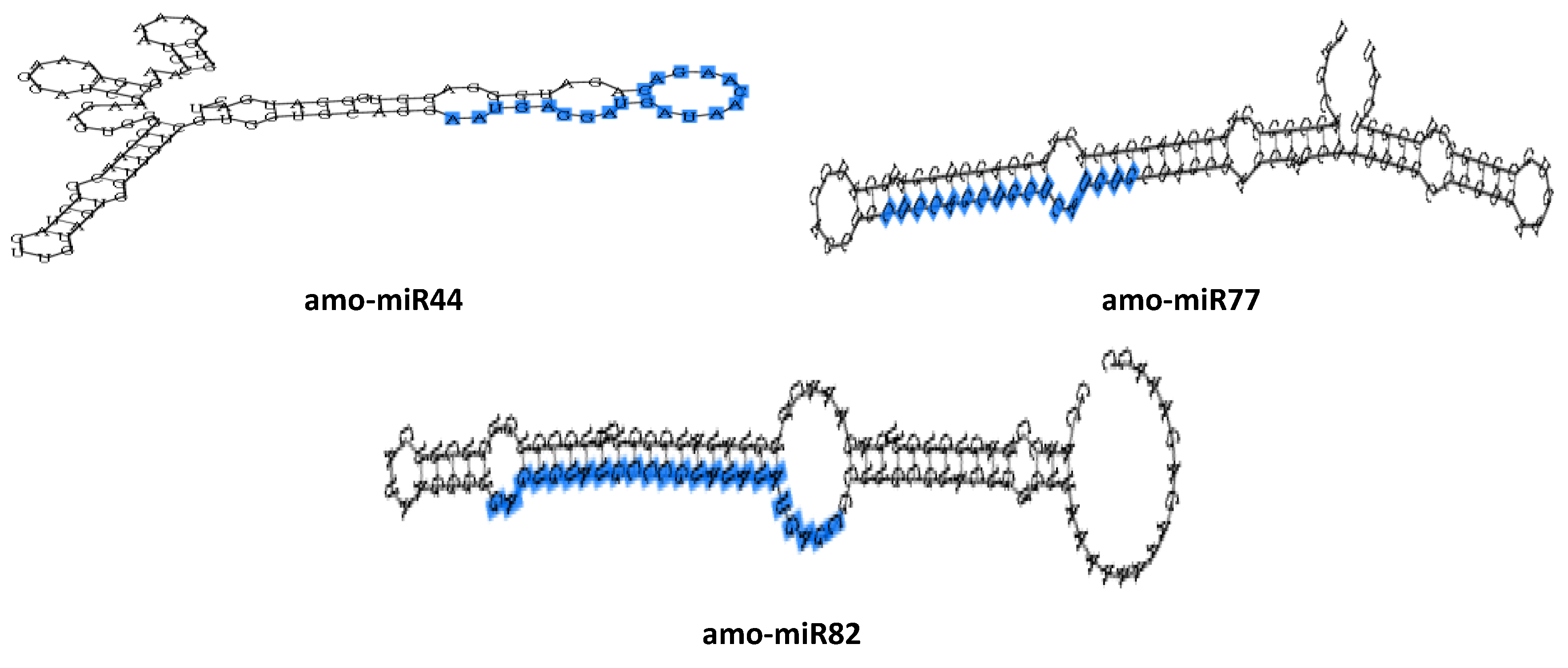
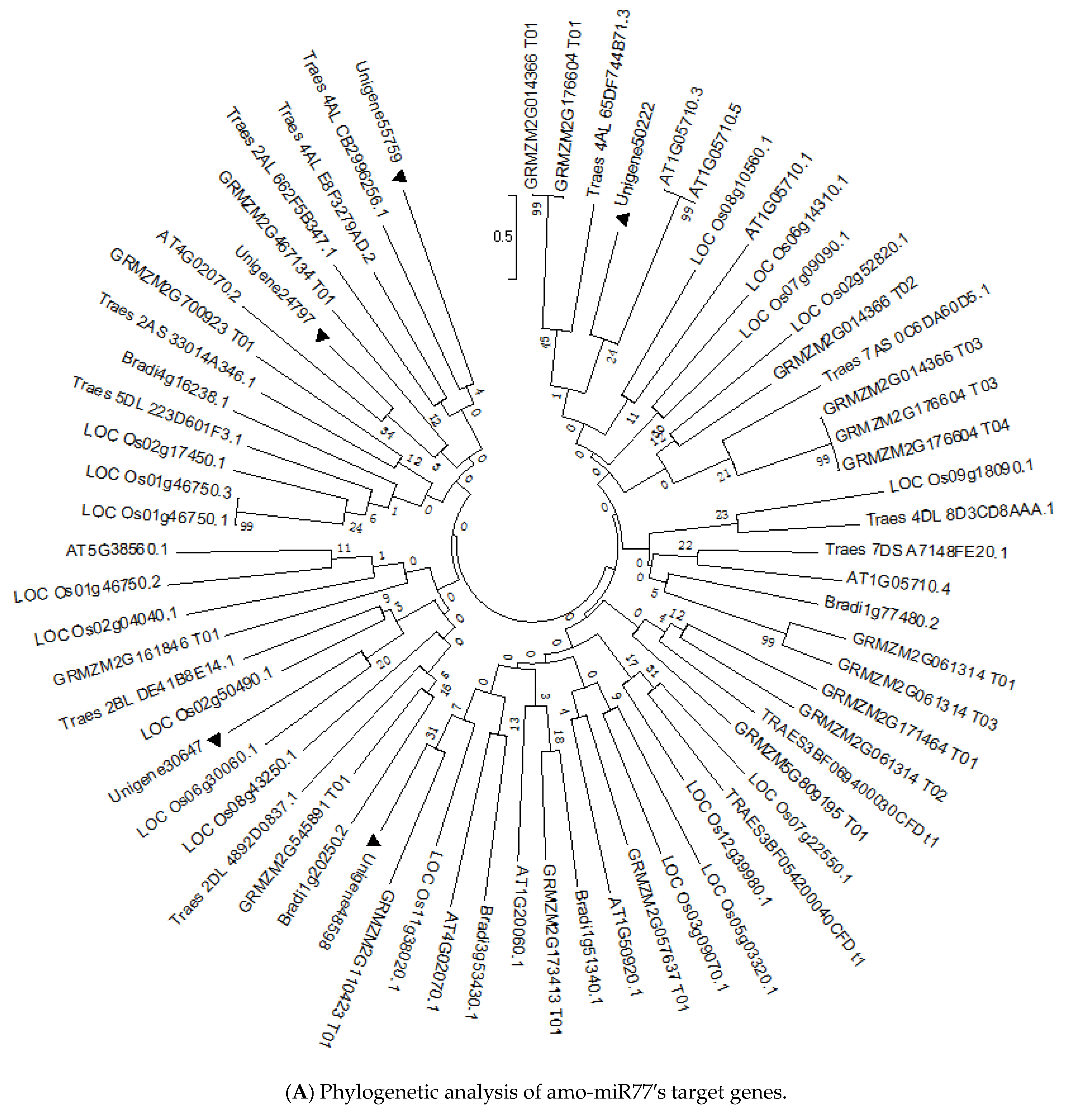
3.4. Target Gene Prediction and Functional Classification of Drought-Responsive miRNAs
3.5. Expression Profiles of Seven Drought-Responsive miRNAs in A. mongolicum
3.6. Amo-miR21, Amo-miR5, Amo-miR62 Transgenic Arabidopsis Plants Were Tolerant to Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dewey, D.R. Forage resources and research in northern China. In Agronomy Abstracts; ASA: Madison, WI, USA, 1981; p. 60. [Google Scholar]
- Che, Y.H.; Li, L.H. Genetic diversity of prolamines in Agropyron mongolicum Keng indigenous to northern China. Genet. Resour. Crop Evol. 2007, 54, 1145–1151. [Google Scholar] [CrossRef]
- Han, H.; Liu, W.; Lu, Y.; Zhang, J.; Yang, X.; Li, X.; Hu, Z.; Li, L. Isolation and application of P genome-specific DNA sequences of Agropyron Gaertn. in Triticeae. Planta 2017, 245, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.S.; Zhou, R.H.; Xu, S.J.; Li, L.H.; Cauderon, Y.; Wang, R.R.-C. Desirable characteristics in perennial Triticeae collected in China for wheat improvement. Hereditas 1992, 116, 175–178. [Google Scholar] [CrossRef]
- Zhao, Y.; Gao, C.P.; Shi, F.L.; Yun, L.; Jia, Y.S.; Wen, J.Q. Transcriptomic and proteomic analyses of drought responsive genes and proteins in Agropyron mongolicum Keng. Curr. Plant Biol. 2018, 14, 19–29. [Google Scholar] [CrossRef]
- Du, J.C.; Li, X.Q.; Li, T.T.; Yu, D.Y.; Han, B. Genome-wide transcriptome profiling provides overwintering mechanism of Agropyron mongolicum. BMC Plant Biol. 2017, 17, 38–151. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yun, J.F.; Shi, F.M.; Wang, J.J.; Yang, Q.C.; Chao, Y.H. Molecular cloning and characterization of a group 3 LEA gene from Agropyron mongolicum Keng. Afr. J. Biotechnol. 2010, 9, 6040–6048. [Google Scholar] [CrossRef]
- Ao, T.G.B.Y.; Gao, L.J.; Wang, L.C.; Li, Y.Q.; Zhao, Y.; Lang, M.L.; Yang, X.J. Cloning and Expression Analysis of AP2/EREBP Transcription Factor Gene (MwAP2/EREBP) in Agropyron mongolicum Keng. Nanosci. Nanotechnol. Lett. 2017, 9, 2031–2038. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, X.; Yu, Z.; Sun, F.; Li, X.; Li, X. RNA-Seq of Agropyron mongolicum Keng in response to drought stress. Grassl. Sci. 2018, 64, 3–15. [Google Scholar] [CrossRef]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef]
- Pashkovskiy, P.P.; Ryazansky, S.S. Biogenesis, evolution, and functions of plant microRNAs. Biochemistry (Mosc.) 2013, 78, 627–637. [Google Scholar] [CrossRef]
- Shalgi, R.; Lieber, D.; Oren, M.; Pilpel, Y. Global and local architecture of the mammalian microRNA transcription factor regulatory network. PLoS Comput. Biol. 2017, 3, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Su, N.F.; Qian, M.P.; Wang, Y.F.; Deng, M.H. Combinatorial regulation of transcription factors and microRNAs. BMC Syst. J. Biol. 2010, 4, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Tian, X.; Li, Y.J.; Wu, C.A.; Zheng, C.C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. Rna-a Publ. Rna Soc. 2008, 14, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cui, X.; Meng, Z.L.; Huang, X.H.; Xie, Q.; Wu, H.; Jin, H.L.; Zhang, D.B.; Liang, W.Q. Transcriptional regulation of Arabidopsis MIR168a and argonaute1 homeostasis in abscisic acid and abiotic stress responses. Plant Physiol. 2012, 158, 1279–1292. [Google Scholar] [CrossRef]
- Li, W.X.; Oono, Y.; Zhu, J.H.; He, X.J.; Wu, J.M.; Lida, K.; Lu, X.Y.; Cui, X.P.; Jin, H.L.; Zhu, J.K. The Arabidopsis NFYA5 transcription factor is regulated transcriptionally and posttranscriptionally to promote drought resistance. Plant Cell 2008, 20, 2238–2251. [Google Scholar] [CrossRef]
- Song, J.B.; Gao, S.; Sun, D.; Li, H.; Shu, X.X.; Yang, Z.M. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef]
- Reyes, J.L.; Chua, N.H. ABA induction of miR159 controls transcript levels of two MYB factors during Arabidopsis seed germination. Plant J. 2007, 49, 592–606. [Google Scholar] [CrossRef]
- Candar-cakir, B.; Arican, E.; Zhang, B. Small RNA and degradome deep sequencing reveals drought and tissue-specific microRNAs and their important roles in drought-sensitive and drought tolerant tomato genotypes. Plant Biotechnol. J. 2016, 14, 1727–1746. [Google Scholar] [CrossRef]
- Gao, F.; Wang, N.; Li, H.; Liu, J.; Fu, C.; Xiao, Z.; Wei, C.; Lu, X.; Feng, J.; Zhou, Y. Identification of drought-responsive microRNAs and their targets in Ammopiptanthus mongolicus by using high-throughput sequencing. Sci. Rep. 2016, 6, 34601. [Google Scholar] [CrossRef]
- Trindade, I.; Capitao, C.; Dalmay, T.; Fevereiro, M.P.; Santos, D.M.D. miR398 and miR408 are up-regulated in response to water deficit in Medicago truncatula. Planta 2010, 231, 705–716. [Google Scholar] [CrossRef]
- Wei, L.Y.; Zhang, D.F.; Xiang, F.; Zhang, Z.X. Differentially expressed miRNAs potentially involved in the regulation of defense mechanism to drought stress in maize seedlings. Int. J. Plant Sci. 2009, 170, 979–989. [Google Scholar] [CrossRef]
- Chen, Q.S.; Li, M.; Zhang, Z.C.; Tie, W.W.; Chen, X.; Jin, L.F.; Zhai, N.; Zheng, Q.X.; Zhang, J.F.; Wang, R.; et al. Integrated mRNA and microRNA analysis identifies genes and small miRNA molecules associated with transcriptional and post-transcriptional-level responses to both drought stress and re-watering treatment in tobacco. BMC Genom. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Jung, H.; Jeong, D.H.; Ha, S.H.; Choi, Y.D.; Kim, J.K. Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genom. 2016, 17, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Kantar, M.; Lucas, S.J.; Budak, H. miRNA expression patterns of Triticum dicoccoides in response to shock drought stress. Planta 2011, 233, 471–484. [Google Scholar] [CrossRef]
- Yao, Y.; Guo, G.; Ni, Z.; Sunkar, R.; Du, J.; Zhu, J.K.; Sun, Q. Cloning and characterization of microRNAs from wheat (Triticum aestivum L.). Genome Biol. 2007, 8, 1–17. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Guo, C.J.; Li, X.J.; Duan, W.W.; Ma, C.Y.; Chan, H.M.; Wen, Y.L.; Lu, W.J.; Xiao, K. Characterization and expression pattern analysis of microRNAs in wheat under drought stress. Biol. Plant. 2015, 59, 37–46. [Google Scholar] [CrossRef]
- Zhang, B.H.; Pan, X.P.; Wang, Q.L.; George, P.C.; Anderson, T.A. Identification and characterization of new plant microRNAs using EST analysis. Cell Res. 2005, 15, 336–360. [Google Scholar] [CrossRef]
- Zhang, B.H.; Pan, X.P.; Cox, S.B.; Cobb, G.P.; Anderson, T.A. Evidence that miRNAs are different from other RNAs. Cell. Mol. Life Sci. 2006, 63, 246–254. [Google Scholar] [CrossRef]
- Fahlgren, N.; Howell, M.D.; Kasschau, K.D.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Law, T.F.; Grant, S.R.; Dangl, J.L.; et al. High-throughput sequencing of Arabidopsis microRNAs: Evidence for frequent birth and death of MIRNA genes. PLoS ONE 2007, 2, 219. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, 199–205. [Google Scholar] [CrossRef]
- Dai, X.B.; Zhuang, Z.H.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ridzon, D.A.; Broomer, A.J.; Zhou, Z.; Lee, D.H.; Nguyen, J.T.; Barbisin, M.; Xu, N.L.; Mahuvakar, V.R.; Andersen, M.R.; et al. Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res. 2005, 33, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Ghedira, R.; Buck, S.D.; Nolf, J.; Depicker, A. The efficiency of Arabidopsis Thaliana floral dip transformation is determined not only by the Agrobacterium strain used but also by the physiology and the ecotype of the dipped plant. Mol. Plant-Microbe Interact. MPMI 2013, 26, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Sunkar, R.; Zhou, C.; Shen, H.; Zhang, J.Y.; Matts, J.; Wolf, J.; Mann, D.G.; Stewart, C.N., Jr.; Tang, Y.; et al. Overexpression of miR156 in switchgrass (Panicum virgatum L.) results in various morphological alterations and leads to improved biomass production. Plant Biotechnol. J. 2012, 10, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Bent, A. Arabidopsis thaliana Floral Dip Transformation Method. Methods Mol. Biol. (MIMB) 2006, 343, 87–104. [Google Scholar] [CrossRef]
- Noman, A.; Fahad, S.; Aqeel, M.; Ali, U. miRNAs: Major modulators for crop growth and development under abiotic stresses. Biotechnol. Lett. 2017, 39, 685–700. [Google Scholar] [CrossRef]
- Dong, Z.; Zhang, J.; Zhu, Q.; Zhao, L.; Sui, S.; Li, Z.; Zhang, Y.; Wang, H.; Tian, D.; Zhao, Y. Identification of microRNAs involved in drought stress responses in early-maturing cotton by high-throughput sequencing. Genes Genom. 2018, 40, 305–314. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, S.; Zhu, C.; Chang, X.; Yue, C.; Wang, Z.; Lin, Y.; Lai, Z. Identification of drought-responsive miRNAs and physiological characterization of tea plant (Camellia sinensis L.) under drought stress. BMC Plant Biol. 2017, 17, 211–231. [Google Scholar] [CrossRef]
- Ao, T.G.B.Y.; Lang, M.L.; Li, Y.Q.; Zhao, Y.; Wang, L.C.; Yang, X.J. Cloning and expression analysis of cysteine protease gene (MwCP) in Agropyron mongolicum Keng. Genet. Mol. Res. 2016, 15, 1–18. [Google Scholar] [CrossRef]
- Zhou, L.G.; Liu, Y.H.; Liu, Z.C.; Kong, D.Y.; Duan, M.; Luo, L.J. Genome-wide identification and analysis of drought-responsive microRNAs in Oryza sativa. J. Exp. Bot. 2010, 61, 4157–4168. [Google Scholar] [CrossRef]
- Guo, H.S.; Xie, Q.; Fei, J.F.; Chua, N.H. MicroRNA directs mRNA cleavage of the transcription factor NAC1 to downregulate auxin signals for arabidopsis lateral root development. Plant Cell 2005, 17, 1376–1386. [Google Scholar] [CrossRef]
- Fang, Y.J.; Xie, K.B.; Xiong, L.Z. Conserved miR164-targeted NAC genes negatively regulate drought resistance in rice. J. Exp. Bot. 2014, 65, 2119–2135. [Google Scholar] [CrossRef] [PubMed]
- Sobkowiak, A.; Jończyk, M.; Adamczyk, J.; Szczepanik, J.; Solecka, D.; Kuciara, I.; Hetmańczyk, K.; Trzcinska-Danielewicz, J.; Grzybowski, M.; Skoneczny, M.; et al. Molecular foundations of chilling-tolerance of modern maize. BMC Genom. 2016, 17, 125–147. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Li, Q.; Yan, C.; Liu, Y.; Liu, J.; Yu, F.; Wang, Z.; Long, J.; He, J.; Wang, H.W.; et al. Structural basis for the modular recognition of single-stranded RNA by PPR proteins. Nature 2013, 504, 168–183. [Google Scholar] [CrossRef] [PubMed]
- Ling, Q.H.; Jarvis, P. Functions of plastid protein import and the ubiquitin-proteasome system in plastid development. Biochim. Biophys. Acta 2015, 1847, 939–948. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. BBA-Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef]
- Kohli, D.; Joshi, G.; Deokar, A.A. Identification and characterization of wilt and salt stress-responsive microRNAs in chickpea through high-throughput sequencing. PLoS ONE 2014, 9, 108851. [Google Scholar] [CrossRef]
- Soderlund, C.; Descour, A.; Kudrna, D.; Bomhoff, M.; Boyd, L.; Currie, J.; Angelova, A.; Collura, K.; Wissotski, M.; Ashley, E.; et al. Sequencing, mapping, and analysis of 27,455 maize full-length cDNAs. PLoS Genet. 2009, 5, 1000740. [Google Scholar] [CrossRef]
- Drechsel, G.; Raab, S.; Hoth, S. Arabidopsis zinc-finger protein 2 is a negative regulator of ABA signaling during seed germination. Plant Physiol. 2010, 167, 1418–1421. [Google Scholar] [CrossRef]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The Basic Helix-Loop-Helix Transcription Factor Family in Plants: A Genome-Wide Study of Protein Structure and Functional Diversity. Mol. Biol. Evol. 2003, 20, 735–747. [Google Scholar] [CrossRef]
- Ni, W.; Xu, S.L.; González-Grandío, E.; Chalkley, R.J.; Huhmer, A.F.; Burlingame, A.L.; Wang, Z.Y.; Quail, P.H. PPKs mediate direct signal transfer from phytochrome photoreceptors to transcription factor PIF3. Nat. Commun. 2017, 8, 15236. [Google Scholar] [CrossRef] [PubMed]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Primer Sequence (5′—3′) |
|---|---|
| amo-miR21 | Forward: ACACTCCAGCTGGG GAGTGTATGCCCGTATATAT |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG ATAGTTT | |
| amo-miR82 | Forward: ACACTCCAGCTGGG ATGCTCGCTCCTCTTTCTGT |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG GCTCATA | |
| amo-miR62 | Forward: ACACTCCAGCTGGG CACGTGCTCGATGAAAT |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG AGTCATT | |
| amo-miR44 | Forward: ACACTCCAGCTGGG AATGAGGATGATAACA |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG GTCTTGT | |
| amo-miR5 | Forward: ACACTCCAGCTGGG GGCGGATGTAGC |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAG TCCACTT | |
| amo-miR17 | Forward: ACACTCCAGCTGGG CCCAACGGGCGGTG |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGCCCCAC | |
| amo-miR77 | Forward: ACACTCCAGCTGGG CTGCTCCAGCTGCTCA |
| stem-loop RT primer: CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACATGA |
| Primer | Primer Sequences (5′—3′) |
|---|---|
| amo-miR21-F | CGggatccACACTCCAGCTGGG GAGTGTATGCCCGTATATAT |
| amo-miR21-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATAGTTT |
| amo-miR82-F | CGggatccACACTCCAGCTGGG ATGCTCGCTCCTCTTTCTGT |
| amo-miR82-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGCTCATA |
| amo-miR17-F | CGggatccACACTCCAGCTGGG CCCAACGGGCGGTG |
| amo-miR17-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGCCCCAC |
| amo-miR5-F | CGggatccACACTCCAGCTGGG GGCGGATGTAGC |
| amo-miR5-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCCACTT |
| amo-miR44-F | CGggatccACACTCCAGCTGGG AATGAGGATGATAACA |
| amo-miR44-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGTCTTGT |
| amo-miR77-F | CGggatccACACTCCAGCTGGG CTGCTCCAGCTGCTCA |
| amo-miR77-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACATGA |
| amo-miR62-F | CGggatccACACTCCAGCTGGG CACGTGCTCGATGAAAT |
| amo-miR62-RE | CgagctcCTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGTCATT |
| Type | Control Plants | Drought-Stressed Plants | ||
|---|---|---|---|---|
| Reads Number | Percentage | Reads Number | Percentage | |
| Total reads number | 16,887,273 | 100% | 30,258,338 | 100% |
| Filter low quality reads | 0 | 0% | 0 | 0% |
| Filter having ‘N’ reads | 1138 | 0.01% | 3408 | 0.01% |
| Length <18 | 6,681,521 | 39.57% | 10,989,611 | 36.32% |
| Length >30 | 2,159,377 | 12.79% | 2,858,115 | 9.45% |
| Clean Reads | 8,045,237 | 47.64% | 16,407,204 | 54.22% |
| miRNA | Sequence | Length (nt) | Regulation Mode | Precursor Length (nt) | MFE (kcal/mol) | MFEI | Conservatism | Homologous miRNA | Expression |
|---|---|---|---|---|---|---|---|---|---|
| amo-miR5 | ggcggauguagccaagugga | 20 | up | 109 | −48.84 | 0.86 | unconservative | qRT-PCR | |
| amo-miR14 | cgcggcggcgggggcggug | 19 | down | 108 | −63.70 | 0.76 | unconservative | ||
| amo-miR17 | cccaacgggcgguggggc | 18 | down | 111 | −66.15 | 0.81 | unconservative | qRT-PCR | |
| amo-miR18 | gauggcgcgcaggcggacg | 19 | down | 108 | −62.43 | 0.72 | unconservative | ||
| amo-miR21 | aaagugucguagaaaaaacuau | 22 | up | 111 | −32.73 | 1.09 | conservative | bdi-miR5180b | qRT-PCR |
| amo-miR29 | gacugaugucgguauggaaccagu | 24 | up | 110 | −41.70 | 0.76 | unconservative | ||
| amo-miR32 | cuacgcgucggaugcacugcgu | 22 | up | 111 | −60.18 | 0.85 | unconservative | ||
| amo-miR35 | aucaaggaauuugugagg | 18 | down | 108 | −59.86 | 1.33 | unconservative | ||
| amo-miR40 | uuugaacuggugguugaaugc | 21 | down | 110 | −42.76 | 0.82 | conservative | bdi-miR7743-3p | |
| amo-miR43 | cgcggcgacgggggcgug | 18 | down | 108 | −61.76 | 0.74 | unconservative | ||
| amo-miR44 | aaugaggaugauaacaagac | 20 | down | 110 | −34.09 | 0.62 | conservative | ath-miR854a | qRT-PCR |
| amo-miR46 | auccgucgugauaugaaaaccagc | 24 | up | 114 | −61.36 | 0.93 | unconservative | ||
| amo-miR49 | cgccggagcugcaaugaagc | 20 | up | 109 | −62.25 | 0.93 | unconservative | ||
| amo-miR62 | cacgugcucgaugaaaugacu | 21 | up | 110 | −46.04 | 0.92 | conservative | zma-miR164g-3p | qRT-PCR |
| amo-miR63 | uugcgucaaagguccuagau | 20 | up | 110 | −32.50 | 0.93 | unconservative | ||
| amo-miR77 | cugcuccagcugcucaugug | 20 | up | 108 | −66.79 | 1.04 | conservative | bdi-miR7725b-5p.2 | qRT-PCR |
| amo-miR82 | gaguguaugcccguauauaugagc | 24 | up | 114 | −33.62 | 0.69 | conservative | bdi-miR5066 | qRT-PCR |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Fan, B.; Yu, Z.; Nie, L.; Zhao, Y.; Yu, X.; Sun, F.; Lei, X.; Ma, Y. Functional Analysis of Three miRNAs in Agropyron mongolicum Keng under Drought Stress. Agronomy 2019, 9, 661. https://doi.org/10.3390/agronomy9100661
Zhang X, Fan B, Yu Z, Nie L, Zhao Y, Yu X, Sun F, Lei X, Ma Y. Functional Analysis of Three miRNAs in Agropyron mongolicum Keng under Drought Stress. Agronomy. 2019; 9(10):661. https://doi.org/10.3390/agronomy9100661
Chicago/Turabian StyleZhang, Xuting, Bobo Fan, Zhuo Yu, Lizhen Nie, Yan Zhao, Xiaoxia Yu, Fengcheng Sun, Xuefeng Lei, and Yanhong Ma. 2019. "Functional Analysis of Three miRNAs in Agropyron mongolicum Keng under Drought Stress" Agronomy 9, no. 10: 661. https://doi.org/10.3390/agronomy9100661
APA StyleZhang, X., Fan, B., Yu, Z., Nie, L., Zhao, Y., Yu, X., Sun, F., Lei, X., & Ma, Y. (2019). Functional Analysis of Three miRNAs in Agropyron mongolicum Keng under Drought Stress. Agronomy, 9(10), 661. https://doi.org/10.3390/agronomy9100661