Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seed Coating Treatment
2.3. Experimental Design for Field Trials
2.4. Effect of Seed Coating with PsJN Strain and Thyme Oil on Plant Physiology, Disease Control and Yield Components
2.5. Effect of Seed Coating with PsJN Strain and Thyme Oil on Total Nitrogen and Carbon Content and Stable Carbon and Nitrogen Isotope Composition
2.6. Statistical Analysis
3. Results
3.1. Climatic Features and Sources of Variances of 3 Years of Study
3.2. Effect of SLB Severity on Wheat Yield Components in Control Plants
3.3. Effect of Seed Coating Treatment on SLB Severity and Yield Components
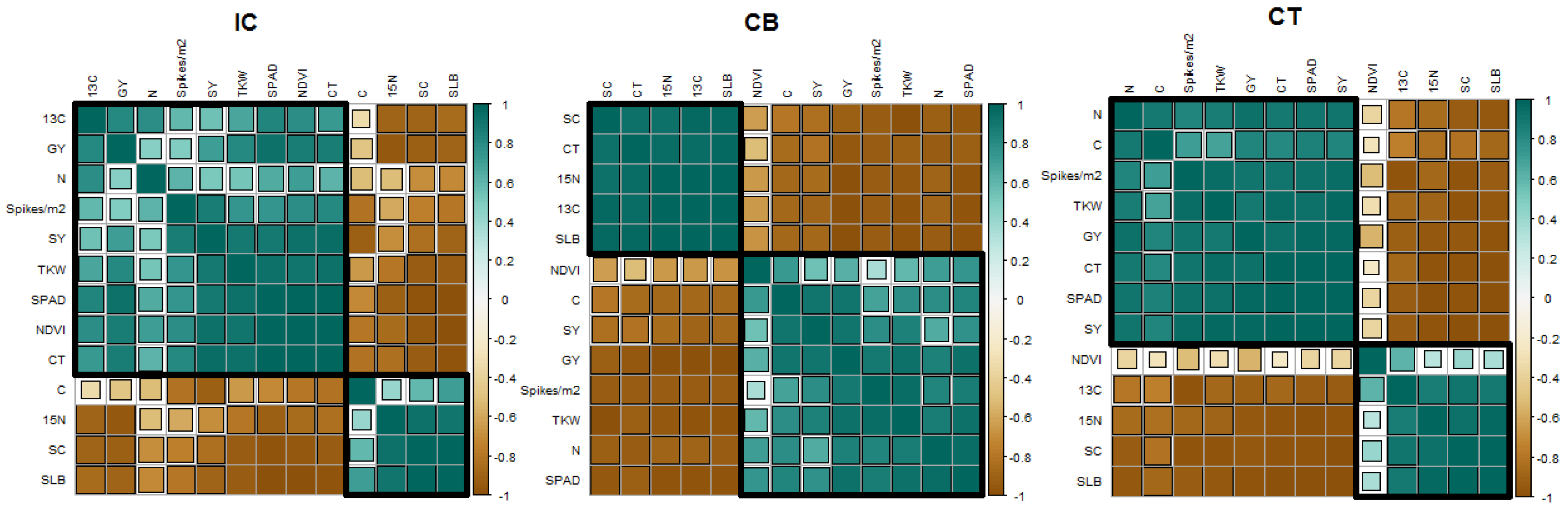
3.4. Effect of Seed CoatingTreatment and SLB Severity on Physiological Traits, Yield Components and Grain Isotopic Composition
3.4.1. Effect of SLB in Control Plants
3.4.2. Effect of Seed Coating with PsJN Strain
3.4.3. Effect of Seed Coating with Thyme Oil
4. Discussion
4.1. Effect of Climate on Variability of SLB Severity, Yield Components among the 3 Years
4.2. Effect of SLB on PhysiologicalTraits, Yield Components, and Stable Isotopic Composition
4.3. Effect of PsJN Strain
4.4. Effect of Thyme Oil
4.5. Comparison between Treatments and Insight to Cost/Gain Balance
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Curtis, B.C. Wheat in the World. In Bread Wheat Improvement and Production; Curtis, B.C., Rajaram, S., Macpherson, H.G., Eds.; Plant Prod Prot Series; FAO: Roma, Italy, 2002; Volume 30, pp. 1–18. [Google Scholar]
- McDonald, B.A.; Mundt, C.C. How knowledge of pathogen population biology informs management of Septoria tritici blotch. Phytopathology 2016, 106, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.N.; Jahn, M.; Clark, B.; Antichi, D.; Góral, T.; Schepers, H.; Lucas, P.; Gouache, D.; Hornok, L. Report on Best Control Practices of Diseases in Winter Wheat in 8 Countries in the EU; ENDURE Project Report DR1.2; Endure: Aarhus, Denmark, 2008. [Google Scholar]
- Matzen, N.; Jørgensen, J.R.; Holst, N.; Jørgensen, L.N. Grain quality in wheat—Impact of disease management. Eur. J. Agron. 2019, 103, 152–164. [Google Scholar] [CrossRef]
- Goodwin, S.B.; Mbarek, S.B.; Dhillon, B.; Wittenberg, A.H.; Crane, C.F.; Hane, J.K.; Foster, A.J.; Van der Lee, T.A.J.; Grimwood, J.; Aerts, A.; et al. Finished genome of the fungal wheat pathogen Mycosphaerella graminicola reveals dispensome structure, chromosome plasticity, and stealth pathogenesis. PLoS Genet. 2011, 7, e1002070. [Google Scholar] [CrossRef] [PubMed]
- Matysiak, K.; Miziniak, W.; Kaczmare, S.; Kierzek, R. Herbicides with natural and synthetic biostimulants in spring wheat. Ciênc. Rural 2018, 48, e20180405. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Djebali, N.; Kthiri, Z.; Ait-Barka, E.; Jacquard, C.; Hamada, W. Seed coating with Paraburkholderia phytofirmans PsJN strain or thyme essential oil enhances durum wheat seedling growth and modulates peroxidase activity and phenolics content. Acta Physiol. Plant. Under review.
- Ben-Jabeur, M.; Kthiri, Z.; Somai-Jammali, L.; Ait-Barka, E.; Jacquard, C.; Hamada, W. Triggering Systemic Resistance by seed coating with thyme oil and Paraburkholderia phytofirmans PsJN strain against Septoria leaf blotch of durum wheat. J. Plant Dis. Prot. Under review.
- Francl, L.J. The Disease Triangle: A plant pathological paradigm revisited. Plant Health Instr. 2001. [Google Scholar] [CrossRef]
- Gouache, D.; Bensdoun, A.; Brun, F.; Oage, C.; Makowski, D.; Wallach, D. Modelling climate change impact on Septoria tritici Blotch (STB) in France: Accounting for climate model and disease model uncertainty. Agric. Meteorol. 2013, 170, 242–252. [Google Scholar] [CrossRef]
- Eastburn, D.M.; McElrone, A.J.; Bilgin, D.D. Influence of atmospheric and climatic change on plant-pathogen interactions. Plant Pathol. 2011, 60, 54–69. [Google Scholar] [CrossRef]
- Ferjaoui, S.; MBarek, S.; Bahri, B.; Slimane, R.; Hamza, S. Identification of resistance sources to septoria tritici blotch in old Tunisian durum wheat germplasm applied for the analysis of the Zymoseptoria tritici-durum wheat interaction. J. Plant Pathol. 2015, 97, 471–481. [Google Scholar]
- Oerke, E.C.; Steiner, U. Potential of Digital Thermography for Disease Control. In Precision Crop Protection—The Challenge and Use of Heterogeneity; Oerke, E.C., Gerhards, R., Menz, G., Sikora, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 5, pp. 167–182. [Google Scholar]
- Ghashghaie, J.; Tcherkez, G. Isotope Ratio Mass Spectrometry Technique to Follow Plant Metabolism: Principles and Applications of 12C/13C Isotopes. In Advances in Botanical Research: Metabolomics Coming of Age with its Technological Diversity; Rolin, D., Ed.; Academic Press Elsevier: London, UK, 2013; Volume 67, pp. 377–405. [Google Scholar]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Heil, M.; Baldwin, I.T. Fitness costs of induced resistance: Emerging experimental support for a slippery concept. Trends Plant Sci. 2002, 7, 61–67. [Google Scholar] [CrossRef]
- Travaglia, C.; Cohen, A.C.; Reinoso, H.; Castillo, C.; Bottini, R. Exogenous abscisic acid increases carbohydrate accumulation and redistribution to the grains in wheat grown under field conditions of soil water restriction. J. Plant Growth Regul. 2007, 26, 285–289. [Google Scholar] [CrossRef]
- Araus, J.L.; Amaro, T.; Zuhair, Y.; Nachit, M.M. Effect of leaf structure and water status on carbon isotope discrimination in field-grown durum wheat. Plant Cell Environ. 1997, 20, 1484–1494. [Google Scholar] [CrossRef]
- Sanchez-Bragado, R.; Serret, M.D.; Araus, J.L. The nitrogen contribution of different plant parts to wheat grains: Exploring genotype, water, and nitrogen effects. Front. Plant Sci. 2017, 7, 19–86. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.D.; Brooks, J.R.; Lajtha, K. Sources of variation in the stable isotopic composition of plants. In Stable Isotopes in Ecology and Environmental Science, 2nd ed.; Michener, R., Lajtha, K., Eds.; Wiley-Blackwell: Malden, MA, USA, 2007; Volume 2, pp. 22–60. [Google Scholar]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef]
- Ben Slimane, R. Effets de la Septoriose Foliaire sur la Sénescence et les Flux D’azote Pendant le Remplissage des Grains Chez le blé Tendre; Agronomie, AgroParisTech: Paris, Françe, 2010. [Google Scholar]
- Garry, G.; Tivoli, B.; Jeuffroy, M.H.; Citharel, J. Effects of Ascochyta blight by Mycosphaerella pinodes on the translocation of carbohydrates and nitrogenous compounds from the leaf and hull to the seed of dried-pea. Plant Pathol. 1996, 45, 769–777. [Google Scholar] [CrossRef]
- McKendry, A.L.; Henke, G.E.; Finney, P.L. Effects of Septoria leaf blotch on soft red winter wheat milling and baking quality. Cereal Chem. 1995, 72, 142–146. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving nitrogen use efficiency in crops for a sustainable agriculture. Sustainability 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Vessey, J.K.; Henry, L.T.; Chaillou, S.; Raper, C.D. Root-zone acidity affects relative uptake of nitrate and ammonium from mixed nitrogen sources. J. Plant Nutr. 1990, 13, 95–116. [Google Scholar] [CrossRef] [PubMed]
- Boots, M.; Best, A. The evolution of constitutive and induced defences to infectious disease. Proc. R. Soc. B 2018, 285, 20180658. [Google Scholar] [CrossRef] [PubMed]
- Ben Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against gray mold and Fusarium wilt. Plant Phys. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Eyal, Z.; Scharen, A.L.; Prescott, J.M.; van Ginkel, M. The Septoria Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: EI Batan, Mexico, 1987. [Google Scholar]
- Stewart, E.L.; McDonald, B.A. Measuring quantitative virulence in the wheat pathogen Zymoseptoria tritici using high-throughput automated image analysis. Phytopathology 2014, 104, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Coplen, T.B. Explanatory Glossary of Terms Used in Expression of Relative Isotope Ratios and Gas Ratios. IUPAC Provisional Recommendations. Inorganic Chemistry Division, Comission on Isotopic Abundances and Atomic Weights. Available online: http://old.iupac.org/reports/provisional/abstract08/coplen_prs.pdf (accessed on 25 November 2017).
- Fiche de Données de Sécurité: Agicote Rouge Cereals; Quick-FDS [18207-41570-12577-010076]; Aegilops Applications: Val de Reuil, France, 5 December 2017; Available online: http://www.utdf.coop/alfresco/service/api/node/content/workspace/SpacesStore/ec008e4e-1c48-4401-9a42-6df5cffcd4cb/101098.pdf?guest=true (accessed on 16 February 2018).



{kind=link}
{kind=link}
| Climatic Factors | Precipitation (mm) | T Min (°C) | T Max (°C) | Humidity (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| year | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 |
| October | 59.2 | 77.5 | 32.0 | 15.03 | 17.5 | 14.76 | 28.92 | 27.10 | 28.85 | 73.5 | 75.4 | 76.5 |
| November | 39.2 | 108.8 | 60.0 | 10.6 | 14.06 | 9.58 | 24.0 | 20.14 | 21.56 | 72.9 | 86.6 | 84.8 |
| December | 105.6 | 21.4 | 40.8 | 6.89 | 11.44 | 8.04 | 16.28 | 17.83 | 17.16 | 86.2 | 90.0 | 92.8 |
| January | 136.2 | 65 | 119.2 | 5.18 | 5.12 | 3.46 | 15.81 | 17.06 | 13.31 | 83.5 | 88.7 | 81.1 |
| February | 189.0 | 39.2 | 96.4 | 5.20 | 6.35 | 4.78 | 13.69 | 17.78 | 17.80 | 87.1 | 86.2 | 76.9 |
| March | 77.3 | 115.6 | 25.6 | 7.48 | 6.52 | 6.37 | 17.75 | 18.63 | 20.72 | 83.4 | 86.1 | 71.7 |
| April | 5.2 | 23.4 | 42.4 | 4.58 | 6.15 | 7.52 | 23.68 | 24.57 | 22.71 | 72.0 | 78.4 | 69.4 |
| May | 25.0 | 40.4 | 23.4 | 12.9 | 9.57 | 11.44 | 29.07 | 27.86 | 29.71 | 65.9 | 70.7 | 56.3 |
| Sum/Average * | 636.7 | 491.3 | 439.8 | 8.48 | 9.58 | 8.24 | 21.15 | 21.37 | 21.47 | 78.06 | 82.76 | 76.18 |
| pH | 7.2 | ||||
| Soil type | Vertosol (texture: Clay loam) | ||||
| Composition of Soil | |||||
| Depth | Clay (%) | Loam (%) | Sand (%) | Mineral N (ppm) | Total N (%) |
| 0–20 | 67.5 | 22.5 | 10 | 859 | 0.17 |
| 20–40 | 65 | 23.7 | 11.3 | 934.7 | 0.16 |
| SLB Severity (%) | SY (Mg ha−1) | GY (Mg ha−1) | TKW (g) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | NIC | IC | CB | CT | NIC | IC | CB | CT | NIC | IC | CB | CT | NIC | IC | CB | CT |
| Year | ||||||||||||||||
| 2015 | 36.0cd | 50.0ab | 20.0ef | 28.0de | 8.7c | 6.0ef | 13.3a | 13.9a | 1.8ef | 1.6fg | 2.0de | 1.6g | 49.8cd | 37.4f | 59.1a | 48.7d |
| 2016 | 15.0fg | 34.5cd | 10.6fg | 9.0fg | 8.1cd | 7.1de | 11.4b | 6.6ef | 2.4c | 2.1d | 3.0a | 2.5bc | 51.3cd | 48.7d | 57.8a | 50.2cd |
| 2017 | 43.5bc | 59.1a | 10.8fg | 5.9g | 4.5gh | 3.7h | 5.7fg | 6.1ef | 1.7fg | 1.2h | 2.4c | 2.7ab | 53.0bc | 44.4e | 55.6ab | 53.2bc |
| LSD | ||||||||||||||||
| 2015 | 16.46 | 18.85 | 10.54 | 11.35 | 1.47 | 0.60 | 0.90 | 0.98 | 0.06 | 0.07 | 0.02 | 0.05 | 6.50 | 2.46 | 1.64 | 2.10 |
| 2016 | 6.66 | 18.62 | 2.83 | 2.10 | 0.09 | 0.44 | 1.21 | 0.55 | 0.04 | 0.02 | 0.26 | 0.11 | 0.20 | 0.25 | 3.01 | 0.19 |
| 2017 | 10.04 | 25.83 | 6.80 | 2.95 | 0.30 | 0.20 | 0.95 | 0.08 | 0.12 | 0.14 | 0.15 | 0.31 | 0.40 | 2.66 | 0.66 | 1.40 |
| ANOVA | ||||||||||||||||
| Treatment (T) | 45.348 *** | 58.35 *** | 55.38 *** | 43.929 *** | ||||||||||||
| Year (Y) | 16.629 *** | 146.48 *** | 80.97 *** | 5.766 ** | ||||||||||||
| Interaction (T × Y) | 4.879 *** | 20.60 *** | 18.47 *** | 6.371 *** | ||||||||||||
| 2017 | SLB Severity (%) | SPAD | NDVI | Canopy Temperature (°C) | Stomatal Conductance (mmol.m−²·s−1) | Spikes.m−2 | Straw Yield (Mg. ha−1) | Grain Yield (Mg. ha−1) | Thousand Kernel Weigh (g) | Ngrain (%, g DW) | Isotopic Composition δ15Ngrain (‰) | Cgrain (%, g DW) | Isotopic Composition δ13Cgrain (‰) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NIC | 43.56b | 49.46b | 0.78a | 19.70a | 157.50d | 176.33bc | 4.50b | 1.75b | 53.06a | 1.79b | 0.77b | 35.87b | −24.62a |
| IC | 59.19a | 43.20c | 0.75b | 18.00b | 282.36a | 154.00c | 3.78b | 1.21c | 44.44b | 1.71b | 1.34a | 40.81a | −24.75b |
| CB | 10.82c | 53.20a | 0.76b | 16.99c | 234.53b | 232.00a | 5.77a | 2.48a | 55.69a | 1.97a | 0.05c | 44.22a | −25.31d |
| CT | 5.94d | 48.63b | 0.74b | 19.43a | 204.80c | 193.66b | 6.18a | 2.77a | 53.20a | 1.96a | 0.62b | 42.49a | −24.99c |
| LSD | |||||||||||||
| NIC | 1.20 | 0.37 | 0.05 | 0.36 | 12.82 | 14.57 | 0.30 | 0.12 | 1.40 | 0.04 | 0.15 | 4.81 | 0.05 |
| IC | 0.46 | 0.36 | 0.00 | 0.17 | 10.26 | 10.14 | 0.20 | 0.14 | 2.66 | 0.03 | 0.14 | 0.77 | 0.03 |
| CB | 0.58 | 1.99 | 0.01 | 0.15 | 3.29 | 24.97 | 0.95 | 0.15 | 0.66 | 0.07 | 0.01 | 1.40 | 0.02 |
| CT | 0.22 | 0.30 | 0.01 | 0.15 | 4.55 | 9.81 | 0.08 | 0.31 | 0.40 | 0.06 | 0.05 | 0.27 | 0.09 |
| ANOVA Treatment | 3872.00 *** | 46.89 *** | 10.03 ** | 95.03 *** | 109.30 *** | 12.58 ** | 14.07 ** | 37.08 *** | 30.10 *** | 16.11 *** | 69.74 *** | 6.039 * | 79.45 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Jabeur, M.; Kthiri, Z.; Harbaoui, K.; Belguesmi, K.; Serret, M.D.; Araus, J.L.; Hamada, W. Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat. Agronomy 2019, 9, 586. https://doi.org/10.3390/agronomy9100586
Ben-Jabeur M, Kthiri Z, Harbaoui K, Belguesmi K, Serret MD, Araus JL, Hamada W. Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat. Agronomy. 2019; 9(10):586. https://doi.org/10.3390/agronomy9100586
Chicago/Turabian StyleBen-Jabeur, Maissa, Zayneb Kthiri, Kalthoum Harbaoui, Karima Belguesmi, Maria Dolores Serret, Jose Luis Araus, and Walid Hamada. 2019. "Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat" Agronomy 9, no. 10: 586. https://doi.org/10.3390/agronomy9100586
APA StyleBen-Jabeur, M., Kthiri, Z., Harbaoui, K., Belguesmi, K., Serret, M. D., Araus, J. L., & Hamada, W. (2019). Seed Coating with Thyme Essential Oil or Paraburkholderia phytofirmans PsJN Strain: Conferring Septoria Leaf Blotch Resistance and Promotion of Yield and Grain Isotopic Composition in Wheat. Agronomy, 9(10), 586. https://doi.org/10.3390/agronomy9100586