Growth Performance and Antioxidant Enzyme Activities of Advanced Mutant Rice Genotypes under Drought Stress Condition

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Site
2.2. Experimental Layout
2.3. Drought Stress Treatment
2.4. Growth Measurement
2.4.1. Vegetative Growth
2.4.2. Dry Weight Production and Partitioning
2.5. Biochemical Analysis
2.5.1. Chlorophyll Content Assay
2.5.2. Proline Content Assay
2.5.3. Enzyme Extract Preparation and Antioxidant Enzymes Assay
2.6. Yield and Yield Components
2.7. Statistical Analysis
3. Results
3.1. Plant Height, Number of Tillers, and Flag Leaf Area
3.2. Culms and Leaves Dry Weight, Panicles Dry Weight, and Total Dry Weight
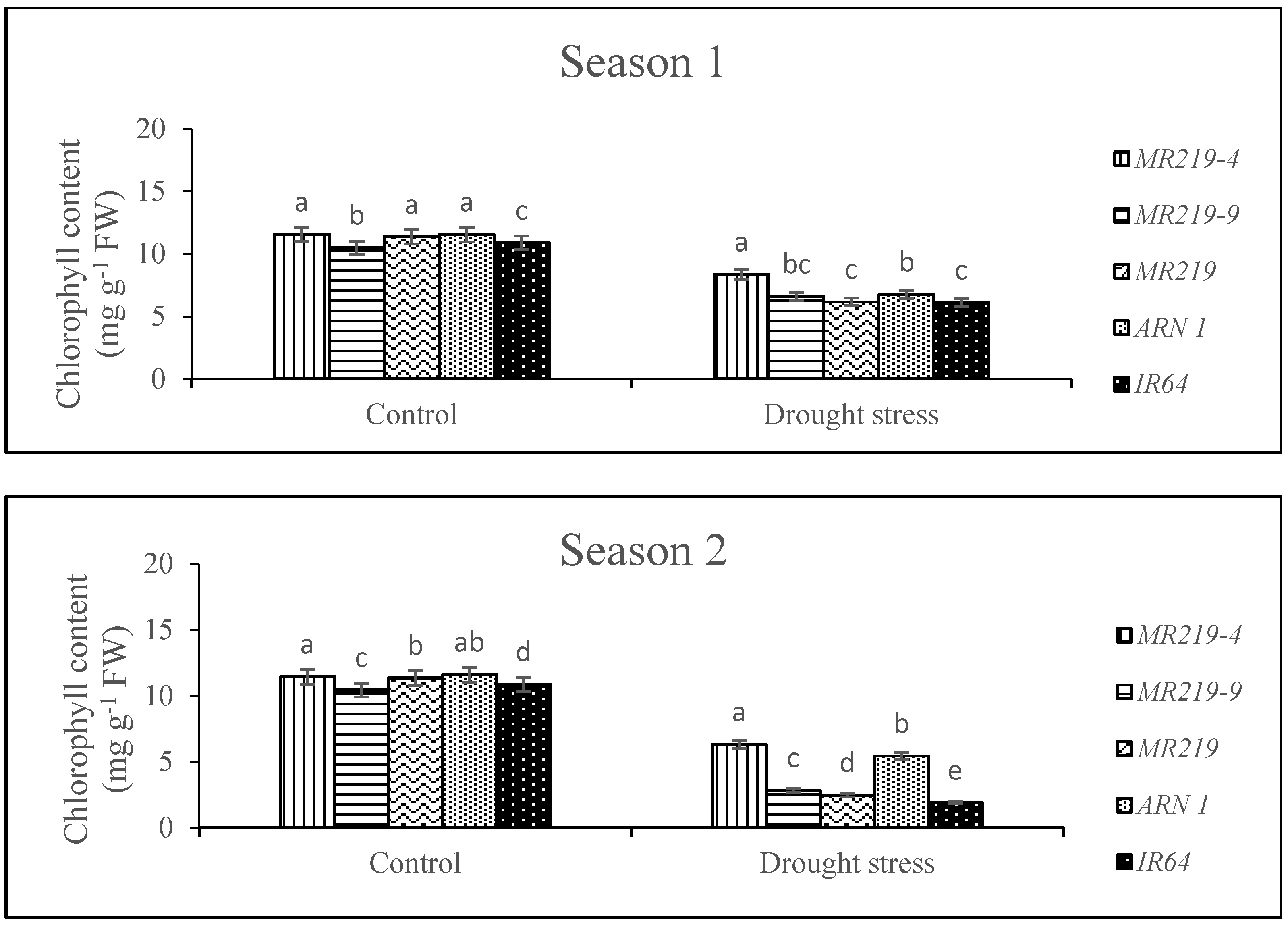
3.3. Chlorophyll Content
3.4. Proline Content
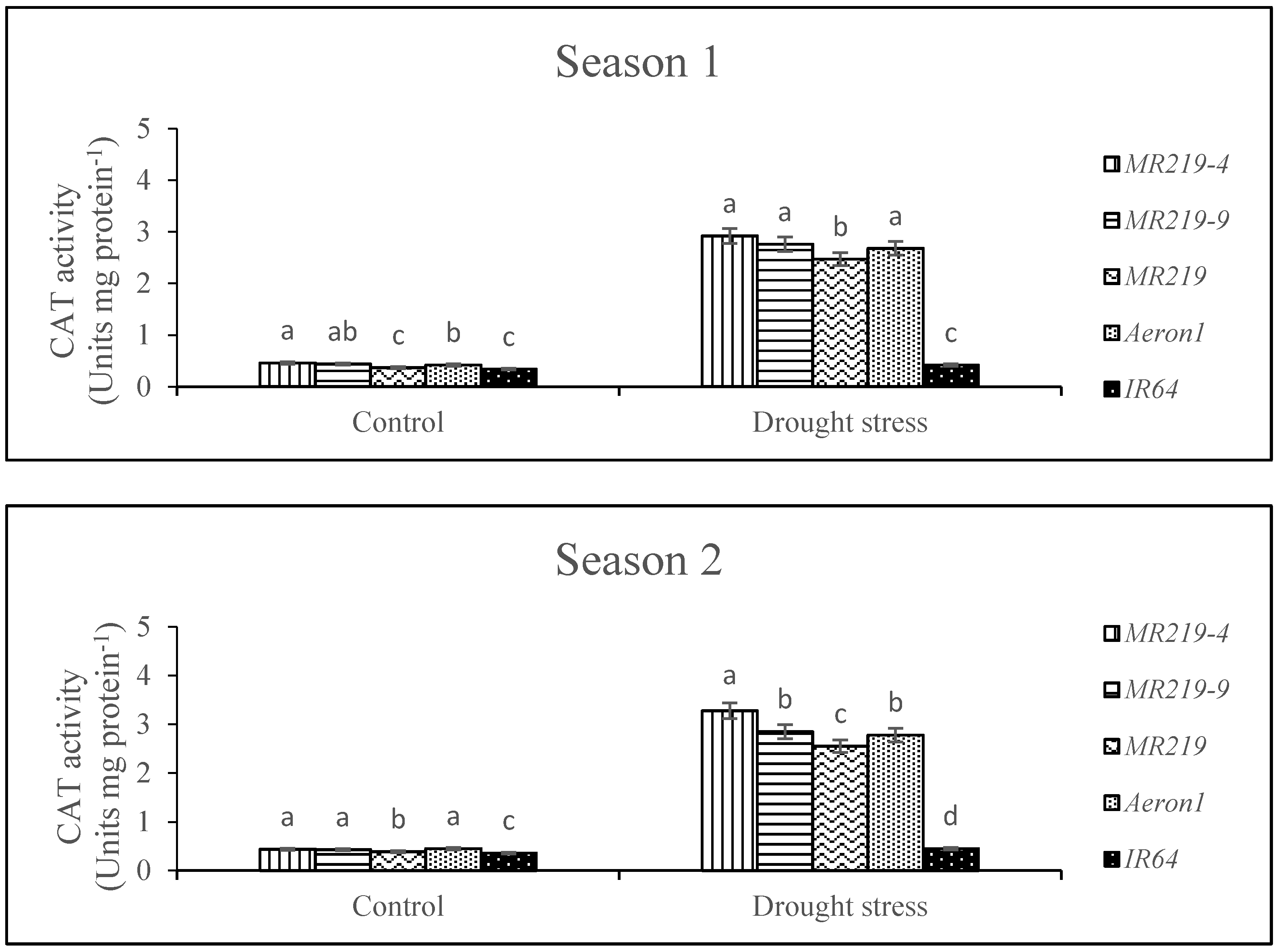
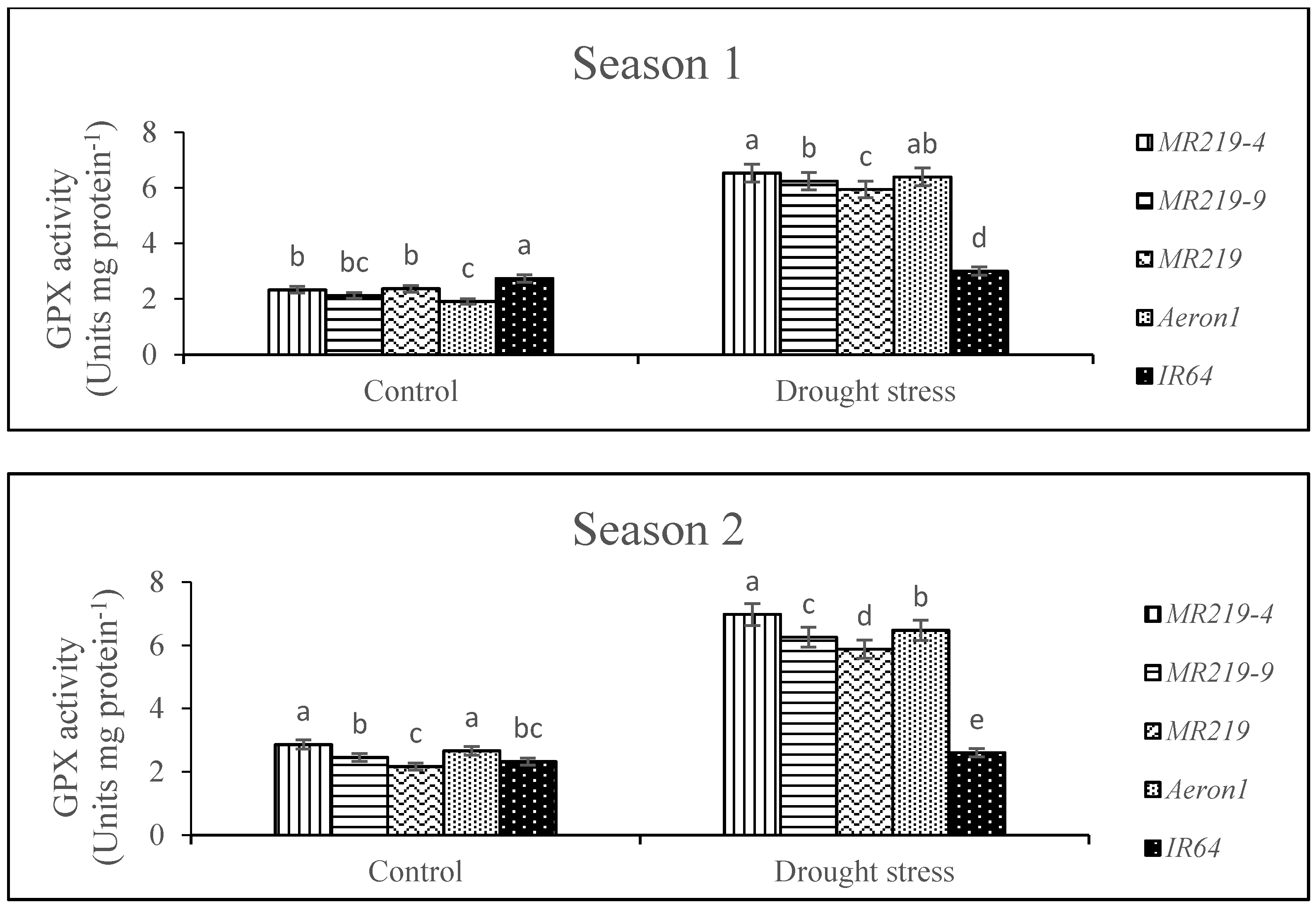
3.5. Activities of Antioxidant Enzymes
3.6. Yield and Yield Components
4. Discussion
4.1. Effects of Drought on Rice Vegetative Growth
4.2. Effects of Drought Stress on Dry Weight Production and Partitioning
4.3. Effects of Drought Stress on Chlorophyll Content
4.4. Effects of Drought Stress on Proline Content
4.5. Effects of Drought Stress on Activity of Antioxidant Enzymes
4.6. Effects of Drought Stress on Yield and Yield Components
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GIEWS—Global Information and Early Warning System. Available online: http://www.fao.org/giews/countrybrief/country.jsp?code=MYS (accessed on 15 December 2017).
- Tiara, H.; Erik, H.M.; Asgar, A.W. Rice production and climate change: A case study of Malaysian rice. Pertanika J. Trop. Agric. Sci. 2015, 38, 321–328. [Google Scholar]
- Dortje, G.; Chao, L.; Harikrishnan, M.; Nina, P. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Abdul, W.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress from Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. ISBN 978-3-642-32653-0. [Google Scholar]
- Joseph, E.A.; Radhakrishnan, V.V.; Mohanan, K.V. A Study on the accumulation of proline-an osmoprotectant amino acid under salt stress in some native rice cultivars of North Kerala, India. Univers. J. Agric. Res. 2015, 3, 15–22. [Google Scholar] [CrossRef]
- Pallavi, S.; Rama, S.D. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Reg. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Gregor, J.S.; Stefan, K.A. Osmotic adjustment under drougth conditions. In Plant Responses to Drought Stress; Springer: Berlin/Heidelberg, Germany, 2012; pp. 199–229. [Google Scholar]
- Sara, I.; Zandalinas, R.M.; Damián, B.; Vicent, A.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2017, 162, 2–12. [Google Scholar] [CrossRef]
- Sankar, B.; Jaleel, C.A.; Manivanna, P.; Kishorekumar, A.; Somasundaram, R.; Panneerselvam, R. Effect of paclobutrazol on water stress amelioration through antioxidants and free radical scavenging enzymes in Arachis hypogaea L. Colloids Surf. B 2007, 60, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Hojati, M.; Modarres-Sanavy, S.A.M.; Karimi, M.; Ghanati, F. Responses of growth and antioxidant systems in Carthamus tinctorius L. under water deficit stress. Acta Physiol. Plant. 2011, 33, 105–112. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 2, 185–212. [Google Scholar] [CrossRef]
- Clifford, S.C.; Kadzere, I.; Jones, H.G.; Jackson, J.E. Field comparisons of photosynthesis and leaf conductance in Ziziphus mauritiana and other fruit tree species in Zimbabwe. Trees 1997, 11, 449–455. [Google Scholar] [CrossRef]
- IRRI—International Rice Research Institute. Standard Evaluation System for Rice (SES). Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf (accessed on 28 August 2012).
- Ashraf, M.Y.; Akhtar, K.; Sarwar, G.; Ashraf, M. Role of rooting system in salt tolerance potential of different guar accessions. Agron. Sustain. Dev. 2005, 25, 243–249. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.K. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–208. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P.; Murr, D.P.; Watkins, C.B. Influence of salicylic acid on H2O2 production, oxidative stress, and H2O2-metabolizing enzymes. Salicylic acid-mediated oxidative damage requires H2O2. Plant Physiol. 1997, 115, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell. Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Bhupinder, S.; Kambham, R.R.; Edilberto, D.R.; Timothy, W. Screening of rice cultivars for morpho-physiological responses to early-season soil moisture stress. Rice Sci. 2017, 24, 322–335. [Google Scholar] [CrossRef]
- Henry, A.; Wehler, R.; Grondin, A.; Franke, R.; Quintana, M. Environmental and physiological effects on grouping of drought-tolerant and susceptible rice varieties related to rice (Oryza sativa) root hydraulics under drought. Ann. Bot. 2016, 118, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Solomon, Z.; Mats, O.; Masresha, F. Growth, gas exchange, chlorophyll a fluorescence, biomass accumulation and partitioning in droughted and irrigated plants of two enset (Ensete ventricosum Welw, Cheesman) clones. J. Agron. 2007, 6, 499–508. [Google Scholar] [CrossRef]
- Remi, L.; Sylvain, L.C.; Rossitza, A.; Fabienne, D.; Thierry, A.; Nathalie, P.; Jean-Louis, B.; Maryse, L.; Pierre, C.T.; Laurence, M.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 1–21. [Google Scholar] [CrossRef]
- Cox, J.A.; Conran, J.G. The effect of water stress on the life cycles of Erodium crinitum Carolin and Erodium cicutarium (L.) L’Hérit. ex Alton (Geraniaceae). Aust. J. Ecol. 1996, 21, 235–240. [Google Scholar] [CrossRef]
- Rodrigues, M.L.; Pacheco, C.M.A.; Chaves, M.M. Soil-plant water relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. J. Exp. Bot. 1995, 46, 947–956. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Noitsakis, B.; Therios, I. Photosynthesis, plant growth and dry matter distribution in kiwifruit as influenced by water deficits. Irrigation Sci. 1993, 14, 1–5. [Google Scholar] [CrossRef]
- Hamblin, A.; Tennant, D.; Perry, M.W. The cost of stress: Dry matter partitioning changes with seasonal supply of water and nitrogen to dryland wheat. Plant Soil 1990, 122, 47–58. [Google Scholar] [CrossRef]
- Kumar, S.; Dey, P. Effects of different mulches and irrigation methods on root growth, nutrient uptake, water-use efficiency and yield of strawberry. Sci. Hort. 2011, 127, 318–324. [Google Scholar] [CrossRef]
- Samuel, D.S.P.; Anna, E.D.S.; Marco, A.O.; Eduardo, G.P. Oxidative damage and photosynthetic impairment in tropical rice cultivars upon exposure to excess iron. Sci. Agric. 2015, 73, 217–226. [Google Scholar] [CrossRef]
- Mohammad Reza, N.R.; Mihdzar, A.K.; Mohd Rafii, Y. Genetic behaviour for plant capacity to produce chlorophyll in wheat (Triticum aestivum) under drought stress. Aust. J. Crop. Sci. 2012, 6, 415–420. [Google Scholar]
- Nurul, H.; Triadiati; Iswandi, A. Photosynthesis and transpiration rates of rice cultivated under the system of rice intensification and the effects on growth and yield. HAYATI J. Biosci. 2016, 23, 67–72. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Mahbod, S.; Mohamed, M.H.; Rafii, M.Y.; Mahmud, T.M.M.; Parisa, A.; Mohama, O.; Rambod, A.; Sima, T.; Nahid, K.; Shabanimofrad, M.; et al. Improvement of drought tolerance in rice (Oryza sativa L.): Genetics, genomic tools, and the WRKY gene family. BioMed Res. Intern. 2018, 2018, 1–20. [Google Scholar] [CrossRef]
- Lum, M.S.; Hanafi, M.M.; Rafii, M.Y.; Akmar, A.S.N. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Anjum, S.A.; Farooq, M.; Xie, X.Y.; Liu, X.J.; Ijaz, M.F. Antioxidant defense system and proline accumulation enables hot pepper to perform better under drought. Sci. Hortic. 2012, 140, 66–73. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Lelandais, M.; Korth, H.G.; Foyer, C.H. Ascorbate is the natural substrate for plant peroxidases. FEBS J. 1996, 378, 203–206. [Google Scholar] [CrossRef]
- Shamsun, N.; Lakshminarayana, R.V.; Lingaraj, S.; Bhaben, T. Antioxidant protection mechanisms reveal significant response in drought-induced oxidative stress in some traditional rice of Assam, India. Rice Sci. 2018, 25, 185–196. [Google Scholar] [CrossRef]
- Nadzariah, K.Z.; Mohd, Y.A.; Sariam, O.; Nadzirah, K.Z. Growth and physiological performance of aerobic and lowland rice as affected by water stress at selected growth stages. Rice Sci. 2018, 25, 82–93. [Google Scholar] [CrossRef]
- Mayumi, K.; Daigo, M.; Naoya, A.; Akira, M.; Yoshinori, Y. Growth and yield responses of upland NERICAs to variable water management under field conditions. Plant Prot. Sci. 2017, 20, 36–46. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Guan, Y.S.; Yan, S.; Li, Z.K. Whole plant responses, key processes, and adaptation to drought stress: The case of rice. J. Exp. Bot. 2007, 58, 169–175. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | Mean Squares | ||
|---|---|---|---|---|
| Plant Height | Number of Tillers | Flag Leaf Area | ||
| Seasons (S) | 1 | 35.25 * | 316.01 ** | 622.11 ** |
| Replications within season (R/S) | 6 | 3.39 | 0.80 | 91.81 |
| Water treatments (W) | 1 | 338.25 ** | 132.61 ** | 371.13 ** |
| S × W | 1 | 30.63 | 9.11 | 300.66 |
| W × (R/S) | 6 | 5.48 | 0.63 | 28.21 |
| Genotypes (G) | 4 | 599.94 ** | 147.89 ** | 120.04 ** |
| G × S | 4 | 71.86 ** | 49.73 ** | 71.05 ** |
| G × W | 4 | 231.60 ** | 8.21 ** | 108.30 ** |
| G × S × W | 4 | 32.22 ** | 2.77 * | 49.97 ** |
| Error | 48 | 3.07 | 0.98 | 7.38 |
| Treatment/ Genotype | Vegetative Growth | |||||
|---|---|---|---|---|---|---|
| Plant Height (cm) | Number of Tillers | Flag Leaf Area (cm) | ||||
| Control | Drought Stress | Control | Drought Stress | Control | Drought Stress | |
| Season 1 | ||||||
| MR219-4 | 98.4b | 94.1b | 15a | 13a | 41.3a | 40.7a |
| MR219-9 | 99.3b | 90.5b | 16a | 10a | 43.0a | 34.0ab |
| MR219 | 98.7b | 92.2b | 14b | 7b | 39.4a | 36.6ab |
| Aeron1 | 112.1a | 101.8a | 11c | 9b | 44.1a | 38.9ab |
| IR64 | 94.5c | 84.1c | 14b | 5c | 30.7a | 12.2b |
| Mean | 100.6 | 92.5 | 14 | 9 | 39.7 | 32.5 |
| CV (%) | 3.10 | 10.53 | 25.66 | 28.86 | 21.16 | 24.98 |
| Season 2 | ||||||
| MR219-4 | 97.1b | 91.6b | 18a | 13a | 38.8a | 29.9a |
| MR219-9 | 99.9b | 93.3bc | 15b | 9a | 37.3a | 24.7a |
| MR219 | 94.0c | 88.8c | 11b | 7ab | 39.7a | 19.3b |
| Aeron1 | 111.7a | 105.7a | 13b | 11a | 32.3a | 27.8a |
| IR64 | 96.5b | 82.0d | 17ab | 4b | 32.9a | 10.1c |
| Mean | 99.8 | 92.3 | 15 | 9 | 36.2 | 22.4 |
| CV (%) | 5.33 | 8.74 | 37.45 | 40.43 | 18.92 | 21.37 |
| Source of Variation | df | Mean Squares | ||
|---|---|---|---|---|
| Culms and Leaves Dry Weight | Panicles Dry Weight | Total Dry Weight | ||
| Seasons (S) | 1 | 177.34 ** | 897.80 ** | 705.97 ** |
| Replications within season (R/S) | 6 | 0.40 | 3.37 | 5.19 |
| Water treatment (W) | 1 | 2193.57 ** | 361.25 ** | 11,336.18 ** |
| S × W | 1 | 0.11 * | 101.25 ** | 465.08 ** |
| W × (R/S) | 6 | 0.18 | 6.72 | 4.28 |
| Genotypes (G) | 4 | 153.04 ** | 1472.39 ** | 3492.92 ** |
| G × S | 4 | 16.54 ** | 176.18 ** | 86.40 ** |
| G × W | 4 | 10.27 ** | 294.75 ** | 1360.41 ** |
| G × S× W | 4 | 3.49 ** | 55.06 ** | 153.91 ** |
| Error | 48 | 0.19 | 5.14 | 4.52 |
| Treatment/ Genotype | Dry Weight Production and Partitioning | |||||
|---|---|---|---|---|---|---|
| Culms and Leaves Dry Weight (g Plant−1) | Panicles Dry Weight (g Plant−1) | Total Dry Weight (g Plant−1) | ||||
| Control | Drought Stress | Control | Drought Stress | Control | Drought Stress | |
| Season 1 | ||||||
| MR219-4 | 18.46c | 13.22a | 79.00b | 76.00a | 68.86b | 58.56a |
| MR219-9 | 25.54a | 8.74b | 81.50a | 72.50a | 81.47a | 51.28b |
| MR219 | 24.41b | 8.28b | 72.75c | 51.25c | 69.65b | 42.30c |
| Aeron1 | 17.86c | 12.49a | 57.75d | 68.50b | 59.86c | 54.29b |
| IR64 | 14.52d | 5.33c | 73.50c | 51.75d | 69.99b | 40.25c |
| Mean | 20.16 | 9.61 | 71.70 | 65.20 | 69.97 | 49.34 |
| CV (%) | 21.30 | 31.44 | 15.12 | 14.89 | 29.09 | 15.26 |
| Season 2 | ||||||
| MR219-4 | 13.72d | 8.50a | 86.50a | 81.25a | 89.71b | 77.89a |
| MR219-9 | 20.62a | 6.76b | 88.75a | 75.00b | 97.61a | 63.80b |
| MR219 | 20.51a | 4.99c | 79.00b | 72.48b | 70.77b | 57.51c |
| Aeron1 | 14.80c | 8.36a | 78.00b | 60.25c | 73.27b | 65.71b |
| IR64 | 15.88b | 4.92c | 73.50c | 55.75d | 71.64b | 47.30d |
| Mean | 17.11 | 6.71 | 76.15 | 74.15 | 79.09 | 64.09 |
| CV (%) | 17.54 | 24.48 | 13.12 | 15.09 | 25.26 | 17.96 |
| Source of Variation | df | Mean Squares | ||||
|---|---|---|---|---|---|---|
| Relative Chlorophyll Content | Proline Content | CAT Activity | APX Activity | GPX Activity | ||
| Seasons (S) | 1 | 45.52 ** | 0.08 ** | 0.09 ** | 0.25 ** | 0.09 * |
| Replications within season (R/S) | 6 | 0.08 | 0.01 | 0.03 | 0.04 | 0.01 |
| Water treatments (W) | 1 | 689.68 ** | 6.78 ** | 90.19 ** | 64.87 ** | 261.44 ** |
| S × W | 1 | 44.21 ** | 0.14 ** | 0.07 ** | 0.62 ** | 0.34 * |
| WT × (R/S) | 6 | 0.05 | 0.04 | 0.04 | 0.07 | 0.01 |
| Genotypes (G) | 4 | 3.04 ** | 0.21 ** | 1.64 ** | 4.24 ** | 2.47 ** |
| G × S | 4 | 5.44 ** | 0.24 ** | 0.01 * | 0.10 ** | 0.83 ** |
| G × W | 4 | 6.58 ** | 0.14 ** | 1.25 ** | 2.91 ** | 2.83 ** |
| G × S × W | 4 | 5.60 ** | 0.22 ** | 0.02 ** | 0.13 ** | 0.01 * |
| Error | 48 | 0.04 | 0.07 | 0.03 | 0.09 | 0.02 |
| Source of Variation | df | Mean Squares | |||
|---|---|---|---|---|---|
| Grain Yield | 1000 Grain Weight | Spikelets/Panicle | Harvest Index | ||
| Seasons (S) | 1 | 5134.89 ** | 12.91 ** | 1209.01 ** | 0.551 ** |
| Replications within season (R/S) | 6 | 10.79 | 0.33 | 25.83 | 0.002 |
| Water treatments (W) | 1 | 266.49 ** | 72.41 ** | 308.11 ** | 0.105 ** |
| S × W | 1 | 665.34 ** | 0.69 | 262.81 ** | 0.002 |
| W × (R/S) | 6 | 5.81 | 0.72 | 13.93 | 0.002 |
| Genotypes (G) | 4 | 1878.03 ** | 81.39 ** | 1625.18 ** | 0.010 |
| G × S | 4 | 495.85 ** | 4.43 * | 264.95 ** | 0.036 ** |
| G × W | 4 | 323.75 ** | 1.87 * | 951.30 ** | 0.024 ** |
| G × S × W | 4 | 539.71 ** | 2.23 * | 50.00 * | 0.008 |
| Error | 48 | 8.00 | 0.61 | 7.60 | 0.001 |
| Treatment/ Genotype | Yield and Yield Attributes | |||||||
|---|---|---|---|---|---|---|---|---|
| Grain Yield (g Plant−1) | 1000 Grain Weight (g) | Spikelets/Panicle | Harvest Index | |||||
| Control | Drought Stress | Control | Drought Stress | Control | Drought Stress | Control | Drought Stress | |
| Season 1 | ||||||||
| MR219-4 | 34.00bc | 32.58b | 29.02b | 28.06a | 60b | 59a | 0.50b | 0.60ab |
| MR219-9 | 59.81a | 34.38ab | 30.32a | 28.44a | 68a | 62a | 0.49b | 0.67a |
| MR219 | 36.62bc | 35.03a | 27.94c | 26.25b | 51c | 46b | 0.53b | 0.60ab |
| Aeron1 | 30.52c | 25.36c | 27.53c | 26.17b | 44d | 43b | 0.51b | 0.63ab |
| IR64 | 47.72b | 24.73c | 23.69d | 21.00c | 68a | 33c | 0.68a | 0.58b |
| Mean | 37.14 | 35.02 | 27.70 | 25.98 | 57 | 49 | 0.54 | 0.62 |
| CV (%) | 21.31 | 34.31 | 8.54 | 10.97 | 16.10 | 22.04 | 14.25 | 34.31 |
| Season 2 | ||||||||
| MR219-4 | 53.15b | 50.88b | 28.11a | 27.10a | 69b | 63b | 0.76a | 0.80a |
| MR219-9 | 92.17a | 65.49a | 29.02a | 26.67a | 81a | 76a | 0.79a | 0.84a |
| MR219 | 47.69c | 45.87bc | 26.15b | 24.84b | 62c | 56c | 0.65c | 0.80a |
| Aeron1 | 46.82c | 44.83c | 28.00a | 23.81c | 64bc | 50d | 0.71b | 0.83a |
| IR64 | 46.22c | 29.89d | 24.13c | 22.56d | 61bc | 28d | 0.65c | 0.63b |
| Mean | 56.81 | 47.39 | 27.08 | 24.99 | 61 | 55 | 0.71 | 0.78 |
| CV (%) | 32.54 | 25.61 | 7.09 | 7.39 | 15.31 | 30.88 | 8.78 | 12.05 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamarudin, Z.S.; Yusop, M.R.; Tengku Muda Mohamed, M.; Ismail, M.R.; Harun, A.R. Growth Performance and Antioxidant Enzyme Activities of Advanced Mutant Rice Genotypes under Drought Stress Condition. Agronomy 2018, 8, 279. https://doi.org/10.3390/agronomy8120279
Kamarudin ZS, Yusop MR, Tengku Muda Mohamed M, Ismail MR, Harun AR. Growth Performance and Antioxidant Enzyme Activities of Advanced Mutant Rice Genotypes under Drought Stress Condition. Agronomy. 2018; 8(12):279. https://doi.org/10.3390/agronomy8120279
Chicago/Turabian StyleKamarudin, Zarifth Shafika, Mohd Rafii Yusop, Mahmud Tengku Muda Mohamed, Mohd Razi Ismail, and Abdul Rahim Harun. 2018. "Growth Performance and Antioxidant Enzyme Activities of Advanced Mutant Rice Genotypes under Drought Stress Condition" Agronomy 8, no. 12: 279. https://doi.org/10.3390/agronomy8120279
APA StyleKamarudin, Z. S., Yusop, M. R., Tengku Muda Mohamed, M., Ismail, M. R., & Harun, A. R. (2018). Growth Performance and Antioxidant Enzyme Activities of Advanced Mutant Rice Genotypes under Drought Stress Condition. Agronomy, 8(12), 279. https://doi.org/10.3390/agronomy8120279