Development and Use of Simple Sequence Repeats (SSRs) Markers for Sugarcane Breeding and Genetic Studies




Abstract
1. Introduction
2. Development of Microsatellite (SSR) Markers
2.1. Next-Generation Sequencing or High Throughput Sequencing
2.2. Expressed Sequence Tag (EST) Libraries
2.3. Enriched Genomic Libraries
3. Detection Systems for SSRs
3.1. Capillary Electrophoresis
3.2. Polyacrylamide Gel Electrophoresis
4. Application of SSR markers
4.1. Cultivars Identification
4.2. Genetic Diversity/Phylogenetic Relationship
4.3. Genome Mapping
4.4. Quantitative Trait Loci (QTL) Analysis
4.5. Paternity Analysis
4.6. Cross-Species Transferability
4.7. Segregation Analysis
4.8. SSR-Assisted Identification of Wild Cross Hybrids
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- UN Food and Agriculture Organization. Sugarcane Production in 2016, Crops/Regions/World List/Production Quantity (Pick Lists); Corporate Statistical Database (FAOSTAT): Rome, Italy, 2017. [Google Scholar]
- Li, X.Y.; Sun, H.D.; Rott, P.C.; Wang, J.D.; Huang, M.T.; Zhang, Q.Q.; Gao, S.J. Molecular identification and prevalence of Acidovorax avenae subsp. avenae causing red stripe of sugarcane in China. Plant Pathol. 2018, 67, 929–937. [Google Scholar]
- Kumar, U.; Priyanka; Kumar, S. Genetic Improvement of Sugarcane through Conventional and Molecular Approaches; Molecular Breeding for Sustainable Crop Improvement; Springer: Cham, Switzerland, 2016; pp. 325–342. [Google Scholar] [CrossRef]
- D’Hont, A.; Grivet, L.; Feldmann, P.; Glaszmann, J.C.; Rao, S.; Berding, N. Characterisation of the double genome structure of modern sugarcane cultivars (Saccharum spp.) by molecular cytogenetics. Mol. Gen. Genet. 1996, 250, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.B. Databasing molecular identities of sugarcane (Saccharum spp.) clones constructed with microsatellite (SSR) DNA markers. Am. J. Plant Sci. 2010, 1, 87–94. [Google Scholar] [CrossRef]
- Lu, Y.H.; D’Hont, A.; Walker, D.I.T.; Rao, P.S.; Feldmann, P.; Glaszmann, J.C. Relationships among ancestral species of sugarcane revealed with RFLP using single copy maize nuclear probes. Euphytica 1994, 78, 7–18. [Google Scholar]
- Nawaz, S.; Khan, F.A.; Tabasum, S.; Zakria, M.; Saeed, A.; Iqbal, M.Z. Phylogenetic relationships among Saccharum clones in Pakistan revealed by RAPD markers. Genet. Mol. Res. 2010, 9, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Besse, P.; Taylor, G.; Carroll, B.; Berding, N.; Burner, D.; McIntyre, C.L. Assessing genetic diversity in a sugarcane germplasm collection using an automated AFLP analysis. Genetica 1998, 104, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, X.; Huang, F.; Huang, Y.; Liu, X.; Wu, J.; Wang, Q.; Deng, Z.; Chen, R.; Zhang, M. A new method based on SNP of nrDNA-ITS to identify Saccharum spontaneum and its progeny in the genus Saccharum. PLoS ONE 2018, 13, e0197458. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Pan, Y.B.; Tew, T.L.; Schnell, R.J.; Viator, R.P. Microsatellite DNA marker-assisted selection of Saccharum spontaneum cytoplasm-derived germplasm. Sugar Tech. 2006, 8, 23–29. [Google Scholar] [CrossRef]
- Ahmad, A.; Wang, J.-D.; Pan, Y.-B.; Deng, Z.-H.; Chen, Z.-W.; Chen, R.-K.; Gao, S.-J. Molecular Identification and Genetic diversity analysis of chinese sugarcane (Saccharum spp. Hybrids) varieties using SSR markers. Trop. Plant Biol. 2017, 10, 194–203. [Google Scholar]
- Pan, Y.B. Development and Integration of an SSR-Based Molecular Identity Database into Sugarcane Breeding Program. Agronomy 2016, 6, 28. [Google Scholar] [CrossRef]
- Pan, Y.B.; Scheffler, B.S.; Richard, E.P., Jr. High throughput molecular genotyping of commercial sugarcane clones with microsatellite (SSR) DNA markers. Sugar Tech. 2007, 9, 176–181. [Google Scholar]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2006, 9, 615–629. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Barkley, N.A.; Jenkins, T.M. Microsatellite markers in plants and insects. Part I. Applications of biotechnology. Genes Genomes Genomics 2009, 3, 54–67. [Google Scholar]
- Soranzo, N.; Provan, J.; Powell, W. An example of microsatellite length variation in the mitochondrial genome of conifers. Genome 1999, 42, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Weising, K.; Gardner, R.C. A set of conserved PCR primers for the analysis of simple sequence repeat polymorphisms in chloroplast genomes of dicotyledonous angiosperms. Genome 1999, 42, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Vettore, A.L.; da Silva, F.R.; Kemper, E.L.; Souza, G.M.; da Silva, A.M.; Ferro, M.I.; Henrique-Silva, F.; Giglioti, É.A.; Lemos, M.V.; Coutinho, L.L.; et al. Analysis and functional annotation of an expressed sequence tag collection for tropical crop sugarcane. Genome Res. 2003, 13, 2725–2735. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.R.; Oliveira, K.M.; Ulian, E.C.; Garcia, A.A.F.; De Souza, A.P. Survey in the sugarcane expressed sequence tag database (SUCEST) for simple sequence repeats. Genome 2004, 47, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.R.; Oliveira, K.M.; Marconi, T.; Garcia, A.A.F.; Ulian, E.C.; Souza, A.P. Characterization of novel sugarcane expressed sequence tag microsatellites and their comparison with genomics SSRs. Plant Breed. 2006, 125, 378–384. [Google Scholar] [CrossRef]
- Oliveira, K.M.; Pinto, L.R.; Marconi, T.G.; Mollinari, M.; Ulian, E.C.; Chabregas, S.M.; Falco, M.C.; Burnquist, A.A.F.; Garcia, A.A.F.; Souza, A.P. Characterization of new polymorphic functional markers for sugarcane. Genome 2009, 52, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Vettore, A.L.; da Silva, F.R.; Kemper, E.L.; Arruda, P. The libraries that made SUCEST. Genet. Mol. Biol. 2001, 24, 1–7. [Google Scholar] [CrossRef]
- Cordeiro, G.M.; Taylor, G.O.; Henry, R.J. Characterization of microsatellite markers from sugarcane (Saccharum sp.), a highly polyploid species. Plant Sci. 2000, 155, 161–168. [Google Scholar] [CrossRef]
- Cordeiro, G.M.; Casu, R.; Mclntyre, C.L.; Manners, J.M.; Henry, R.J. Microsatellite markers from sugarcane (Saccharum sp.) ESTs cross transferable to Erianthus and Sorghum. Plant Sci. 2001, 160, 1115–1123. [Google Scholar] [CrossRef]
- Kalia, R.; Manoj, R.; Sanjay, K.; Rohtas, S.; Ashok, D. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Singh, R.K.; Jena, S.N.; Khan, S.; Yadav, S.; Banarjee, N.; Raghuvanshi, S.; Bhardwaj, V.; Dattamajumder, S.K.; Kapur, R.; Solomon, S.; et al. Development, cross-species/genera transferability of novel EST-SSR markers and their utility in revealing population structure and genetic diversity in sugarcane. Gene 2013, 524, 309–329. [Google Scholar] [CrossRef] [PubMed]
- Kalwade, S.; Devarumath, R.; Harinath, B.K. Functional Microsatellite Marker Development from the Sugarcane Sequence Tag Database (SUCEST) using Bioinformatics Tools. In Proceedings of the International Plant & Animal Genome Conference XXI, San Diego, CA, USA, 12–16 January 2013. [Google Scholar]
- Kushwah; Singh, R.B. Development of Novel EST derived Microsatellite markers for high sucrose content and their Application in Sugarcane molecular breeding. In Proceedings of the STAI Annual Convention and International Sugar Expo, Delhi, India, 28–30 July 2016. [Google Scholar]
- Ul Haq, S.; Kumar, P.; Singh, R.K.; Verma, K.S.; Bhatt, R.; Sharma, M.; Kachhwaha, S.; Kothari, S.L. Assessment of functional EST-SSR markers (Sugarcane) in cross-species transferability, genetic diversity among poaceae plants, and bulk segregation analysis. Genet. Res. Int. 2016, 2016, 7052323. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.K.; Kalia, S.K.; Sunita, K.; Dalal, V.; Hemaprabha, G.; Selvi, A.; Pandit, A.; Singh, A.; Gaikwad, K.; Sharma, T.R.; et al. Informative genomic microsatellite markers for efficient genotyping applications in sugarcane. Theor. Appl. Genet. 2009, 118, 327–338. [Google Scholar] [CrossRef] [PubMed]
- James, B.T.; Chen, C.; Rudolph, A.; Swaminathan, K.; Murray, J.E.; Na, J.K.; Spence, A.K.; Smith, B.; Hudson, M.E.; Moose, S.P.; et al. Development of microsatellite markers in autopolyploid sugarcane and comparative analysis of conserved microsatellites in sorghum and sugarcane. Mol. Breed. 2012, 30, 661–669. [Google Scholar] [CrossRef]
- Parshant, S.; Manish, D.S.; Upma, D.; Sundip, K. Development of Microsatellite Markers from Enriched Genomic Library of ISH 100 (Saccharum hybrid) for Genetic Variability of Sugarcane. Indian J. Plant Genet. Res. 2014, 27, 178–180. [Google Scholar]
- Tsuruta, S.I.; Ebina, M.; Kobayashi, M.; Takahashi, W.; Terajima, Y. Development and validation of genomic simple sequence repeat markers in Erianthus arundinaceus. Mol. Breed. 2017, 37, 71. [Google Scholar] [CrossRef]
- Cardoso-Silva, C.B.; Costa, E.A.; Mancini, M.C.; Balsalobre, T.W.A.; Canesin, L.E.C.; Pinto, L.R.; Carneiro, M.S.; Garcia, A.A.F.; de Souza, A.P.; Vicentini, R. De novo assembly and transcriptome analysis of contrasting sugarcane varieties. PLoS ONE 2014, 9, e88462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Yan, J.J.; Shen, X.Y.; Bai, S.Q.; Li, D.X.; Zhang, Y.; You, M.H. Development of polymorphic microsatellite markers and their use in collections of Erianthus arundinaceus in China. Grassland Sci. 2017, 63, 54–60. [Google Scholar] [CrossRef]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.L.; Botha, F.C. Preliminary Analysis of Expressed Sequence Tags for Sugarcane. Crop Sci. 2000, 40, 1769–1779. [Google Scholar] [CrossRef]
- Carson, D.; Botha, F.C. Genes expressed in sugarcane maturing internodal tissue. Plant Cell Rep. 2002, 20, 1075–1081. [Google Scholar]
- Cato, S.A.; Gardner, R.C.; Kent, J.; Richardson, T.E. A rapid PCR-based method for genetically mapping ESTs. Theor. Appl. Genet. 2001, 102, 196–306. [Google Scholar] [CrossRef]
- Telles, G.P.; Braga, M.D.V.; Dias, Z.; Lin, T.L.; Quitzau, J.A.A.; Silva, F.R.; Meidanis, J. Bioinformatics of the sugarcane EST project. Genet Mol. Biol. 2001, 24, 9–15. [Google Scholar] [CrossRef]
- Vicentini, R.; Bem, L.E.V.; Sluys, M.A.; Nogueira, F.T.S.; Vincentz, M. Gene Content Analysis of Sugarcane Public ESTs Reveals Thousands of Missing Coding-Genes and an Unexpected Pool of Grasses Conserved ncRNAs. Trop Plant Biol. 2012, 5, 199–205. [Google Scholar] [CrossRef]
- Silva, J.A.G. Preliminary analysis of microsatellite markers derived from sugarcane expressed sequence tags (ESTs). Genet Mol. Biol. 2001, 24, 155–159. [Google Scholar] [CrossRef]
- Pan, Y.B.; Cordeiro, G.M.; Richard, E.P., Jr.; Henry, R.J. Molecular genotyping of sugarcane clones with microsatellite DNA markers. Maydica 2003, 48, 319–329. [Google Scholar]
- Aitken, K.S.; Jackson, P.A.; McIntyre, C.L. A combination of AFLP and SSR markers provide extensive map coverage and identification of homo(eo)logous linkage groups in a sugarcane cultivar. Theor. Appl. Genet. 2005, 110, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Shinde, D.; Lai, Y.; Sun, F.; Arnheim, N. Taq DNA polymerase slippage mutation rates measured by PCR and quasi-likelihood analysis: (CA/GT)n and (A/T)n microsatellites. Nucleic Acids Res. 2003, 31, 974–980. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.B. Highly polymorphic microsatellite DNA markers for sugarcane germplasm evaluation and variety identity testing. Sugar Tech. 2006, 8, 246–256. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005, 10, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Brownie, J.; Shawcross, S.; Theaker, J.; Whitcombe, D.; Ferrie, R.; Newton, C.; Little, S. The elimination of primer-dimer accumulation in PCR. Nucleic Acids Res. 1997, 25, 3235–3241. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.R.; Butler, J.M.; Vallone, P.M. A 26plex autosomal STR assay to aid human identity testing. J. Forensic Sci. 2009, 54, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.M. Novel non-templated nucleotide addition-reactions catalyzed by prokaryotic and eukaryotic DNA-polymerases. Nucleic Acids Res. 1988, 16, 9677–9686. [Google Scholar] [CrossRef] [PubMed]
- Esselink, G.D.; Smulders, M.J.M.; Vosman, B. Identification of cut rose (Rosa hybrida) and rootstock varieties using robust sequence tagged microsatellite site markers. Theor. Appl. Genet. 2003, 106, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mo, J.; Xie, T.; Cai, P.; Zou, X. Application of spline wavelet self-convolution in processing capillary electrophoresis overlapped peaks with noise. Anal. Chim. Acta 2001, 437, 151–156. [Google Scholar] [CrossRef]
- Fu, Y.-H.; Pan, Y.-B.; Lei, C.-Y.; Xie, H.-J.; Lei, S.-F.; Lu, J.-J. SSR-CE/FD assessment of Guizhou approved sugarcane cultivars and regional materials. Chin. Agric. Sci. Bull. 2017, 33, 5–12. [Google Scholar]
- Wang, X.W.; Trigiano, R.N.; Windham, M.T.; DeVries, R.E.; Scheffler, B.E.; Rinehart, T.A.; Spiers, J.M. A simple PCR procedure for discovering microsatellites from small insert libraries. Mol. Ecol. Notes 2007, 7, 558–561. [Google Scholar] [CrossRef]
- Andru, S.; Pan, Y.-B.; Thongthawee, S.; Burner, D.M.; Kimbeng, C.A. Genetic analysis of the sugarcane (Saccharum spp.) cultivar ‘LCP 85-384’. I. Linkage mapping using AFLP, SSR, and TRAP markers. Theor. Appl. Genet. 2011, 123, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Singh, R.B.; Singh, S.P.; Sharma, M.L. Identification of sugarcane microsatellites associated to sugar content in sugarcane and transferability to other cereal genomes. Euphytica 2011, 182, 335–354. [Google Scholar] [CrossRef]
- Edmé, S.J.; Glynn, N.G.; Comstock, J.C. Genetic segregation of microsatellite markers in Saccharum officinarum and S. spontaneum. Heredity 2006, 97, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, G.M.; Pan, Y.B.; Henry, R.J. Sugarcane microsatellites for the assessment of genetic diversity in sugarcane germplasm. Plant Sci. 2003, 165, 181–189. [Google Scholar] [CrossRef]
- Joshi, S.V.; Albertse, E.H. Development of a DNA fingerprinting database and cultivar identification in sugarcane using a Genetic analyzer. Proc. S. Afr. Sugarcane Technol. Assoc. 2013, 86, 200–212. [Google Scholar]
- Hameed, U.; Pan, Y.B.; Muhammad, K.; Afghan, S.; Iqbal, J. Use of simple sequence repeat markers for DNA fingerprinting and diversity analysis of sugarcane (Saccharum spp) cultivars resistant and susceptible to red rot. Genet. Mol. Res. 2012, 11, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Pan, Y.B.; Xu, L.P.; Gao, S.W.; Wang, Q.N.; Su, Y.C.; Yang, Y.Q.; Wu, Q.B.; Zhou, D.G.; Que, Y.X. Genetic diversity analysis of sugarcane germplasm based on fluorescence-labeled simple sequence repeat markers and a capillary electrophoresis-based genotyping platform. Sugar Tech. 2016, 18, 380–390. [Google Scholar] [CrossRef]
- Singh, R.K.; Mishra, S.K.; Singh, S.P.; Mishra, N.; Sharma, M.L. Evaluation of microsatellite markers for genetic diversity analysis among sugarcane species and commercial hybrids. Aust. J. Crop Sci. 2010, 4, 116–125. [Google Scholar]
- Fu, Y.; Pan, Y.; Lei, C.; Grisham, M.P.; Yang, C.; Meng, Q. Genotype-Specific Microsatellite (SSR) Markers for the Sugarcane Germplasm from the Karst Region of Guizhou, China. Am. J. Plant Sci. 2016, 7, 2209–2220. [Google Scholar] [CrossRef]
- Santos, J.M.D.; Barbosa, G.V.D.S.; Ramalho Neto, C.E.; Almeida, C. Efficiency of biparental crossing in sugarcane analyzed by SSR markers. Crop Breed. Appl. Biotechnol. 2014, 14, 102–107. [Google Scholar] [CrossRef]
- Liu, P.; Chandra, A.; Que, Y.; Chen, P.H.; Grisham, M.P.; White, W.H.; Dalley, C.D.; Tew, T.L.; Pan, Y.-B. Identification of quantitative trait loci controlling sucrose content based on an enriched genetic linkage map of sugarcane (Saccharum spp. hybrids) cultivar ‘LCP 85-384’. Euphytica 2016, 207, 527–549. [Google Scholar] [CrossRef]
- Siraree, A.; Banerjee, N.; Kumar, S.; Khan, M.S.; Singh, P.K.; Sharma, S.; Singh, R.K.; Singh, J. Identification of marker-trait associations for morphological descriptors and yield component traits in sugarcane. Physiol. Mol. Biol. Plants 2017, 23, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhou, H.; Pan, Y.B.; Chen, C.Y.; Zhu, J.R.; Chen, P.H.; Li, Y.R.; Cai, Q.; Chen, R.K. Segregation analysis of microsatellite (SSR) markers in sugarcane polyploids. Genet. Mol. Res. 2015, 14, 18384–18395. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.B.; Liu, P.; Que, Y. Independently segregating simple sequence repeats (SSR) alleles in polyploid sugarcane. Sugar Tech. 2015, 17, 235–242. [Google Scholar] [CrossRef]
- Gao, Y.J.; Liu, X.H.; Zhang, R.H.; Zhou, H.; Liao, J.X.; Duan, W.X.; Zhang, G.M. Verification of Progeny from Crosses between Sugarcane (Saccharum spp.) and an Intergeneric Hybrid (Erianthus arundinaceus × Saccharum spontaneum) with Molecular Makers. Sugar Tech. 2015, 17, 31–35. [Google Scholar] [CrossRef]
- Liu, X.; Fang, F.; Zhang, R.; Song, H.; Yang, R.; Gao, Y.; Ou, H.; Lei, J.; Luo, T.; Duan, W.; et al. Identification of progenies from sugarcane× Narenga porphyrocoma (Hance) Bor. by SSR marker. Southwest China J. Agric. Sci. 2012, 25, 38–43. [Google Scholar]
- Cai, Q.; Aitken, K.S.; Fan, Y.H.; Piperidis, G.; Jackson, P.; McIntyre, C.L. A preliminary assessment of the genetic relationship between Erianthus Rockii and the ‘‘Saccharum complex’’ using microsatellite (SSR) and AFLP markers. Plant Sci. 2005, 169, 976–984. [Google Scholar] [CrossRef]
- Nayak, S.N.; Song, J.; Villa, A.; Pathak, B.; Ayala-Silva, T.; Yang, X.; Todd, J.; Glynn, N.C.; Kuhn, D.N.; Glaz, B.; et al. Promoting utilization of saccharum spp. genetic resources through genetic diversity analysis and core collection construction. PLoS ONE 2014, 9, e110856. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.D.; Dobhal, U.; Singh, P.; Kumar, S.; Gaur, A.K.; Singh, S.P.; Jeena, A.S.; Koshy, E.P.; Kumar, S. Assessment of genetic diversity among sugarcane cultivars using novel microsatellite markers. Afr. J. Biotechnol. 2014, 8, 1444–1451. [Google Scholar] [CrossRef]
- Tena, E.; Mekbib, F.; Ayana, A. Analysis of genetic diversity and population structure among exotic sugarcane (Saccharum spp.) cultivars in Ethiopia using simple sequence repeats (SSR) molecular markers. Afr. J. Biotechnol. 2014, 13, 4308–4319. [Google Scholar]
- Creste, S.; Sansoli, D.M.; Tardiani, A.C.S.; Silva, D.N.; Goncalves, F.K.; Favero, T.M.; Medeiros, C.N.F.; Festucci, C.S.; Carlini-Garcia, L.A.; Landell, M.G.A.; et al. Comparison of AFLP, TRAP and SSRs in the estimation of genetic relationships in sugarcane. Sugar Tech. 2010, 12, 150–154. [Google Scholar] [CrossRef]
- Gouy, M.; Rousselle, Y.; Chane, A.T.; Anglade, A.; Royaert, S.; Nibouche, S.; Costet, L. Genome wide association mapping of agro-morphological and disease resistance traits in sugarcane. Euphytica 2015, 202, 269. [Google Scholar] [CrossRef]
- Aitken, K.S.; Hermann, S.; Karno, K.; Bonnett, G.D.; McIntyre, L.C.; Jackson, P.A. Genetic control of yield related stalk traits in sugarcane. Theor. Appl. Genet. 2008, 117, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.R.; Garcia, A.A.F.; Pastina, M.M.; Teixeira, L.H.M.; Bressiani, J.A.; Ulian, E.C.; Bidoia, M.A.P.; Souza, A.P. Analysis of genomic and functional RFLP derived markers associated with sucrose content, fiber and yield QTLs in a sugarcane (Saccharum spp.) commercial cross. Euphytica 2010, 172, 313–327. [Google Scholar] [CrossRef]
- Al-Janabi, S.M.; Honeycutt, R.J.; McClelland, M.; Sobral, B.W.S. A genetic linkage map of Saccharum spontaneum L. ‘SES 208’. Genetics 1993, 134, 1249–1260. [Google Scholar] [PubMed]
- da Silva, J.A.G.; Sorrells, M.E.; Burnquist, W.; Tanksley, S.D. RFLP linkage map and genome analysis of Saccharum spontaneum. Genome 1993, 36, 782–791. [Google Scholar] [CrossRef]
- Ming, R.; Lin, S.C.; Lin, Y.R.; da Silva, J.; Wilson, W. Detailed alignment of Saccharum and Sorghum chromosomes: Comparative organization of closely related diploid and polyploid genomes. Genetics 1998, 150, 1663–1682. [Google Scholar] [PubMed]
- Guimaraes, C.T.; Sills, G.R.; Sobral, B.W.S. Comparative mapping of Andropogoneae: Saccharum L. (sugarcane) and its relation to sorghum and maize. Proc. Natl. Acad. Sci. USA 1997, 94, 14262–14266. [Google Scholar] [CrossRef]
- Aitken, K.S.; McNeil, M.D.; Hermann, S.; Bundock, P.C.; Kilian, A.; Heller-Uszynska, K.; Henry, R.J.; Li, J. A comprehensive genetic map of sugarcane that provides enhanced map coverage and integrates high-throughput Diversity Array Technology (DArT) markers. BMC Genomics 2014, 15, 152. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.A.F.; Kido, E.A.; Meza, A.N.; Souza, H.M.B.; Pinto, L.R.; Pastina, M.M.; Leite, C.S.; da Silva, J.A.G.; Ulian, E.C.; Figueira, A.V.O.; et al. Development of an integrated genetic map of a sugarcane (Saccharum spp.) commercial cross based on a maximum-likelihood approach for estimation of linkage and linkage phases. Theor. Appl. Genet. 2006, 112, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Alwala, S.; Kimbeng, C.A.; Veremis, J.C.; Gravois, K.A. Linkage mapping and genome analysis in Saccharum interspecific cross using AFLP, SRAP and TRAP markers. Euphytica 2008, 164, 37–51. [Google Scholar] [CrossRef]
- Pastina, M.M.; Malosetti, M.; Gazaffi, R.; Mollinari, M.; Margarido, G.R.A.; Oliveira, K.M.; Pinto, L.R.; Souza, A.P.; van Eeuwijk, F.A.; Garcia, A.A.F. A mixed model QTL analysis for sugarcane multiple-harvest-location trial data. Theor. Appl. Genet. 2012, 124, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, C.T.; Honeycutt, R.J.; Sills, G.R.; Sobral, B.W.S. Genetic linkage maps of Saccharum officinarum L. and Saccharum robustum Brandes & Jew. Ex Grassl. Genet. Mol. Biol. 1999, 22, 125–132. [Google Scholar]
- Ming, R.; Wang, Y.W.; Draye, X.; Moore, P.H.; Irvine, J.E.; Paterson, A.H. Molecular dissection of complex traits in autopolyploids: Mapping QTLs affecting sugar yield and related traits in sugarcane. Theor. Appl. Genet. 2002, 105, 332–345. [Google Scholar] [PubMed]
- Aitken, K.S.; Jackson, P.A.; McIntyre, C.L. QTL identified for sugar related traits in a sugarcane (Saccharum spp.) cultivar 9 S. officinarum population. Theor. Appl. Genet. 2007, 112, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, K.M.; Pinto, L.R.; Marconi, T.G.; Margarido, G.R.A.; Pastina, M.M.; Teixeira, L.H.M.; Figueira, A.V.; Ulian, E.C.; Garcia, A.A.F.; Souza, A.P. Functional integrated genetic linkage map based on EST markers for a sugarcane (Saccharum spp.) commercial cross. Mol. Breed. 2007, 20, 189–208. [Google Scholar] [CrossRef]
- Singh, R.K.; Banerjee, N.; Khan, M.S.; Yadav, S.; Kumar, S.; Duttamajumder, S.K.; Lal, R.J.; Patel, J.D.; Guo, H.; Zhang, D.; et al. Identification of putative candidate genes for red rot resistance in sugarcane (Saccharum species hybrid) using LD-based association mapping. Mol. Genet. Genom. 2016, 291, 1363–1377. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.A.; Anoni, C.O.; Mancini, M.C.; Santos, F.R.C.; Marconi, T.G.; Gazaffi, R.; Pastina, M.M.; Perecin, D.; Mollinari, M.; Xavier, M.A.; et al. QTL mapping including co-dominant SNP markers with ploidy level information in a sugarcane progeny. Euphytica 2016, 211, 1–16. [Google Scholar] [CrossRef]
- Ming, R.; Liu, S.C.; Moore, P.H.; Irvine, J.E.; Paterson, A.H. QTL analysis in a complex autopolyploid: Genetic control of sugar content in sugarcane. Genome Res. 2001, 11, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Hoarau, J.Y.; Grivet, L.; Offmann, B.; Raboin, L.M.; Diorflar, J.P.; Payet, J.; Hellmann, M.; D’Hont, A.; Glaszmann, J.C. Genetic dissection of a modern sugarcane cultivar (Saccharum spp.). II. Detection of QTLs for yield components. Theor. Appl. Genet. 2002, 105, 1027–1037. [Google Scholar] [PubMed]
- Nunes, C.D.M.C. Mapeamento de QTL em Cana-de-Açúcar (Saccharum spp.) Utilizando Marcadores DArt (Diversity Arrays Technology) e Microssatélites; 191 f. Tese (Doutorado em Agronomia); Universidade Federal de Goiás: Goiânia, Brazil, 2013. [Google Scholar]
- Gutierrez, A.; Hoy, J.; Kimbeng, C.; Baisakh, N. Identification of Genomic Regions Controlling Leaf Scald Resistance in Sugarcane Using a Bi-parental Mapping Population and Selective Genotyping by Sequencing. Front. Plant Sci. 2018, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Tew, T.L.; Pan, Y.B. Microsatellite (SSR) Marker-Based Paternity Analysis of a Seven-Parent Poly crosses in Sugarcane. Crop Sci. 2010, 50, 1401–1408. [Google Scholar] [CrossRef]
- Berding, N.; Hogarth, D.M.; Cox, M.C. Plant Improvement of Sugarcane; Glyn, J., Ed.; Blackwell Science Ltd.: Oxford, UK, 2004; pp. 20–53. [Google Scholar]
- Xavier, M.A.; Pinto, L.R.; Fávero, T.M.; Perecin, D.; Carlini-Garcia, L.A.; Landell, M.G.A. Paternity identification in sugarcane polycrosses by using microsatellite markers. Genet. Mol. Res. 2014, 13, 2268–2277. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.H.; Pan, Y.B.; Chen, R.K.; Xu, L.P.; Chen, Y.Q. SSR marker based analysis of genetic relatedness among sugarcane cultivars (Saccharum spp. hybrids) from breeding programs in China and other countries. Sugar Tech. 2009, 11, 347–354. [Google Scholar] [CrossRef]
- Gupta, V.; Raghuvanshi, S.; Gupta, A.; Saini, N.; Gaur, A.; Khan, M.S.; Gupta, R.S.; Singh, J.; Duttamajumder, S.K.; Srivastava, S.; et al. The water-deficit stress-and red-rot-related genes in sugarcane. Funct. Integr. Genom. 2010, 10, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Selvi, A.; Nair, N.V.; Balasundaram, N.; Mohapatra, T. Evaluation of maize micro satellite markers for genetic diversity analysis and fingerprinting in sugarcane. Genome 2003, 46, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Dillon, S.L.; Shapter, F.M.; Henry, R.J.; Cordeiro, G.; Izquierdo, L.; Slade Lee, L. Domestication to crop improvement: Genetic resources for Sorghum and Saccharum (Andropogoneae). Ann. Bot. 2007, 100, 975–989. [Google Scholar] [CrossRef] [PubMed]
- Alwala, S.; Kimbeng, C.A.; Veremis, J.C.; Gravois, K.A. Identification of molecular markers associated with sugar related traits in a Saccharum inter-specific cross. Euphytica 2009, 167, 127–142. [Google Scholar] [CrossRef]
- Bremer, G. Problems in breeding and cytology of sugar cane. Euphytica 1961, 10, 59–78. [Google Scholar] [CrossRef]
- Tew, T.L.; Pan, Y.B. Molecular assessment of the fidelity of sugarcane crosses with high-throughput microsatellite genotyping. J. Am. Soc. Sugar Cane Technol. 2005, 25, 119. [Google Scholar]
- D’Hont, A.; Rao, P.S.; Feldmann, P.; Grivet, L.; Islam-Faridi, N.; Taylor, P.; Glaszmann, J.C. Identification and characterisation of sugarcane intergeneric hybrids, Saccharum officinarum × Erianthus arundinaceus, with molecular markers and DNA in situ hybridisation. Theor. Appl. Genet. 1995, 91, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Aitken, K.S.; Deng, H.H.; Chen, X.W.; Fu, C.; Jackson, P.; McIntyre, C.L. Verification of the introgression of Erianthus arundinaceus germplasm into sugarcane using molecular markers. Plant Breed. 2005, 124, 322–328. [Google Scholar] [CrossRef]
- Tao, L.A.; Chu, L.B.; Jing, Y.F.; Liu, X.L.; An, R.D.; Dong, L.H.; Yang, L.H.; Zhou, Q.M.; Duan, H.F. Identification of genuine hybrids from the cross of chewing cane and Saccharum spontaneum in Yunnan by SSR Markers. J. Plant Genet. Res. 2009, 10, 132–135. [Google Scholar]
- Alix, K.; Paulet, F.; Glaszmann, J.C.; D’Hont, A. Inter-Alu like species-specific sequences in the Saccharum complex. Theor. Appl. Genet. 1992, 99, 962–968. [Google Scholar] [CrossRef]
- Piperidis, G.; Christopher, M.J.; Carroll, B.J.; Berding, N.; D’Hont, A. Molecular contribution to selection of intergeneric hybrids between sugarcane and the wild species Erianthus arundinaceus. Genome 2000, 43, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Fang, F.X.; Gao, Y.J.; Zhang, R.H.; Song, H.Z.; Yang, R.Z.; Fang, W.K.; Duan, W.X.; Luo, T.; Zhang, G.M.; et al. Identification and genetic analysis of hybrid from cross between Erianthus arundinacius (Retz.) Jesws. and Saccharum spontaneum L. Acta Agron. Sin. 2012, 38, 914–920. [Google Scholar] [CrossRef]



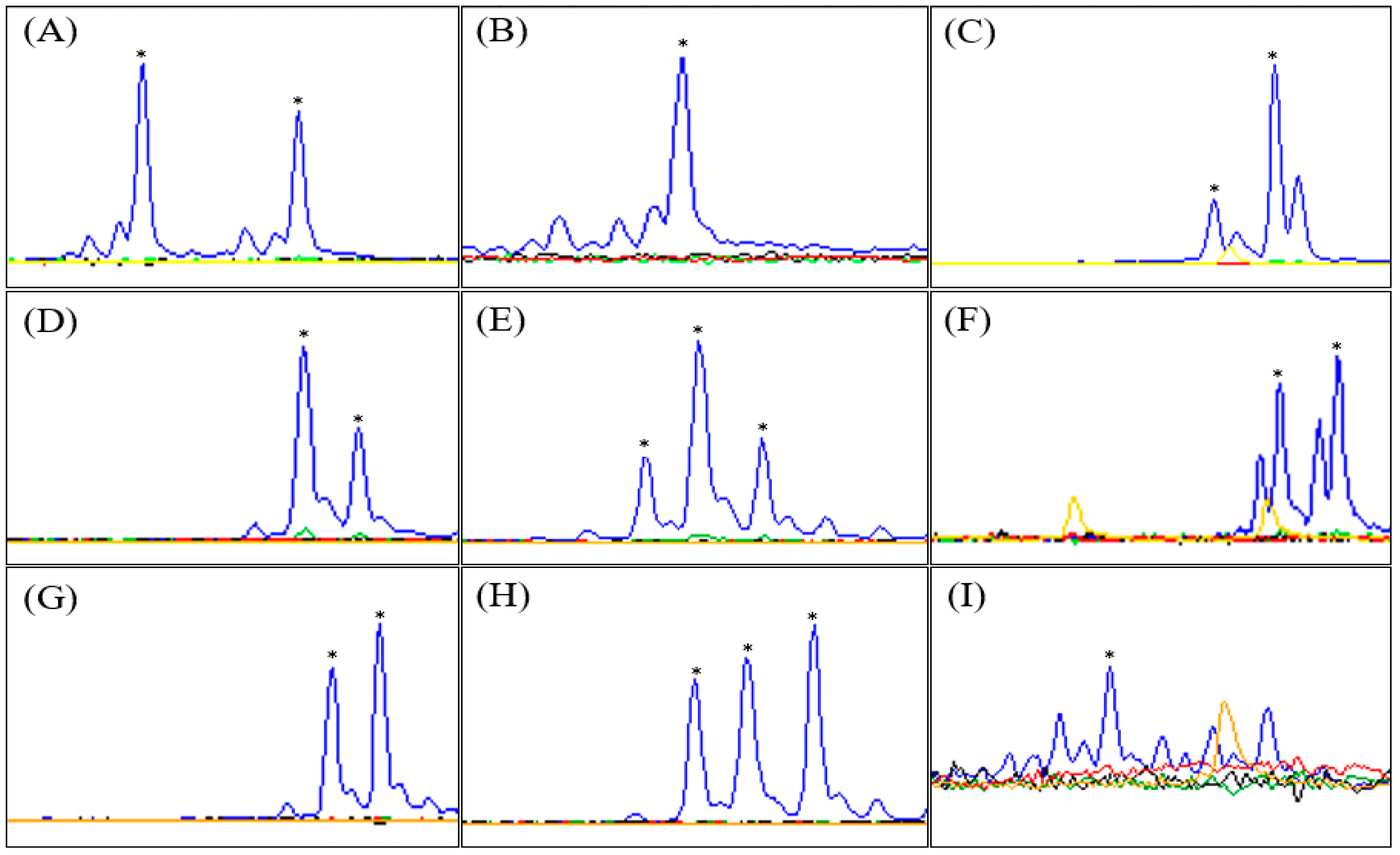{kind=link}
{kind=link}
| Classification | Motif |
|---|---|
| (A) Based on the number of nucleotides per repeats: | |
| Mono (A)10 | A |
| Di (CT)5 | CTCTCTCTCT |
| Tri (CTG)3 | CTGCTGCTG |
| Tetra (CAGA)4 | CAGACAGACAGACAGA |
| Penta (AAATT)5 | AAATTAAATTAAATT |
| Hexa (CTTTAA)6 | CTTTAACTTTAACTTTAA |
| (B) Based on the arrangement of nucleotides in the repeat motifs: | |
| Perfect repeat | CTCTCTCTCTCT |
| Compound repeat | CTCTCTCACACA |
| Imperfect repeat | CTCTCTACTCTCT |
| Region of cryptic simplicity | GTGTACACAGT |
| (C) Based on the location of SSRs in the genome: | |
| Nuclear (nuSSRs) | |
| Chloroplast (cpSSRs) | |
| Mitochondrial (mtSSRs) | |
| Method | Description | Reference |
|---|---|---|
| ESTs libraries | Development of 51 EST-SSRs from SUCEST database for the comparison of EST-SSRs with genomic SSR’s to establish relationship among 18 sugarcane clones. | [21] |
| ESTs libraries | Development of 351 EST-SSRs from the 4085 non redundant EST sequences of two Indian sugarcane cultivars, out of which 227 were evaluated in sugarcane. | [27] |
| ESTs libraries | Oliveira designed 342 EST-SSR primer pairs, of which 224 amplified polymorphic bands in 18 sugarcane varieties. | [22] |
| ESTs libraries | A total of 2335 EST’s functional microsatellite marker developed from the sugarcane sequence tag database (SUCEST) using bioinformatics tools. | [28] |
| ESTs libraries | Development of 722 EST-SSRs from the 8760 sucrose related EST sequences harvested from NCBI database and their application in sugarcane molecular breeding. | [29] |
| ESTs libraries | Development of 267 EST-SSRs markers through computational approach from 10,000 ESTs sequences for genetic diversity, cross-species transferability among Poaceae plants and bulk segregation analysis. | [30] |
| Enriched-genomic libraries | Development of 21 microsatellite markers from sugarcane (Saccharum spp.) ESTs and tested on 5 sugarcane clones and cross transferable to Erianthus and sorghum. | [24] |
| Enriched-genomic libraries | Development of informative genomic microsatellite markers from two hybrid sugarcane cultivars enriched with 18 different repeat-motifs for efficient genotyping applications in sugarcane. | [31] |
| Enriched-genomic libraries | Development of a total of 5675 microsatellite markers in autopolyploid sugarcane and comparative analysis of conserved microsatellites in sorghum and sugarcane | [32] |
| Enriched-genomic libraries | Development of 26 SSR’s from a sugarcane interspecific hybrid of ISH 100 to study the population genetics structure across the species. SSR’s primer pairs were assessed on 8 individuals sampled from one population. | [33] |
| Next-generation sequencing | A total of 1682 candidate loci were used to developed 174 primer pairs and validate on eight Erianthus arundinaceus accessions. | [34] |
| Next-generation sequencing | Illumina RNA-Seq platform were used on the de novo assembly of the sugarcane transcriptome, and a large number of molecular markers were found, including 5106 SSRs and 708,125 SNPs. | [35] |
| Next-generation sequencing | Development of 15 polymorphic SSR markers using 3730 XL Automated DNA Sequencer for 164 individuals of 18 populations of Chinese wild E. arundinaceus. | [36] |
| EST-SSRs | Genic-SSRs | AFLPs | RFLPs | RAPDs/ISSRs | SNP | |
|---|---|---|---|---|---|---|
| Classification | PCR-Based | PCR-Based | PCR-Based | Hybridization based | PCR-Based | Sequence Based |
| Degree of polymorphism | Low | High | Low-Moderate | Low | Low-Moderate | High |
| Dominance | Co-dominant | Co-dominant | Dominant | Co-dominant | Dominant | Co-dominant |
| Interspecific transferability | High | Moderate | Low-Moderate | Moderate-High | Low-Moderate | High |
| Utility among commonly used markers | High | High | Low-Moderate | Moderate | Low-Moderate | Moderate-High |
| Production cost and labor involved | Low | High | Low-Moderate | High | Low-Moderate | Moderate |
| Application | Phylogeny, Gene mapping, Fingerprinting, Genetic diversity | Disease resistance, Genetic diversity, Starch contents, DNA fingerprinting | Genetic mapping and QTL linkage, QTL mapping, Paternity tests | Fingerprinting Genome mapping, Disease analysis, Genetic diversity, | Phylogeny, Genetic identity, Parentage, Clone strains identification, Gene mapping | Haplotype mapping, Linkage Disequilibrium, Disease and Trait association |
| Usage in plant species | Sugarcane, Rice, Maize, Tomato, Apple, Cereals | Rice, Sugarcane, Maize, Pineapple, Fruits crops | Palm, Sugarcane, Wheat, Rice, Fruits Crops | Wheat, Melon, Sugarcane, Barley, Potato | Broccoli, Mulberry, Cherry, Cucurbits, Sugarcane, Radish | Rice, Maize, Wheat, Sugarcane, Brassica rapa |
| Application | Description | Reference |
|---|---|---|
| Cultivar identification | A total of 84 clones were identified, including 58 commercial varieties, 17 clones in the final phase of the SASRI selection program for release and 9 imported varieties, using four SSR markers. | [60] |
| Cross-species transferability | Validation and polymorphism study of 227 EST-SSRs with 124 important Indian sugarcane cultivars for cross-species transferability and their utility in revealing population structure and genetic diversity. | [27] |
| Fingerprinting and genetic diversity | Establishment of the molecular identities (ID) of 91 nationally or provincially released Chinese sugarcane varieties and to evaluate the extent of genetic diversity among these varieties using SSR DNA markers and two fingerprinting systems. | [12] |
| Fingerprinting and genetic diversity | Use of 21 simple sequence repeat (SSR) markers for DNA fingerprinting and diversity analysis on 20 sugarcane (Saccharum spp) cultivars resistant and susceptible to red rot. | [61] |
| Genetic diversity | Fluorescence-labeled seven gSSR and eight EST-SSR markers were used for the genetic diversity among 181 sugarcane genotypes. | [62] |
| Genetic diversity | A total of 84 genotypes from the Saccharum barberi, S. spontaneum, S. officinarum, Indian and non-Indian commercial cultivars were evaluated for the genetic diversity using seven sugarcane cDNA derived SSRs, nine gSSRs and 16 uni-gene SSRs markers. | [63] |
| Genetic variability | Eighteen sugarcane genotypes including 13 active cultivars and five elite QT-series clones bred locally were screened for genetic variability with 21 SSR primer pairs. | [64] |
| Genome mapping | Genetic variability of 42 E. arundinaceus accessions native to Japan based on nuclear DNA content and 31 simple sequence repeat (SSR) markers | [34] |
| Genome mapping | Low cost single strand conformation polymorphism of 16 genomic and 12 EST-SSRs marker and its utility in genetic evaluation of 22 sugarcane genotypes. | [28] |
| Paternity analysis | Paternity analysis of 76 sugarcane progenies from four crosses was analyzed using three highly polymorphic SSR markers. | [65] |
| QTL mapping | Identification of quantitative trait loci controlling sucrose content based on an enriched genetic linkage map of sugarcane (Saccharum spp. hybrids) cultivar ‘LCP 85–384 using 65 microsatellites markers. | [66] |
| QTL mapping | Identification of marker-trait associations for morphological descriptors and yield component traits in sugarcane using 174 SSRs in a panel of 92 sugarcane varieties from sub-tropical India. | [67] |
| Segregation analysis | Investigation of SSR marker segregation among 964 single pollens and 288 self-progenies (S1) of sugarcane cultivar LCP 85–384. | [68] |
| Segregation analysis | Segregation of a multiallelic sugarcane SSR marker SMC336BS among single pollens of a sugarcane cultivar L99–233 as well as its F1 progenies of a bi-parental cross between HoCP 00–950 (female) and L 99–233 (male). | [69] |
| SSR-assisted identification | Identification of the crosses of sugarcane (Saccharum spp.) × an intergeneric hybrid (Erianthus arundinaceus × Saccharum spontaneum) and their progenies by SSR and sequence-related amplified polymorphism (SRAP) molecular markers. | [70] |
| SSR-assisted identification | Identification of 57 progenies of sugarcane × Narenga porphyrocoma (Hance) Bor. by nine pairs of SSR primers. | [71] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Wang, J.-D.; Pan, Y.-B.; Sharif, R.; Gao, S.-J. Development and Use of Simple Sequence Repeats (SSRs) Markers for Sugarcane Breeding and Genetic Studies. Agronomy 2018, 8, 260. https://doi.org/10.3390/agronomy8110260
Ahmad A, Wang J-D, Pan Y-B, Sharif R, Gao S-J. Development and Use of Simple Sequence Repeats (SSRs) Markers for Sugarcane Breeding and Genetic Studies. Agronomy. 2018; 8(11):260. https://doi.org/10.3390/agronomy8110260
Chicago/Turabian StyleAhmad, Ali, Jin-Da Wang, Yong-Bao Pan, Rahat Sharif, and San-Ji Gao. 2018. "Development and Use of Simple Sequence Repeats (SSRs) Markers for Sugarcane Breeding and Genetic Studies" Agronomy 8, no. 11: 260. https://doi.org/10.3390/agronomy8110260
APA StyleAhmad, A., Wang, J.-D., Pan, Y.-B., Sharif, R., & Gao, S.-J. (2018). Development and Use of Simple Sequence Repeats (SSRs) Markers for Sugarcane Breeding and Genetic Studies. Agronomy, 8(11), 260. https://doi.org/10.3390/agronomy8110260