Breeding for Quality Protein Maize (QPM) Varieties: A Review

 ,
,
Abstract
:1. Introduction
2. Nutritional Quality and Impact of QPM
3. Breeding Approaches
4. Conventional Breeding Approaches
4.1. Screening of QPM Genotypes for Abiotic Stresses
4.2. Screening of QPM Genotypes for Tolerance to Biotic Stresses
5. Combining Ability Studies of QPM Genotypes Traits for Tolerance to Biotic and Abiotic Stresses
6. Molecular Approaches
7. QPM Varieties Released in Some Countries around the World
8. Adoption of QPM and Ways to Enhance Adoption
9. Isolation Requirement for QPM Production
10. Participatory Plant Breeding and Variety Selection
11. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prasanna, B.M.; Vasal, S.K.; Kassahun, B.; Singh, N.N. Quality protein maize. Curr. Sci. 2001, 81, 1308–1319. [Google Scholar]
- Vasal, S.K. Quality protein maize development: An exciting experience. In Integrated Approaches to Higher Maize Productivity in the New Millennium, Proceedings of the Seventh Eastern and Southern Africa Regional Maize Conference, Nairobi, Kenya, 5–11 February 2002; Friesen, D., Palmer, A.F.E., Eds.; CIMMYT (International Maize and Wheat Improvement Center) and KARI (Kenya Agricultural Research Institute): Nairobi, Kenya, 2002; pp. 2–9. [Google Scholar]
- Mbuya, K.; Nkongolo, K.K.; Kalonji-Mbuyi, A. Nutritional analysis of quality protein maize varieties selected for agronomic characteristics in a breeding program. Int. J. Plant Breed. Genet. 2011, 5, 317–327. [Google Scholar]
- Sofi, P.A.; Wani, A.S.; Rather, A.G.; Wani, H.S. Review article: Quality protein maize (QPM): Genetic manipulation for the nutritional fortification of maize. J. Plant Breed. Crop Sci. 2009, 1, 244–253. [Google Scholar]
- Krivanek, A.F.; Groote, H.D.; Gunaratna, N.S.; Diallo, A.O.; Friesen, D. Breeding and disseminating quality protein maize (QPM) for Africa. Afr. J. Biotechnol. 2007, 6, 312–324. [Google Scholar]
- Müller, O.; Krawinkel, M. Malnutrition and health in developing countries. Can. Med. Assoc. J. 2005, 173, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Dudley, J.W.; Lambert, R.J. Genetic variability after 65 generations of selection in Illinois high oil, low oil, high protein and low protein strains of Zea mays L. Crop Sci. 1969, 9, 179–181. [Google Scholar] [CrossRef]
- Bjarnason, M.; Vasal, S.K. Breeding for quality protein maize. Plant Breed. Rev. 1992, 9, 181–216. [Google Scholar]
- Zaidi, P.H.; Vasal, S.K.; Maniselvan, P.; Jha, G.C.; Mehrajjudin; Singh, R.P. Stability in performance of quality protein maize under abiotic stress. Maydica 2008, 53, 249–260. [Google Scholar]
- Machida, L.; Derera, J.; Tongoona, P.; Mutanga, O.; Macrobert, J. Geostatistical analysis of quality protein maize outcrossing with pollen from adjacent normal endosperm maize varieties. Crop Sci. Soc. Am. 2012, 52, 1235–1245. [Google Scholar] [CrossRef]
- Qi, G.; Diao, Q.; Tu, Y.; Wu, S.; Zhang, S. Nutritional evaluation of quality protein maize (QPM) in animal feed. In Protein Source for Animal Feed Industry: Expert Workshop and Consultation, Bangkok, 29 April–3 May 2002; FAO Animal Production and Health Proceedings; FAO: Rome, Italy, 2004; pp. 185–198. [Google Scholar]
- Vivek, B.S.; Krivanek, A.F.; Palacios-Rojas, N.; Twumasi-Afriyie, S.; Diallo, A.O. Breeding Quality Protein Maize (QPM): Protocols for Developing QPM Cultivars; Mexico, D.F., Ed.; CIMMYT: Texcoco, Mexico, 2008; p. 5. [Google Scholar]
- Tripathy, S.K.; Ithape, D.M.; Maharana, M.; Prusty, A.M. Quality Protein Maize (QPM): Genetic basis and breeding perspective. Trop. Plant Res. 2017, 4, 145–152. [Google Scholar] [CrossRef]
- Pixley, K.V. The Development and Promotion of Quality Protein Maize in Sub-Saharan Africa; Progress Report; CIMMYT: Harare, Zimbabwe, 2003; 65p. [Google Scholar]
- Teklewold, A.; Wegary, D.; Tadesse, A.; Tadesse, B.; Bantte, K.; Friesen, D.; Prasanna, B.M. Quality Protein Maize (QPM): A Guide to the Technology and Its Promotion in Ethiopia; CIMMYT: Addis Ababa, Ethiopia, 2015; p. 440. [Google Scholar]
- Pixley, K.V.; Bjarnason, M.S. Stability of grain yield, endosperm modification and protein quality of hybrid and open-pollinated Quality Protein Maize (QPM) cultivars. Crop Sci. 2002, 42, 1882–1890. [Google Scholar] [CrossRef]
- Bello, O.B.; Ganiyu, O.T.; Wahab, M.K.A.; Azeez, M.A.; Abdulmaliq, S.Y.; Ige, S.A.; Mahmood, J.; Oluleye, F.; Afolabi, M.S. Yield and disease reactions of quality protein maize varieties in the Southern Guinea Savanna agro-ecology of Nigeria. Int. J. Agric. For. 2012, 2, 203–209. [Google Scholar]
- Pfunde, C.N.; Mutengwa, C.S. Combining ability of quality protein maize inbred lines for seedling tolerance to drought stress. Philipp. J. Crop Sci. 2016, 41, 1–12. [Google Scholar]
- Bisen, P.; Dadheech, A.; Namrata, N.O.; Meena, R.K. Exploitation of heterosis in single hybrids of quality protein maize (Zea may L.) for yield and quality traits. Int. J. Bio-Resour. Stress Manag. 2017, 8, 12–19. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Annor, B.; Oyekunle, M.; Akinwale, R.O.; Fakorede, M.A.B.; Talabi, A.O.; Akaogu, I.C.; Melaku, G.; Fasanmade, Y. Grouping of early maturing quality protein maize inbreds based on SNP markers and combining ability under multiple environments. Field Crops Res. 2015, 183, 169–183. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Talabi, A.O.; Oyekunle, M.; Akaogu, I.C.; Akinwale, R.O.; Annor, B.; Melaku, G.; Fasanmade, Y.; Aderounmu, M. Gene action and heterotic groups of early white quality protein maize inbreds under multiple stress environments. Crop Sci. 2016, 56, 183–199. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fontem, L.A. The pattern of grain yield response of normal and quality protein maize cultivars in stress and nonstress environments. Agron. J. 2010, 102, 381–394. [Google Scholar] [CrossRef]
- Musila, N.R.; Diallo, O.A.; Makumbi, D.; Njoroge, K. Combining ability of early-maturing quality protein maize inbred lines adapted to Eastern Africa. Field Crops Res. 2010, 119, 231–237. [Google Scholar] [CrossRef]
- Parihar, A.K.; Godawat, S.L.; Singh, D.; Parihar, C.M.; Jat, M.L. Behaviour of quality protein maize (QPM) genotypes under well irrigated and water stress conditions in subtropical climate. Maydica 2012, 57, 293–299. [Google Scholar]
- Pfunde, C.N. Parent Characterization of Quality Protein Maize (Zea Mays L.) and Combining Ability for Tolerance to Drought Stress. Master’s Thesis, University of Fort Hare, Alice, South Africa, 2012. [Google Scholar]
- Annor, B.; Badu-Apraku, B. Gene action controlling grain yield and other agronomic traits of extra-early quality protein maize under stress and non-stress conditions. Euphytica 2016, 212, 213–228. [Google Scholar] [CrossRef]
- Chiuta, N.E. Breeding Investigations into Heat and Drought Stress Tolerance of Yellow Quality Protein Maize (QPM) Inbred Lines. Master’s Thesis, University of Fort Hare, Alice, South Africa, 2016. [Google Scholar]
- Pfunde, C.N. Investigation into the Genetic Diversity, Physiology, Proteomics and Combining Ability of Quality Protein Maize Inbred Lines under Drought and Heat Stress. Ph.D. Thesis, University of Fort Hare, Alice, South Africa, 2016. [Google Scholar]
- Mertz, E.T.; Bates, L.S.; Nelson, O.E. Mutant gene that changes protein composition and increases lysine content of maize endosperm. Science 1964, 145, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Paes, M.C.D.; Bicudo, M.H. Nutritional perspectives of quality protein maize. In Quality Protein Maize: 1964–1994, Proceedings of the International Symposium on Quality Protein Maize, Sete Lagoas, MG, Brazil, 1–3 December 1994; Purdue University Press: West Lafayette, IN, USA, 1997. [Google Scholar]
- Vasal, S.K. Quality Protein Maize Story, Improving Human Nutrition through Agriculture: The Role of International Agricultural Research; A Workshop Hosted by the International Rice Research Institute, Los Banos, Philippines; International Food Policy Research Institute: Washington, DC, USA, 1999; Volume 16. [Google Scholar]
- Vietmeyer, N.D. A drama in three long acts: The story behind the story of the development of quality-protein maize. Diversity 2000, 16, 29–32. [Google Scholar]
- Ignjatovic-Micic, D.; Kostadinovic, M.; Stankovic, G.; Markovic, K.; Vancetovic, J.; Bozinovic, S.; Andjelkovic, V. Biochemical and agronomic performance of quality protein maize hybrids adapted to temperate regions. Maydica 2013, 58, 311–317. [Google Scholar]
- Twumasi-Afriye, S.; Palacios Rojas, N.; Friesen, D.; Teklewold, A.; Gissa, D.W.; De Groote, H.; Prasanna, B.M. Guidelines for the Quality Control of Quality Protein Maize (QPM) Seed and Grain; CGIAR, CIMMYT: Addis Ababa, Ethiopia, 2016; p. 38. [Google Scholar]
- De Groote, H.; Gunaratna, N.; Ergano, K.; Friesen, D. Extension and adoption of biofortified crop: Quality protein maize in East Africa. In Proceedings of the African Agricultural Economics Association Meetings, Cape Town, South Africa, 19–23 September 2010. [Google Scholar]
- Bressani, R. Protein quality of high-lysine maize for humans. Cereal Foods World 1991, 36, 806–811. [Google Scholar]
- Akuamoa-Boateng, A. Quality Protein Maize Infant Feeding Trials in Ghana; Ghana Health Service Report; Ghana Health Service: Ashanti, Ghana, 2002.
- Akalu, G.; Taffesse, S.; Gunaratna, N.S.; De Groote, H. The effectiveness of quality protein maize in improving the nutritional status of young children in the Ethiopian Highlands. Food Nutr. Bull. 2010, 31, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Gunaratna, N.S.; De Groote, H.; Nestel, P.; Pixley, K.V.; Mccabe, G.P. A meta-analysis of community-based studies on quality protein maize. Food Policy 2010, 35, 202–210. [Google Scholar] [CrossRef]
- Nuss, E.T.; Tanumihardjo, S.A. Quality protein maize for Africa: Closing the protein inadequacy gap in vulnerable populations. Adv. Nutr. 2011, 2, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Mpofu, I.D.T.; Sibanda, S.; Shonihwa, A.; Pixley, K. The nutritional value of quality protein maize for weaner pigs. J. Pet. Environ. Biotechnol. 2012, 3, 129. [Google Scholar]
- Emerson, R.A.; Beadle, G.W.; Fraser, A.C. A Summary of Linkage Studies in Maize; Cornell University Agricultural Experiment Station Memoirs: Ithaca, NY, USA, 1935; Volume 180, pp. 1–80. [Google Scholar]
- Nelson, O.E.; Mertz, E.T.; Bate, L.S. Second mutation gene affecting amino acid pattern of maize endosperm proteins. Science 1965, 150, 1469–1470. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. Improving the protein content and composition of cereal grain. J. Cereal Sci. 2007, 46, 239–250. [Google Scholar] [CrossRef]
- Menkir, A.; Liu, W.; White, W.S.; Maziya-Dixon, B.; Rocheford, T. Carotenoid diversity in tropical-adapted yellow maize inbred lines. Food Chem. 2008, 109, 521–529. [Google Scholar] [CrossRef]
- Pillay, K.; Derera, J.; Siwela, M.; Veldman, F.J. Consumer acceptance of yellow provitamin a-biofortified maize in Kwazulu-Natal. S. Afr. J. Clin. Nutr. 2011, 24, 186–191. [Google Scholar] [CrossRef]
- Vasal, S.K. The quality protein maize story. Food Nutr. Bull. 2000, 21, 445–450. [Google Scholar] [CrossRef]
- Villegas, E.; Vasal, S.K.; Bjarnason, M. Quality protein maize—What is it and how was it developed? In Quality Protein Maize; Mertz, E.T., Ed.; American Association of Cereal Chemists: Eagan, MN, USA, 1992; pp. 27–48. [Google Scholar]
- Nepir, G.; Wegary, D.; Zeleke, H. Heterosis and combining ability of highland quality protein maize inbred lines. Maydica 2015, 60, 1–12. [Google Scholar]
- Ababulgu, D.A. Genetic Diversity and Combining Ability of Selected Quality Protein Maize (QPM) Inbred Lines Adapted to the Highland Agro-Ecology of Ethiopia. Ph.D. Thesis, University of KwaZulu-Natal, Durban, South Africa, 2014. [Google Scholar]
- Henry, A.M.; Manicacci, D.; Falque, M.; Damerval, C. Molecular evolution of the opaque-2 gene in Zea mays L. J. Mol. Evol. 2005, 61, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Woo, Y.; Kim, C.; Larkins, B. Quantitative trait locus mapping of loci influencing elongation factor 1 alpha content in maize endosperm. Plant Physiol. 2001, 125, 1271–1282. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Lou, X.; Ma, C.; Wang, X.; Larkins, B.; Casella, G. An improved genetic model generates high resolution mapping of qtl for protein quality in maize. Proc. Natl. Acad. Sci. USA 2002, 99, 11281–11286. [Google Scholar] [CrossRef] [PubMed]
- Gibbon, C.B.; Larkins, A.B. Molecular genetic approaches to developing quality protein maize. Trends Genet. 2005, 21, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Nkongolo, K.K.; Daniel, G.; Mbuya, K.; Michael, P.; Theriault, G. Characterization of new quality protein maize (QPM) varieties from a breeding program: Analysis of amino acid profiles and development of a variety-diagnostic molecular marker. Br. Biotechnol. J. 2015, 6, 101–112. [Google Scholar] [CrossRef]
- Nkongolo, K.K.; Mbuya, K.; Mehes-Smith, M.; Kalonji-Mbuyi, A. Molecular analysis of quality protein (QPM) and normal maize varieties from the DR-Congo breeding program. Afr. J. Biotechnol. 2011, 10, 14293–14301. [Google Scholar]
- Manna, R.; Okello, D.K.; Imanywoha, J.; Pixley, K.V.; Edema, R. Enhancing introgression of the opaque-2 trait into elite maize lines using simple sequence repeats. Afr. Crop Sci. J. 2005, 13, 215–226. [Google Scholar]
- Badu, R.; Prasanna, B.M. Molecular breeding for Quality Protein Maize (QPM). In Genomics of Plant Genetic Resources; Tuberosa, R., Graner, A., Frison, E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 489–505. [Google Scholar]
- Zhang, Z.; Yang, J.; Wu, Y. Transcriptional regulation of zein gene expression in maize through the additive and synergistic action of opaque-2, prolamine-box binding factor, and o2 heterodimerizing proteins. Plant Cell 2015, 21, 1–11. [Google Scholar]
- Zhou, Z.; Song, L.; Zhang, X.; Li, X.; Yan, N.; Xia, R.; Zhu, H.; Weng, J.; Hao, Z.; Zhang, D.; et al. Introgression of opaque-2 into waxy maize causes extensive biochemical and proteomic changes in endosperm. PLoS ONE 2016, 11, e0158971. [Google Scholar]
- Gupta, H.S.; Agarwal, P.K.; Mahajan, V. Quality protein maize for nutritional security: Rapid development of short duration hybrids through molecular marker assisted breeding. Curr. Sci. 2009, 96, 230–237. [Google Scholar]
- Gupta, H.S.; Badu, R.; Agarwal, P.K.; Mahajan, V. Accelarated development of quality protein maize hybrids through marker-assisted introgression of opaque-2 allele. Plant Breed. 2013, 132, 77–82. [Google Scholar] [CrossRef]
- DAFF (Department of Agriculture Forestry and Fisheries). South African Varietal List as Maintained by the Registrar of Plant Improvement. Seed Crops, November 2015; p. 38. Available online: http://www.nda.agric.za (accessed on 24 October 2017).
- FAO. Faostat. Statistical Database of the Food and Agriculture of the United Nations, 2012. Available online: http://faostat.fao.org/site/567/ (accessed on 31 March 2012).
- Nedi, G.; Alamerew, S.; Tulu, L. Review on Quality Protein Maize breeding for Ethiopia. J. Biol. Agric. Healthc. 2016, 6, 84–96. [Google Scholar]
- Atlin, G.N.; Palacios, N.; Babu, R.; Das, B.; Twumasi-Afriye, S.; Friesen, D.K.; De Groote, H.; Vivek, B.; Pixley, K.V. Quality protein maize: Progress and prospects. Plant Breed. Rev. 2011, 34, 83–129. [Google Scholar]
- Gregory, T.; Sewando, P. Determinants of the probability of adopting of Quality Protein Maize (QPM) technology in Tanzania: A logistic regression analysis. Int. J. Dev. Sustain. 2013, 2, 729–746. [Google Scholar]
- Bateman, A.J. Contamination of seed crops II. Wind pollination. Heredity 1947, 1, 235–246. [Google Scholar] [CrossRef]
- Twumasi-Afriyie, S.; Dzah, B.D.; Ahenkora, K. Why QPM moved in Ghana. In Maize Productivity Gains through Research and Productivity Dissemination, Proceedings of the Fifth Eastern and Southern Africa Maize Conference, Arusha, Tanzania, 3–7 June 1996; Ransom, J.K., Palmer, A.F.E., Mduruma, Z.O., Waddington, S.R., Pixley, K.V., Jewel, D.C., Eds.; CIMMYT: Addis Ababa, Ethiopia, 1997; pp. 28–31. [Google Scholar]
- Burris, J.S. Adventitious pollen intrusion into hybrid maize seed production fields. In Proceedings of the Fifty-Sixth Annual Corn and Sorghum Research Conference, Chicago, IL, USA, 5–7 December 2001; Falasca, R.J., Ed.; American Seed Trade Association Inc.: Washington, DC, USA, 2001. [Google Scholar]
- Baltazar, M.B.; Schoper, J.B. Crop to crop gene flow: Dispersal of transgenes in maize, during field tests and commercialization. In Proceedings of the 7th International Symposium on the Biosafety of Genetically Modified Organisms, Beijing, China, 10–16 October 2002; pp. 21–28. [Google Scholar]
- Sperling, L.; Ashby, J.A.; Smith, M.E.; Weltzien, E.; Mcguire, S. A framework for analysing participatory plant breeding approaches and results. Euphytica 2001, 122, 439–450. [Google Scholar] [CrossRef]
- Weltzien, E.; Smith, M.E.; Meitzner, L.S.; Sperling, L. Technical and Institutional Issues in Participatory Plant Breeding-from the Perspective of Formal Plant Breeding: A Global Analysis of Issues, Results, and Current Experience; PPB Monograph Cali; PRGA: Cali, Colombia, 2003. [Google Scholar]
- Kisembo, L.G. The Impact of Participatory Plant Breeding and Selection on Adoption of Improved Sweetpotato Varieties in Uganda. Master’s Thesis, Makerere University, Kampala, Uganda, 2009. [Google Scholar]
- Olaoye, G.; Ajala, S.O.; Adedeji, S.A. Participatory selection of maize (Zea mays L.) variety for the control of stem borers in a Southeastern Nigeria location. J. Food Agric. Environ. 2009, 7, 508–512. [Google Scholar]
- Asumadu, H.; Ewool, M.B.; Obeng-Antwi, K. Participatory evaluation of drought tolerant maize varieties using mother-baby trial model: A case study in the Forest-Savannah transition zone of Ghana. J. Agron. 2011, 10, 68–73. [Google Scholar] [CrossRef]
- Tadesse, D.; Medhin, Z.G.; Ayalew, A. Participatory on farm evaluation of improved maize varieties in Chilga district of North Western Ethiopia. Int. J. Agric. For. 2014, 4, 402–407. [Google Scholar]
- FAO. Developing Sustainable Food Value Chains—Guiding Principles; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Mayne, J.; Johnson, N. Using theories of change in the CGIAR Research Program on Agriculture for Nutrition and Health. Evaluation 2015, 21, 407–428. [Google Scholar] [CrossRef]


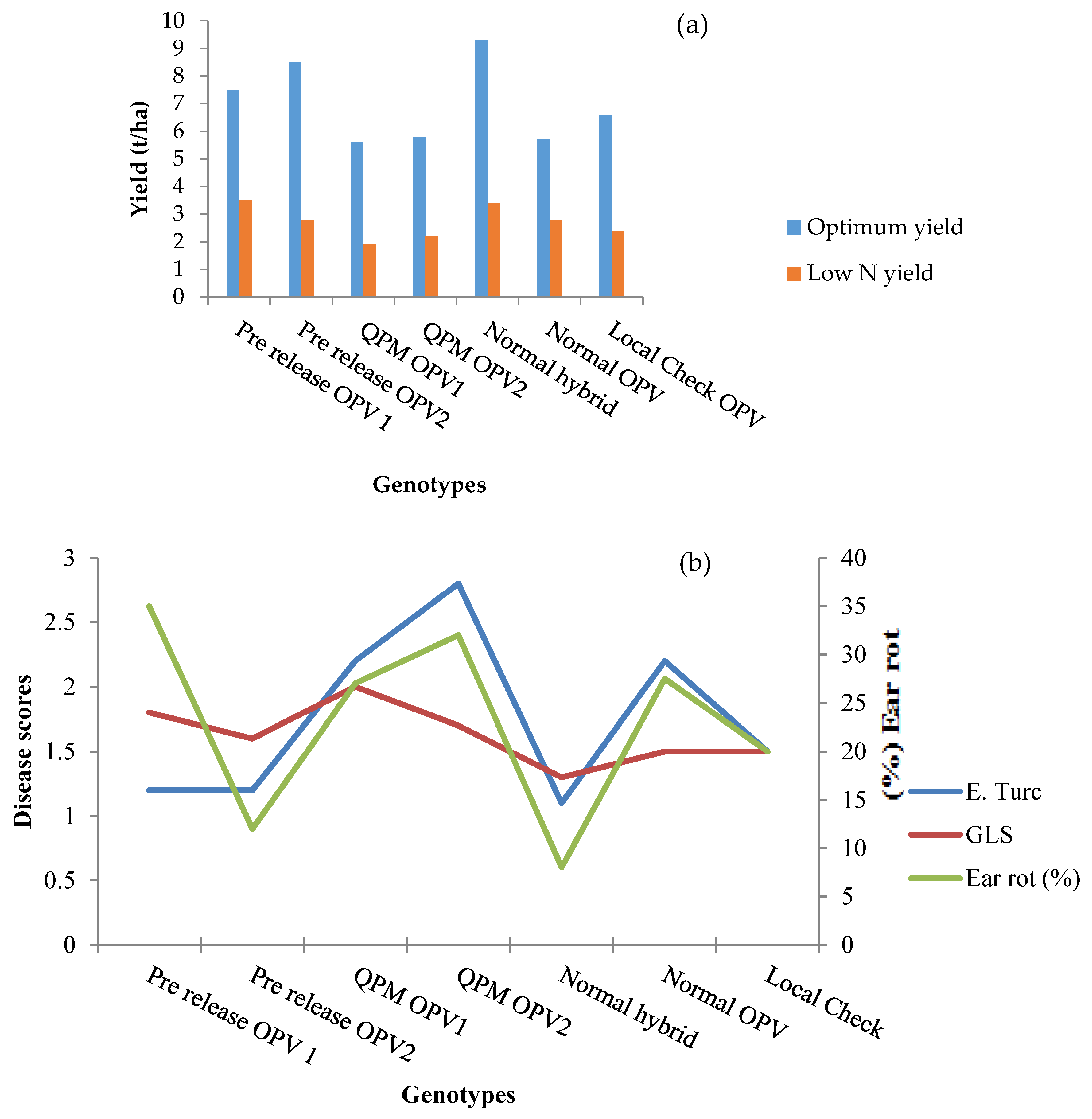{kind=link}
| Type of Variety | QPM Yield Range (t/ha) | Non-QPM Yield Range (t/ha) | Reference |
|---|---|---|---|
| OPVs and synthetics | 2–7.3 | >4 | [14,16] |
| Hybrids | 3–13.9 | >5 | [1,14,17,18,19] |
| Total Protein | Lysine | Tryptophan | |
|---|---|---|---|
| QPM (%) | >9 | 4.1 | 1 |
| Non-QPM (maize flour) | >9 | 2.7 | 0.6 |
| FAO recommendations (g/100 g total protein) | |||
| Children | 6.6 | 1.7 | |
| Adults | 1.6 | 0.5 | |
| Pedigree | Grain Yield (t/ha) | Ear Rot (%) | Silking (50%) | Endosperm Hardness * | Tryptophan (%) |
|---|---|---|---|---|---|
| CML-141 × CML-144 | 6.4 | 5.5 | 55 | 1.6 | 8.8 |
| (CML-141 × CML-144) CML-142 | 6.4 | 6.2 | 55 | 1.7 | 8.1 |
| CML-142 × CML-146 | 6.3 | 6.3 | 55 | 2.2 | 10 |
| CML-142 × CML150 | 6.2 | 7.8 | 55 | 2.0 | 8.9 |
| (CML-142 × CML150) CML-176 | 6.1 | 7.5 | 55 | 2.0 | 8.6 |
| CLQ-6203 × CML-150 | 5.8 | 7.2 | 55 | 2.3 | 9.0 |
| CML-144 × CML-159 | 5.9 | 5.9 | 56 | 1.9 | 9.3 |
| (CML-144 × CML-159) CML-176 | 5.6 | 6.0 | 56 | 1.7 | 9.4 |
| Local check | 5.9 | 7.6 | 55 | 1.9 | 5.0 |
| Plant Material | Environment | Combinability | Reference |
|---|---|---|---|
| Hybrids and inbred lines | Ethiopia | GCA effects were larger than SCA suggesting the effects of additive genes. | [49] |
| Single-cross hybrids | Well-watered, drought stress and low nitrogen conditions in Eastern Africa | GCA effects were significant indicating the effects of additive genes. | [23] |
| Extra-early QPM single-crosses | Drought and low-N environments in Nigeria. | GCA effects contributed more suggesting the effects of additive genes. | [26] |
| Single hybrids | Striga infested, drought, low-N and optimal environments in Nigeria. | GCA effects were more important suggesting the effects of additive gene action. | [20] |
| QPM hybrids | Drought, low soil N, and Striga infested environments in Nigeria | SCA effects were higher than GCA indicated the action of nonadditive genes. | [21] |
| Inbred lines and single cross hybrids | Combined drought and heat stresses in South Africa. | Grain yield was mostly controlled by non-additive gene action. | [17] |
| Single cross hybrids | Combined drought and heat stress in the Eastern Cape province of South Africa. | Additive genetic effects influenced most traits except grain yield, which was influenced by non-additive gene effects. | [27] |
| QPM hybrids | Rainfed condition in Ethiopia | Larger addidive genetic variation for grain yield. | [50] |
| Technic | Plant Material | Aim | Reference |
|---|---|---|---|
| Simple sequence repeat markers (SSR) | QPM and non-QPM | Marker assisted introgression of the o2 trait | [57] |
| ISSR primers and variety-diagnostic markers | QPM and non-QPM | Molecular breeding program development | [55] |
| SDS-PAGE | QPM grain | Protein identification | [59,60] |
| Proteomic analysis | QPM inbred lines | Identification of proteins associated with tolerance to combined drought and heat stress. | [28] |
| Inter-simple sequence repeat (ISSR) and random amplified polymorphic DNA (RAPD) markers. | QPM and non-QPM | Assess the level of genetic variation | [56] |
| SNP makers | QPM lines | Genetic diversity studies | [28] |
| SNP-based genetic distance | QPM inbred lines | Heterotic grouping | [20] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
N. Tandzi, L.; S. Mutengwa, C.; L. M. Ngonkeu, E.; Woïn, N.; Gracen, V. Breeding for Quality Protein Maize (QPM) Varieties: A Review. Agronomy 2017, 7, 80. https://doi.org/10.3390/agronomy7040080
N. Tandzi L, S. Mutengwa C, L. M. Ngonkeu E, Woïn N, Gracen V. Breeding for Quality Protein Maize (QPM) Varieties: A Review. Agronomy. 2017; 7(4):80. https://doi.org/10.3390/agronomy7040080
Chicago/Turabian StyleN. Tandzi, Liliane, Charles S. Mutengwa, Eddy L. M. Ngonkeu, Noé Woïn, and Vernon Gracen. 2017. "Breeding for Quality Protein Maize (QPM) Varieties: A Review" Agronomy 7, no. 4: 80. https://doi.org/10.3390/agronomy7040080
APA StyleN. Tandzi, L., S. Mutengwa, C., L. M. Ngonkeu, E., Woïn, N., & Gracen, V. (2017). Breeding for Quality Protein Maize (QPM) Varieties: A Review. Agronomy, 7(4), 80. https://doi.org/10.3390/agronomy7040080