
Biological Control of Spreading Dayflower (Commelina diffusa) with the Fungal Pathogen Phoma commelinicola

Abstract
:1. Introduction

2. Materials and Methods
2.1. Seed Sources, Test Plant Propagation
2.2. Isolation and Culture of Phoma commelinicola
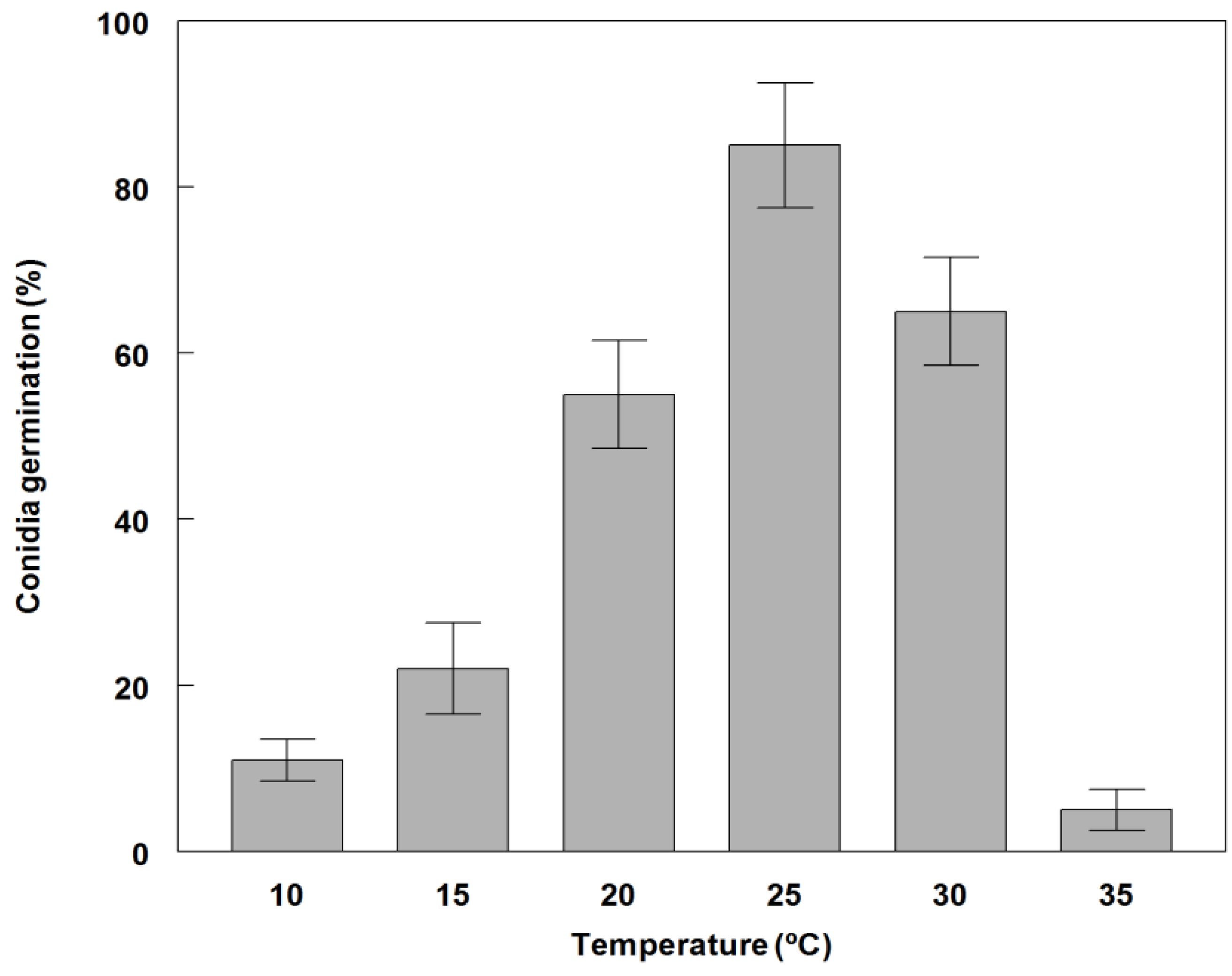
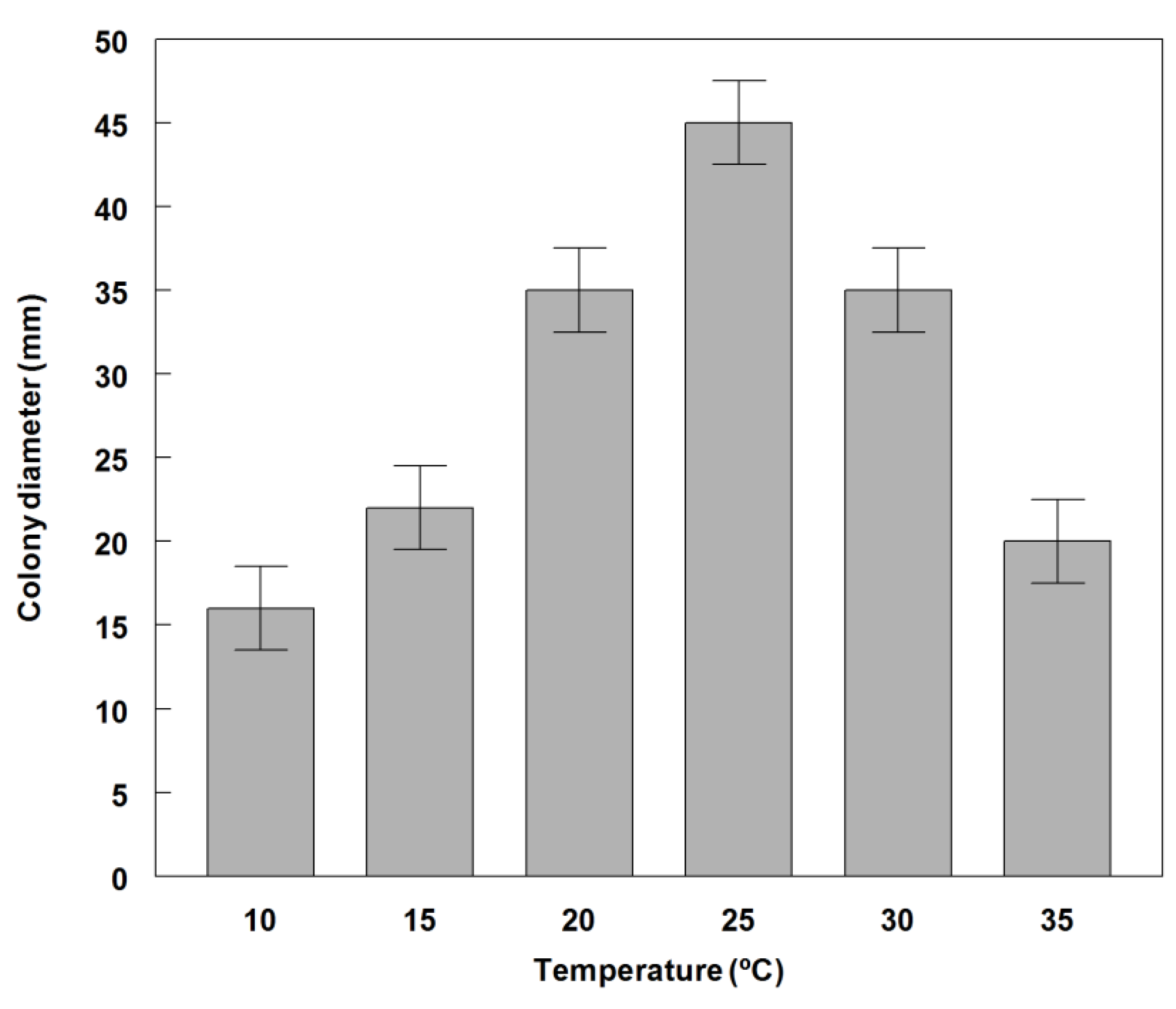
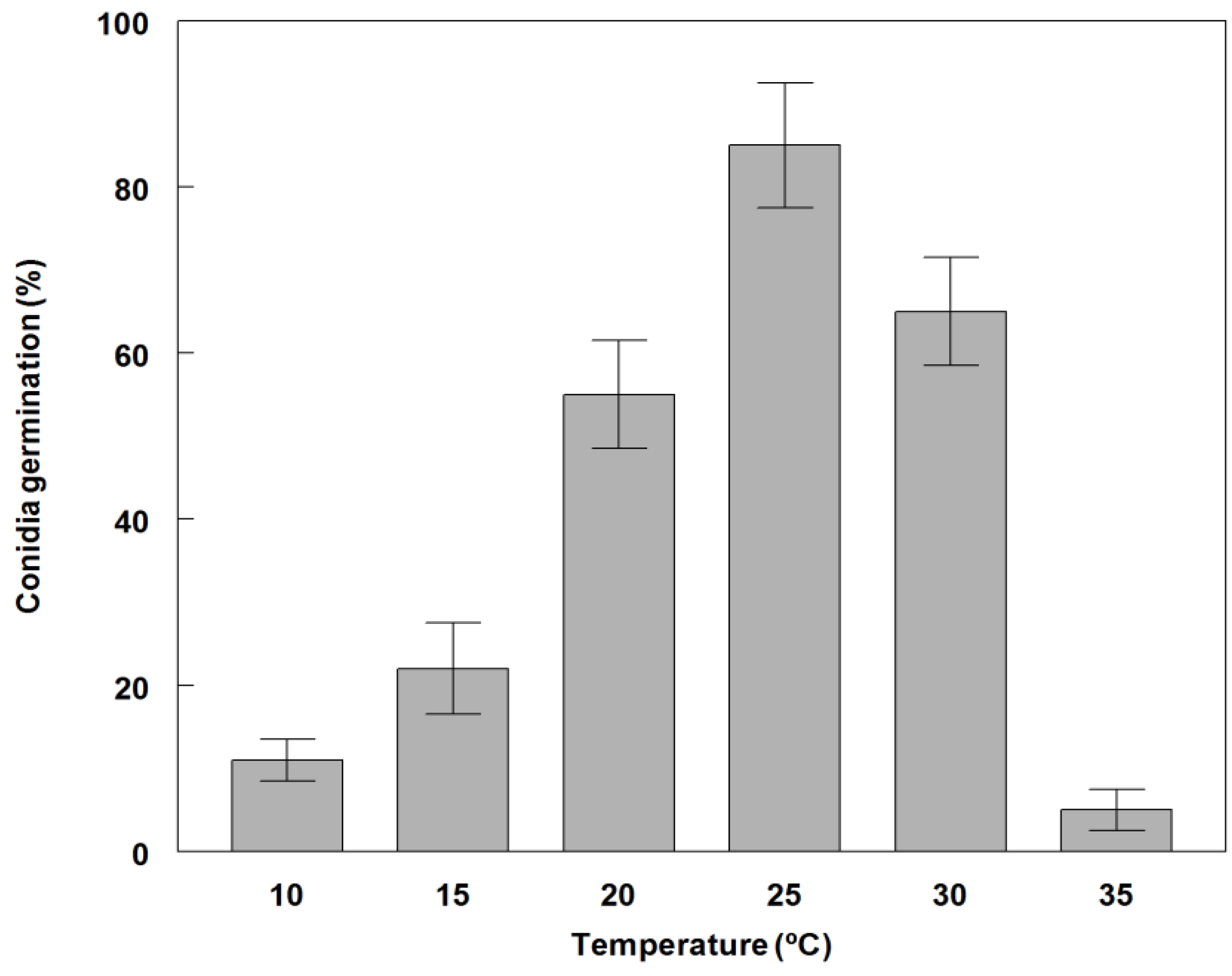
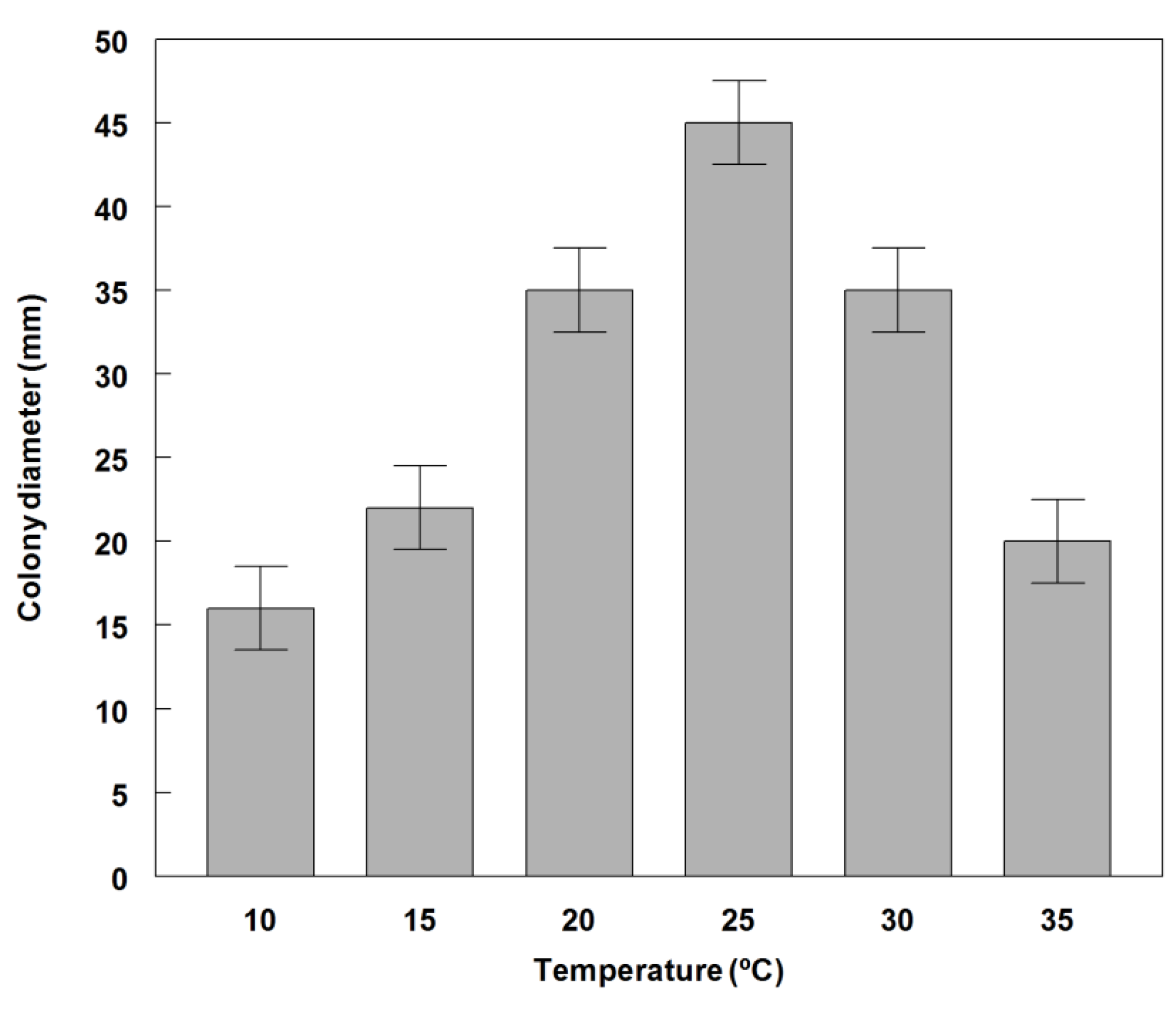
2.3. Effect of Temperature on Conidial Germination and Radial Growth Rate
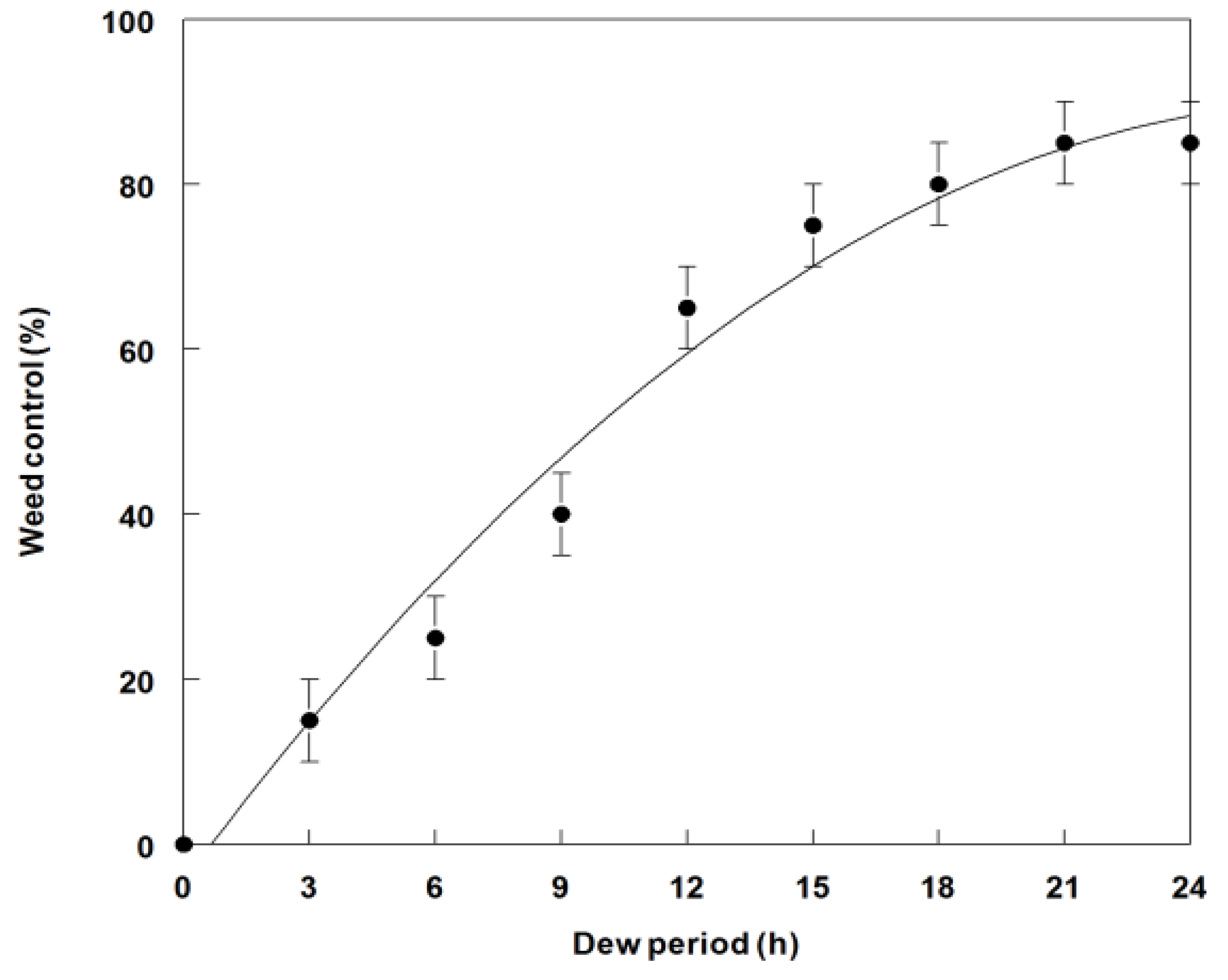
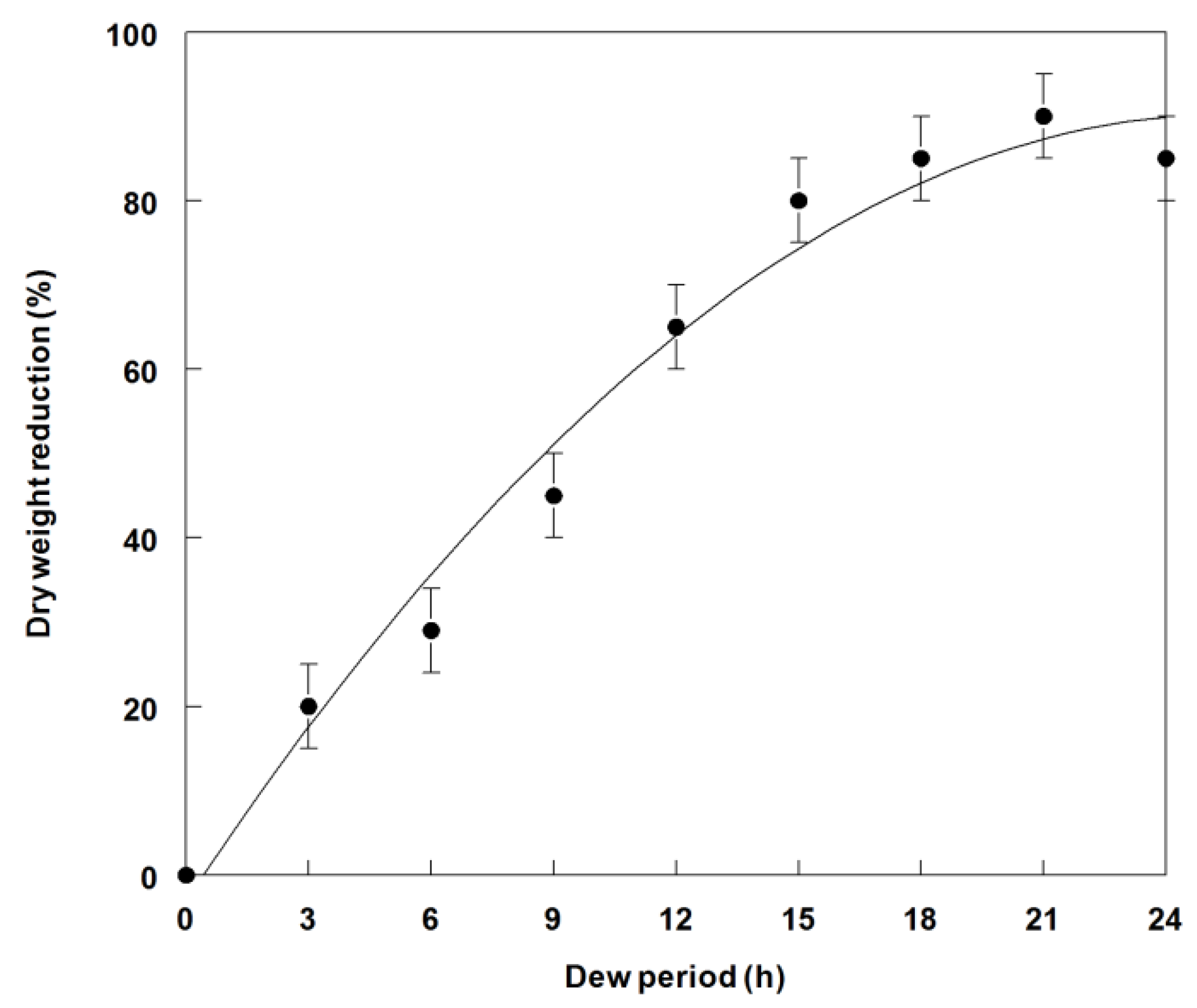
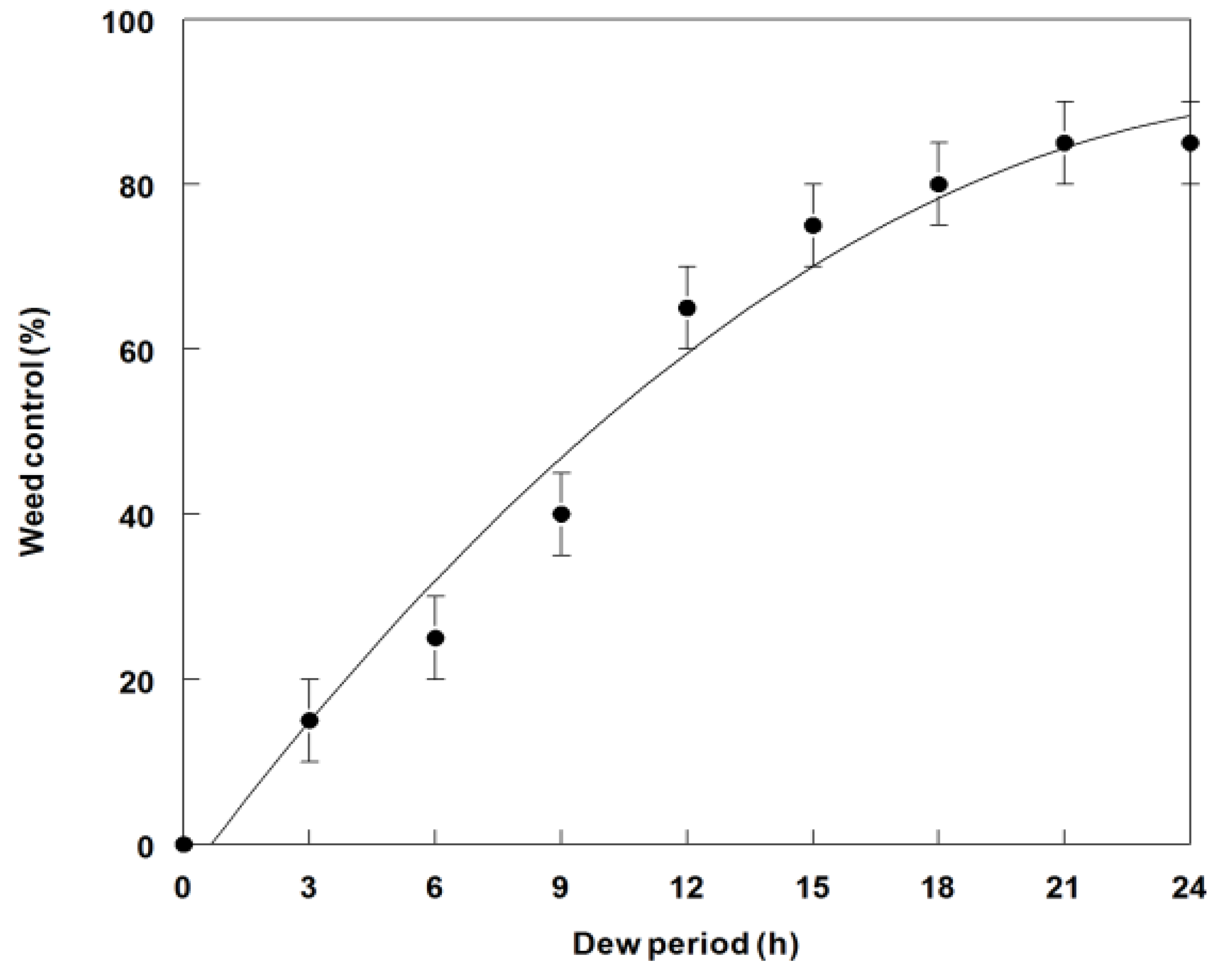
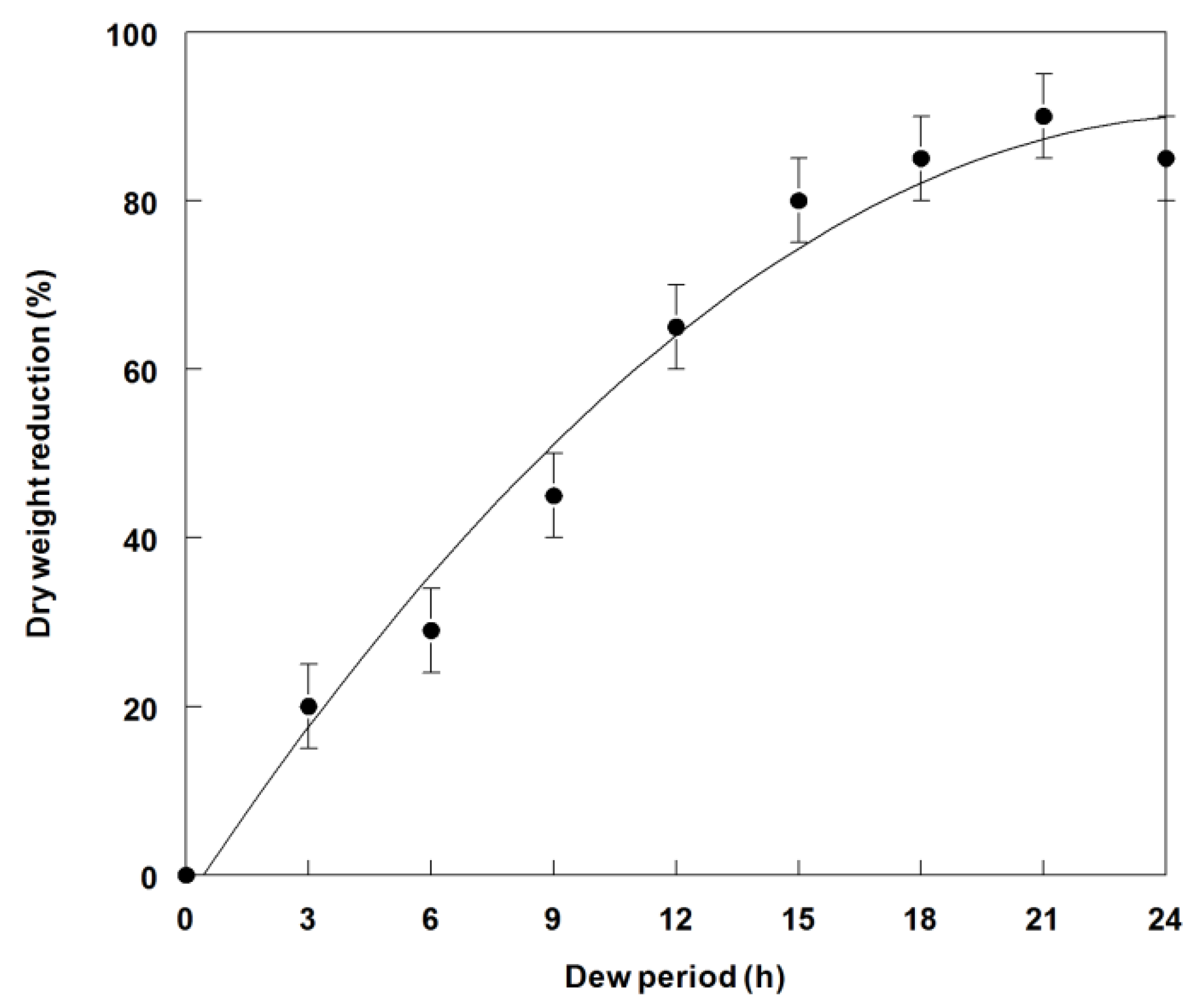
2.4. Effect of Dew Period Duration on Weed Control and Dry Weight Reduction of C. diffusa Seedlings
2.5. Effect of Inoculum Concentration and Plant Growth Stage
2.6. Effects of Phoma commelinicola on Crop Seedlings
2.7. Field Experiments
2.8. Statistical Procedures
3. Results and Discussion
3.1. Isolation of Phoma commelinicola
3.2. Germination and Growth of Phoma commelinicola

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth | Radial Growth (mm·day−1) | Conidia (108 plate−1) | ||||
|---|---|---|---|---|---|---|
| Light | Dark | Light/Dark | Light | Dark | Light/Dark | |
| WA | - | - | - | - | - | - |
| DFA | 10.2 a,c | 9.0 a | 10.5 a | 4.2 a | 2.8 a | 6.0 a |
| PDA | 9.5 b | 8.0 b | 9.9 b | 3.0 b | 1.3 b | 5.0 b |
| V8A | 8.4 c | 7.0 c | 8.5 c | 2.1 c | 0.9 c | 3.9 c |
| YEA | 7.5 d | 6.9 c | 7.4 d | 1.0 d | 0.3 d | 2.3 d |
| CDA | 4.0 e | 2.8 d | 4.1 e | 1.0 d | 0.2 d | 1.2 e |
3.3. Effects of Dew Period Duration on Weed Control and Dry Weight Reduction of C. diffusa Inoculated with P. commelinicola under Greenhouse Conditions
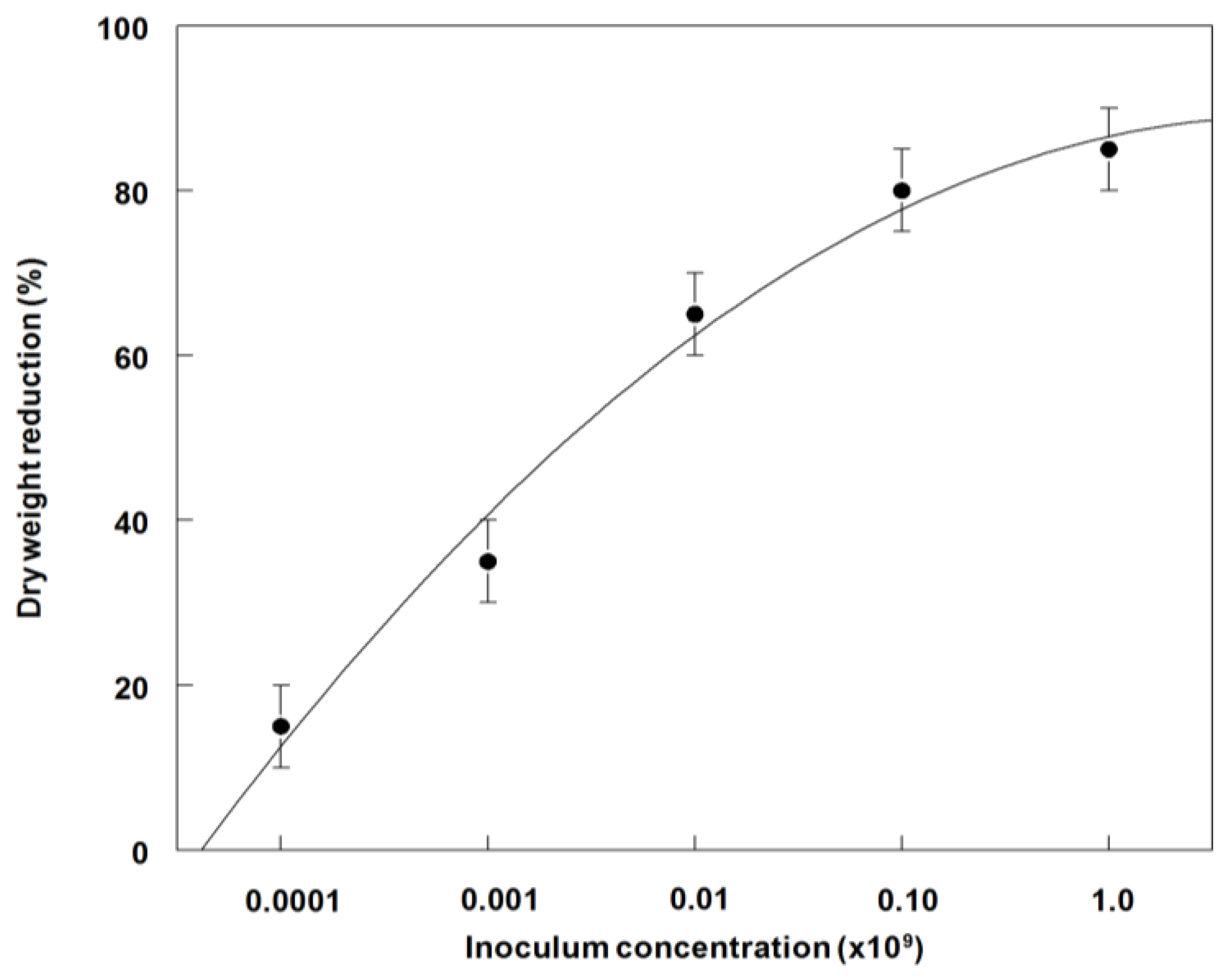
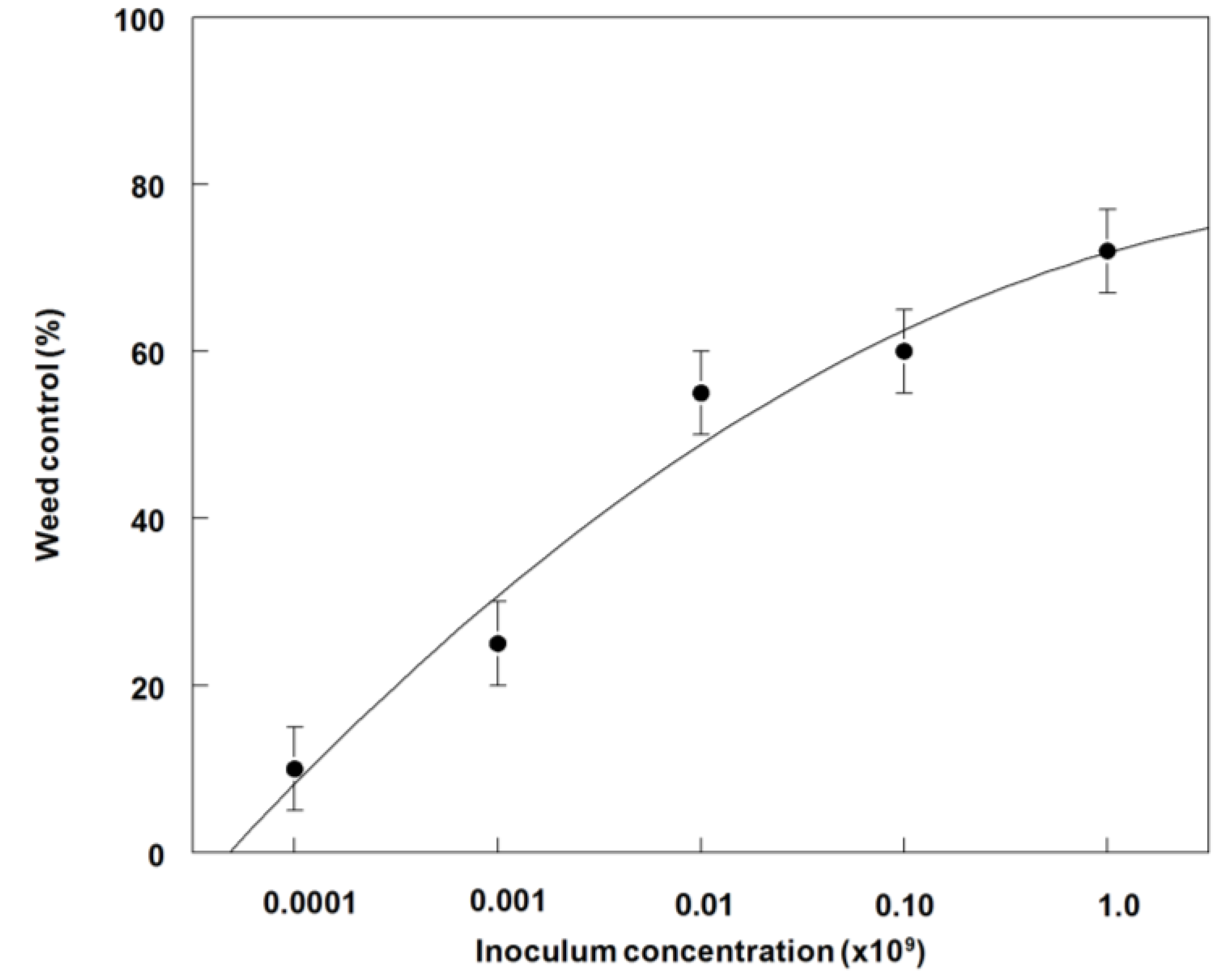
3.4. Effects of P. commelinicola Inoculum Concentration on Weed Control and Dry Weight Reduction of C. diffusa under Greenhouse Conditions
3.5. Disease Progression of P. commelinicola on C. diffusa under Greenhouse Conditions
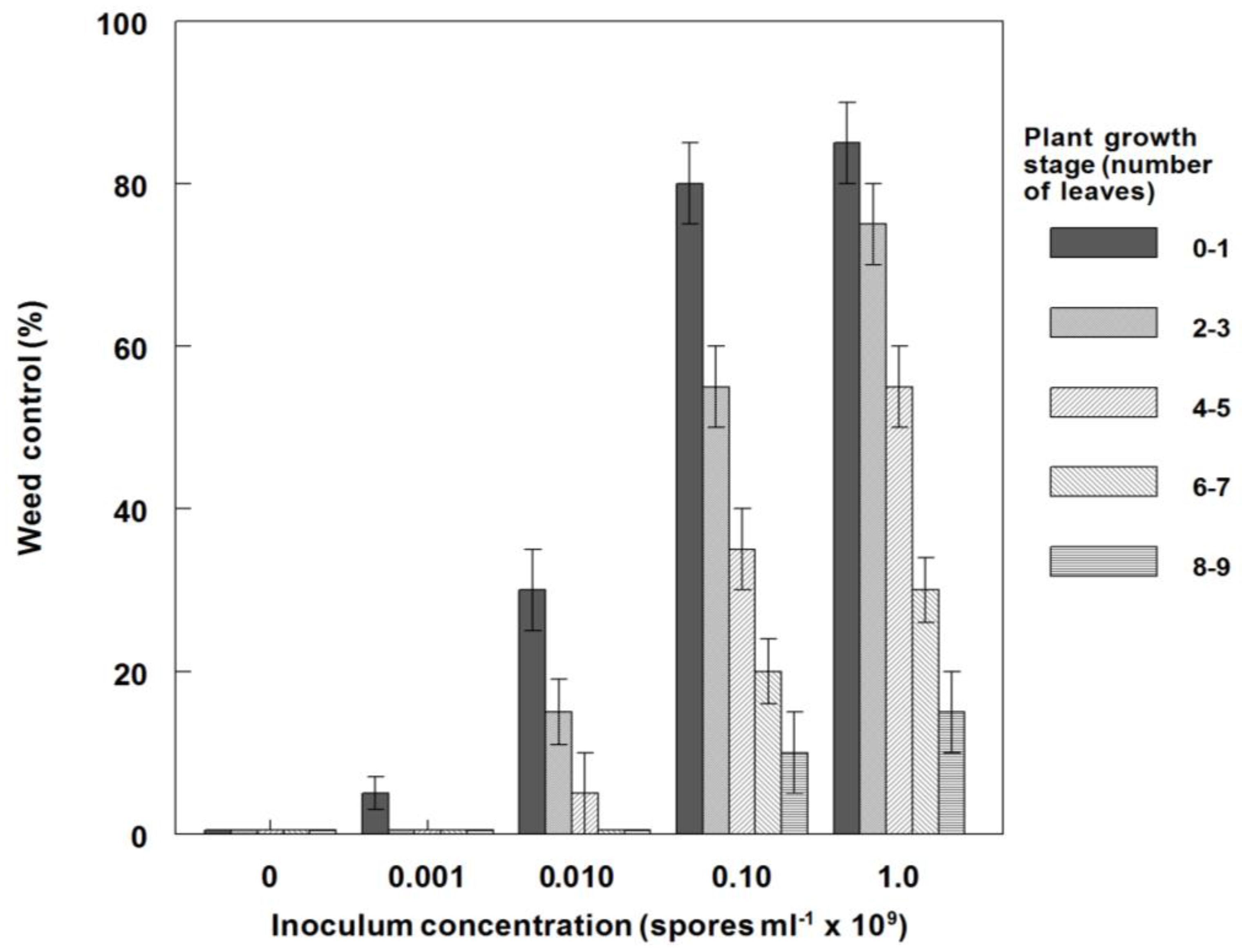
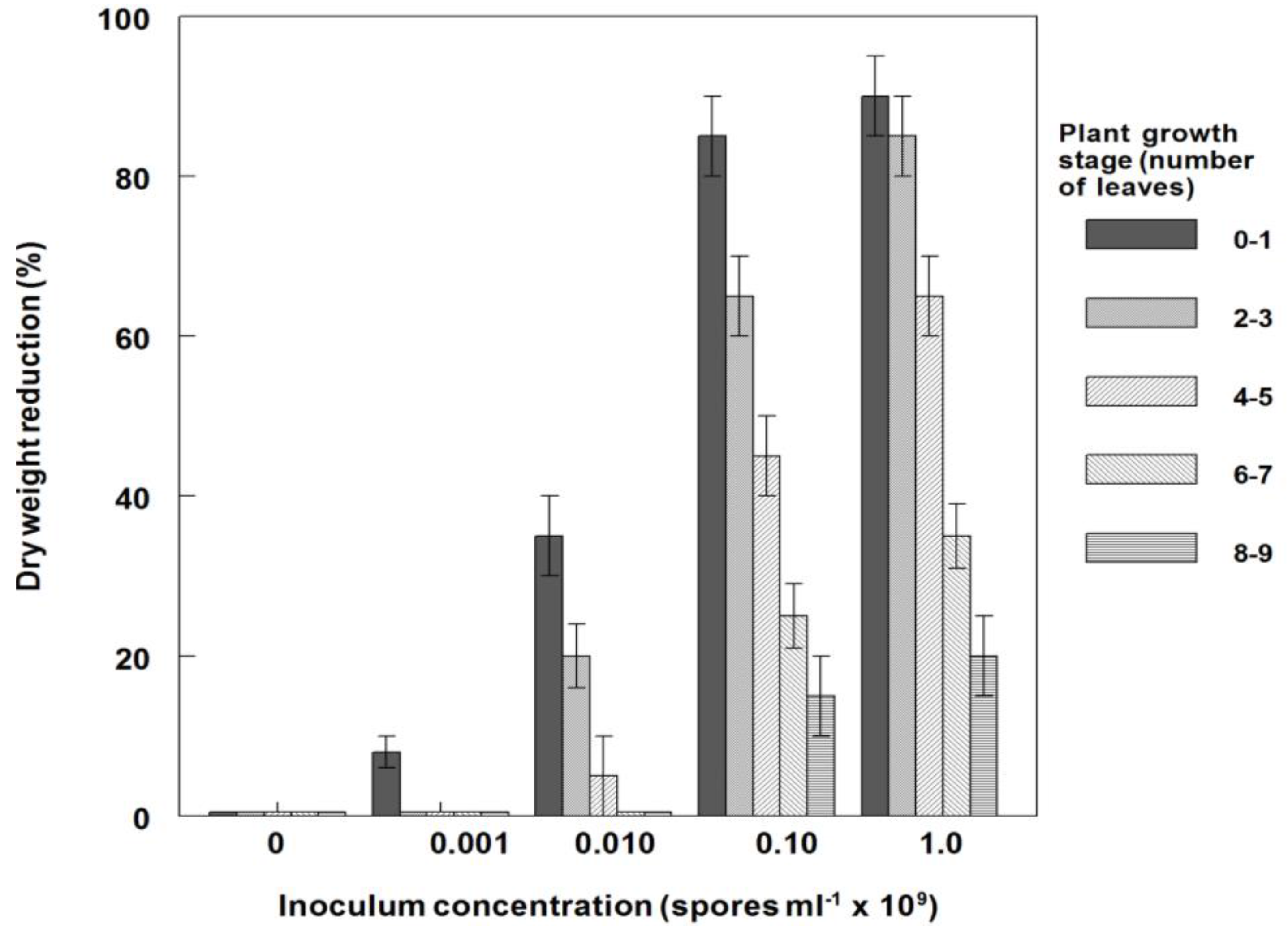
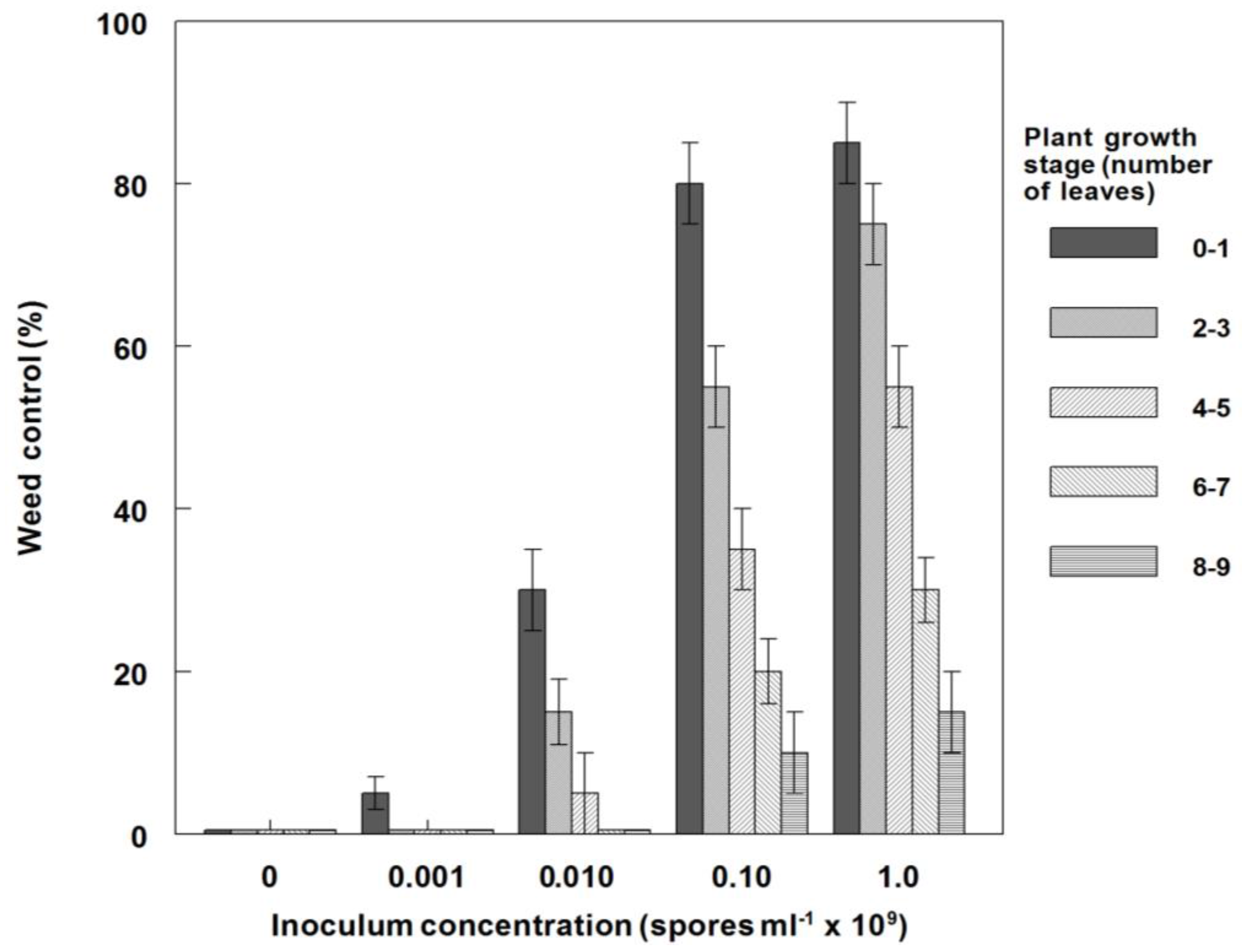
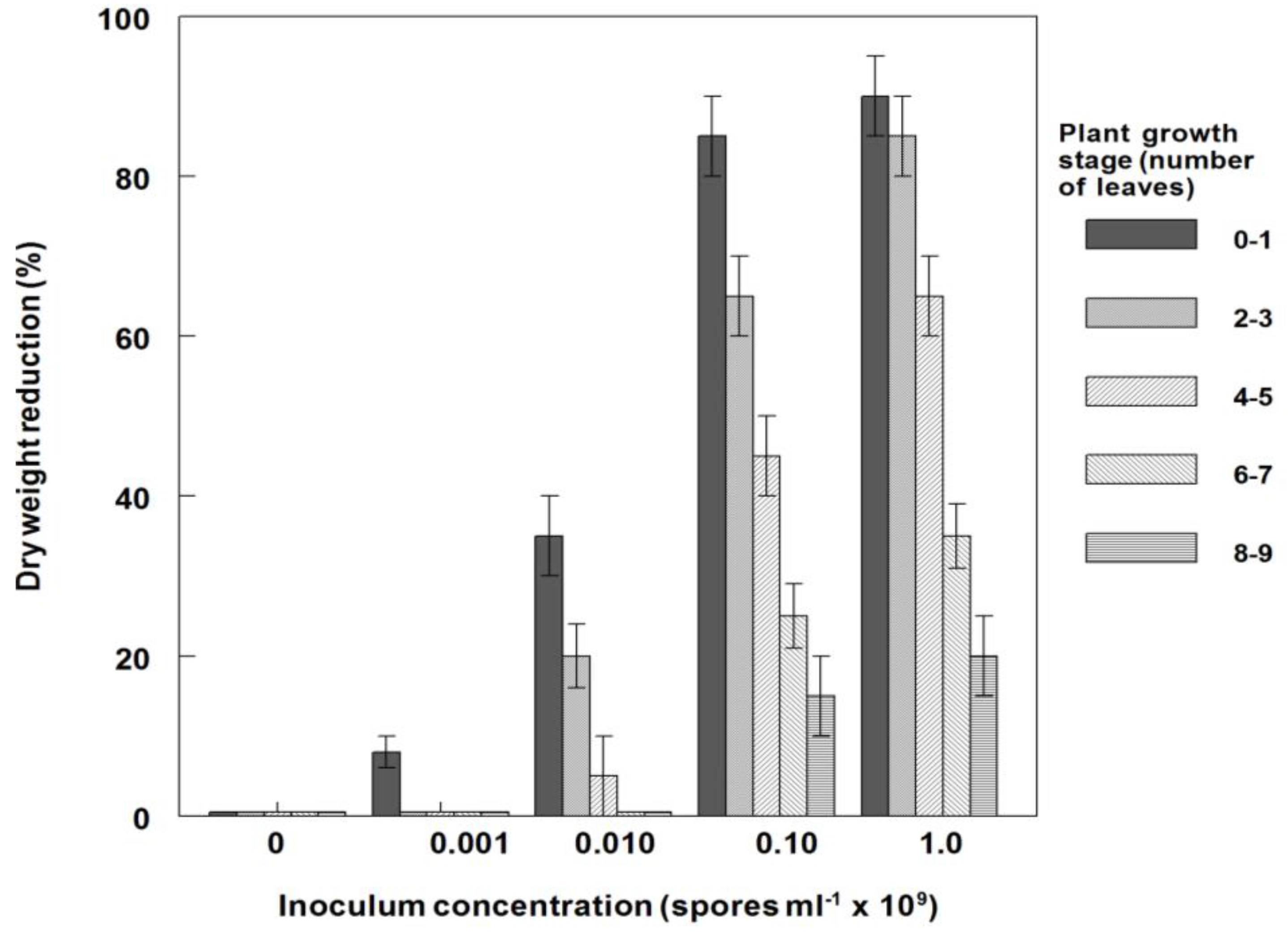
3.6. Effects of P. commelinicola Inoculum Concentration and C. diffusa Growth Stage on Weed Control and Dry Weight Reduction under Field Conditions
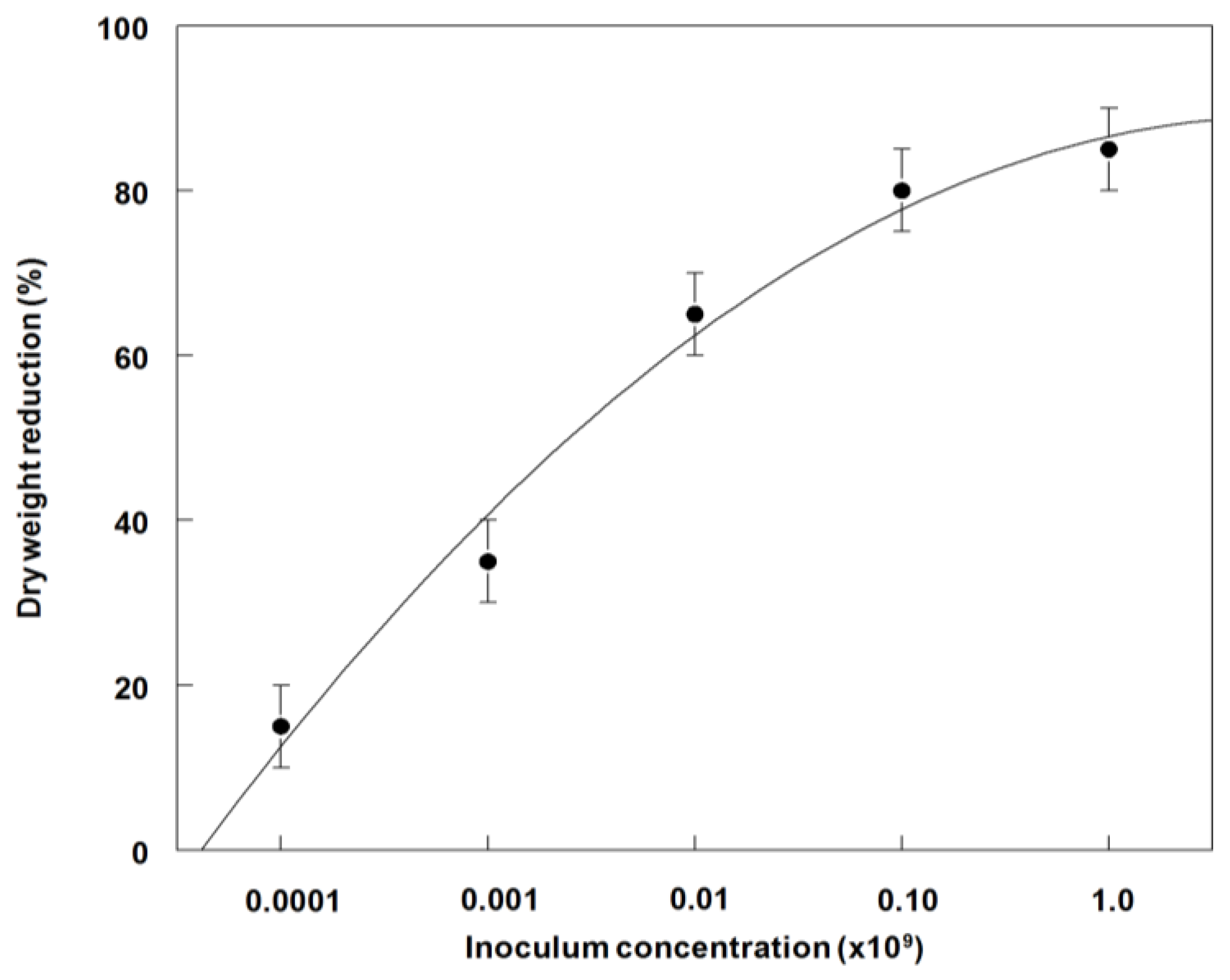
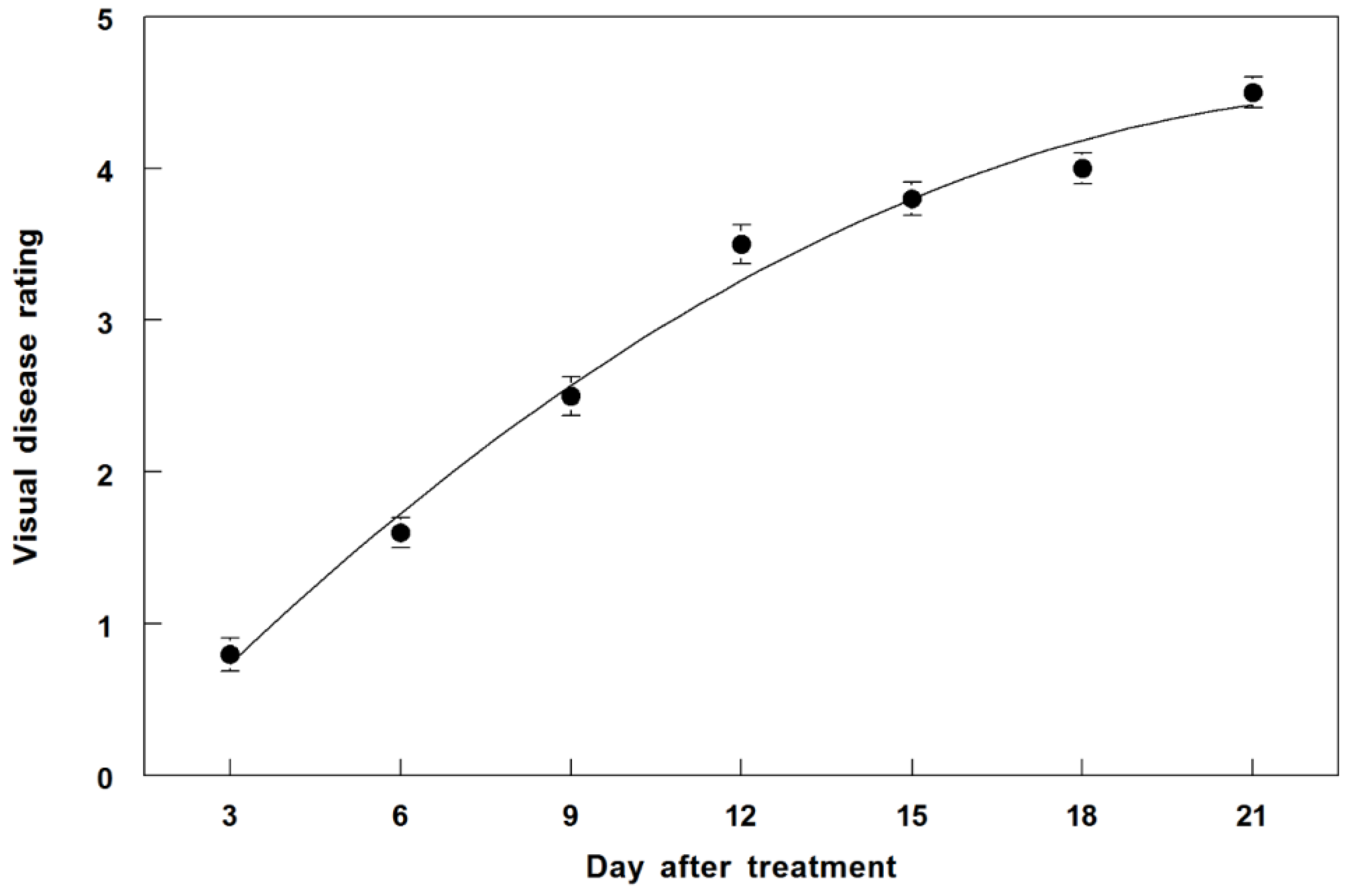
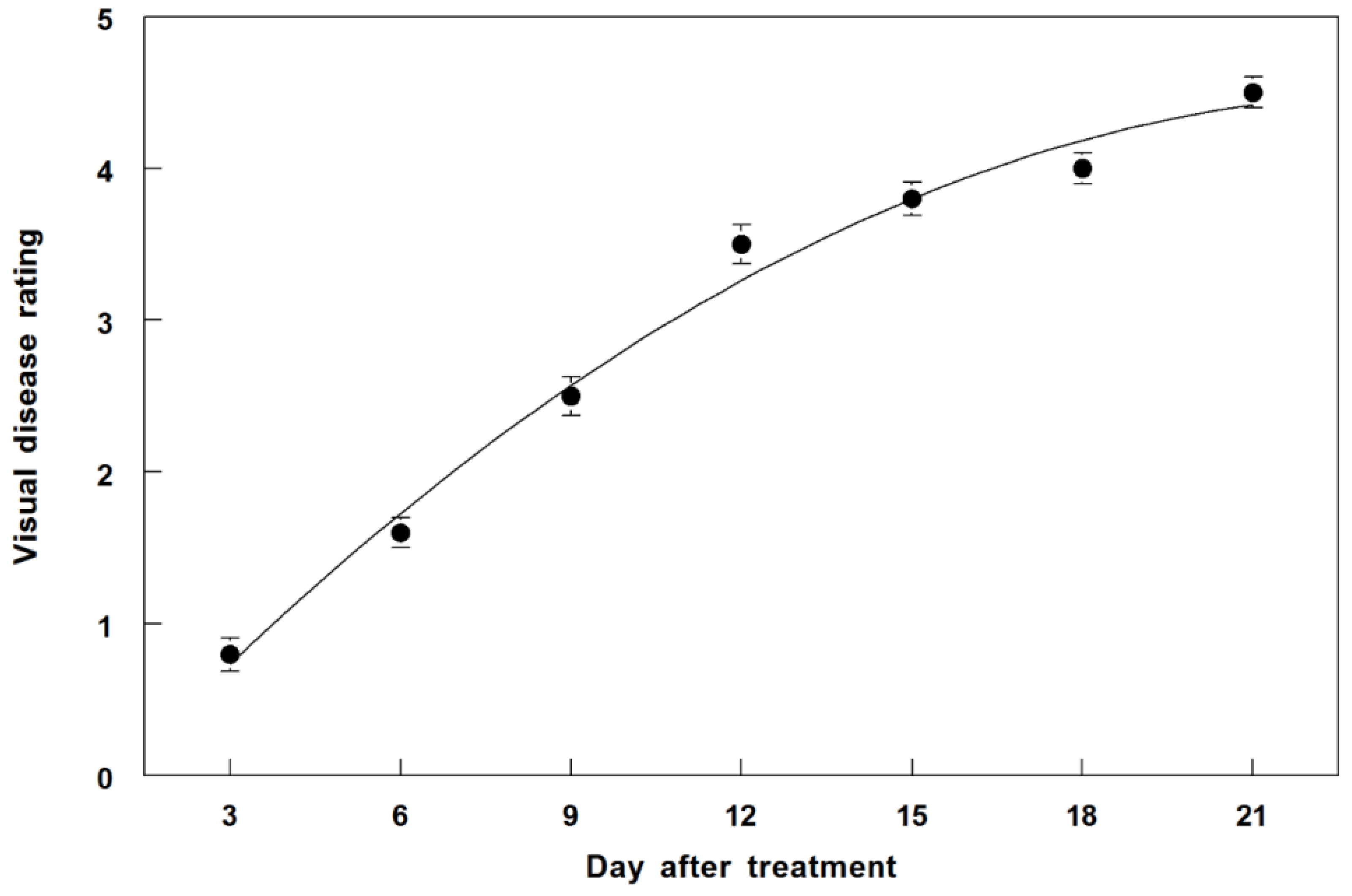
3.7. Disease Progression of P. commelinicola on C. diffusa under Field Conditions
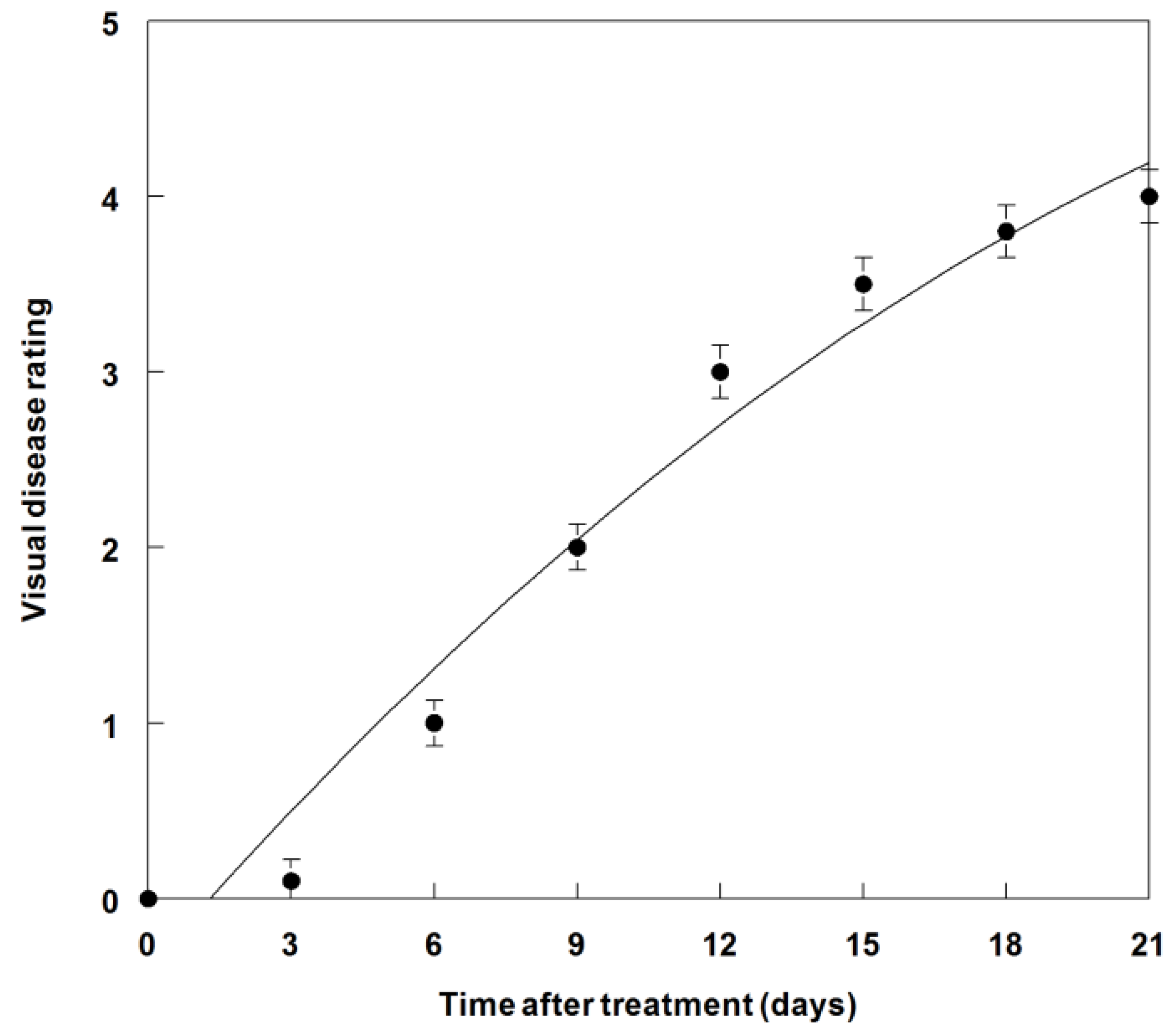
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bryson, C.T.; DeFelice, M.S. Weeds of the South; University of Georgia: Athens, GA, USA, 2009; p. 325. [Google Scholar]
- Isaac, W.A.; Gao, Z.; Li, M. Managing Commelina species: Prospects and limitations. Herbic. Curr. Res. Case Stud. Use 2013, 543–561. [Google Scholar] [CrossRef]
- Lanyasunya, T.P.; Wang, R.H.; Abdulrazak, S.A.; Mukisira, E.A. The potential of the weed, Commelina diffusa L., as a fodder crop for ruminants. S. Afr. J. Anim. Sci. 2006, 36, 28–32. [Google Scholar] [CrossRef]
- Invasive Species Compendium: Commelina diffusa (Spreading Dayflower). Available online: http://www.cabi.org/isc/datasheet/14979 (accessed on 2 July 2015).
- Webster, T.M. Weed survey—Southern States 2005: Broadleaf Crops Subsection (Cotton, Peanut, Soybean, Tobacco, and Forestry). Proc. South. Weed Sci. Soc. 2005, 58, 291–306. [Google Scholar]
- Norsworthy, J.K.; Burgos, N.R.; Scott, R.C.; Smith, K.L. Consultant perspectives on weed management needs in Arkansas rice. Weed Technol. 2007, 21, 832–839. [Google Scholar] [CrossRef]
- Singh, G.; Singh, Y.; Singh, V.P.; Johnson, D.E.; Mortimer, M. System level effects in weed management in rice-wheat cropping in India. In Proceedings of the British Crop Protection Conference International Congress on Crop Science and Technology (Glasgow, UK), Alton, Hampshire, UK, 2005; pp. 545–550.
- Queneherve, P.; Chabrier, C.; Auwerkerken, A.; Topart, P.; Martiny, B.; Martie-Luce, S. Status of weeds as reservoirs of plant parasitic nematodes in banana fields in Martinique. Crop Prot. 2006, 25, 860–867. [Google Scholar] [CrossRef]
- Robinson, A.F.; Inserra, R.N.; Caswell-Chen, E.P.; Vovlas, N.; Troccoli, A. Rotylenchulus species: Identification, distribution, host ranges and crop plant resistance. Nematropica 1997, 15, 165–170. [Google Scholar]
- Rich, J.R.; Brito, J.A.; Kaur, R.; Ferrell, J.A. Weed species as hosts of Meloidogyne: A review. Nematropica 2008, 39, 157–185. [Google Scholar]
- Richard, A.; Farreyrol, K.; Rodier, B.; Leoce-Mouk-San, K.; Wong, M.; Pearson, M.; Grisoni, M. Control of virus diseases in intensively cultivated vanilla plots of French Polynesia. Crop Protect. 2009, 28, 870–877. [Google Scholar] [CrossRef]
- Sosnoskie, L.M.; Hanson, B. Herbicide Resistance and Its Management. Available online: http://ucanr.edu/blogs/blogcore/postdetail.cfm?postnum=12365 (accessed on 6 July 2015).
- Isaac, W.A.P.; Brathwaite, R.A.I. Commelina species—A review of its weed status and possibilities for alternative weed management in the tropics. Agro. Thesis 2007, 5, 3–18. [Google Scholar]
- Wilson, A.K. Commelinaceae—A review of the distribution, biology and control of the important weeds belonging to this family. Int. J. Pest Manag. 1981, 27, 405–418. [Google Scholar] [CrossRef]
- Anonymous. CABI Crop Protection Compendium, Global Module; CAB International: Wallingford, UK, 2007; Available online: http://www.cabi/compendia/cpc/index.htm (accessed on 21 June 2015).
- Dodds, D.; Calcote, K.; Byrd, J. Weed Control Guidelines for Mississippi; Mississippi State University Extension Service and Mississippi Agricultural and Forestry Experimental Station: Starkville, MS, USA, 2015; p. 64. [Google Scholar]
- Scott, R.C.; Boyd, J.W.; Selden, G.; Norsworthy, J.K.; Burgos, N. Recommended Chemicals for Weed and Brush Control—MP44; University of Arkansas Extension Publication: Fayetteville, AR, USA, 2015; p. 79. [Google Scholar]
- Fawcett, J. Glyphosate tolerant Asiatic dayflower (Commelina communis) control in no-till soybeans. Proc. North Cent. Weed Sci. Soc. 2002, 57, 183. [Google Scholar]
- Santos, L.D.T.; Meira, R.M.S.A.; Santos, I.C.; Ferreira, F.A. Effect of glyphosate on the morpho-anatomy of leaves and stems of C. diffusa and C. benghalensis. Planta Daninha 2004, 22, 101–107. [Google Scholar]
- Webster, T.M.; Burton, M.G.; Culpepper, A.S.; York, A.C.; Prostko, E.P. Tropical spiderwort (Commelina benghalensis): A tropical invader threatens agroecosystems of the Southern United States. Weed Technol. 2005, 19, 501–508. [Google Scholar] [CrossRef]
- Culpepper, A.S. Glyphosate-induced weed shifts. Weed Technol. 2006, 20, 277–281. [Google Scholar] [CrossRef]
- Isaac, W.A.P.; Brathwaite, R.A.I.; Cohen, J.E.; Bekele, I. Effects of alternative weed management strategies on Commelina diffusa Burm. infestations in fair trade banana (Musa spp.) in St. Vincent and the Grenadines. Crop Prot. 2007, 26, 1219–1225. [Google Scholar] [CrossRef]
- Webster, T.M.; Faircloth, W.H.; Flanders, J.T.; Prostko, E.P.; Grey, T.L. The critical period of Bengal dayflower (Commelina benghalensis) control in peanut. Weed Sci. 2007, 55, 359–364. [Google Scholar] [CrossRef]
- Ulloa, S.M.; Owen, M.D.K. Response of Asiatic dayflower (Commelina communis) to glyphosate and alternatives in soybean. Weed Sci. 2009, 57, 74–80. [Google Scholar] [CrossRef]
- Gomez, J.M. Glyphosate-tolerant Asiatic Dayflower (Commelina communis L.): Ecological, Biological and Physiological Factors Contributing to Its Adaptation to Iowa Agronomic Systems. Master Thesis, Iowa State University, Ames, IA, USA, 2012. [Google Scholar]
- Charudattan, R. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology. Biol. Control 2001, 46, 229–260. [Google Scholar]
- Charudattan, R. Ecological, practical, and political inputs into selection of weed targets: What makes a good biological control target? Biol. Control 2005, 35, 183–196. [Google Scholar] [CrossRef]
- Glare, T.; Caradus, J.; Gelemter, W.; Jackson, T.; Keyhani, N.; Kohl, J.; Marrone, P.; Morin, L.; Stewart, A. Have biopesticides come of age? Trends Biotechnol. 2012, 30, 250–258. [Google Scholar] [CrossRef]
- Weaver, M.A.; Lyn, M.E.; Boyette, C.D.; Hoagland, R.E. Bioherbicides for weed control. In Non-Chemical Weed Management; Updhyaya, M.K., Blackshaw, R.E., Eds.; CABI International: Cambridge, MA, USA, 2007; pp. 93–110. [Google Scholar]
- Duke, S.O.; Scheffler, B.E.; Boyette, C.D.; Dayan, F.E. Biotechnology in weed control. In Encyclopedia of Chemical Technology; Kirk, O., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2015; pp. 1–25. [Google Scholar]
- De Gruyter, J. Contributions towards a monograph of Phoma (Coelomycetes) IX. Section Macrospora. Persoonia 2002, 18, 85–102. [Google Scholar]
- Boyetchko, S.M.; Peng, G. Challenges and strategies for development of mycoherbicides. In Fungal Biotechnology in Agricultural, Food, and Environment Applications; Arora, D.K., Ed.; Marcel and Dekker: New York, NY, USA, 2004; pp. 111–121. [Google Scholar]
- Tuite, J. Plant Pathological Methods: Fungi and Bacteria; Burgess Publication Co.: Minneapolis, MN, USA, 1969. [Google Scholar]
- Steele, R.G.D.; Torrey, J.H.; Dickeys, D.A. Multiple Comparisons. In Principles and Procedures of Statistics—A Biometrical Approach; McGraw Hill: New York, NY, USA, 1997; p. 365. [Google Scholar]
- Horsfall, J.G.; Barratt, R.W. An improved grading system for measuring diseases. Phytopathology 1945, 35, 655. [Google Scholar]
- Zhao, S.; Shamoun, S.F. Effects of culture media, temperature, pH, and light on growth, sporulation, germination, and bioherbicidal efficacy of Phoma exigua, a potential biological control agent for salal (Gaultheria shallon). Biocontrol Sci. Technol. 2006, 16, 1043–1055. [Google Scholar] [CrossRef]
- Heiny, D.K.; Templeton, G.E. Effects of spore concentration, temperature, and dew period on disease of field bindweed caused by Phoma proboscis. Phytopathology 1991, 81, 905–909. [Google Scholar] [CrossRef]
- Auld, B.A.; Morin, L. Constraints in the development of bioherbicides. Weed Technol. 1995, 9, 638–652. [Google Scholar]
- Heiny, D.K. Phoma probocis sp. nov. pathogenic on Convolvoulus arvensis. Mycotaxon 1990, 36, 457–471. [Google Scholar]
- Heiny, D.K. Field survival of Phoma proboscis and synergism with herbicides for control of field bindweed. Plant Dis. 1994, 78, 1156–1164. [Google Scholar] [CrossRef]
- Gu, Z.M.; Ji, M.S.; Li, X.H.; Qi, Z.Q. Effects of environmental factors on effectiveness of Phoma herbarum strain SYAU-06 against Commelina communis. Chin. J. Biol. Control 2009, 25, 355–358. [Google Scholar]
- Rajak, R.C.; Farkya, S.; Hasija, S.K.; Pandey, A.K. Fungi associated with congress weed (Parthenium hysterophorus L.). Proc. Nat. Acad. Sci. India 1990, 60, 165–168. [Google Scholar]
- Pandey, A.K.; Luka, R.S.; Hasija, S.K.; Rajak, R.C. Pathogenicity of some fungi to Parthenium and obnoxious weed in Madhya Pardesh. J. Biol. Control 1991, 5, 113–115. [Google Scholar]
- Pandey, S.; Pandey, A.K. Mycoherbicidal potential of some fungi against Lantana camara L.: A preliminary observation. J. Trop. For. 2000, 16, 28–32. [Google Scholar]
- Pandey, A.K. Microorganism associated with weeds: Opportunities and challenges for their exploitation as herbicides. Int. J. Mendel 2000, 17, 59–62. [Google Scholar]
- Deshmukh, P.; Rai, M.K.; Kövics, G.; Irinyi, L.M.; Karaffa, E.M. Phomas—Can these fungi be used as biocontrol agents and sources of secondary metabolites? In Proceedings of the 4th International Plant Protection Symposium at Debrecen University and 11th Trans-Tisza Plant Protection Forum, Debrecen, Hungary, 18–19 October 2006; pp. 224–232.
- Quarles, W. New Biopesticides for IPM and Organic Production. IPM Pract. 2011, 33, 1–20. [Google Scholar]
- Neumann, S.; Boland, G.J. Influence of host and pathogen variables on the efficacy of Phoma herbarum, a potential biological control agent of Taraxacum officinale. Can. J. Bot. 2002, 80, 425–429. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M.; Boland, G.J. Selected cultural and environmental parameters influence disease severity of dandelion caused by the potential bioherbicidal fungi, Phoma herbarum and Phoma exigua. Biocontrol Sci. Technol. 2004, 14, 561–569. [Google Scholar] [CrossRef]
- Burgos, N.R.; Singh, V.; Tseng, T.-M.; Black, H.; Young, N.D.; Huang, Z.; Hyma, K.E.; Gealy, D.R.; Caicedo, A.L. The impact of herbicide-resistant rice technology on phenotypic diversity and population structure of United States weedy rice. Plant Physiol. 2014, 166, 1208–1220. [Google Scholar] [CrossRef]
- Scott, R.C.; Meins, K.B.; Smith, K.L. Tank-mix partners with Newpath herbicide for hemp sesbania control in a Clearfield rice-production system. Ark. Agric. Res. Ser. 2005, 54, 225–229. [Google Scholar]
- Norsworthy, J.K.; Ward, S.M.; Shaw, D.R.; Llewellyn, R.S.; Nichols, R.L.; Webster, T.M.; Bradley, K.W.; Frisvold, G.; Powles, S.B.; Burgos, N.R.; et al. Reducing the risks of herbicide resistance: Best management practices and recommendations. Weed Sci. 2012, 60, 31–62. [Google Scholar] [CrossRef]
- Hoagland, R.E.; Boyette, C.D.; Vaughn, K.C. Interactions of quinclorac with a bioherbicidal strain of Myrothecium verrucaria. Pest Technol. 2011, 5, 88–96. [Google Scholar]
- Boyette, C.D.; Hoagland, R.E.; Weaver, M.A.; Stetina, K.C. Interaction of the bioherbicide Myrothecium verrucaria and glyphosate for kudzu control. Am. J. Plant Sci. 2014, 5, 3943–3956. [Google Scholar] [CrossRef]
- Cartwright, K.; Boyette, C.D.; Roberts, M. Lockdown: Collego bioherbicide gets a second act. Phytopathology 2010, 100. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyette, C.D.; Hoagland, R.E.; Stetina, K.C. Biological Control of Spreading Dayflower (Commelina diffusa) with the Fungal Pathogen Phoma commelinicola. Agronomy 2015, 5, 519-536. https://doi.org/10.3390/agronomy5040519
Boyette CD, Hoagland RE, Stetina KC. Biological Control of Spreading Dayflower (Commelina diffusa) with the Fungal Pathogen Phoma commelinicola. Agronomy. 2015; 5(4):519-536. https://doi.org/10.3390/agronomy5040519
Chicago/Turabian StyleBoyette, Clyde D., Robert E. Hoagland, and Kenneth C. Stetina. 2015. "Biological Control of Spreading Dayflower (Commelina diffusa) with the Fungal Pathogen Phoma commelinicola" Agronomy 5, no. 4: 519-536. https://doi.org/10.3390/agronomy5040519
APA StyleBoyette, C. D., Hoagland, R. E., & Stetina, K. C. (2015). Biological Control of Spreading Dayflower (Commelina diffusa) with the Fungal Pathogen Phoma commelinicola. Agronomy, 5(4), 519-536. https://doi.org/10.3390/agronomy5040519