Abstract
Monilinia laxa is the causal agent of brown rot disease on stone fruits, and also causes blossom wilt and twig canker. The common practice used to manage this disease is through fungicide treatments. However the demand to reduce fungicide inputs has been increasing and there is a growing number of reports of M. laxa strains that are resistant to fungicides. There is an urgent need to search for an alternative strategy to control the disease. This study focused on the isolation and characterisation of biological control agents (BCAs) using indigenous isolates isolated from cherries and plums collected within the UK. A total of 192 isolates were screened against two strains of M. laxa in a series of in vitro dual culture tests. From this in vitro screen, 12 isolates were selected for a subsequent in vivo screen on detached fruits, which then narrowed these isolates down to two potential BCAs. These two strains were identified as Bacillus amyloliquefaciens/subtilis (isolate B91) and Aureobasidium pullulans (isolate Y126). The capability of these two potential BCAs to grow and survive at a range of temperatures likely to be experienced under field and storage conditions was studied in order to gain knowledge for product formulation and field application. Bacillus sp. B91 was shown to be a mesophilic bacterium that could grow at 10–25 °C but suffered significant mortality at 0 and 5 °C, while A. pullulans Y126 was both mesophilic and psychrotolerant as it grew between 0–25 °C with the optimum at 20 °C. When all nutrients were removed, Y126 was able to survive for several weeks in all test temperatures (0–25 °C) but showed significant mortality at 25 °C. The capability of B91 to survive at 20 and 25 °C was higher than at low temperatures (0–15 °C). In addition, the modes of action of the potential BCAs were studied. B91 was shown to produce soluble and volatile organic compounds that inhibited M. laxa, while A. pullulans Y126 did not produce inhibitory compounds, but appeared to inhibit the pathogen via competition for nutrients. This study shows that microbial antagonists against M. laxa can be found from indigenous sources and that they are capable of preventing brown rot disease in controlled conditions, thus demonstrating a potential to be developed into commercial products.
1. Introduction
Economically important stone fruit species of Prunus include peach (P. persica var. persica), nectarine (P. persica var. nucipersica), apricot (P. armeniaca), plum (P. domestica) and sweet cherry (P. avium). Cherries and plums are high value crops in the UK and are susceptible to many fungal pathogens including Monilinia spp., Alternaria spp., Botrytis cinerea, Mucor piriformis, Pencillium expansum and Rhizopus spp. [1,2]. Of these pathogens, Monilinia spp., causing pre- and post-harvest brown rot, are the most destructive pathogens [2]. In North America and Australasia M. fructicola (G. Winter) is the major species responsible for the disease [3,4]. In Europe there are primarily two causal agents of brown rot, M. fructigena (Aderhold and Ruhland) and M. laxa (Aderhold and Ruhland); a third agent, M. fructicola, has been detected in many European countries although it is still considered a quarantine pathogen [5]. Monilinia laxa is most problematic as it can infect blossom, twigs, wounded and intact fruits, whilst M. fructigena can only infect wounded fruits. Fungicide treatment is the common pre-harvest practice to control brown rot disease [1], since post-harvest fungicide usage is not allowed [6]. In addition, strains of Monilinia spp. resistant to fungicides have emerged due to repeated use of fungicides [7,8]. Alternative control measures are needed for sustainable fruit production. In the last two decades, biological control agents (BCAs) have been increasingly studied worldwide and used to control brown rot disease, with most research focusing on M. fructicola [9,10,11,12,13] on peaches and nectarines. These studies have focused on bacteria [14], yeasts [11] and filamentous fungi [15], alone or integrated with physical methods [16]. In Europe, studies of biological control targeting brown rot pathogens have been carried out mainly in Spain [17,18,19] and Italy [20]. However, BCAs which have bioactivity against M. fructicola may not be able to control M. laxa, and the biological control of this pathogen on cherries and plums in the UK has been rarely studied. The candidate BCAs that have been developed specifically for brown rot disease originated in other countries outside the UK, and may thus be categorised as alien species for use within the UK, which may incur further legislative barriers [21,22]. For the purpose of registration, indigenous strains are preferred over alien ones.
Resident microbes that live epiphytically on plant surfaces are widely reported to be the most abundant source of antagonistic microbes against plant pathogens [10]. Bacteria and yeasts are favoured as BCAs because they are a rich source of bioactive compounds and require minimal nutrients and growth conditions which make them good candidates for development on an industrial scale [10,23]. For example Bacillus sublitis and its capability to produce iturin, an antibiotic used against pathogenic fungi, has been well studied and shown to be a promising BCA against Monilinia spp. [24,25,26]. Studies of many aspects of BCAs are needed for developing potential BCAs into commercial products. For example, when BCAs are introduced into natural ecosystems, there are many uncontrollable factors affecting the biocontrol efficacy. They might also have unforeseen impacts on humans and the environment. Information about potential BCAs about their identity, ecology, control mechanism, environmental impact, toxicology etc., are required for registration [21,27].
This paper reports studies aimed at finding indigenous microbial antagonists for future development as BCAs of M. laxa on cherry and plum in the UK. Specific objectives were (1) to obtain indigenous isolates and screen them for inhibitory activity against M. laxa; (2) to identify potential biocontrol isolates; (3) to study growth and survival of potential biocontrol isolates at a range of temperatures likely to be experienced in the biocontrol environment; and (4) to understand the modes of actions of the potential biocontrol isolates in inhibiting spore germination and mycelial growth.
2. Results and Discussion
2.1. In Vitro and in Vivo Screening of Microbes against M. laxa
A total of 192 microbes (92 bacterial strains and 100 yeast strains) were isolated and tested for mycelial inhibition against two strains of M. laxa on MEA. Thirty microbes were able to inhibit both strains of M. laxa with more than 50% inhibition. Fifty-seven microbes showed more than 50% inhibition against at least one of the two M. laxa strains and these 57 isolates were tested again against both strains of M. laxa on PDA. Although there was a positive relationship between inhibition of fungal mycelia on MEA and PDA, the levels of inhibition on MEA were generally higher (40%–80%) than on PDA (20%–60%). There were 12 strains that had levels of inhibition greater than the averages (60% on MEA and 40% on PDA) among the 57 isolates and these 12 strains were chosen for further in vivo screening on detached fruit. Plain and simple growth media were used rather than selective media as the latter contain selective and enrichment components which might favour or interfere with the bioactivity of the BCAs or the target pathogen. In vitro screening is normally conducted on one agar medium [28,29]. However the efficacy of BCAs varies and depends on the composition of culture media [30]. Therefore testing on at least two media may ensure that their bioactivity is less likely to be nutrient dependent.
In vitro screening for biocontrol efficacy may be conducted directly on fruits. For example, Zhang et al. [20] tested 210 isolates on peaches. However this approach is labour-intensive and hence may restrict the number of microbes that can be screened. We used a dual culture technique on agar media as a primary screen. In this study, epiphytes, the diverse group of microbes colonising on “phyllosphere”—the surface of the aerial parts of plants, were targeted [10]. The phyllosphere environment has been considered as a harsh and hostile environment for epiphytes because they are exposed to rapid fluctuations of environment as well as limited amounts of nutrients [31]. The ability to tolerate such harsh conditions is a prerequisite for a successful BCA. Based on these facts, a simple in vitro test was used here, and was used successfully to select a number of promising isolates against M. laxa, supporting the finding by Janisiewicz et al. [10,32] that plant material is a good source of BCAs, especially when the microbes share the same habitat with the pathogen.
The second screen was conducted in vivo, using wounded fruits inoculated with the pathogen. The percentage of M. laxa-infected cherry and plum wounds treated with Indar and isolates B91 and Y126 was close to zero and significantly (p < 0.001) lower than treatment with sterile distilled water (SDW) (Figure 1a,b). Isolates Y101, Y106 and Y110 significantly reduced percentage infection (p < 0.001) compared to SDW. For those few wounds that were infected, Indar, B91 and Y126 treatments led to smaller (p < 0.001) lesions than those in the SDW treatment (Figure 1c). The failure of the other 10 isolates to perform well in the in vivo screening may reflect differences in the conditions under which the screening were conducted. For example, nutrients leaked from artificial wounds on fruit surfaces may have affected the mechanism of inhibition used by the BCAs. This high level of attrition of potential candidates between in vitro and in vivo screens is common [20].
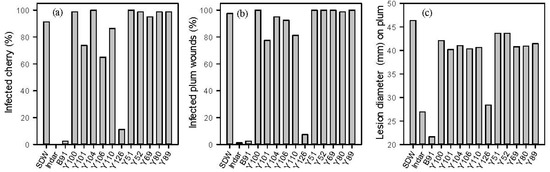
Figure 1.
Percentage of infected detached cherry and plum fruit when wounded, treated with 12 microbes (with concentrations between 107–108 CFU mL−1) and then inoculated with a conidial suspension of M. laxa (1 × 105 spores mL−1): on cherry (a), on plums (b), and average of lesion diameter (mm) on plum (c). There were two controls (1) Indar, a standard fungicide and (2) sterile distilled water (SDW). There were 80 cherry fruits each with one wound, and 40 plums each with two wounds. The inoculated fruits were incubated at room tempeature and the infection of M. laxa was assessed within 7 days after the inoculation with the pathogen.
2.2. Identification
The SSU sequence of isolate B91 (1148 bp) showed 100% DNA similarity to three Bacillus species, B. amyloliquefaciens, B. licheniformis and B. subtilis. In most cases, the SSU region is sufficient to identify bacterial isolates [33], but in this study the identification of B91 was limited to these three closely related species. As a consequence, a morphological study was carried out and a set of biochemical tests was used to further determine the species identity of isolate B91. The cells stained Gram-positive, were rod-shaped and the organism was a facultative anaerobe. Colonies on NA at 25 °C were creamy to light yellow, opaque, dull, spreading, velvety, smooth to slimy at a young stage but became rough and wrinkled by 14 days. These morphological characters are shared by B. subtilis and B. amyloliquefaciens and thus the exact identity of B91 cannot easily be determined. Alvarez et al. [34] also reported the difficulty and confusion of identification of these two species. Bacillus amyloliquefaciens used to be classified as sub-group of B. subtilis but was proposed as a new species from biochemical tests, gas liquid chromatography, DNA-DNA hybridisation and sequence analysis of recA and gyrA genes [35,36]. To differentiate these two species, at least two of these techniques should be used.
The LSU sequence of isolate Y126 (770 bp) showed a close similarity (97%–100%) with various fungal taxa including Aureobasidium, Botryosphaeria, Cladosporium, Dothidea and Mycosphaerella. The similarity of ITS sequence of Y126 (518 bp) with sequences of Aureobasidium pullulans (accession no. JN886796, JN886797, JN886798, HM849057, HM849619, EF690466 and AM160630) was between 99%–100%; A. pullulans was the only species showing this close affinity with Y126. Colonies of Y126 on PDA and MEA at 25 °C attained a diameter of 18 and 23 mm after seven days, respectively. Colonies were smooth, slimy and round and the colony colours of Y126 were compared to the colour chart of Kornerup & Wanscher [37]. On PDA, the surface colour of the colonies was light orange to pale orange (colour code 6A3/5A4), with the reverse orange white (colour code 5A6); the surface colour on MEA was pinkish-white to orange white (colour code 7A2/6A2), with the reverse pale yellow (colour code 4A3). The vegetative hyphae were hyaline, and stained in cotton blue, and were smooth, thin-walled and septate. Conidia were ellipsoidal or ovoidal, one-celled, smooth and produced in dense groups at terminal and subterminal ends of hyphae. Budding of conidia was observed, with secondary conidia usually smaller than primary conidia. These characters confirm that Y126 is A. pullulans and the tendency for the colony to darken with age suggests it is A. pullulans var. pullulans. It is worth noting that descriptions usually state that conidia are usually produced under “normal conditions”, for example on PDA or MEA incubated at 25 °C in the dark. In this study, isolate Y126 did not produce conidia under such conditions, but did so if cultured on PDA and incubated under UV light for seven days.
Cultures of Bacillus sp. B91 and A. pullulans Y126 were deposited in the National Centre for Industrial, Food and Marine Bacteria (NCIMB) and the National Collection of Fungus Cultures, CAB International under deposition codes, NCIMB30287 and IMI501717, respectively. The DNA sequences of the two strains were deposited in the GenBank database (accession numbers KC161970 for B91, and KC161971 and KC161972 for Y126).
2.3. Effect of Temperature on Antagonists B91 and Y126
2.3.1. Growth of B91 and Y126
Temperature had a highly significant effect on the growth of B91 [F(5,28) = 40.62, p < 0.001] and there were highly significant interactions between temperature and time [F(30,180) = 17.26, p < 0.001] (Figure 2a). At low temperatures (0 and 5 °C), the number of viable cells sharply decreased and remained stable after two weeks. At 10 °C the number of living cells decreased slowly initially for two weeks then gradually increased over the remaining four weeks. At 15, 20 and 25 °C, the number of living cells increased for the first week and remained steady thereafter. As B91, temperature also had significant effect on growth of Y126 [F(5,28) = 18.73, p < 0.001] and there were significant interactions between temperature and time, [F(30,180) = 12.58, p < 0.001] (Figure 2b). At 20 and 25 °C, the CFU number increased sharply during the first week, while there was also an increase but to a lesser extent in the number of living cells at 15 °C. After 2 weeks, the number of living cells remained stable and similar among the six temperatures.
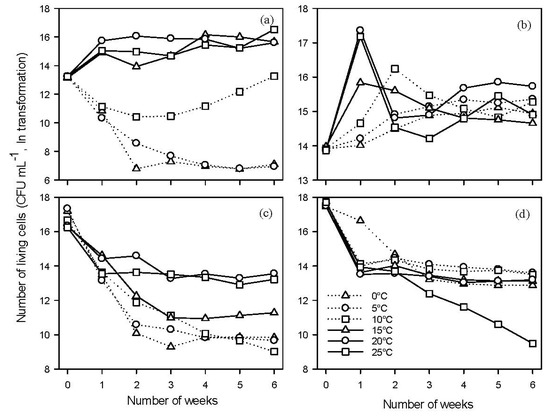
Figure 2.
The effects of temperature on the growth and survival of antagonists B91 and Y126. Liquid broth (NB and MEB) and phosphate buffer solution (PBS) were used in growth and survival studies, respectively. The cultures were incubated under a static condition over a period of six weeks: (a) growth of B91 in NB (b) growth of Y126 in MEB, (c) survival of B91 in PBS, and (d) survival of Y126 in PBS. Numbers of viable cells were quantified using a serial dilution method. Each point is an average of six replicates, three from each repeat experiment.
Studying the ability of BCAs to grow at a range of temperatures provides information for BCA development and application since BCAs must be able to grow and colonise expanding plant tissues under field conditions. The temperature range for active growth of B91 was narrower than that for Y126. B91 grew well at 15, 20 and 25 °C but the growth of B91 was limited at 0 and 5 °C. There was some recovery, however, of cell counts of B91 at 10 °C. This might have been an adaptation of bacterial cells to low temperatures. It is generally accepted that there are two phases of cold stress response in bacteria [38]: (1) an immediate response and (2) subsequent delayed response. At 10 °C, B91 took two weeks to adapt to cold then it was able to grow gradually over time.
In contrast, growth of Y126 was limited by lower temperatures but not prevented and, even at the lowest test temperature (0 °C) it still grew slowly. The growth rate of Y126 was highest at 20 and 25 °C for the first week, and then sharply decreased, which might be due to the fact that Y126 multiplied rapidly such that nutrients were possibly exhausted, resulting in the decline in numbers of living cells over the following week. According to their ability to grow at high, intermediate and low temperature, microbes are divided into three categories: thermophiles, mesophiles and psychophiles, respectively [39]. We can conclude that B91 is a mesophilic bacterium, while Y126 is both mesophilic and psychrotolerant.
2.3.2. Mortality of Antagonists B91 and Y126
Temperature had significant effects on survival of Bacillus sp. B91 [F(5,28) = 12.96, p < 0.001] and its interaction with time was also highly significant [F(30,180) = 6.12, p < 0.001] (Figure 2c). During the first week, the CFU number decreased sharply at all test temperatures. Thereafter, the CFU number at 20 and 25 °C remained steady for the next 4 weeks. In contrast, the CFU number at lower temperatures (at 0–15 °C) continued to decrease for further two weeks and then remained steady. When nutrients were present, B91 grew well if it was incubated at 15–25 °C, but poorly at low temperatures such that the reproduction rate was lower than the mortality rate. As for B91, temperature had a significant effect on survival of Y126 [F(5,28) = 22.70, p < 0.001] and its interaction with time also highly significant [F(30,180) = 13.16, p < 0.001] (Figure 2d). After initial steep decrease except at 0 °C, the CFU number maintained at a relatively high level thereafter except at 25 °C for which the viability steadily declined over the whole 6-week period. Overall, the survival for Y126 at 0, 5, 10, 15 and 20 °C were not significantly different from each other and was greater than at 25 °C.
Temperatures in post-harvest fruit storage conditions and during winter and early spring are usually low. Potential BCAs should be able to survive and maintain competitive numbers of living cells relative to the pathogen under these conditions. Bacillus species are more likely to tolerate high temperatures than low temperatures as they produce endospores which are heat-resistant [40]. The ability of BCAs to grow or survive at different temperatures, especially low temperatures, is crucial for their successful pre-harvest use during the blossom and early fruiting period and post-harvest use in cold stores. Both B. subtilis and B. amyloliquefaciens are known to live epiphytically on plant surfaces and in the upper layers of soil, and thus it might be expected that they are adapted to field conditions.
Although low temperatures initially reduced the viable counts of B91, it was still able to survive and maintain a lower but stable concentration for many weeks in all test temperatures. This result is supported by Tokuda et al. [41] who studied the survival of B. subtilis in sterile soil incubated at 15 and 25 °C. The cells of B. subtilis initially declined in the first week and then stabilised over the 50 day incubation period. Survival was higher if B91 was incubated at room temperature (20–25 °C), but began to decline faster at temperatures of 15 °C and below. It is likely that 15 °C is a critical temperature that triggers a cold response for B. subtilis [42]. Y126 was tolerant to a wide range of temperatures between 0–20 °C. To survive at 25 °C, it is likely that some basic nutrients have to be available. Some strains of A. pullulans have been reported to have higher temperature requirements, for example 30 °C was preferred for strains of A. pullulans isolates from India [43]. Y126 was isolated in the UK where the climate is colder, so this isolate may be less well adapted to high temperatures. Y126 managed to maintain a high concentration of cells at 0 °C throughout the study emphasising the psychrotolerant nature of A. pullulans. This observation is supported by strains of A. pullulans which have repeatedly been isolated from cold environments such as Arctic ice [44].
2.4. Possible Modes of Action
2.4.1. Inhibition of Spore Germination via Antibiosis and Competition for Nutrients
There was no significant difference in efficacy of BCAs conducted on PDB and WA [F(1,71) = 1.51, p = 0.224], suggesting the efficacy of BCAs did not depend on media used in this study. Therefore results presented here are combined from both media. There was a significant difference in the percentage of spore germination of M. laxa between the control and treatments with BCAs [F(1,97) = 88.92, p < 0.001]. Percentage germination for the control and BCA treatments were 88.5% and 20.3%, respectively. BCAs and culture type (broth vs. living cells) had significant effects on the percentage germination [F(1,71) = 168.86, p < 0.001 and F(1,71) = 256.11, p < 0.001, respectively]. There was an interaction between BCAs and culture type [F(1,71) = 223.21, p < 0.001]. Both the culture broth and living cells of B91 equally reduced the germination (Figure 3), suggesting that bioactivity of B91 did not depend on living cells being present. In contrast, bioactivity of Y126 was dependent on living cells being present, as the cell-free extract of Y126 did not inhibit spore germination.
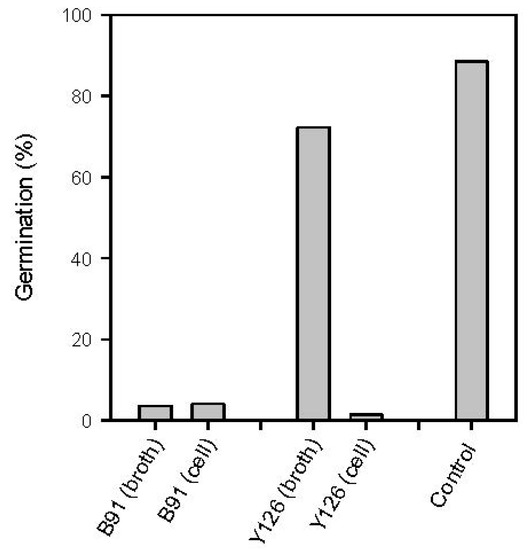
Figure 3.
Percentage germination of M. laxa spores treated with cell-free culture filtrate and living cells from two BCAs when tested on WA and PDB. The living cells were harvested by ultracentrifugation and the culture filtrate was filter-sterilised. Sterile PDB was used as a control. The mixture of M. laxa spores (1 × 105 spores mL−1) and each treatment was incubated at 25 °C in the dark and the germination of spores was assessed 12 h after the incubation. Each value was based on 2000 spores over two repeat experiments.
Cell-free culture filtrates of B91 were able to reduce spore germination, but living cells of Y126 were needed to inhibit the spore germination of M. laxa. This suggests the involvement of soluble, diffusible, bioactive substances produced and secreted into the culture filtrate by B91 in the inhibition of germination of M. laxa conidia. Nutrient competition might also have a contribution but this cannot easily be confirmed unless all target antibiotics are identified and excluded from the tests. Yanez-Mendizabal et al. [45] suggested that antibiosis was a major factor for bioactivity of B. subtilis CPA-8 in controlling brown rot disease on peaches caused by M. laxa and M. fructicola. They showed that cyclic lipopeptides, termed iturins, produced by CPA-8 were responsible for inhibitory activity. Iturins have also been reported from other isolates of B. subtilis and may well be responsible for the effects noted here [24,46].
On the other hand, living cells of Y126 were essential for bioactivity and that bioactive compounds were not detected in the cell-free filtrates. However, it might be that any bioactive compounds against M. laxa were produced but were not stable after they were secreted at room temperature. Zhang et al. [47] reported that A. pullulans PL5 completely inhibited spore germination of M. laxa tested in PDB, whereas dead cells or cell-free broth had no effect. Bioactivity of A. pullulans has been assigned to two mechanisms: (1) secretion of lytic enzymes and (2) competition for nutrients [48]. Enzymes produced by A. pullulans were intensively studied for their bioactivity [47,48] but these enzymes were enriched using optimum conditions to maximise the quantity of enzymes produced—which it is not likely to happen in a natural situation. Vero et al. [49] suggested that enzymes did not seem to play a major role in inhibiting pathogens or their spore germination because A. pullulans took at least 48 h to produce them in the log phase or even longer (7 to 10 days) if the culture was incubated at 5 °C. In this study, the bioactivity of A. pullulans Y126 potentially came from competition for nutrients rather than through production of enzymes as inhibition was manifested within 12 h of incubation.
Previous experiments have shown that A. pullulans inhibited P. expansum and M. laxa in low concentrations of apple and peach juices, respectively [47,50]. However when the concentrations of juices were higher, nutrient competition was no longer an issue and A. pullulans was unable to control the pathogens. This shows the important of competition for nutrients and to maximise efficacy of A. pullulans against pathogens, nutrients should be in low concentration—a situation common in the natural environment. Media used for screening process should contain enough nutrients to maintain growth of a BCA and the pathogen but should be a minimal medium in order to favour mechanisms based on nutrient competition. Therefore relatively simple agar such as PDB and WA were used in this study.
2.4.2. Inhibition of Mycelium Growth by Microbial VOCs
Two-way ANOVA showed that colonies of M. laxa co-cultured with either B91 or Y126 were significantly smaller than the colonies in the control treatment [F(2,89) = 597.05, p < 0.001]. An interaction between BCAs and isolates of M. laxa was present [F(2,89) = 25.29, p < 0.001]. The effect of VOCs produced by B91 was highly inhibitory against the two test isolates of M. laxa, while the inhibitory activity of VOCs from Y126 varied across the two test isolates of M. laxa. Although Y126 reduced colony diameters (mean 50.8 mm) of both M. laxa isolates compared to the control treatment (mean 66.8 mm), this effect was not as pronounced as that with B91 (mean 11.3 mm). Bioactivity of VOCs produced by various groups of microbes against Monilinia species have been reported elsewhere [9]. Further research is needed to understand the nature of these VOCs and assess their potential use in an industrial scale.
3. Experimental Section
3.1. In Vitro and In Vivo Screening
3.1.1. Microbial Isolation
Mummified fruits, healthy leaves and green fruits of cherries and plums were collected from seven orchards in the county of Kent, UK, during 2010–2011 for isolating microbes based on a published method [51]. The plant materials were submerged in 100 mL of 1% Triton X-100 contained in a Duran bottle. The bottle was shaken at 200 rpm for 1 h on a shaking incubator at room temperature. A serial dilution technique was used to isolate microbes. A 20 µL aliquot of each dilution was spread onto the isolation medium. Nutrient agar (NA) amended with cyclohexamine (0.05 g L−1) was used to isolate bacterial strains and malt extract agar (MEA) amended with streptomycin (0.05 g L−1) and chloramphenicol (0.05 g L−1) was used to isolate yeast strains. The plates were incubated at 25 °C for 3 days. Microbes were randomly selected and streaked on fresh NA and MEA without antibiotics and incubated under similar growth conditions. Pure cultures of the selected microbes were maintained at −80 °C until further use.
3.1.2. Dual Culture Screening
Two isolates of M. laxa, R92/10/44 and R118/10/19 from cherries and plums, respectively, were isolated by Dr. Angela Berrie at East Malling Research, from commercial orchards in Kent, UK. They were maintained on potato dextrose agar (PDA), and stored at 4 °C in the dark until required. An in vitro test of mycelium inhibition was adapted from the published dual culture methodology [51], in which the test bacterial or yeast isolates were inoculated 1 cm away from the margin of a Petri dish containing MEA. The plates were incubated at 25 °C for 3 days. An agar plug cut from actively growing mycelium of M. laxa using a cork borer (no. 2: diameter 5 mm) was placed centrally. The plates were incubated at 25 °C for a further 14 days. There were three replicate plates per treatment and per strain of M. laxa. Mycelium growth of M. laxa was measured. To confirm bioactivity, strains of microbes yielding >50% inhibition on MEA were tested on PDA using the same growth conditions as MEA. There were five replicate plates per treatment.
3.1.3. Production of M. laxa Inoculum
Plums bought from a supermarket were surface-sterilised by submerging in 70% ethanol for 5 min and then dried in a flow cabinet for 1 h. Four plum cultivars (Early Red, Pioneer, Ruby Nel and Lady Red) were used at different times of the year. Plums were wounded on the shoulders in a triangle shape of 1 cm width and 0.5 cm depth using a sterile blade. An agar plug with mycelium (0.5 cm3) from an actively-growing colony of M. laxa was inoculated into each wound. Four different strains of M. laxa were used, the 2 strains used in in vitro screen and 2 additional strains, R35/10/10 and R118/10/42, originating from cherries and plums, respectively. Inoculated plums were then placed in plastic boxes (two plums per box) and incubated at room temperature for 2–4 weeks. Conidiophores of M. laxa were harvested en masse using sterile forceps and suspended in a tube of 5 mL of sterile distilled water (SDW) resulting in inoculum comprising a mixture of all four strains. Spore concentration was estimated using a haemocytometer and adjusted to 1 × 105 conidia mL−1.
3.1.4. In Vivo Tests on Detached Fruits
There were 12 candidate BCA isolates selected from the dual culture screening [one bacterium (B91) and 11 yeasts (Y51, Y52, Y69, Y80, Y89, Y100, Y101, Y104, Y106, Y10 and Y126)] with two controls including Indar, a standard fungicide (a.i. fenbuconazole), and sterile distilled water (SDW). Single colonies of the microbes were inoculated into 100 mL of NB and PDB (for a bacterium and yeasts, respectively), and the cultures were incubated at 25 °C on a shaking incubator at 200 rpm for 4 days. A concentration of BCAs between 107–108 CFU mL−1 was used; the cell numbers were quantified using a serial dilution technique. Apparently healthy cherries cv. of Merchant and plums cv. of Victoria were picked from commercial orchards in Kent in summer 2011 and 2012. Fruits were randomly picked from different trees and stored at cold storage at 4 °C and used within 2 (cherry) and 4 (plum) weeks of picking. Cherries were surface-sterilised using 0.5% sodium hypochlorite [52]. After air-drying in a flow cabinet for 1 h, each cherry was stabbed with a sterile 200 µL tip (one wound per cherry, ca. 2 mm depth) and 5 µL cell suspension of each candidate BCA was pipetted into wounds. Treated cherries were left to dry in a flow cabinet for 1 h and then 5 µL M. laxa spore suspension (1 × 105 spores mL−1) was pipetted into the same wound.
Treated cherries were placed in round plastic pots (5 cherries per plastic pot) lined with filter paper (7 mm diameter) soaked with sterile distilled water (SDW) to maintain high humidity. Cherries were arranged by leaving a small distance between fruit to ensure that there was no direct contact between neighbours. The lids were put on and cherries were incubated at room temperature on a bench in a completely randomised design. Rot development was assessed within 7 days after inoculation with M. laxa. When brown rot or other contamination was first noted, infected cherries were removed from the pots. The experiment was carried out twice with ten and six replicated plastic pots per treatment (i.e., 50 and 30 fruits in total, respectively).
The same set of treatments was tested similarly on plums with some modification. Plums were used without surface sterilization, and stabbed on two opposite sites on the shoulder (two wounds per plum, ca. 2 mm depth). A droplet of 10 µL (instead of 5 µL) of the cell suspension of each candidate and the spore suspension of M. laxa was pipetted into each wound. Treated plums were placed in Phytatrays (Sigma Aldrich, 2 plums per Phytatray) without filter paper and incubated at room temperature on a bench in a completely randomised design. The experiment was carried out twice with ten replicated Phytatrays per treatment (i.e., 20 fruits in total). Rot development and lesion sizes were assessed within 7 days after inoculation. The lesion size was measured using a digital Vernier caliper by assessing every lesion on the left-hand side wound of each plum. If the wound on the left-hand side was not infected but the right-hand site was, then the lesion on the right-hand side was measured. The diameter of a lesion was the average of its vertical and horizontal diameters.
3.2. Identification
3.2.1. Molecular Identification
Identification was only carried out for the two isolates that showed greatest potential as BCAs in the in vivo tests on detached fruit (the bacterium B91 and the yeast Y126). DNA of the two strains was extracted using a boiling method [53]. Molecular identification was based on ribosomal DNA sequencing. The small subunit (SSU, 1148 bp) of isolate B91 was amplified using primers 27F and 1525R [54], while the large subunit (LSU, 770 bp) and internal transcribed spacers (ITS, 518 bp) of isolate Y126 were amplified using primers LR3R, LR7 [55] and ITS1, ITS4 [56], respectively. The PCR mixture for PCR amplification of B91 (final volume 50.0 µL) contained 3.0 µL of DNA preparation, 23.8 µL of double processed sterile-filtered water (“PCR water”), 4.0 µL of 25 mM MgCl2, 4.0 µL of 2.5 mM dNTPs, 5.0 µL of thermophilic DNA Poly 10× buffer, 5.0 µL of 2 µM of each primer, and 0.2 µL of 1000 U Taq polymerase. Concentrations and volumes of the PCR mixture for PCR amplification of Y126 were similar to those used for B91 but the volumes of the DNA template and PCR water were adjusted to 2.5 and 24.3 µL, respectively.
For the SSU primers, the PCR conditions consisted of initialisation at 94 °C (4 min); 40 cycles of denaturation at 94 °C (30 s), annealing at 50 °C (30 s) and extension at 72 °C (2 min); final extension at 72 °C (10 min) and final hold at 4 °C. The PCR conditions for LSU primers were initialization at 94 °C (4 min); 40 cycles of 94 °C (1 min 30 s), 53 °C (1 min 30 s), 72 °C (2 min 30 s); 72 °C (10 min) and final hold at 4 °C. For the ITS region of Y126, the PCR conditions developed by Martini et al. [57] were used: initialisation at 94 °C (4 min); 40 cycles of 94 °C (1 min), 54 °C (1 min), 72 °C (1 min); 72 °C (10 min) and final hold at 4 °C. The PCR amplification was performed using a DNAEngine DYAD Thermocyler PTC-220 (MJ Research). PCR Products were purified using a QIAquick PCR purification kit following the standard protocol. The purified PCR products were sequenced by Qiagen Sequencing Services, Germany. The DNA sequences of B91 and Y126 were compared with sequences deposited in the GenBank DNA database using the BLAST search tool. Thirty sequences showing highest similarity values to the sequence of B91 or Y126 were selected; sequence similarity was computed using BioEdit version 7.1.3 [58].
3.2.2. Conventional Identification
Isolates B91 and Y126 were tentatively identified through the observation of colony and cell morphology. B91 was grown on NA and incubated at 25 °C for 7 days in the dark. Its morphological characters and physiology were compared to the description in Bergey’s Manual of Systematic Bacteriology [59]. Y126 was grown on PDA and MEA incubated at 25 °C for 14 days in the dark. The conventional identification of Y126 was based on the Atlas of Clinical Fungi [60]. Conidial induction of Y126 was studied using a slide culture technique [44] with incubation at room temperature under UV light for 7 days.
3.3. Temperature Effects on Growth and Mortality of B91 and Y126
3.3.1. Growth
Inoculum of B91 was transferred into 100 mL of NB and incubated at 25 °C for 4 days at 200 rpm; by day 4 the culture was in log phase. This culture was then inoculated into fresh NB by pipetting 0.5 mL of the log phase culture into Duran bottles containing 100 mL of NB. The cultures were incubated at six different temperatures (0, 5, 10, 15, 20 and 25 °C) without shaking. There were triplicate bottles for each temperature (18 bottles in total). The cultures were sampled weekly for six weeks. Before sampling, the bottle was manually shaken in order to disperse cells. A serial dilution technique was used to quantify the viable number of cells. Forty µL of liquid culture from each bottle was taken and serially diluted with 360 µL of normal saline solution (a 10-fold dilution). All samples were spread onto duplicate NA plates. Plates were incubated at 25 °C for 3 days and resultant colonies were counted using a digital counting pen. Viable cell numbers were expressed as colony forming units per ml (CFU mL−1). The experiments were carried out twice. The growth of Y126 was investigated using the same protocol except that MEA and MEB were used as growth media.
3.3.2. Survival
B91 was inoculated into six Duran bottles containing 100 mL of NB and incubated at 25 °C at 200 rpm. After 4 days growth, the liquid cultures were centrifuged at 14,000 rpm for 10 min. Supernatant was discarded and the cell pellets were washed twice with phosphate buffer solution (PBS) and pooled together. The final volume of cell suspension was adjusted to 20 mL and the concentrated cell suspension was aliquoted (1.0 mL) into each of 18 Duran bottles each containing 100 mL of PBS (i.e., no nutrients were provided) and the bottles were incubated at six different temperatures (0, 5, 10, 15, 20 and 25 °C). There were triplicate bottles per temperature. Number of viable cells was quantified weekly for six weeks using the same serial dilution method described previously. The experiments were carried out twice. The survival of Y126 was similarly investigated.
3.4. Modes of Action
3.4.1. Inhibition of Spore Germination
The effect of B91 and Y126 on spore germination of M. laxa was assessed using the method modified from Zhang et al. [47]. B91 and Y126 were grown in NB and PDB, respectively, at 25 °C for 4 days at 200 rpm. The cultures of BCAs were centrifuged at 14,000 rpm for 10 min and the supernatant was filter-sterilised using a 0.2 µM filter and transferred to a new tube while the cell pellets were washed twice with PBS and re-suspended in 5 mL of PBS (108–109 CFU mL−1). Then 0.5 mL of spore suspension of M. laxa (1 × 105 spores mL−1) was mixed with either 0.5 mL of living BCA cell suspension or cell free-culture filtrate in a screw cap tube. Sterile PDB was used as control treatment. Additional 1 mL of sterile PDB was added to provide nutrients for initial microbial growth. After mixing thoroughly with a vortex mixer, 100 µL of the mixture of each treatment was transferred onto water agar (WA) and the plates were sealed with parafilm to maintain high humidity. The tubes containing the rest of the mixture were inclined and the caps were loosened to increase surface area and air circulation. The plates of WA and the tubes of PDB were incubated at 25 °C in the dark. Twelve hours later, germination of spores of M. laxa was assessed by counting 100 spores per replicate. Spores were considered to have germinated when the germ tube was equal to or longer than the smallest diameter of spores. In total there were 10 treatments: 2 media (WA & PDB) × 2 microbes (B91 & Y126) × 2 activity (living cells and cell-broth) and two control treatments (WA & PDB). There were five replicates (plates or tubes) per treatment. The experiment was carried out twice and a completely randomized designed was used.
3.4.2. Production of Volatile Organic Compounds (VOCs)
The ability of candidate BCAs to produce VOCs against the same two strains of M. laxa used in in vitro screening (Section 3.1.2) was assessed using the co-culture technique [61]. Spread plates of B91 and Y126 were prepared on NA and PDA, respectively, by pipetting 0.5 mL of a 24-hour culture onto agar plates before spreading with a sterile triangular glass rod. The plates were incubated at 25 °C for 2 days. Plates of PDA were inoculated centrally with an agar plug cut from the edge of an actively growing mycelial culture of M. laxa on PDA using a sterile cork borer (No. 2: diameter 5 mm). The lids were removed from the Petri dishes and the PDA plate inoculated with M. laxa was inverted and placed on top of either the NA or PDA plate seeded with B91 or Y126, respectively, and the plates were taped together and incubated at 25 °C for 2 weeks. The control treatment was PDA plates inoculated with a sterile agar plug taped together with a M. laxa plate. Diameters of M. laxa colonies were measured with a digital vernier calliper. In total there were six treatments: (B91, Y126, control) × 2 M. laxa isolates. There were eight replicate (plates) per treatment, and the diameter for each replicate plate was an average from two measurements across the vertical and horizontal angles. The experiment was carried out twice.
3.5. Statistical Analysis
All statistical analyses were performed using GenStat (13th Edition) [62]; repeat experiments were treated as a blocking factor. The percentage inhibition in the in vitro screen was calculated by comparing the radius of M. laxa in the presence of antagonists with the radius of M. laxa in the control. For statistical analysis using Analysis of Variance (ANOVA), normality and homogeneity of variance were tested before statistical analysis using the Shapiro-Wilk test and Bartlett’s test, respectively. Percentage of inhibition was arcsin transformed before ANOVA and lesion size was analysed in ANOVA without transformation. Generalised linear models (GLM) were used to analyse disease incidence data (percentage of infected fruit (or wound) and germination) assuming a binomial distribution for the number of infected fruit (or wounds) and spores germinated. For the effects of temperature on microbial grown and survival, the numbers of living cells (expressed as CFU mL−1) were transformed using natural logarithms. The transformed data were subjected to Repeated Measures ANOVA. In all analyses, only when there was an overall significant treatment effect in ANOVA and GLM, pairwise treatment comparisons were then made using the least significant differences for ANOVA or the t-test for GLM at the significance level at p = 0.05.
4. Conclusions
This study shows that microbial antagonists against M. laxa can be found from indigenous sources in the UK and that that they are capable of preventing brown rot disease in controlled conditions, thus demonstrating a potential to be developed into commercial products. Two potential BCAs with strong inhibitory potential were selected: one (B91) was identified as Bacillus amyloliquefaciens/subtilis sp. and the other (Y126) Aureobasidium pullulans. They were able to grow and survive under a range of temperatures likely to be experienced under field and post-harvest storage conditions. Competition for nutrients played a major role in bioactivity of A. pullulans Y126 against M. laxa while Bacillus sp. B91 had several modes of action including production of bioactive compounds, volatile organic compounds and possibly competition for nutrients.
Acknowledgments
This project was funded under the HortLink (project No. HL0189) scheme by the Department of Environment and Rural Affairs (DEFRA). The first author (N.R.) would like to thank DEFRA and the International Office, University of Kent, for funding. John Leigh-Pemberton is thanked for allowing us to collect the cherries and plums used in in vivo studies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Borve, J.; Stensvand, A. Use of a plastic rain shield reduces fruit decay and need for fungicides in sweet cherry. Plant Dis. 2003, 87, 523–528. [Google Scholar] [CrossRef]
- Snowdon, A.L. A Colour Atlas of Post Harvest Diseases and Disorders of Fruits and Vegetables, Volume 1: General Introduction & Fruits; Wolfe Scientific Limited: Barcelona, Spain, 1990; Volume 1, pp. 218–236. [Google Scholar]
- Wherrett, A.D.; Sivasithamparam, K.; Kumar, S. Detection of possible systemic fungicide resistance in Western Australian Monilinia populations. Phytopathology 2001, 91, S95. [Google Scholar]
- Chen, F.; Liu, X.; Schnabel, G. First report of brown rot caused by Monilinia fructicola in sweet cherry in Maryland. Plant Dis. 2012, 97, 145. [Google Scholar]
- European and Mediterranean Plant Protection Organization (EPPO). Diagnostics: Monilinia fructicola. EPPO Bull. 2009, 39, 337–343. [CrossRef]
- Adaskaveg, J.E.; Forster, H.; Gubler, W.D.; Teviotdale, B.L.; Thompson, D.F. Reduced-risk fungicides help manage brown rot and other fungal diseases of stone fruit. Calif. Agric. 2005, 59, 109–114. [Google Scholar] [CrossRef]
- Malandrakis, A.; Koukiasas, N.; Veloukas, T.; Karaoglanidis, G.; Markoglou, A. Baseline sensitivity of Monilinia laxa from Greece to fenhexamid and analysis of fenhexamid-resistant mutants. Crop Prot. 2013, 46, 13–17. [Google Scholar] [CrossRef]
- Thomidis, T.; Michailides, T.; Exadaktylou, E. Contribution of pathogens to peach fruit rot in northern Greece and their sensitivity to iprodione, carbendazim, thiophanate-methyl and tebuconazole fungicides. J. Phytopathol. 2009, 157, 194–200. [Google Scholar] [CrossRef]
- Pimenta, R.S.; da Silva, J.F.M.; Buyer, J.S.; Janisiewicz, W.J. Endophytic fungi from plums (Prunus domestica) and their antifungal activity against Monilinia fructicola. J. Food Prot. 2012, 75, 1883–1889. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Jurick Ii, W.M.; Vico, I.; Peter, K.A.; Buyer, J.S. Culturable bacteria from plum fruit surfaces and their potential for controlling brown rot after harvest. Postharvest Biol. Technol. 2013, 76, 145–151. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Baykal, N. Biological control of postharvest diseases of peaches and nectarines by yeasts. J. Phytopathol. 2003, 151, 130–134. [Google Scholar] [CrossRef]
- Zhou, T.; Schneider, K.E.; Li, X. Development of biocontrol agents from food microbial isolates for controlling post-harvest peach brown rot caused by Monilinia fructicola. Int. J. Food Microbiol. 2008, 126, 180–185. [Google Scholar] [CrossRef]
- Yang, H.; Zhao, X.; Zhao, X.; Liu, Z.; Yang, H.Q.; Zhao, X.M.; Zhao, X.Y.; Liu, Z.P. Antifungal mechanism against monilinia fruticola and stability of the antagonistic substance from bacterium strain CE. J. Fruit Sci. 2011, 28, 204–208. [Google Scholar]
- Bonaterra, A.; Mari, M.; Casalini, L.; Montesinos, E. Biological control of Monilinia laxa and Rhizopus stolonifer in postharvest of stone fruit by Pantoea agglomerans EPS125 and putative mechanisms of antagonism. Int. J. Food Microbiol. 2003, 84, 93–104. [Google Scholar] [CrossRef]
- Mari, M.; Torres, R.; Casalini, L.; Lamarca, N.; Mandrin, J.F.; Lichou, J.; Larena, I.; de Cal, M.A.; Melgarejo, P.; Usall, J. Control of post-harvest brown rot on nectarine by Epicoccum nigrum and physico-chemical treatments. J. Sci. Food Agric. 2007, 87, 1271–1277. [Google Scholar] [CrossRef]
- Karabulut, O.A.; Baykal, N. Evaluation of the use of microwave power for the control of postharvest diseases of peaches. Postharvest Biol. Technol. 2002, 26, 237–240. [Google Scholar] [CrossRef]
- Casals, C.; Teixido, N.; Vinas, I.; Silvera, E.; Lamarca, N.; Usall, J. Combination of hot water, Bacillus subtilis CPA-8 and sodium bicarbonate treatments to control postharvest brown rot on peaches and nectarines. Eur. J. Plant Pathol. 2010, 128, 51–63. [Google Scholar] [CrossRef]
- Guijarro, B.; Melgarejo, P.; Torres, R.; Lamarca, N.; Usall, J.; de Cal, A. Penicillium frequentans population dynamics on peach fruits after its applications against brown rot in orchards. J. Appl. Microbiol. 2008, 104, 659–671. [Google Scholar] [CrossRef]
- Larena, I.; de Cal, A.; Melgarejo, P. Enhancing the adhesion of Epicoccum nigrum conidia to peach surfaces and its relationship to the biocontrol of brown rot caused by Monilinia laxa. J. Appl. Microbiol. 2010, 109, 583–593. [Google Scholar]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Selection and evaluation of new antagonists for their efficacy against postharvest brown rot of peaches. Postharvest Biol. Technol. 2010, 55, 174–181. [Google Scholar] [CrossRef]
- European Union (EU). Regulation (EC) No 1007/2009 of the European Parliament and of the Council of 16 September 2009. Off. J. Eur. Community 2009, L309, 1–50.
- European Union (EU). Council Directive 91/414/EEC. Off. J. Eur. Community 1991, L230, 1–32.
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A Review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Gueldner, R.C.; Reilly, C.C.; Pusey, P.L.; Costello, C.E.; Arrendale, R.F.; Cox, R.H.; Himmelsbach, D.S.; Crumley, F.G.; Cutler, H.G. Isolation and identification of iturins as antifungal peptides in biological control of peach brown rot with Bacillus subtilis. J. Agric. Food Chem. 1988, 36, 366–370. [Google Scholar] [CrossRef]
- Pusey, P.L.; Wilson, C.L. Postharvest biological control of stone fruit brown rot by Bacillus subtilis. Plant Dis. 1984, 68, 753–756. [Google Scholar]
- Pusey, P.L.; Wilson, C.L.; Hotchkiss, M.W.; Fraklin, J.D. Compatibility of Bacillus subtilis for postharvest control of peach brown rot with commercial fruit waxes, dicloran and cold storage conditions. Plant Dis. 1986, 70, 587–590. [Google Scholar] [CrossRef]
- CRD Chemicals Regulation Directorate: Biopesticides. Available online: http://www.pesticides.gov.uk/guidance/industries/pesticides/user-areas/biopesticides-home (accessed on 19th October 2013).
- Wang, X.; Li, G.; Jiang, D.; Huang, H.-C. Screening of plant epiphytic yeasts for biocontrol of bacterial fruit blotch (Acidovorax avenae subsp. citrulli) of hami melon. Biol. Control 2009, 50, 164–171. [Google Scholar] [CrossRef]
- Raspor, P.; Miklic-Milek, D.; Avbelj, M.; Cadez, N. Biocontrol of grey mould disease on grape caused by Botrytis cinerea with autochthonous wine yeasts. Food Technol. Biotechnol. 2010, 48, 336–343. [Google Scholar]
- Whipps, J.M. Effect of media on growth and interactions between a range of soil-borne glasshouse pathogens and antagonistic fungi. New Phytol. 1987, 107, 127–142. [Google Scholar] [CrossRef]
- Magan, N. Ecophysiology of Biocontrol Agents for Improved Completence in the Phyllophere. In Microbial Ecology of Aerial Plant Surfaces; Bailey, M.J., Lilley, A.K., Timms-Wilson, T.M., Spencer-Phillips, P.T.N., Eds.; CAB International: Wallingford, UK, 2006; pp. 149–164. [Google Scholar]
- Janisiewicz, W.J.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with nectarines and their potential for biological control of brown rot. Yeast 2010, 27, 389–398. [Google Scholar] [CrossRef]
- Mignard, S.; Flandrois, J.P. 16S rRNA sequencing in routine bacterial identification: A 30-month experiment. J. Microbiol. Methods 2006, 67, 574–581. [Google Scholar] [CrossRef]
- Alvarez, F.; Castro, M.; Principe, A.; Borioli, G.; Fischer, S.; Mori, G.; Jofre, E. The plant-associated Bacillus amyloliquefaciens strains MEP218 and ARP23 capable of producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in biocontrol of sclerotinia stem rot disease. J. Appl. Microbiol. 2012, 112, 159–174. [Google Scholar] [CrossRef]
- Chun, J.; Bae, K.S. Phylogenetic analysis of Bacillus subtilis and related taxa based on partial gyrA gene sequences. Antonie Van Leeuwenhoek 2000, 78, 123–127. [Google Scholar] [CrossRef]
- O’Donnell, A.G.; Norris, J.R.; Berkeley, R.C.W.; Claus, D.; Kaneko, T.; Logan, N.A.; Nozaki, R. Characterization of Bacillus subtilis, Bacillus pumilus, Bacillus licheniformis, and Bacillus amyloliquefaciens by pyrolysis gas-liquid chromatography, deoxyribonucleic acid-deoxyribonucleic acid hybridization, biochemical tests, and API systems. Int. J. Syst. Bacteriol. 1980, 30, 448–459. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; p. 252. [Google Scholar]
- Klein, W.; Weber, M.H.W.; Marahiel, M.A. Cold shock response of Bacillus subtilis: Isoleucine-dependent switch in the fatty acid branching pattern for membrane adaptation to low temperatures. J. Bacteriol. 1999, 181, 5341–5349. [Google Scholar]
- D’Amico, S.; Collins, T.; Marx, J.-C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Warth, A.D. Relationship between the heat resistance of spores and the optimum and maximum growth temperatures of Bacillus species. J. Bacteriol. 1978, 134, 699–705. [Google Scholar]
- Tokuda, Y.; Ano, T.; Shoda, M. Survival of Bacillus subtilis NB22 and its transformant in soil. Appl. Soil Ecol. 1995, 2, 85–94. [Google Scholar] [CrossRef]
- Budde, I.; Steil, L.; Scharf, C.; Volker, U.; Bremer, E. Adaptation of Bacillus subtilis to growth at low temperature: A combined transcriptomic and proteomic appraisal. Microbiology 2006, 152, 831–853. [Google Scholar] [CrossRef]
- Sugumaran, K.R.; Gowthami, E.; Swathi, B.; Elakkiya, S.; Srivastava, S.N.; Ravikumar, R.; Gowdhaman, D.; Ponnusami, V. Production of pullulan by Aureobasidium pullulans from Asian palm kernel: A novel substrate. Carbohydr. Polym. 2013, 92, 697–703. [Google Scholar] [CrossRef]
- Zalar, P.; Gostincar, C.; de Hoog, G.S.; Ursic, V.; Sudhadham, M.; Gunde-Cimerman, N. Redefinition of Aureobasidium pullulans and its varieties. Stud. Mycol. 2008, 61, 21–38. [Google Scholar] [CrossRef]
- Yanez-Mendizabal, V.; Zeriouh, H.; Vinas, I.; Torres, R.; Usall, J.; de Vicente, A.; Perez-Garcia, A.; Teixido, N. Biological control of peach brown rot (Monilinia spp.) by Bacillus subtilis CPA-8 is based on production of fengycin-like lipopeptides. Eur. J. Plant Pathol. 2012, 132, 609–619. [Google Scholar] [CrossRef]
- Zeriouh, H.; Romero, D.; Garcia-Gutierrez, L.; Cazorla, F.M.; de Vicente, A.; Perez-Garcia, A. The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol. Plant Microbe Interact. 2011, 24, 1540–1552. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Efficacy of the antagonist Aureobasidium pullulans PL5 against postharvest pathogens of peach, apple and plum and its modes of action. Biol. Control 2010, 54, 172–180. [Google Scholar] [CrossRef]
- Castoria, R.; de Curtis, F.; Lima, G.; Caputo, L.; Pacifico, S.; de Cicco, V. Aureobasidium pullulans (LS-30) an antagonist of postharvest pathogens of fruits: Study on its modes of action. Postharvest Biol. Technol. 2001, 22, 7–17. [Google Scholar] [CrossRef]
- Vero, S.; Garmendia, G.; Belen Gonzalez, M.; Fernanda Garat, M.; Wisniewski, M. Aureobasidium pullulans as a biocontrol agent of post-harvest pathogens of apples in Uruguay. Biocontrol Sci. Technol. 2009, 19, 1033–1049. [Google Scholar] [CrossRef]
- Bencheqroun, S.K.; Bajji, M.; Massart, S.; Bentata, F.; Labhilili, M.; Achbani, H.; El Jaafari, S.; Jijakli, M.H. Biocontrol of blue mold on apple fruits by Aureobasidium pullulans (strain Ach 1–1): In vitro and in situ evidence for the possible involvement of competition for nutrients. Commun. Agric. Appl. Biol. Sci. 2006, 71, 1151–1157. [Google Scholar]
- Swadling, I.R.; Jeffries, P. Isolation of microbial antagonists for biocontrol of grey mould disease of strawberries. Biocontrol Sci. Technol. 1996, 6, 125–136. [Google Scholar] [CrossRef]
- Xu, X.-M.; Bertone, C.; Berrie, A. Effects of wounding, fruit age and wetness duration on the development of cherry brown rot in the UK. Plant Pathol. 2007, 56, 114–119. [Google Scholar]
- Fang, H.; Hedin, G. Rapid screening and identification of methicillin-resistant Staphylococcus aureus from clinical samples by selective-broth and real-time PCR assay. J. Clin. Microbiol. 2003, 41, 2894–2899. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–147. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cyptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar]
- White, T.J.; Bruns, T.L.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogentics. In A Guide to Molecular Methods and Applications; Innis, M.A., Gelfand, D.H., Snisky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Martini, M.; Musetti, R.; Grisan, S.; Polizzotto, R.; Borselli, S.; Pavan, F.; Osler, R. DNA-Dependent detection of the grapevine fungal endophytes Aureobasidium pullulans and Epicoccum nigrum. Plant Dis. 2009, 93, 993–998. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Logan, N.A.; de Vos, P. Bacillus. In Bergey’s Manual of Systematic Bacteriology: The Firmicutes, 2nd ed.; De Vos, P., Garrity, G., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; Volume 3, pp. 21–128. [Google Scholar]
- De Hoog, G.S.; Guarro, J.; Gene, J.; Figueras, M.J. Atlas of Clinical Fungi; Centraalbureau voor Schimmecultures: Utrecht, The Netherlands, 2000; p. 1126. [Google Scholar]
- Swadling, I. Biological Control of Botrytis cinerea in Strawberries. Ph.D. Thesis, University of Kent, Canterbury, UK, July 1994. [Google Scholar]
- Payne, R.; Soutar, D.; Baird, D.; Thompson, R.; Gilmour, A.R.; Todd, A.D.; Harding, S.A.; Wilson, G.T.; Lane, P.W.; Webster, R.; Murray, D.; Welham, S. The Guide to GenStat, Part 2: Statistics; VSN International Ltd.: Hemel Hempstead, UK, 2000; p. 782. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).