1. Introduction
Cabbage (
Brassica oleracea var.
capitata) is one of the most important vegetables grown in tropical and sub-tropical Africa. In Kenya, the farming and marketing of cabbage provides a secure source of continuous income to support family needs. Cabbage production allows small farms to remain financially viable, especially in the rapid growing peri-urban farming sector [
1]. Cabbage production in Kenya has, however, shown mixed trends over the past four years. The area under cabbage production was 14,783 hectares in 2009, 18,477 hectares in 2010 and declined to 12,832 hectares in 2011 [
2]. Total yields showed a similar trend at about 510,000 metric tons in 2009, 600,000 metric tons in 2010 and down to 523,000 metric tons in 2011 [
3] against an annual population growth of about 2.6%. High cost of seed, poor seed germination and seedling performance in the nursery are major constraints to many commercial nursery owners and cabbage growers in Kenya. These constraints substantially increase the production cost, rendering cabbage production non-profitable and, consequently, an unsustainable venture. Although, technologies for enhanced seedling performance exist in many developed countries, the cost of most of these technologies is prohibitive to the many small-scale growers of the tropics.
Seed germination and early seedling vigor are important attributes that impact the success of many crops, especially in the tropics. The germination of most seeds is affected by environmental factors, such as temperature and moisture [
4,
5]. Temperature affects the capacity for germination and the rate of germination of seeds, while soil moisture is essential for germination [
6]. Insufficient moisture results in poor germination. The moisture content of the growing media influences early seedling survival. Relative humidity is also of considerable importance in influencing seedling growth of most annuals [
7].
Apart from temperature and moisture, successful transplant production in the tropics is also constrained by pest and disease injury. Aphids and leaf miners have been documented among major pests of cabbage seedlings in Kenya [
8]. The sucking of seedling sap by aphids creates a lack of vigor in the plant marked by decreased growth rates, mottled leaves, yellowing, stunted growth, curled leaves, browning and wilting. All of these symptoms affect the physiology of seedlings and the subsequent plant growth. Aphids may also transmit diseases, like cabbage mosaic virus, to their hosts. In addition, honeydew, a sugary liquid secreted by aphids as they feed, attracts fungi, which can damage the plants [
9,
10]. Leaf miner larvae, on the other hand, burrow through the leaves, resulting in destruction of the leaf’s mesophyll. Leaf mining depresses the level of photosynthesis in the plant, thereby reducing plant vigor and growth. An indirect effect of feeding by aphids and leaf miners is that the wounds created can serve as avenues for the entry of bacterial and fungal pathogens.
Information on sustainable seedling production is lacking in many sub-Saharan countries. Little is known on cost-effective and environmental friendly nursery production technologies with the potential of increasing transplant numbers and quality through microclimate modification and insect pest exclusion. The use of net coverings in crop production has been shown to mitigate extreme climatic fluctuations and improve canopy vitality [
11]. At the same time, net covers provide physical exclusion for insects, thereby reducing the incidence of direct crop damage and insect-transmitted viral diseases [
12]. This in turn would reduce the farmer’s dependence on chemical pest control. The objective of this study was to assess the effect of EFN covering on cabbage seed germination, seedling physiology, insect pest population and seedling growth and quality.
2. Materials and Methods
2.1. Site Description
The study was conducted over a span of two seasons (March–April 2011 and September–October 2011) at the Horticulture Research and Teaching Field, Egerton University, Kenya. The area lies at altitude 2238 m, latitude 0°23' S and longitude 35°35' E. The average maximum and minimum temperatures range between 19 to 22 °C and 5 to 8 °C, respectively. The area receives an annual rainfall of 1200 to 1400 mm (Egerton Metrological Station, 2009).
2.2. Experimental Design and Treatments
The study was laid using a randomized complete block design with two treatments replicated five times. Individual blocks comprised of two beds each measuring 1 by 2 m. One bed in each block was protected with fine mesh nets (AgroNet 0.4 mm pore diameter) (A to Z Textile Mills Ltd., Arusha, Tanzania), while the other bed was mulched with dry grass until germination, as is the standard practice by most farmers in the region, to serve as the control. Each block measured 4.5 by 1 m with a 1 m buffer separation. Ordinary mild steel was used to make arches to support the nets. Hoops used were 1 m wide and 0.3 m high after being secured 0.2 m into the soil. Two such arches were used per plot mounted in the 1 m opposite ends of the plot.
2.3. Plant Material, Planting, and Seedling Maintenance
Gloria F1 hybrid cabbage seed (Kenya Seed Co., Kitale, Kenya) was used in this study. Gloria F1 is one of the most popular cabbage varieties in Kenya, being preferred for its high yields, superior taste, resistance to splitting upon maturity and excellent storage qualities. Seeds were sown in rows spaced 10 cm apart at a distance of one centimeter within the rows for a total of 2,000 seeds per nursery bed. After planting, the beds were watered thoroughly and the nets mounted on the protected beds. Thereafter, nursery management practices, like weeding and watering, were carried out uniformly to all plots. Weeding was done once using hand hoes two weeks after emergence, while watering was done every morning using watering cans during dry spells.
2.4. Data Collection
Data collection commenced at three days after planting and was continued for a period of four weeks. The variables measured included microclimate modification, days to emergence, percent emergence, seedling growth, leaf chlorophyll content, leaf stomatal conductance, insect pest populations and number of diseased seedlings. Based on the percent emergence and recommended spacing for Gloria F1 cabbage, the seed requirement and cost per unit area was also estimated.
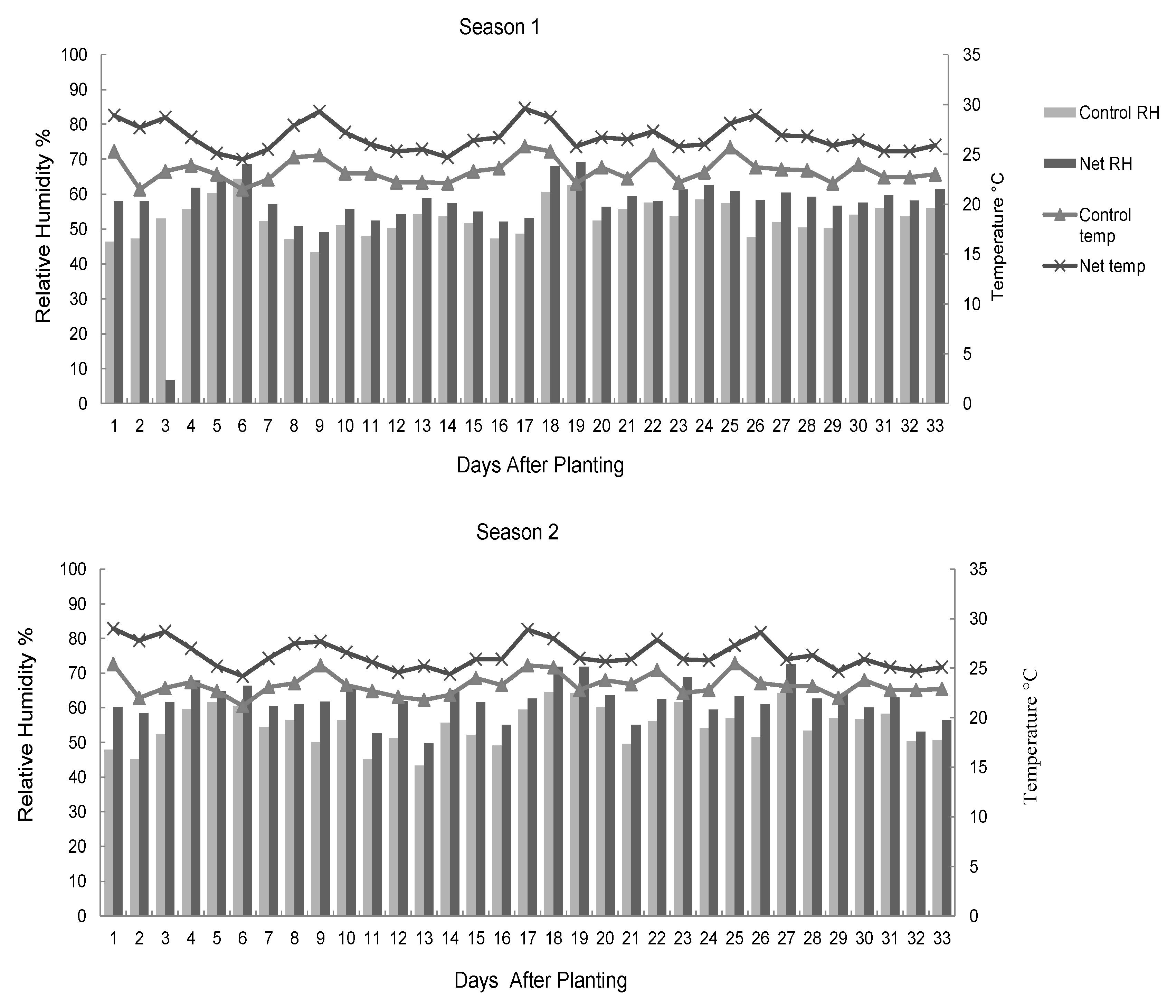
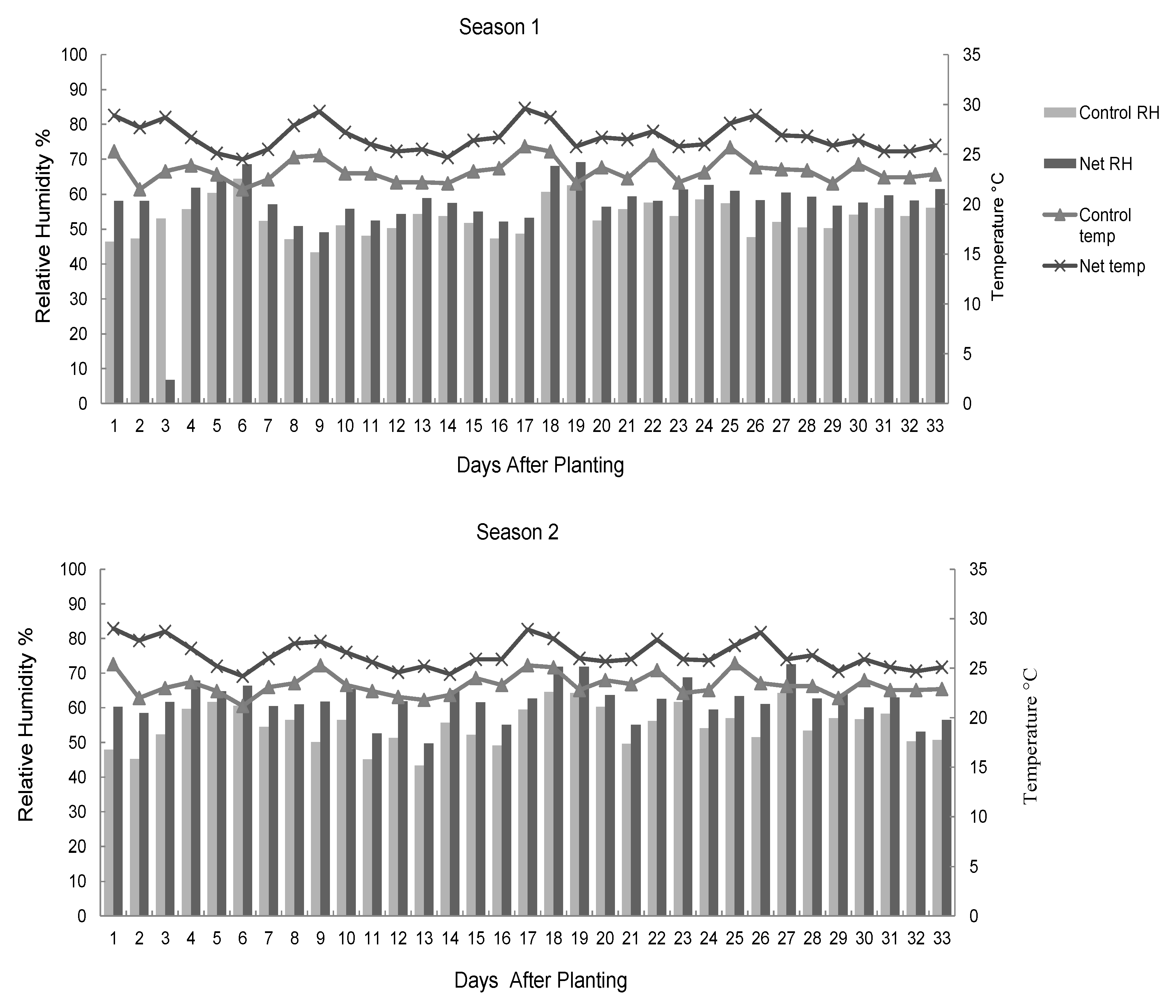
The nursery microclimate was monitored by collecting data on temperatures and relative humidity three times every day (8.00 am, noon, and 4.00 pm) from each plot. A thermo-hygrometer (HM9, Shanghai Precision and Scientific Instrument Co., Shanghai, China) was used to measure the temperature and relative humidity. The daily mean temperature and relative humidity were then obtained as the average of the three readings collected for each variable in each day.
The days to emergence was determined as the mean of the number of days from sowing to first seedling emergence for each treatment in the individual blocks. For percent emergence, the total number of emerged seedlings was counted on the seventh, ninth and eleventh day after planting (DAP) and germination percentages computed [
13]. To estimate seedling growth
, leaves were counted and plant height measured weekly using a ruler from 20 tagged cabbage seedlings in each plot. At the same time, the collar diameter of the 20 tagged seedlings was also measured using a standard calliper (Vernier calliper Series 530 Standard model, Mitutoyo America Corporation, Aurora, IL, USA) at the stem base just above the ground level.
Leaf chlorophyll content estimates were taken in chlorophyll concentration index units (CCI) from a recently fully expanded leaf of each of the 20 tagged cabbage seedling of each treatment using a chlorophyll meter (CCM-200 Plus, Opti-Sciences, Inc. NH, USA). These data were collected weekly between 10 am and 11 am from two weeks after emergence (WAE) to the end of the experiment. Stomatal conductance was also taken from the same leaf used to estimate the chlorophyll content using a steady state leaf porometer (SC-1; Decagon Devices, Pullman, WA, USA). Stomatal conductance readings were taken directly from the leaf porometer and recorded in µmol m−1 sec−2.
The number of aphids and leaf miners were assessed weekly from two leaves (one from the upper canopy and one from the lower canopy) from each of the 20 tagged seedlings. The mean number of each pest per plant was then computed in individual treatments. The total number of seedlings showing disease symptoms was also recorded for each treatment. The nets were open only during data collection times.
Based on the percentage emergence and recommended spacing for Gloria F1 hybrid, the amount of seed required to establish a hectare of cabbage and the cost reduction estimate for the purchase of seed were also computed.
2.5. Data Analysis
Data for the two seasons were pooled and subjected to analysis of variance using SAS (version 9.1; SAS Institute, Cary, NC) software. The statistical model; Yij = μ + ρi + αj + εij was used: where i = 1, 2, 3, 4, 5, j = 1, 2, Yij—Cabbage seedling response, μ—grand mean, ρi—ith blocking effect, αj—jth treatment effect, εij—random error component. Where the F-test was significant at P ≤ 0.05, mean comparison was done using Least Significant Difference at the same probability level.
4. Discussion and Conclusions
Starting cabbage seeds in the nets proved a beneficial and sustainable technology in cabbage transplant production. When compared with the conventional farmer practice, netting resulted in early emergence, as well as higher germination percentages. Cabbage seeds are small and consequently have fewer reserves to sustain a developing seedling before forming a photosynthetic surface. Conditions that make it difficult for the cotyledons of developing seedlings to quickly reach the surface are, therefore, likely to have an adverse effect on the subsequent emergence and development of a seedling [
6]. The chemical and metabolic processes, which enable downward elongation of the radicle and upward elongation of the hypocotyl, tend to be faster in warmer than in cool conditions. Temperature rises within the optimal range of any given seed will therefore tend to trigger faster germination and emergence. Adequate moisture, on the other hand, is necessary for activation of important enzymes involved in germination, which break down stored reserves in the cotyledons to provide energy for the growing points [
14]. In the current study, temperatures and moisture conditions remained higher in the nets compared to the control throughout the study period. The significant differences in germination attributes between the control and the net protected seedlings in the current study attest to the importance of the microclimate of a seed in the seedling germination process. Sub-optimal conditions negatively affect early germination and seedling emergence and
vice versa. Similar to the findings of our study, Song
et al., [
15] observed better germination of chieh-qua (
Benincasa hispida var.
chieh-qua) seeds under shade net covering.
Upon seedling emergence, better growth was also observed in the nets compared to the control treatment. Fajinmi and Fajinmi [
16] similarly recorded increased plant height and plant bearing capacity in okra (
Abelmoschus esculentus) grown under netting compared to control plants. Adequate moisture, besides enhancing nutrient uptake and activating enzymatic activity, is important in cell division and expansion. The differences in soil moisture content and relative humidity between the net covered and the control treatment could possibly account for the differences in seedling growth between the treatments where seedlings in the nets ended up with more leaves and higher stem length and collar diameter compared to the control seedlings. Reduced stomatal conductance and chlorophyll content observed under the control treatment could also be attributed to the low soil moisture conditions in this treatment and possibly reduced nutrient uptake. Stomatal conductance decreases in response to low moisture as a result of which the supply of CO
2 to chloroplasts reduces [
17]. Low stomatal conductance of the control seedlings together with the low chlorophyll content recorded could have lowered photosynthesis of the seedlings, thereby affecting their growth rate.
Net covering has been used as a physical and visual barrier excluding pests from growing plants [
18,
19]. As a result, their feeding and mating habits are distracted, thereby lowering their population. In their review on UV absorbing barriers, Antignus and Ben-Yakir [
20] showed that the light spectrum plays an important role in aspects of insect behavior, including navigation and orientation. In the current study, growing seedlings in white nets significantly reduced populations of leaf miners and aphids, probably because of the brightness of the nets, which could have served as a visual barrier, besides the physical barrier provided. Similar findings have been reported in the control of red pumpkin beetle (
Aulacophora foveicollis) where mosquito net covering provided an effective barrier against the pest [
21]. Leaf miners feed on leafy material, while aphids suck on the sap with both effects negatively affecting the photosynthetic turnover and subsequent growth and normal functioning of the plant. In addition, aphids have been documented as vectors for viruses, like the cabbage mosaic virus [
9,
10], while wounds created can serve as avenues for secondary infection. Slowed emergence together with low moisture may also increase the exposure of a developing seedling to a range of diseases, which could be injurious to plants [
22]. Such a scenario could have rendered the control seedlings in the current study more vulnerable to disease attack compared to the seedlings in the nets. The damp conditions at the seedling bases created by the grass mulch used as part of the control treatment may also have contributed to the higher disease incidences recorded for this treatment.
The use of net covers in seedling production is still comparatively low in Africa compared to the developed world. This could be attributed to the fact that over 80% of vegetable growing is by rural small holder farmers who are not familiar with the technology, which is relatively new in many African countries. Even though some few commercial nurseries exist that employ the technology in seedling production, most small holder farmers do not purchase seedlings from them, because of high seedling and transportation costs. As such, many growers have continued with their traditional nursery practices where cabbage seedlings are started in the open either mulched or in shaded patches that are subject to temperature fluctuations and pest attack. This ultimately affects seedling germination and the length of time the seedlings take in the nursery before they are ready for transplanting.
Cabbage seedlings are ready for transplanting when they have at least four true leaves. While seedlings in the nets were able to attain 4.1 true leaves at four weeks after emergence in the current study, seedlings in the open only had 2.8 leaves by this time. This would imply that a conventional cabbage seedling grower would have to retain the seedlings for a longer period in the nursery before they can achieve the recommended attributes for transplanting. Based on these observations, the use of netting by commercial nursery owners can, therefore, potentially increase the land cropping index, as well as the annual sales of the nursery seedling growers. Netting also presented overriding advantages in terms of seed cost reduction owing to a higher emergence percentage and, as such, a potential reduction on the cost of seed that a grower would incur on a given unit area of land. Together with the enhanced seedling growth in the nets, this technology presents an amicable solution to sustainable and profitable cabbage transplant and/or crop production in sub-Saharan Africa, whose institutions and other practitioners need to play a key role in disseminating. Future studies should look at the full economic analysis of the production systems with the cost of the nets included when they become commercially available.



{kind=link}
{kind=link}
{kind=link}
