
Zea mays-Derived Protein Hydrolysate and Diverse Application Modes Differentially Compose Crop Production and Fruit Quality of Strawberry Cultivated Under Tunnel


 , ,
, ,  , ,
, , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Trial Field and Vegetal Material
2.2. Biostimulant Treatments
2.3. Experimental Design and Statistics
2.4. Yield, Nutritional and Functional Traits, Mineral Profile and NUE
3. Results
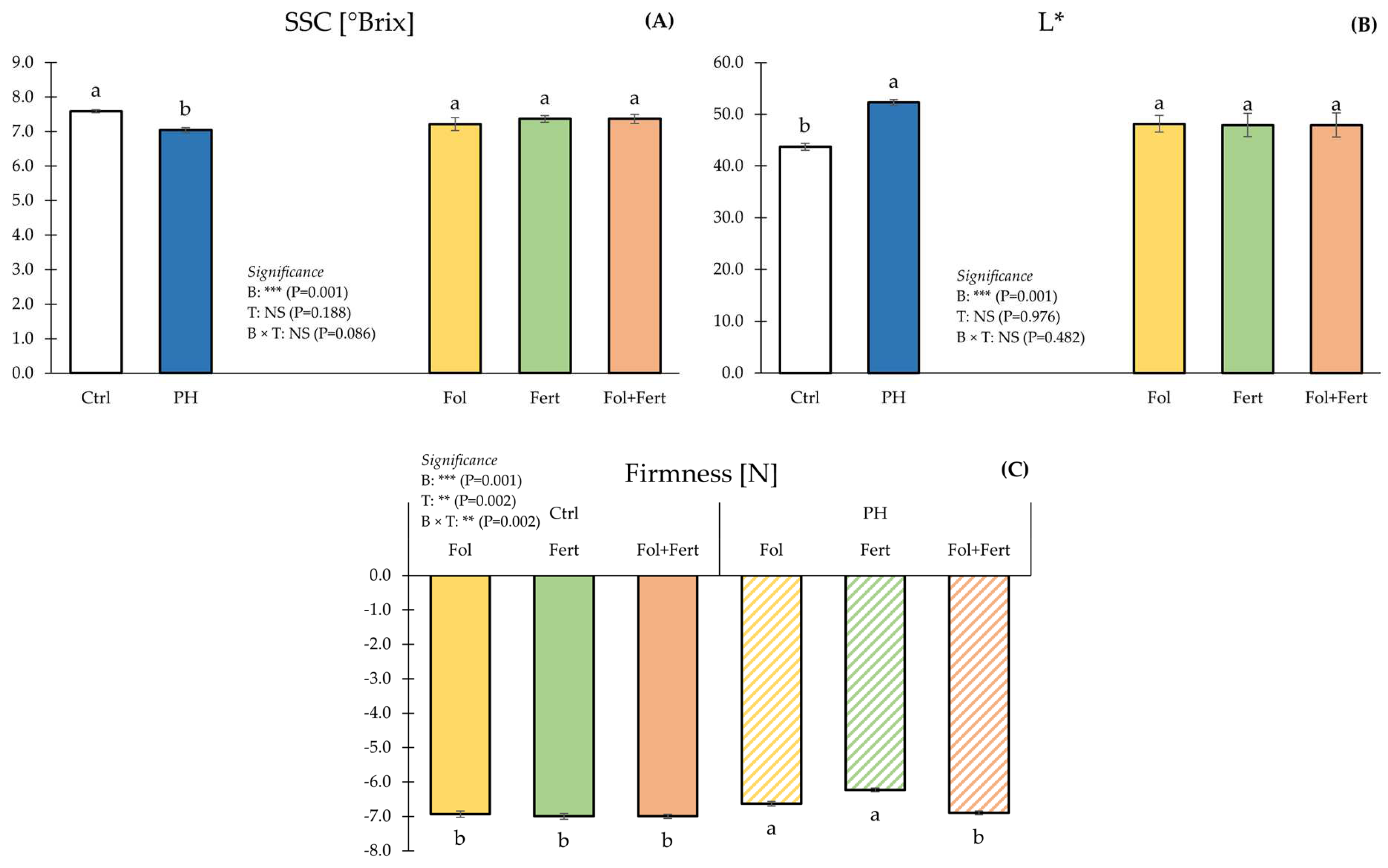
3.1. Crop Productivity and Fruit Quality
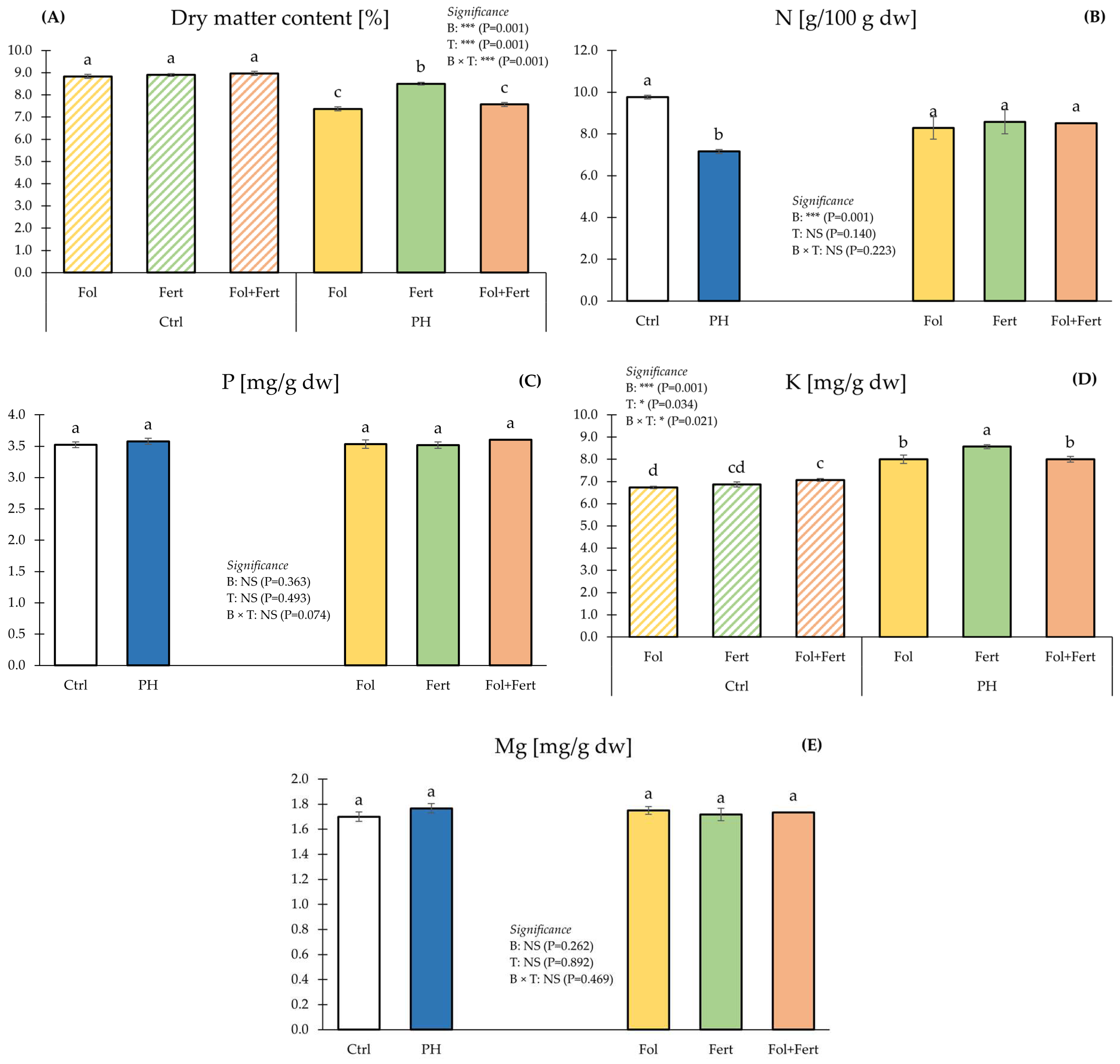
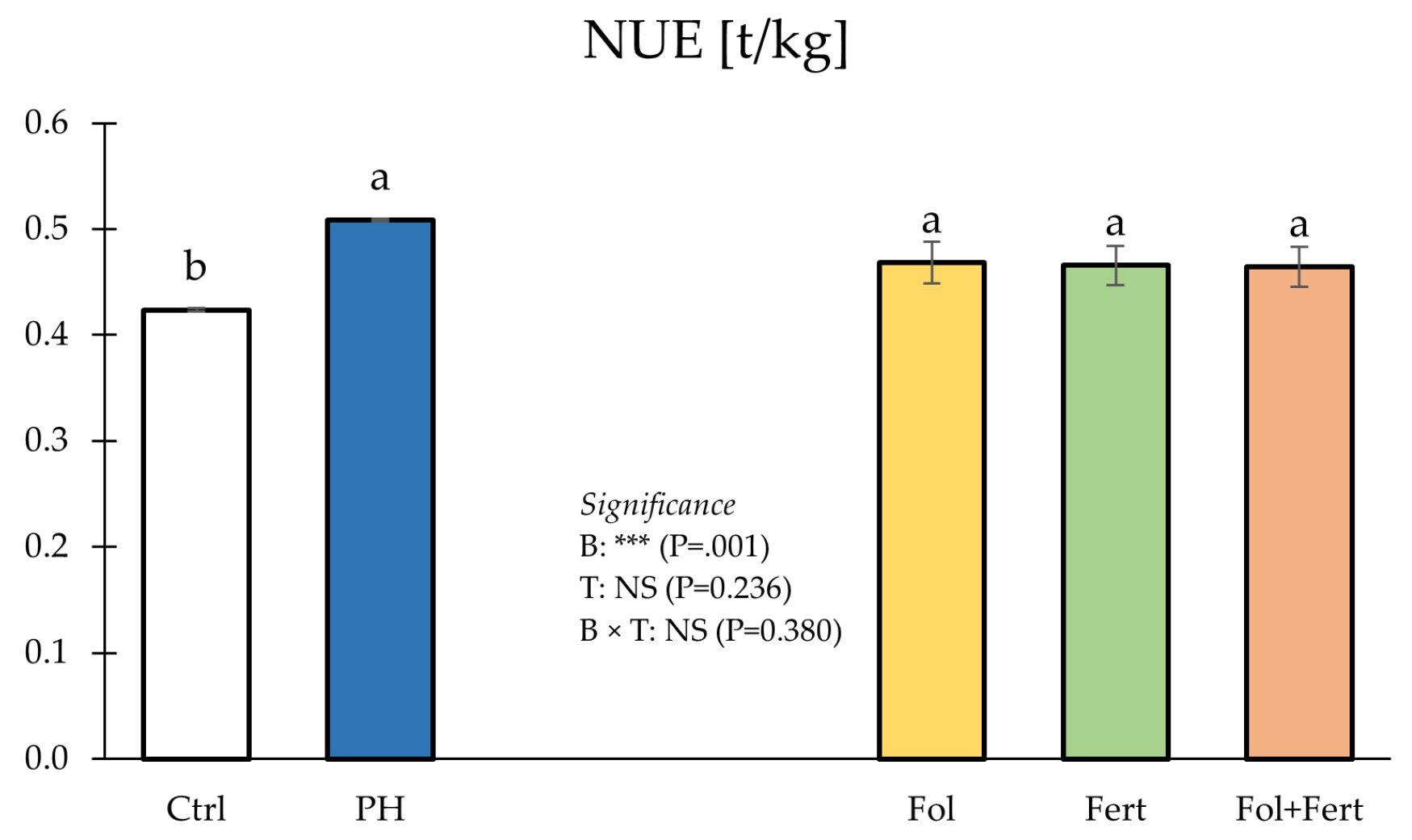
3.2. Fuit Dry Matter, Mineral Profile and NUE
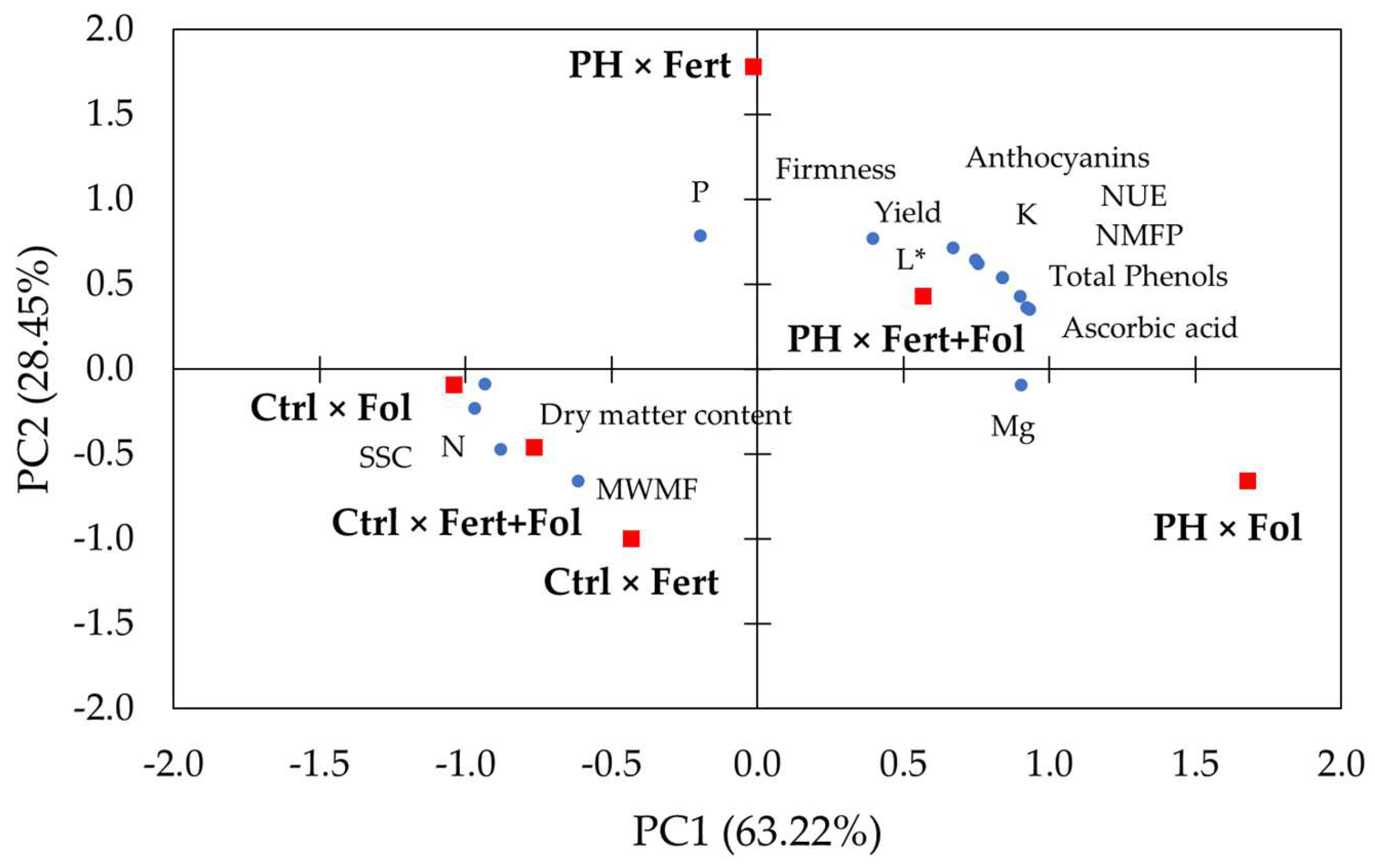
3.3. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Biswas, M.K.; Islam, R.; Hossain, M. Micropropagation and field evaluation of strawberry in Bangladesh. J. Agric. Technol. 2008, 4, 167–182. [Google Scholar]
- Sarıdaş, M.A.; Ağ, E.; Akbaş, F.C.; Akyıldiz, A.; Kargı, S.P. Comparison of superior bred strawberry genotypes with popular cultivars in terms of fruit bioactive compounds during the wide harvest dates. S. Afr. J. Bot. 2022, 147, 142–152. [Google Scholar] [CrossRef]
- FAOSTAT Database. 2023. Available online: https://www.fao.org/faostat/en/#home (accessed on 24 March 2025).
- ISTAT. Available online: https://www.istat.it/ (accessed on 24 February 2025).
- Tesi, R. Colture Protette; Edagricole: Milan, Italy, 2008. [Google Scholar]
- Romero-Gámez, M.; Suárez-Rey, E.M. Environmental footprint of cultivating strawberry in Spain. Int. J. Life Cycle Assess. 2020, 25, 719–732. [Google Scholar] [CrossRef]
- Vultaggio, L.; Allevato, E.; Consentino, B.B.; Bellitto, P.; Napoli, S.; Cannata, C.; Ntatsi, G.; Vasto, S.; Baldassano, S.; La Bella, S.; et al. Joint Action of Trichoderma atroviride and a Vegetal Derived-Protein Hydrolysate Improves Performances of Woodland Strawberry in Italy. Horticulturae 2024, 10, 459. [Google Scholar] [CrossRef]
- Vultaggio, L.; Ciriello, M.; Campana, E.; Bellitto, P.; Consentino, B.B.; Rouphael, Y.; Colla, G.; Mancuso, F.; La Bella, S.; Napoli, S.; et al. Single or Blended Application of Non-Microbial Plant-Based Biostimulants and Trichoderma atroviride as a New Strategy to Enhance Greenhouse Cherry Tomato Performance. Plants 2024, 13, 3048. [Google Scholar] [CrossRef]
- Consentino, B.B.; Vultaggio, L.; Iacuzzi, N.; La Bella, S.; De Pasquale, C.; Rouphael, Y.; Ntatsi, G.; Virga, G.; Sabatino, L. Iodine Biofortification and Seaweed Extract-Based Biostimulant Supply Interactively Drive the Yield, Quality, and Functional Traits in Strawberry Fruits. Plants 2023, 12, 245. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Rotino, G.L.; Palazzolo, E.; Mennella, G.; D’Anna, F. Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ‘Scarlatti’. Agronomy 2019, 9, 223. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed]
- Du Jardin, P. Plant biostimulants: Definition; concept; main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Clément, J.; Delisle-Houde, M.; Nguyen, T.T.A.; Dorais, M.; Tweddell, R.J. Effect of Biostimulants on Leafy Vegetables (Baby Leaf Lettuce and Batavia Lettuce) Exposed to Abiotic or Biotic Stress under Two Different Growing Systems. Agronomy 2023, 13, 879. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; Baldassano, S.; De Pasquale, C.; Ntatsi, G. Ecklonia maxima-derivate seaweed extract supply as mitigation strategy to alleviate drought stress in chicory plants. Sci. Hortic. 2023, 312, 111856. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Fernández, V.; Eichert, T. Uptake of hydrophilic solutes through plant leaves: Current state of knowledge and perspectives of foliar fertilization. Crit. Rev. Plant Sci. 2009, 28, 36–68. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activities of two protein hydrolysates on the growth and nitrogen metabolism in maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Matsumiya, Y.; Kubo, M. Soybean peptide: Novel plant growth promoting peptide from soybean. In Soybean and Nutrition; El-Shemy, R.H., Ed.; Tech Europe Publisher: Giza, Egypt, 2011; pp. 215–230. [Google Scholar]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 96, 28–38. [Google Scholar] [CrossRef]
- Tesi, R. Orticoltura Mediterranea Sostenibile; Pàtron Editore: Bologna, Italy, 2010. [Google Scholar]
- Doumett, S.; Fibbi, D.; Cincinelli, A.; Giordani, E.; Nin, S.; Del Bubba, M. Comparison of Nutritional and Nutraceutical Properties in Cultivated Fruits of Fragaria vesca L. Produced in Italy. Food Res. Int. 2011, 44, 1209–1216. [Google Scholar] [CrossRef]
- Rabino, I.; Mancinelli, A.L. Light, Temperature, and Anthocyanin Production 1. Plant Physiol. 1986, 81, 922–924. [Google Scholar] [CrossRef]
- Fogg, D.N.; Wilkinson, A.N. The colorimetric determination of phosphorus. Analyst 1958, 83, 406–414. [Google Scholar] [CrossRef]
- Choi, S.; Colla, G.; Cardarelli, M.; Kim, H.J. Effects of plant-derived protein hydrolysates on yield, quality, and nitrogen use efficiency of greenhouse grown lettuce and tomato. Agronomy 2022, 12, 1018. [Google Scholar] [CrossRef]
- Martín, M.H.J.; Ángel, M.M.M.; Aarón, S.L.J.; Israel, B.G. Protein hydrolysates as biostimulants of plant growth and development. In Biostimulants: Exploring Sources and Applications; Springer Nature: Singapore, 2022; pp. 141–175. [Google Scholar]
- Heinemann, B.; Hildebrandt, T.M. The role of amino acid metabolism in signaling and metabolic adaptation to stress-induced energy deficiency in plants. J. Exp. Bot. 2021, 72, 4634–4645. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Joung, J.G.; Fei, Z.; Jander, G. Interdependence of threonine, methionine and isoleucine metabolism in plants: Accumulation and transcriptional regulation under abiotic stress. Amino Acids 2010, 39, 933–947. [Google Scholar] [CrossRef]
- Lehmann, S.; Funck, D.; Szabados, L.; Rentsch, D. Proline metabolism and transport in plant development. Amino Acids 2010, 39, 949–962. [Google Scholar] [CrossRef]
- Ji, Y.; Zuo, L.; Wang, F.; Li, D.; Lai, C. Nutritional value of 15 corn gluten meals for growing pigs: Chemical composition, energy content and amino acid digestibility. Arch. Anim. Nutr. 2012, 66, 283–302. [Google Scholar] [CrossRef]
- Tariq, A.; Zeng, F.; Graciano, C.; Ullah, A.; Sadia, S.; Ahmed, Z.; Murtaza, G.; Zhang, Z. Regulation of metabolites by nutrients in plants. In Plant Ionomics: Sensing, Signaling, and Regulation; Wiley: Hoboken, NJ, USA, 2023; pp. 1–18. [Google Scholar]
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Tryptophan: A precursor of signaling molecules in higher plants. In Hormones and Plant Response. Plant in Challenging Environments; Springer: Cham, Switzerland, 2021; Volume 2, pp. 273–289. [Google Scholar]
- Pasković, I.; Popović, L.; Pongrac, P.; Polić Pasković, M.; Kos, T.; Jovanov, P.; Franić, M. Protein Hydrolysates—Production, Effects on Plant Metabolism, and Use in Agriculture. Horticulturae 2024, 10, 1041. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A new protein hydrolysate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H.; Kuang, Y.; Wang, N. Amino acids biostimulants and protein hydrolysates in agricultural sciences. Plants 2024, 13, 210. [Google Scholar] [CrossRef]
- Gan, L.; Song, M.; Wang, X.; Yang, N.; Li, H.; Liu, X.; Li, Y. Cytokinins are involved in regulation of tomato pericarp thickness and fruit size. Hortic. Res. 2022, 9, uhab041. [Google Scholar] [CrossRef] [PubMed]
- Soteriou, G.A.; Rouphael, Y.; Emmanouilidou, M.G.; Antoniou, C.; Kyratzis, A.C.; Kyriacou, M.C. Biostimulatory Action of Vegetal Protein Hydrolysate and the Configuration of Fruit Physicochemical Characteristics in Grafted Watermelon. Horticulturae 2021, 7, 313. [Google Scholar] [CrossRef]
- Cardarelli, M.; Ceccarelli, A.V.; El Nakhel, C.; Rouphael, Y.; Salehi, H.; Ganugi, P.; Zhang, L.; Lucini, L.; Pii, Y.; Choi, S.; et al. Foliar applications of a Malvaceae-derived protein hydrolysate and its fractions differentially modulate yield and functional traits of tomato under optimal and suboptimal nitrogen application. J. Sci. Food Agric. 2024, 104, 7603–7616. [Google Scholar] [CrossRef] [PubMed]
- Broxterman, S.E.; Schols, H.A. Interactions between pectin and cellulose in primary plant cell walls. Carbohydr. Polym. 2018, 192, 263–272. [Google Scholar] [CrossRef]
- Trovato, M.; Funck, D.; Forlani, G.; Okumoto, S.; Amir, R. Amino acids in plants: Regulation and functions in development and stress defense. Front. Plant Sci. 2021, 12, 772810. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Lal, M.A. Concepts of Plant Water Relations. In Plant Physiology, Development and Metabolism; Springer Nature: Singapore, 2023; pp. 3–23. [Google Scholar]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein hydrolysates and mo-biofortification interactively modulate plant performance and quality of ‘canasta’lettuce grown in a protected environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- Malécange, M.; Sergheraert, R.; Teulat, B.; Mounier, E.; Lothier, J.; Sakr, S. Biostimulant Properties of Protein Hydrolysates: Recent Advances and Future Challenges. Int. J. Mol. Sci. 2023, 24, 9714. [Google Scholar] [CrossRef]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Kumari, A.; Kumar, V.; Ovadia, R.; Oren-Shamir, M. Phenylalanine in motion: A tale of an essential molecule with many faces. Biotechnol. Adv. 2023, 68, 108246. [Google Scholar] [CrossRef] [PubMed]
- Ciriello, M.; Campana, E.; De Pascale, S.; Rouphael, Y. Implications of Vegetal Protein Hydrolysates for Improving Nitrogen Use Efficiency in Leafy Vegetables. Horticulturae 2024, 10, 132. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Yield (g/Plant) | NMFP (No./Plant) | MWMF (g) | |||
|---|---|---|---|---|---|---|
| Biostimulant (B) | ||||||
| Ctrl | 635.4 | b | 22.6 | b | 28.2 | a |
| PH | 763.3 | a | 30.3 | a | 25.3 | b |
| Application mode (M) | ||||||
| Foliar (Fol) | 702.4 | a | 25.9 | b | 27.4 | a |
| Fertigation (Fert) | 698.8 | a | 25.7 | b | 25.6 | b |
| Fol+Fert | 696.8 | a | 26.2 | a | 27.3 | a |
| Significance | ||||||
| B | *** (p = 0.001) | *** (p = 0.001) | ** (p = 0.009) | |||
| M | NS (p = 0.236) | ** (p = 0.008) | * (p = 0.042) | |||
| B×M | NS (p = 0.380) | NS (p = 0.108) | NS (p = 0.258) | |||
| Variables | PC1 | PC2 |
|---|---|---|
| Yield | 0.84 | 0.54 |
| NMFP | 0.75 | 0.64 |
| MWMF | −0.61 | −0.66 |
| SSC | −0.97 | −0.23 |
| Dry matter content | −0.93 | −0.09 |
| L* | 0.76 | 0.62 |
| Firmness | 0.39 | 0.77 |
| Ascorbic acid | 0.92 | 0.37 |
| Total phenols | 0.93 | 0.36 |
| Anthocyanins | 0.90 | 0.43 |
| N | −0.88 | −0.47 |
| P | −0.19 | 0.78 |
| K | 0.67 | 0.72 |
| Mg | 0.90 | −0.09 |
| NUE | 0.84 | 0.54 |
| Eigenvalue | ||
| Variance % | 63.22 | 28.45 |
| Cumulative % | 63.22 | 91.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancuso, F.; Vultaggio, L.; Sabatino, L.; Bellitto, P.; Ntatsi, G.; Allevato, E.; La Placa, G.G.; La Bella, S.; Consentino, B.B. Zea mays-Derived Protein Hydrolysate and Diverse Application Modes Differentially Compose Crop Production and Fruit Quality of Strawberry Cultivated Under Tunnel. Agronomy 2025, 15, 1314. https://doi.org/10.3390/agronomy15061314
Mancuso F, Vultaggio L, Sabatino L, Bellitto P, Ntatsi G, Allevato E, La Placa GG, La Bella S, Consentino BB. Zea mays-Derived Protein Hydrolysate and Diverse Application Modes Differentially Compose Crop Production and Fruit Quality of Strawberry Cultivated Under Tunnel. Agronomy. 2025; 15(6):1314. https://doi.org/10.3390/agronomy15061314
Chicago/Turabian StyleMancuso, Fabiana, Lorena Vultaggio, Leo Sabatino, Pietro Bellitto, Georgia Ntatsi, Enrica Allevato, Gaetano Giuseppe La Placa, Salvatore La Bella, and Beppe Benedetto Consentino. 2025. "Zea mays-Derived Protein Hydrolysate and Diverse Application Modes Differentially Compose Crop Production and Fruit Quality of Strawberry Cultivated Under Tunnel" Agronomy 15, no. 6: 1314. https://doi.org/10.3390/agronomy15061314
APA StyleMancuso, F., Vultaggio, L., Sabatino, L., Bellitto, P., Ntatsi, G., Allevato, E., La Placa, G. G., La Bella, S., & Consentino, B. B. (2025). Zea mays-Derived Protein Hydrolysate and Diverse Application Modes Differentially Compose Crop Production and Fruit Quality of Strawberry Cultivated Under Tunnel. Agronomy, 15(6), 1314. https://doi.org/10.3390/agronomy15061314