Carbon Use Efficiency in the Soil Profile: The Role of Soil Acidity Alleviation, Nitrogen, and Phosphorus Availability


Abstract

1. Introduction
2. Materials and Methods
2.1. Soil
2.2. Experiment 1—Microbial C Use Efficiency
2.3. Experiment 2—Stoichiometric Control of C Respiration
2.4. Statistics
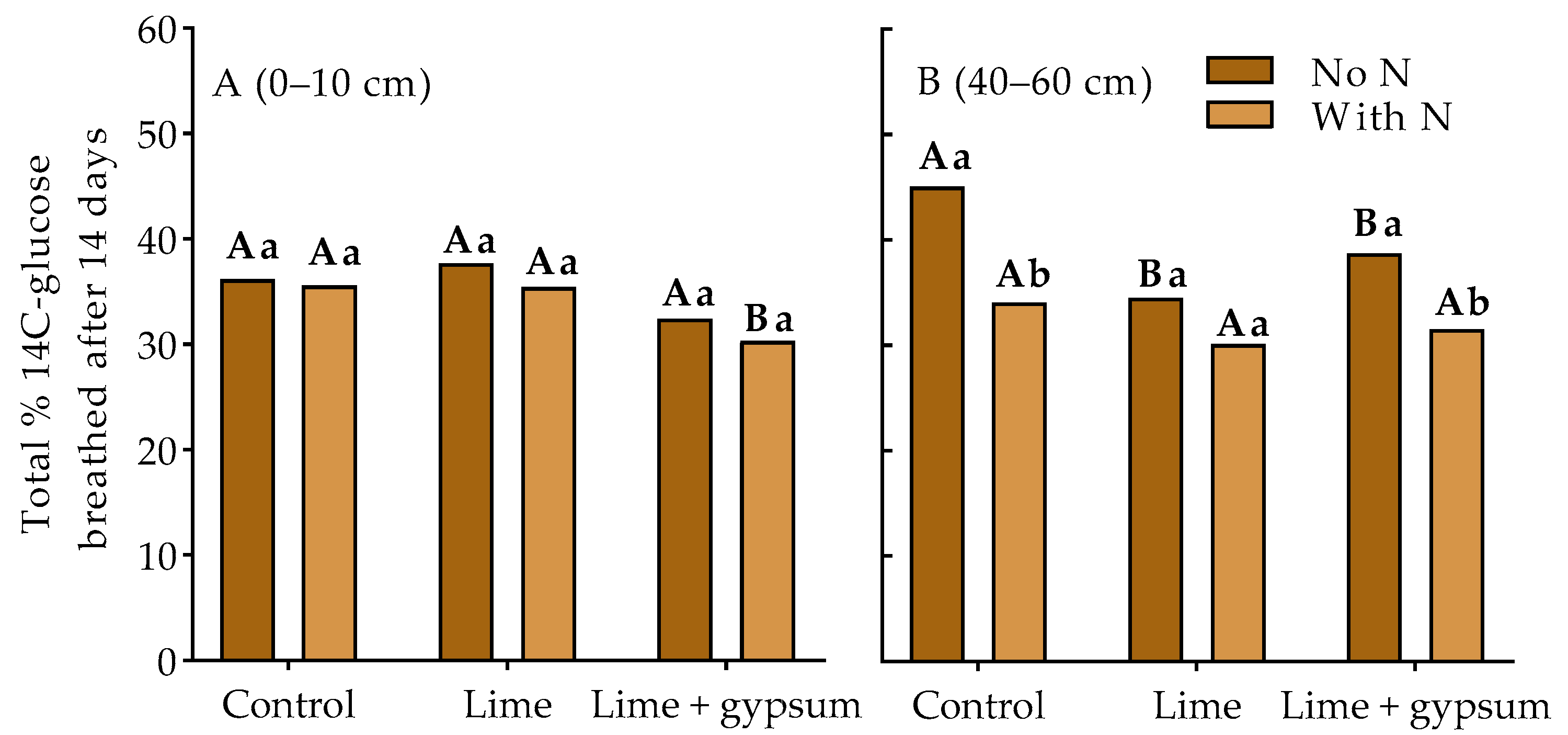
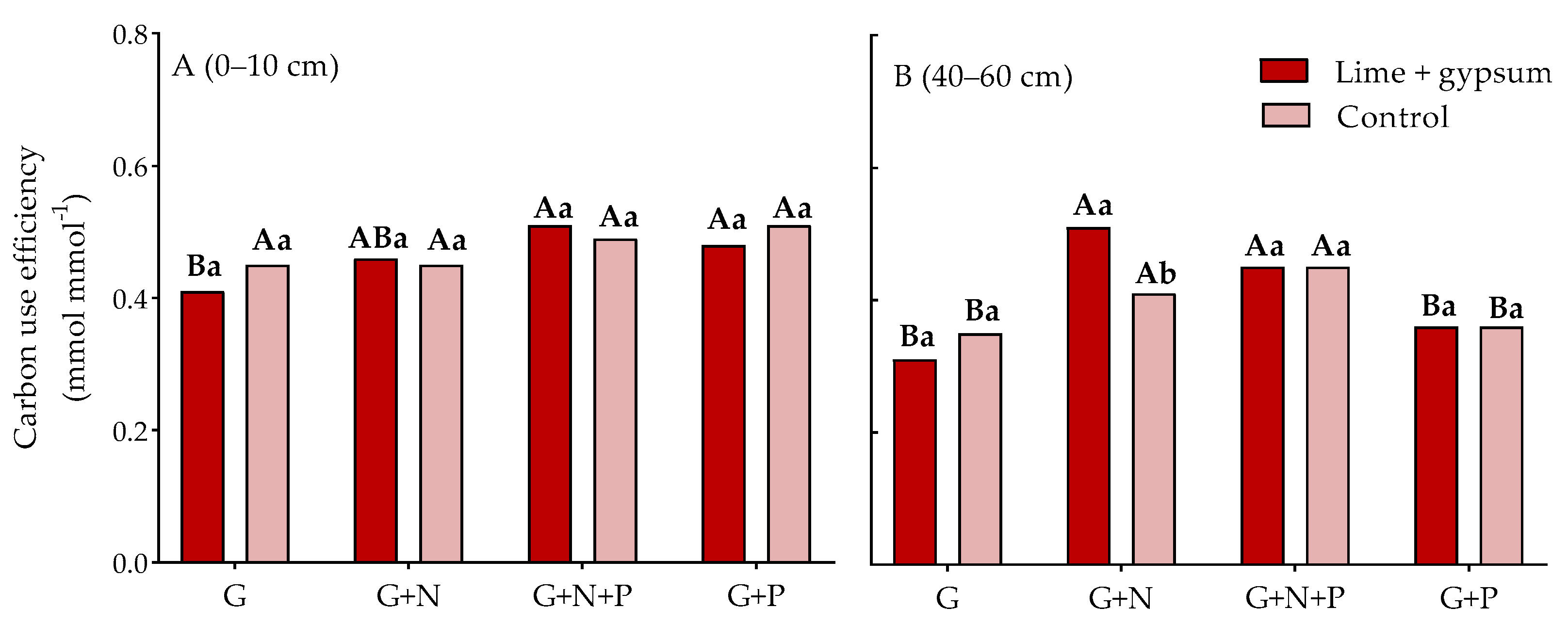
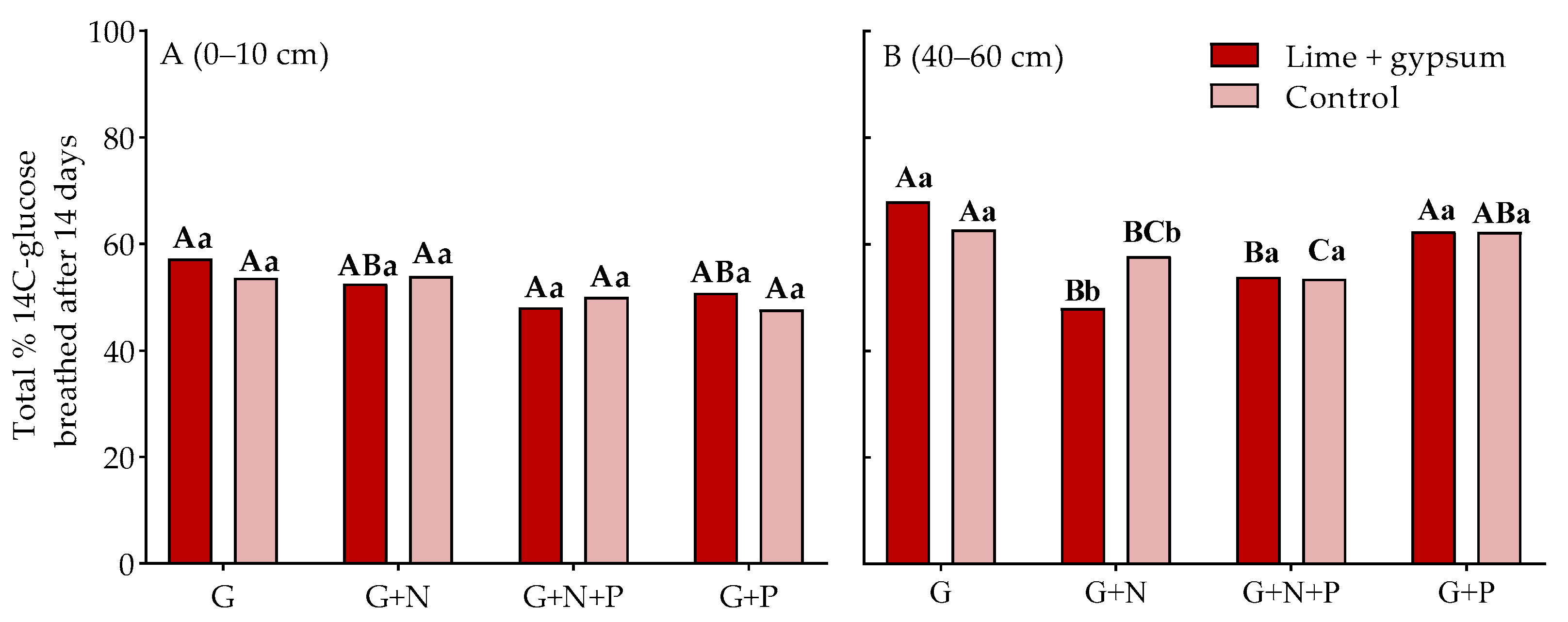
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Christensen, B.T. Matching measurable soil organic matter fractions with conceptual pools in simulation models of carbon turnover: Revision of model structure. In Evaluation of Soil Organic Matter Models Using Existing Long-Term Datasets; Powlson, D.S., Smith, P., Smith, J.U., Eds.; Global Environmental Change; Springer: Berlin/Heidelberg, Germany, 1996; Volume 38, pp. 143–160. [Google Scholar]
- Zelles, L.; Stepper, I.; Zsolnay, A. The effect of liming on microbial activity in spruce (Picea abies L.) forests. Biol. Fertil. Soils 1990, 9, 78–82. [Google Scholar] [CrossRef]
- Acosta-Martinez, V.; Tabatabai, M.A. Enzyme activities in a limed agricultural soil. Biol. Fertil. Soils 2000, 31, 85–91. [Google Scholar] [CrossRef]
- Lal, R. Challenges and opportunities in soil organic matter research. Eur. J. Soil Sci. 2009, 60, 158–169. [Google Scholar] [CrossRef]
- Barcelos, J.P.Q.; Souza, M.; Nascimento, C.A.C.; Rosolem, C.A. Soil acidity amelioration improves N and C cycles in the short term in a system with soybean followed by maize-guinea grass intercropping. Geoderma 2022, 421, 115909. [Google Scholar] [CrossRef]
- Salome, C.; Nunan, N.; Pouteau, V.; Lerch, T.Z.; Chenu, C. Carbon dynamics in topsoil and subsoil may be controlled by different regulatory mechanisms. Glob. Change Biol. 2010, 16, 416–426. [Google Scholar] [CrossRef]
- Sanaullah, M.; Chabbi, A.; Leifeld, J.; Bardoux, G.; Billou, D.; Rumpel, C. Decomposition and stabilization of root litter in top- and subsoil horizons: What is the difference? Plant Soil 2011, 338, 127–141. [Google Scholar] [CrossRef]
- Cook, F.J.; Knight, J.H. Where does oxygen extinction occur in a soil profile? In Proceedings of the 21st International Congress on Modelling and Simulation, Gold Coast, Australia, 29 November–4 December 2015; Available online: https://www.mssanz.org.au/modsim2015/J8/cook.pdf (accessed on 10 April 2025).
- Rovira, P.; Vallejo, V.R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach. Geoderma 2002, 107, 109–141. [Google Scholar] [CrossRef]
- Barcelos, J.P.Q.; Mariano, E.; Jones, D.L.; Rosolem, C.A. Topsoil and subsoil C and N turnover are affected by superficial lime and gypsum application in the short-term. Soil Biol. Biochem. 2021, 163, 108456. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.-H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar] [CrossRef]
- Glanville, H.C.; Hill, P.W.; Schnepf, A.; Oburger, E.; Jones, D.L. Combined use of empirical data and mathematical modelling to better estimate the microbial turnover of isotopically labelled carbon substrates in soil. Soil Biol. Biochem. 2016, 94, 154–168. [Google Scholar] [CrossRef]
- Qiao, Y.; Wang, J.; Liang, G.; Du, Z.; Zhou, J.; Zhu, C.; Huang, K.; Zhou, X.; Luo, Y.; Xia, J. Global variation of soil microbial carbon-use efficiency in relation to growth temperature and substrate supply. Sci. Rep. 2019, 9, 5621. [Google Scholar] [CrossRef] [PubMed]
- Creamer, R.E.; Stone, D.; Berry, P.; Kuiper, I. Measuring respiration profiles of soil microbial communities across Europe using MicroResp™ method. Appl. Soil Ecol. 2016, 97, 36–43. [Google Scholar] [CrossRef]
- Creamer, R.E.; Schulte, R.P.O.; Stone, D.; Gal, A.; Krogh, P.H.; Lo Papa, G.; Murray, P.J.; Peres, G.; Foerster, B.; Rutgers, M.; et al. Measuring basal soil respiration across Europe: Do incubation temperature and incubation period matter? Ecol. Indic. 2014, 36, 409–418. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Liptzin, D. C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 2007, 85, 235–252. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon use efficiency of microbial communities: Stoichiometry, methodology and modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J.; Emmett, B.A.; Gundersen, P.; Kjonaas, O.J.; Koopmans, C.J.; Schleppi, P.; Tietema, A.; Wright, R.F. Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests. Nature 1998, 398, 145–148. [Google Scholar] [CrossRef]
- Magnani, F.; Mencuccini, M.; Borghetti, M.; Berbigier, P.; Berninger, F.; Delzon, S.; Grelle, A.; Hari, P.; Jarvis, P.G.; Kolari, P.; et al. The human footprint in the carbon cycle of temperate and boreal forests. Nature 2007, 447, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Leite, H.M.F.; Calonego, J.C.; Rosolem, C.A.; Mendes, L.W.; Moraes, L.N.; Grotto, R.M.T.; Araujo, F.F.; Pereira, A.P.A.; Melo, V.M.M.; Araujo, A.S.F. Cover crops shape the soil bacterial community in a tropical soil under no-till. Appl. Soil Ecol. 2021, 168, 104166. [Google Scholar] [CrossRef]
- Allison, S.D. Cheaters, diffusion and nutrients constrain decomposition by microbial enzymes in spatially structured environments. Ecol. Lett. 2005, 8, 626–635. [Google Scholar] [CrossRef]
- Frankenberger, W.T.; Johanson, J.B. Effect of pH on enzyme stability in soils. Soil Biol. Biochem. 1982, 14, 433–437. [Google Scholar] [CrossRef]
- Turner, B.L. Variation in pH optima of hydrolytic enzyme activities in tropical rain forest soils. Appl. Environ. Microbiol. 2010, 76, 6485–6493. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.A.; Puissant, J.; Buckeridge, K.M.; Goodall, T.; Jehmlich, N.; Chowdhury, S.; Gweon, H.S.; Peyton, J.M.; Manson, K.E.; van Agtmaal, M.; et al. Land use driven change in soil pH affects microbial carbon cycling processes. Nat. Commun. 2018, 9, 3591. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.; Damatîca, C.; Bolscher, T.; Chenu, C.; Elsgaard, L.; Tebbe, C.C.; Skadell, L.; Poeplau, C. Liming effects on microbial carbon use efficiency and its potential consequences for soil organic stocks. Soil Biol. Biochem. 2024, 191, 109342. [Google Scholar] [CrossRef]
- Craine, J.M.; Morrow, C.; Fierer, N. Microbial nitrogen limitation increases decomposition. Ecology 2007, 88, 2105–2113. [Google Scholar] [CrossRef]
- Billings, S.A.; Ziegler, S.E. Altered patterns of soil carbon substrate usage and heterotrophic respiration in a pine forest with elevated CO2 and N fertilization. Glob. Change Biol. 2008, 14, 1025–1036. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). Keys to Soil Taxonomy, 12th ed.; Soil Survey Staff: Washington, DC, USA, 2014.
- Raij, B.V.; Andrade, J.C.; Cantarella, H.; Quaggio, J.A. Análise Química Para Avaliação da Fertilidade de Solos Tropicais; IAC: Campinas, Brazil, 2001. [Google Scholar]
- Gunina, A.; Kuzyakov, Y. Sugars in soil and sweets for microorganisms: Review of origin, content, composition and fate. Soil Biol. Biochem. 2015, 90, 87–100. [Google Scholar] [CrossRef]
- Haney, R.L.; Franzluebbers, A.J.; Porter, E.B.; Hons, F.M.; Zuberer, D.A. Soil carbon and nitrogen mineralization: Influence of drying temperature. Soil Sci. Soc. Am. J. 2004, 68, 489–492. [Google Scholar] [CrossRef]
- Hill, P.W.; Farrar, J.F.; Jones, D.L. Decoupling of microbial glucose uptake and mineralization in soil. Soil Biol. Biochem. 2008, 40, 616–624. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Banerjee, M.R.; Chapman, S.J. The significance of microbial biomass sulphur in soil. Biol. Fertil. Soils 1996, 22, 116–125. [Google Scholar] [CrossRef]
- He, Z.L.; Wu, J.; O’Donnell, A.G.; Syers, J.K. Seasonal responses in microbial biomass carbon, phosphorus and sulphur in soils under pasture. Biol. Fertil. Soils 1997, 24, 421–428. [Google Scholar] [CrossRef]
- Chowdhury, M.A.H.; Kouno, K.; Ando, T. Correlation among microbial biomass S, soil properties, and other biomass nutrients. Soil Sci. Plant Nutr. 1999, 45, 175–186. [Google Scholar] [CrossRef]
- Chowdhury, M.A.H.; Kouno, K.; Ando, T. Critical sulphur concentration and sulphur requirement of microbial biomass in a glucose and cellulose-amended regosol. Biol. Fertil. Soils 2000, 32, 310–317. [Google Scholar] [CrossRef]
- Kramer, C.; Trumbore, S.; Fröberg, M.; Dozal, L.M.C.; Zhang, D.C.; Xu, X.M.; Santos, G.M.; Hanson, P.J. Recent (4 year old) leaf litter is not a major source of microbial carbon in a temperate forest mineral soil. Soil Biol. Biochem. 2010, 42, 1028–1037. [Google Scholar] [CrossRef]
- Kleber, M.; Eusterhues, K.; Keiluweit, M.; Mikutta, C.; Mikutta, R.; Nico, P.S. Chapter One—Mineral–Organic Associations: Formation, Properties, and Relevance in Soil Environments. Adv. Agron. 2015, 130, 1–140. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of microbial biomass carbon and nitrogen in soil. In Advances in Nitrogen Cycling in Agricultural Systems; Wilson, J.R., Ed.; CAB International: Wallingford, UK, 1998; pp. 368–386. [Google Scholar]
- Fischer, H.; Ingwersen, J.; Kuzyakov, Y. Microbial uptake of low-molecular-weight organic substances out competes sorption in soil. Eur. J. Soil Sci. 2010, 61, 504–513. [Google Scholar] [CrossRef]
- Mota Neto, L.V.; Barros, J.V.S.; Costa, V.E.; Galdos, M.V.; Santos, A.R.P.; Rosolem, C.A. Insights on soil carbon cycling in intercropped maize-forage systems as affected by nitrogen. Geoderma 2024, 449, 116998. [Google Scholar] [CrossRef]
- Malik, A.A.; Thomson, B.C.; Whiteley, A.S.; Bailey, M.; Griffiths, R.I. Bacterial physiological adaptations to contrasting edaphic conditions identified using landscape scale metagenomics. mBio 2017, 8, e0079. [Google Scholar] [CrossRef]
- Puissant, J.; Jones, B.; Goodall, T.; Mang, D.; Blaud, A.; Gweon, H.S.; Malik, A.; Jones, D.L.; Clark, I.M.; Hirsch, P.R. The pH optimum of soil exoenzymes adapt to long term changes in soil pH. Soil Biol. Biochem. 2019, 138, 107601. [Google Scholar] [CrossRef]
- Wang, C.; Kuzyakov, Y. Soil organic matter priming: The pH effects. Glob. Change Biol. 2024, 30, e17349. [Google Scholar] [CrossRef] [PubMed]
- Commichau, F.M.; Forchhammer, K.; Stülke, J. Regulatory links between carbon and nitrogen metabolism. Curr. Opin. Microbiol. 2006, 9, 167–172. [Google Scholar] [CrossRef]
- Conde, E.; Cardenas, M.; Ponce-Mendoza, A.; Luna-Guido, M.L.; Cruz-Mondragón, C.; Dendooven, L. The impacts of inorganic nitrogen application on mineralization of 14C-labelled maize and glucose, and on priming effect in saline alkaline soil. Soil Biol. Biochem. 2005, 37, 681–691. [Google Scholar] [CrossRef]
- Russell, J.B. The energy spilling reactions of bacteria and other organisms. J. Mol. Microbiol. Biotechnol. 2007, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- De Goffau, M.C.; Yang, X.M.; Van Dijl, J.M.; Harmsen, H.J.M. Bacterial pleomorphism and competition in a relative humidity gradient. Environ. Microbiol. 2009, 11, 809–822. [Google Scholar] [CrossRef]
- Lebre, P.H.; De Maayer, P.; Cowan, D.A. Xerotolerant bacteria: Surviving through a dry spell. Nat. Rev. Microbiol. 2017, 15, 285–296. [Google Scholar] [CrossRef]
- Santi, A.; Amado, T.J.C.; Costa, J.A.A. Adubação nitrogenada na aveia preta: Influência na produção de matéria seca e ciclagem de nutrientes sob sistema plantio direto. Rev. Bras Cienc. Solo 2003, 27, 1075–1083. [Google Scholar] [CrossRef]
- Raphael, J.P.; Calonego, J.C.; Milori, D.M.B.; Rosolem, C.A. Soil organic matter in crop rotations under no-till. Soil Tillage Res. 2016, 155, 45–53. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient co-limitation of primary producer communities. Ecol. Lett. 2011, 14, 852–862. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, C.A.; Kirkegaard, J.A.; Richardson, A.E.; Wade, L.J.; Blanchard, C.; Batten, G. Stable soil organic matter: A comparison of C: N:P: S ratios in Australian and other world soils. Geoderma 2011, 163, 197–208. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Hessen, D.O.; Ågren, G.I.; Anderson, T.R.; Elser, J.J.; Ruiter, P.C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology 2004, 85, 1179–1192. [Google Scholar] [CrossRef]
- Ågren, G.I.; Wetterstedt, J.Å.M. What determines the temperature response of soil organic matter decomposition? Soil Biol. Biochem. 2007, 39, 1794–1798. [Google Scholar] [CrossRef]
- Wagai, R.; Kishimoto-Mo, A.W.; Yonemura, S.; Shirato, Y.; Hiradate, S.; Yagasaki, Y. Linking temperature sensitivity of soil organic matter decomposition to its molecular structure, accessibility, and microbial physiology. Glob. Change Biol. 2013, 19, 1114–1125. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, X.; Wang, D.; Wang, M.; Liao, C.; Yang, X.; Liu, F. Decoupled linkage between soil carbon and nitrogen mineralization among soil depths in a subtropical mixed forest. Soil Biol. Biochem. 2017, 109, 135–144. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Manzoni, S.; Pineiro, G.; Jackson, R.B.; Jobbagy, E.G.; Kim, J.H.; Porporato, A. Analytical models of soil and litter decomposition: Solutions for mass loss and time-dependent rates. Soil Biol. Biochem. 2012, 50, 66–76. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth | pH | SOM | P | S | K | Ca | Al | H+Al | CEC | BS |
|---|---|---|---|---|---|---|---|---|---|---|
| m | CaCl2 | g kg−1 | mg dm−3 | --------------------- mmc dm−3 --------------------------- | % | |||||
| 0.0–0.1 | 4.4 | 18 | 13 | 5 | 2.1 | 12 | 9 | 52 | 78 | 32 |
| 0.1–0.2 | 4.3 | 14 | 10 | 5 | 1.1 | 9 | 9 | 48 | 66 | 27 |
| 0.2–0.4 | 4.1 | 11 | 6 | 22 | 0.5 | 4 | 12 | 55 | 63 | 13 |
| 0.4–0.6 | 4.0 | 11 | 6 | 21 | 0.4 | 3 | 20 | 90 | 97 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertolino, K.M.; de Souza, M.; Jones, D.L.; Rosolem, C.A. Carbon Use Efficiency in the Soil Profile: The Role of Soil Acidity Alleviation, Nitrogen, and Phosphorus Availability. Agronomy 2025, 15, 1299. https://doi.org/10.3390/agronomy15061299
Bertolino KM, de Souza M, Jones DL, Rosolem CA. Carbon Use Efficiency in the Soil Profile: The Role of Soil Acidity Alleviation, Nitrogen, and Phosphorus Availability. Agronomy. 2025; 15(6):1299. https://doi.org/10.3390/agronomy15061299
Chicago/Turabian StyleBertolino, Karina Mendes, Murilo de Souza, Davey L. Jones, and Ciro Antonio Rosolem. 2025. "Carbon Use Efficiency in the Soil Profile: The Role of Soil Acidity Alleviation, Nitrogen, and Phosphorus Availability" Agronomy 15, no. 6: 1299. https://doi.org/10.3390/agronomy15061299
APA StyleBertolino, K. M., de Souza, M., Jones, D. L., & Rosolem, C. A. (2025). Carbon Use Efficiency in the Soil Profile: The Role of Soil Acidity Alleviation, Nitrogen, and Phosphorus Availability. Agronomy, 15(6), 1299. https://doi.org/10.3390/agronomy15061299