
Effect of Ethephon on Sensitivity Difference of Lodging Resistance in Different Maize Inbred Lines


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Reagents
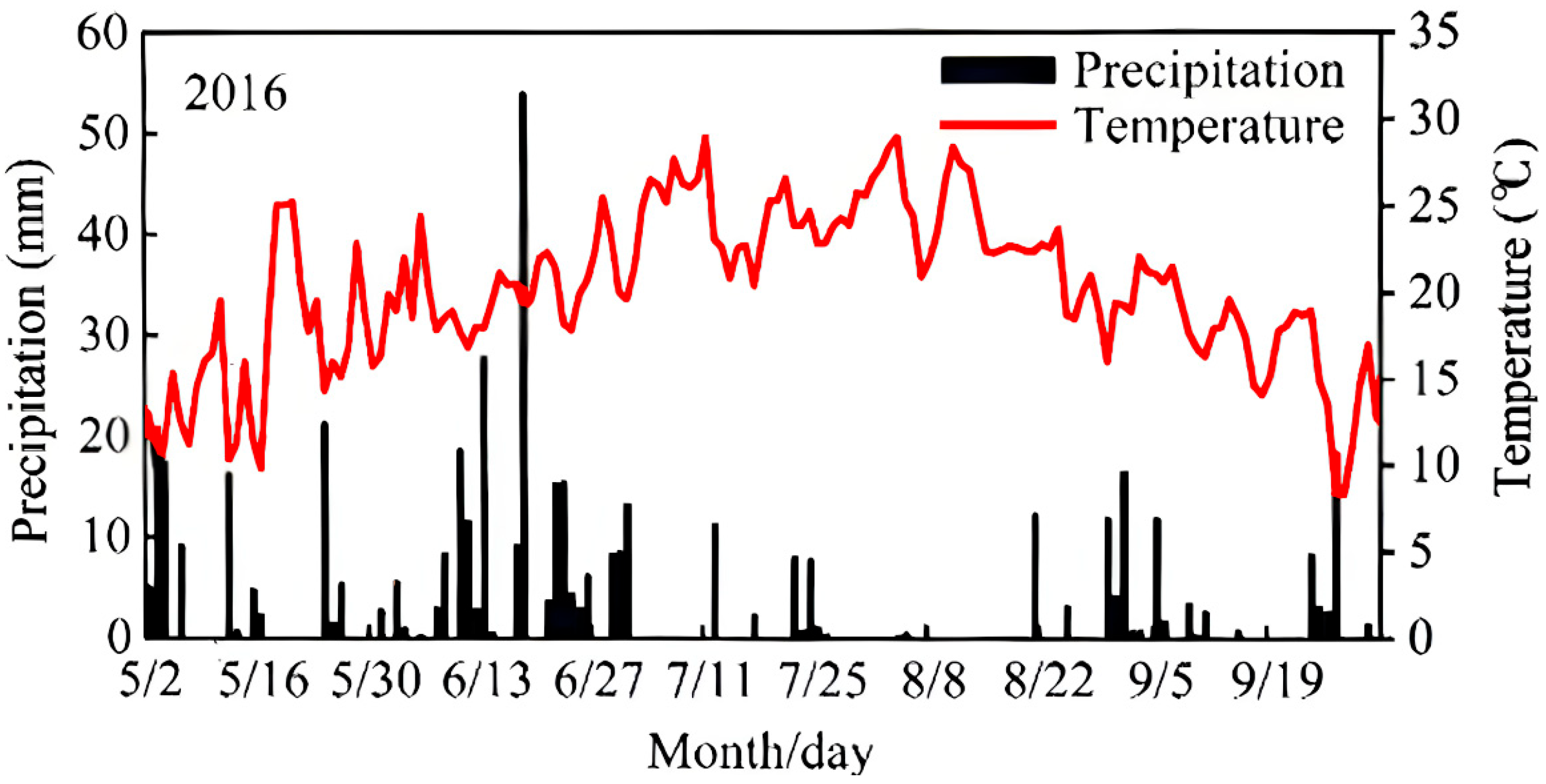
2.2. Experimental Design
2.3. Measurement Items and Methods
2.3.1. Maize Morphological Indicators
2.3.2. Stem Morphological Indicators
2.3.3. Measurement of Stem Strength
2.3.4. Measurement of Lignin Content and Related Enzyme Activities in Stalks
2.3.5. Measurement of Lodging Resistance Index
2.4. Data Analysis
3. Results
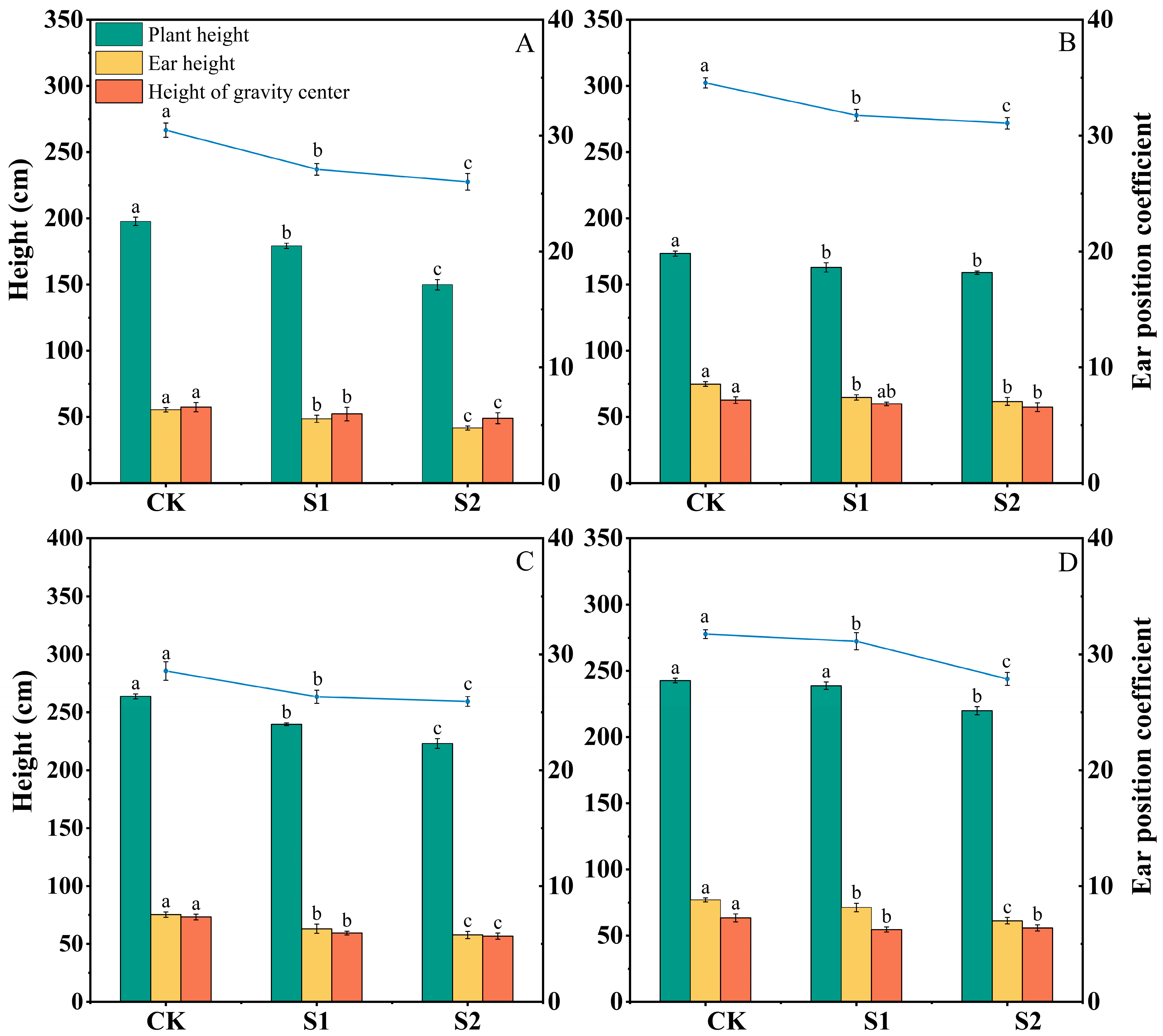
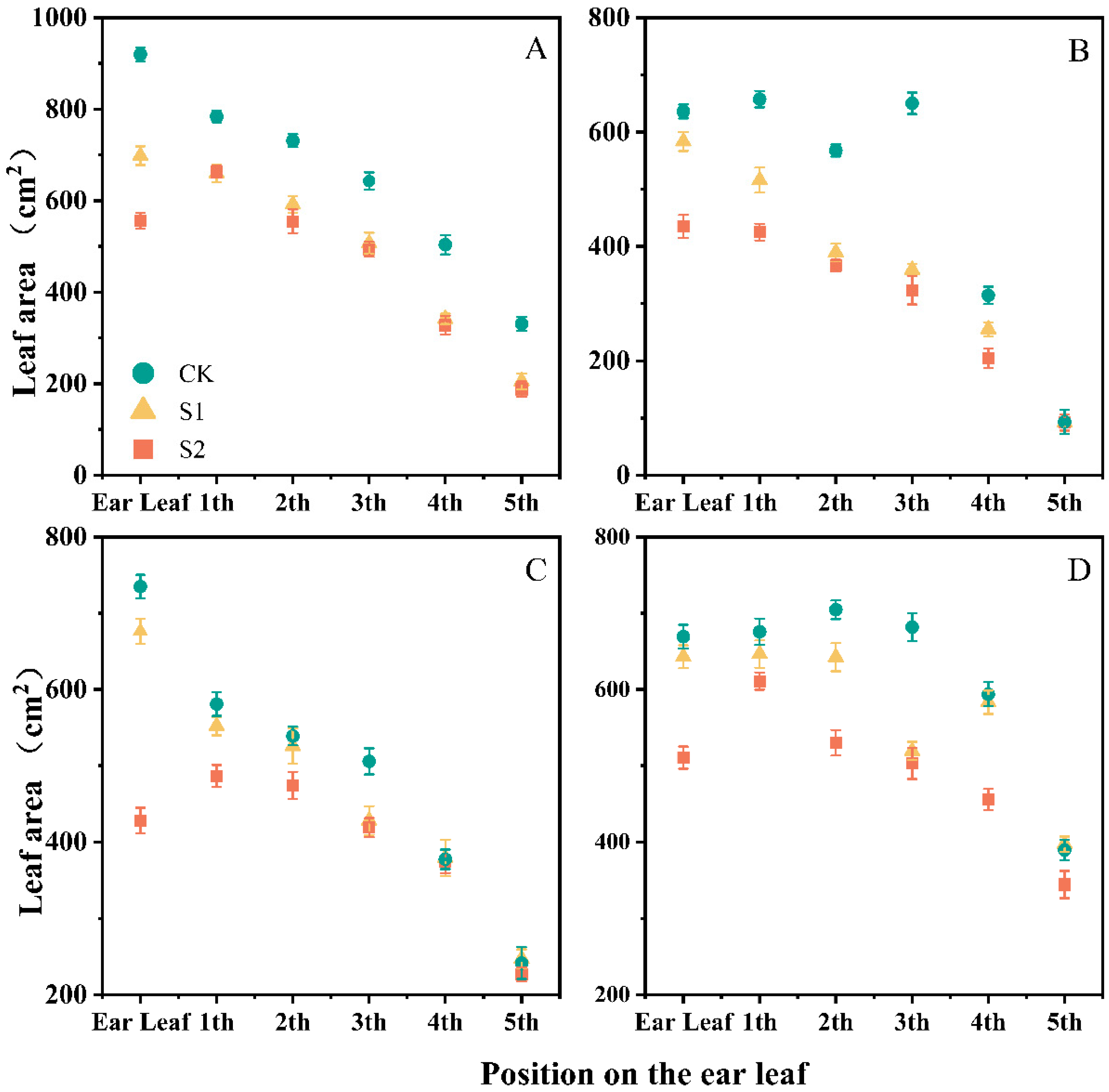
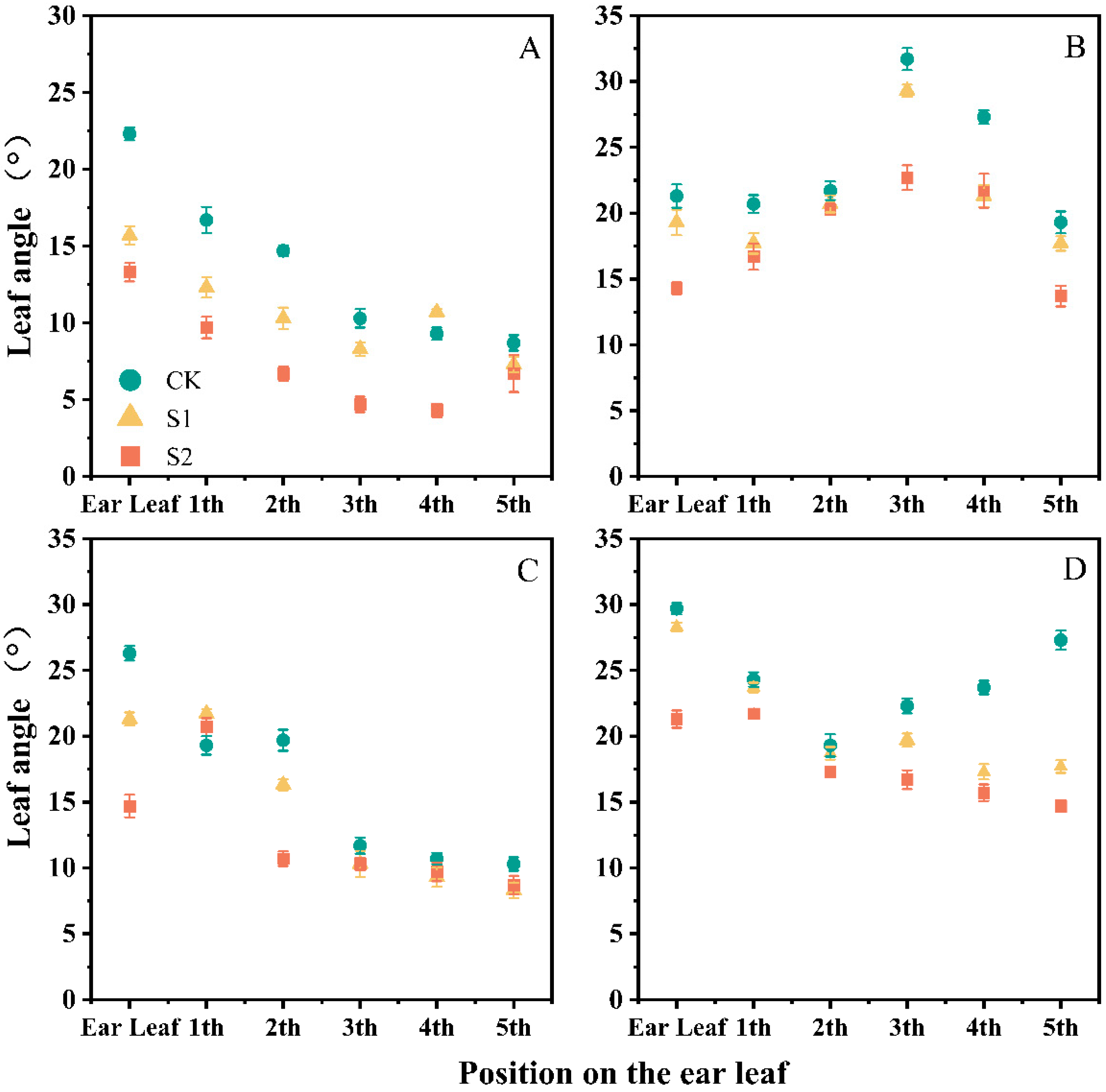
3.1. Effect of Ethephon on Maize Plant Morphology
3.2. Effects of Ethephon on Maize Stalk Morphology and Biomechanical Properties
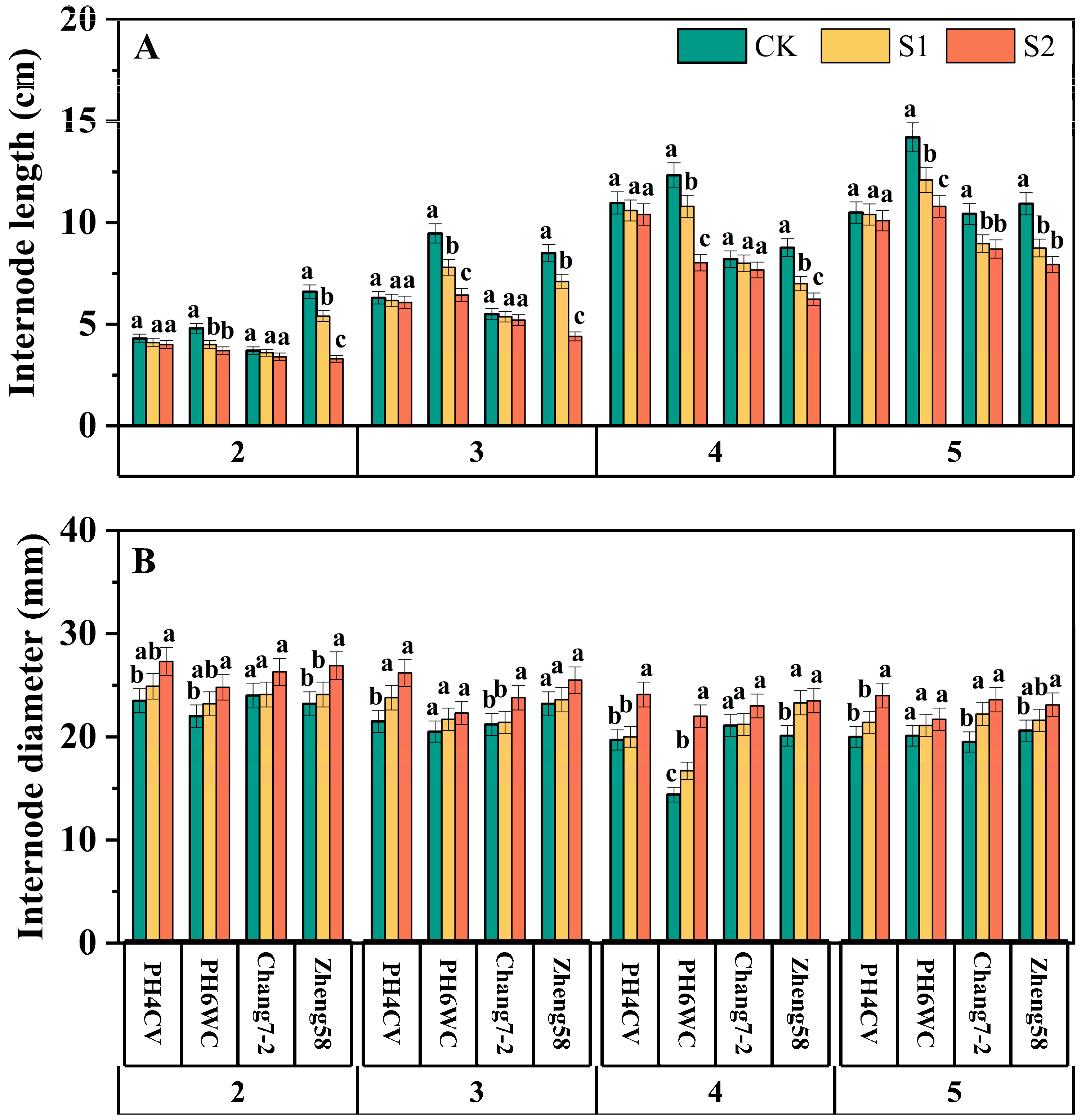
3.2.1. Internode Morphology
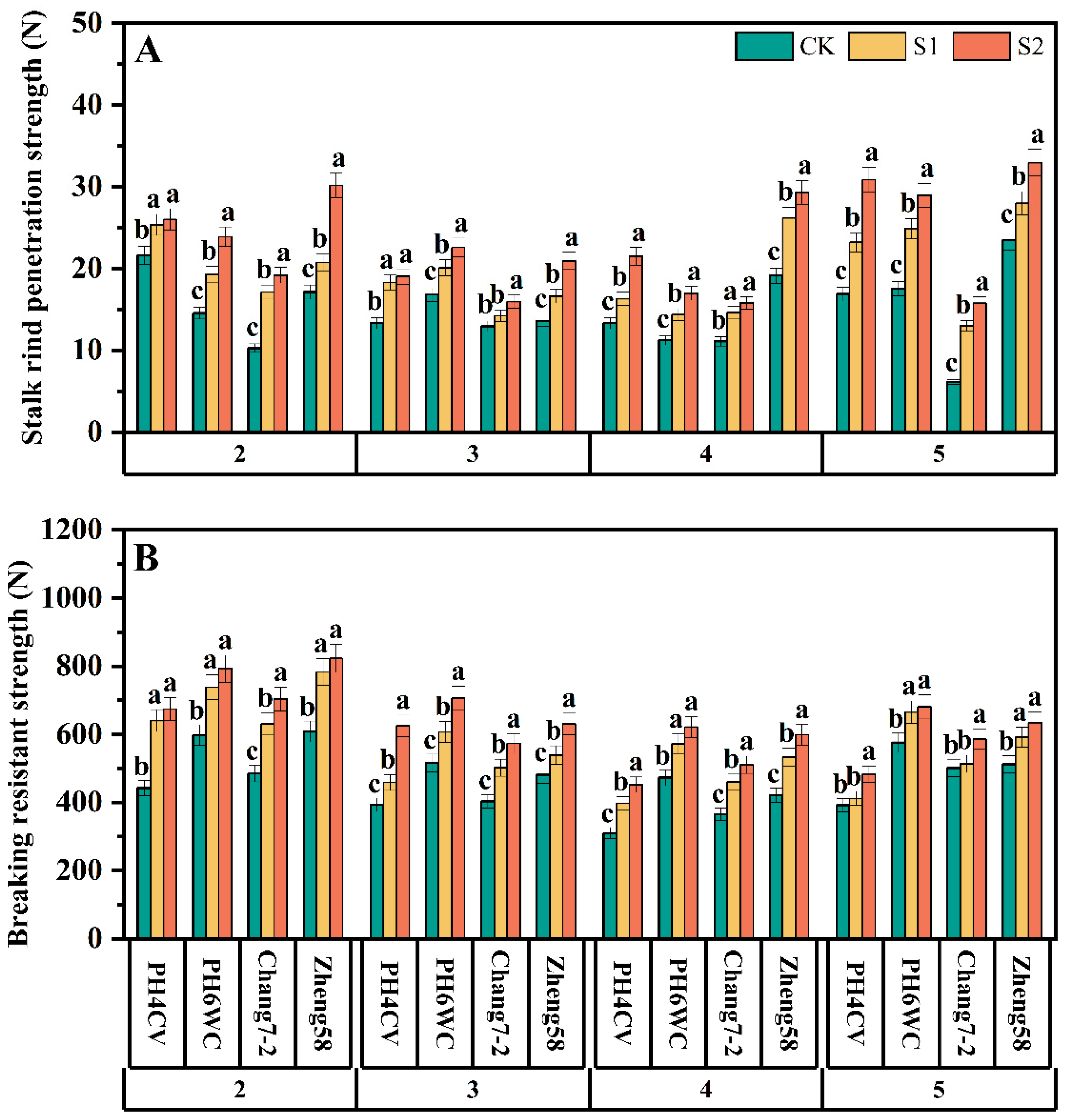
3.2.2. Physical Strength of Internodes
3.3. The Effect of Ethephon on Lignin Content and Related Enzyme Activity in Maize Internodes
3.4. The Effect of Ethephon on Lodging Resistance Index and Yield Components of Maize
3.5. Correlation Analysis of Various Agronomic Traits of Plants with Lodging Resistance Index and Yield
4. Discussion
4.1. Key Characteristics of Lodging Resistance in Maize
4.2. Effects of Ethephon on Lodging Traits of Maize
4.3. Influence of Variety Difference
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Zhang, Z.; Tao, F.L.; Wang, P.; Wei, X. Spatio-temporal patterns of winter wheat yield potential and yield gap during the past three decades in North China. Field Crops Res. 2017, 206, 11–20. [Google Scholar] [CrossRef]
- Hunt, J.R.; Hayman, P.T.; Richards, R.A.; Passioura, J.B. Opportunities to reduce heat damage in rain-fed wheat crops based on plant breeding and agronomic management. Field Crops Res. 2018, 224, 126–138. [Google Scholar] [CrossRef]
- Gong, L.S.; Qu, S.J.; Huang, G.M.; Guo, Y.L.; Zhang, M.C.; Li, Z.H.; Zhou, Y.Y.; Duan, L.S. Improving maize grain yield by formulating plant growth regulator strategies in North China. J. Integr. Agric. 2021, 20, 622–632. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, S. Research progress on traits and assessment methods of stalk lodging resistance in maize. Acta Agron. Sin. 2022, 48, 15–26. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Lv, Y.; Ning, F.; Cao, Y.; Liao, S.; Wang, P.; Huang, S. Optimizing ear-plant height ratio to improve kernel number and lodging resistance in maize (Zea mays L.). Field Crops Res. 2022, 276, 108376. [Google Scholar] [CrossRef]
- Ma, X.; Lu, M.; Xing, J.; Mo, T.; Liu, X.; Wu, Y.; Yuan, J.; Kong, F. Effects of Planting Density on Stalk and Lodging-Resistance in Summer Maize. J. Maize Sci. 2018, 26, 118–125. [Google Scholar]
- Robertson, D.; Smith, S.; Gardunia, B.; Cook, D. An Improved Method for Accurate Phenotyping of Corn Stalk Strength. Crop Sci. 2014, 54, 2038–2044. [Google Scholar] [CrossRef]
- Ghorbani, M.; Amirahmadi, E. Biochar and soil contributions to crop lodging and yield performance—A meta-analysis. Plant Physiol. Biochem. 2024, 215, 109053. [Google Scholar] [CrossRef]
- Yang, J.P.; Li, M.; Yin, Y.; Liu, Y.; Gan, X.K.; Mu, X.H.; Li, H.Q.; Li, J.K.; Li, H.C.; Zheng, J.; et al. Spatial accumulation of lignin monomers and cellulose underlying stalk strength in maize. Plant Physiol. Biochem. 2024, 214, 108918. [Google Scholar] [CrossRef]
- Li, Q.; Fu, C.F.; Liang, C.L.; Ni, X.J.; Zhao, X.H.; Chen, M.; Ou, L.J. Crop Lodging and the Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy 2022, 12, 1795. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Luan, Y.T.; Shi, W.B.; Tang, Y.H.; Huang, X.Q.; Tao, J. Melatonin enhances stem strength by increasing lignin content and secondary cell wall thickness in herbaceous peony. J. Exp. Bot. 2022, 73, 5974–5991. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Asghar, M.A.; Javed, H.H.; Ullah, A.; Cheng, B.; Xu, M.; Wang, W.Y.; Liu, C.Y.; Rahman, A.; Iqbal, T.; et al. Optimum nitrogen improved stem breaking resistance of intercropped soybean by modifying the stem anatomical structure and lignin metabolism. Plant Physiol. Biochem. 2023, 199, 10. [Google Scholar] [CrossRef]
- Rehman, M.; Luo, D.J.; Mubeen, S.; Pan, J.; Cao, S.; Saeed, W.; Chen, P. Progress in Agronomic Crops Lodging Resistance and Prevention: A Review. J. Agron. Crop Sci. 2024, 210, e12785. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Wang, Y.B.; Ye, D.L.; Xing, J.P.; Duan, L.S.; Li, Z.H.; Zhang, M.C. Ethephon-regulated maize internode elongation associated with modulating auxin and gibberellin signal to alter cell wall biosynthesis and modification. Plant Sci. 2020, 290, 110196. [Google Scholar] [CrossRef]
- Chai, M.; Li, Z.; Qin, D.; Liu, Y.; Xu, M.; Dong, L.; Zhang, Q.; Yang, D.; Zhang, P. Effect of Ethephon on Lodging Resistance of Maize Stem. J. Maize Sci. 2017, 25, 63–72. [Google Scholar]
- Geng, W.J.; Sun, Z.C.; Ren, B.Z.; Ren, H.; Zhao, B.; Liu, P.; Zhang, J.W. Spraying Ethephon Effectively Increased Canopy Light Transmittance of Densely Planted Summer Maize, Thus Achieving Synergistic Improvement in Stalk Lodging Resistance and Grain Yield. Plants 2022, 11, 2219. [Google Scholar] [CrossRef]
- Geng, W.; Li, B.; Ren, B.; Zhao, B.; Liu, P.; Zhang, J. Regulation Mechanism of Planting Density and Spraying Ethephon on Lignin Metabolism and Lodging Resistance of Summer Maize. Sci. Agric. Sin. 2022, 55, 307–319. [Google Scholar]
- Zou, X.J.; Xu, J.Y.; Xu, H.S.; Gong, L.; Zhang, L.Z.; Wang, Y.; Feng, L.S.; Sun, Z.X. Ethephon and Diethyl Aminoethyl Mixture (EDAH) Reduces Maize Lodging and Enhances Overall Productivity in Maize/Peanut Intercropping. Agronomy 2025, 15, 84. [Google Scholar] [CrossRef]
- Ren, J.H.; Tang, Q.; Niu, S.D.; Liu, S.S.; Wei, D.J.; Zhang, Y.R.; Gao, Z. High Dose of Plant Growth Regulator Enhanced Lodging Resistance Without Grain Yield Reduction of Maize Under High Density. Int. J. Plant Prod. 2022, 16, 329–339. [Google Scholar] [CrossRef]
- Reddy, M.V.B.; Arul, J.; Angers, P.; Couture, L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seed quality. J. Agric. Food Chem. 1999, 47, 1208–1216. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B. Tissue and method specificities of phenylalanine ammonia-lyase assay. J. Plant Physiol. 2012, 169, 1317–1320. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, K.H.; Hahlbrock, K. Isoenzymes of p-coumarate: CoA ligase from soybean cell suspension cultures (author’s transl). Planta Medica 1975, Suppl, 102–111. [Google Scholar] [CrossRef]
- Morrison, T.A.; Kessler, J.R.; Hatfield, R.D.; Buxton, D.R. Activity of two lignin biosynthesis enzymes during development of a maize internode. J. Sci. Food Agric. 1994, 65, 133–139. [Google Scholar] [CrossRef]
- Ahmad, I.; Ahmad, S.; Yang, X.N.; Meng, X.P.; Yang, B.P.; Liu, T.; Han, Q.F. Effect of uniconazole and nitrogen level on lodging resistance and yield potential of maize under medium and high plant density. Plant Biol. 2021, 23, 485–496. [Google Scholar] [CrossRef]
- Niu, Y.A.; Chen, T.X.; Zhao, C.C.; Zhou, M.X. Improving Crop Lodging Resistance by Adjusting Plant Height and Stem Strength. Agronomy 2021, 11, 2421. [Google Scholar] [CrossRef]
- Han, L.P.; Guo, X.Q.; Yu, Y.J.; Duan, L.S.; Rao, M.S.; Xie, G.H. Effect of prohexadione-calcium, maleic hydrazide and glyphosine on lodging rate and sugar content of sweet sorghum. Res. Crops 2011, 12, 230–238. [Google Scholar] [CrossRef]
- Fuli, C.; Xiong, D.U.; Mengxing, L.I.U.; Xiaoli, J.I.N.; Yanhong, C.U.I. Lodging of Summer Maize and the Effects on Grain Yield. Maize Sci. 2011, 19, 105–108. [Google Scholar]
- Xue, J.; Qi, B.Q.; Ma, B.Y.; Li, B.X.; Gou, L. Effect of altered leaf angle on maize stalk lodging resistance. Crop Sci. 2021, 61, 689–703. [Google Scholar] [CrossRef]
- Shah, A.N.; Tanveer, M.; Abbas, A.; Yildirim, M.; Shah, A.A.; Ahmad, M.I.; Wang, Z.W.; Sun, W.W.; Song, Y.H. Combating Dual Challenges in Maize Under High Planting Density: Stem Lodging and Kernel Abortion. Front. Plant Sci. 2021, 12, 699085. [Google Scholar] [CrossRef]
- Sekhon, R.S.; Joyner, C.N.; Ackerman, A.J.; McMahan, C.S.; Cook, D.D.; Robertson, D.J. Stalk Bending Strength is Strongly Associated with Maize Stalk Lodging Incidence Across Multiple Environments. Field Crops Res. 2020, 249, 107737. [Google Scholar] [CrossRef]
- Erndwein, L.; Cook, D.D.; Robertson, D.J.; Sparks, E.E. Field-based mechanical phenotyping of cereal crops to assess lodging resistance. Appl. Plant Sci. 2020, 8, e11382. [Google Scholar] [CrossRef] [PubMed]
- Jampatong, S. Effect of One- and Two-Eared selection on stalk strength and other characters in maize. Crop Sci. 1999, 40, 605–611. [Google Scholar] [CrossRef]
- Chesang-Chumo, J.J. Direct and Correlated Responses to Divergent Selection for Rind Penetrometer Resistance in MoSCSS Maize Synthetic; University of Missouri-Columbia: Columbia, MO, USA, 1993. [Google Scholar]
- Ling, G.O.U.; Ming, Z.; JianJun, H.; Bin, Z.; Tao, L.I.; Rui, S.U.N. Bending Mechanical Properties of Stalk and Lodging-Resistance of Maize (Zea mays L.). Acta Agron. Sin. 2008, 34, 653–661. [Google Scholar] [CrossRef]
- Wang, K.; Zhao, X.; Yao, X.; Yao, Y.; Bai, Y.; Wu, K. Relationship of stem characteristics and lignin synthesis with lodging resistance of hulless barley. Acta Agron. Sin. 2019, 45, 621–627. [Google Scholar] [CrossRef]
- Wu, L.M.; Zhang, W.J.; Ding, Y.F.; Zhang, J.W.; Cambula, E.D.; Weng, F.; Liu, Z.H.; Ding, C.Q.; Tang, S.; Chen, L.; et al. Shading Contributes to the Reduction of Stem Mechanical Strength by Decreasing Cell Wall Synthesis in Japonica Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 881. [Google Scholar] [CrossRef]
- Wang, X.Q.; Chen, Y.N.; Sun, X.; Li, J.H.; Zhang, R.Y.; Jiao, Y.Y.; Wang, R.H.; Song, W.; Zhao, J.R. Characteristics and candidate genes associated with excellent stalk strength in maize (Zea mays L.). Front. Plant Sci. 2022, 13, 957566. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Li, C.; Li, W.; Chen, X.; Zhang, L.; Wei, S. Dual application of ethephon and DCPTA increases maize yield and stalk strength. Agron. J. 2019, 111, 612–627. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Kumdee, O.; Wongkaew, A.; Khongchiu, P.; Worathongchai, N.; Alam, M.R.; Mahmud, A.A.; Nakasathien, S. Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse. Plants 2023, 12, 4192. [Google Scholar] [CrossRef]
- Huang, G.; Guo, Y.; Tan, W.; Zhang, M.; Li, Z.; Zhou, Y.; Duan, L. Enhancing maize radiation use efficiency under high planting density by shaping canopy architecture with a plant growth regulator. Crop Environ. 2024, 3, 51–63. [Google Scholar] [CrossRef]
- Ahmad, I.; Kamran, M.; Ali, S.; Bilegjargal, B.; Cai, T.; Ahmad, S.; Meng, X.P.; Su, W.N.; Liu, T.N.; Han, Q.F. Uniconazole application strategies to improve lignin biosynthesis, lodging resistance and production of maize in semiarid regions. Field Crops Res. 2018, 222, 66–77. [Google Scholar] [CrossRef]
- Zhao, Y.T.; Lv, Y.J.; Zhang, S.; Ning, F.F.; Cao, Y.B.; Liao, S.H.; Wang, P.; Huang, S.B. Shortening Internodes Near Ear: An Alternative to Raise Maize Yield. J. Plant Growth Regul. 2022, 41, 628–638. [Google Scholar] [CrossRef]
- Xu, C.L.; Gao, Y.B.; Tian, B.J.; Ren, J.H.; Meng, Q.F.; Wang, P. Effects of EDAH, a novel plant growth regulator, on mechanical strength, stalk vascular bundles and grain yield of summer maize at high densities. Field Crops Res. 2017, 200, 71–79. [Google Scholar] [CrossRef]
- Fan, H.; Gu, W.; Yang, D.; Yu, J.; Piao, L.; Zhang, Q.; Zhang, L.; Yang, X.; Wei, S. Effect of Chemical Regulators on Physical and Chemical Properties and Lodging Resistance of Spring Maize Stem in Northeast China. Acta Agron. Sin. 2018, 44, 909–919. [Google Scholar] [CrossRef]
- Liu, X.M.; Gu, W.R.; Li, C.F.; Li, J.; Wei, S. Effects of nitrogen fertilizer and chemical regulation on spring maize lodging characteristics, grain filling and yield formation under high planting density in Heilongjiang Province, China. J. Integr. Agric. 2021, 20, 511–526. [Google Scholar] [CrossRef]
- In, B.C.; Ha, S.T.T.; Lee, Y.S.; Lim, J.H. Relationships between the longevity, water relations, ethylene sensitivity, and gene expression of cut roses. Postharvest Biol. Technol. 2017, 131, 74–83. [Google Scholar] [CrossRef]
- Naing, A.H.; Soe, M.T.; Yeum, J.H.; Kim, C.K. Ethylene Acts as a Negative Regulator of the Stem-Bending Mechanism of Different Cut Snapdragon Cultivars. Front. Plant Sci. 2021, 12, 745038. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, M.; Li, Z.; Duan, L. Differences in Responding Sensitivity to Ethephon among Different Maize Genotypes. Acta Agron. Sin. 2011, 37, 1819–1827. [Google Scholar] [CrossRef]
- Massa, D.; Prisa, D.; Montoneri, E.; Battaglini, D.; Ginepro, M.; Negre, M.; Burchi, G. Application of municipal biowaste derived products in Hibiscus cultivation: Effect on leaf gaseous exchange activity, and plant biomass accumulation and quality. Sci. Hortic. 2016, 205, 59–69. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatment | LRI (%) | Grains per Ear | 1000-Grain Weight (g) | Yield (kg/hm2) |
|---|---|---|---|---|---|
| Zheng58 | CK | 8.51 ± 0.44 c | 624.8 ± 4.02 a | 350.59 ± 0.57 a | 9092.84 ± 54.88 a |
| S1 | 10.34 ± 0.32 b | 614.3 ± 6.14 b | 350.25 ± 0.31 a | 8935.75 ± 67.31 b | |
| S2 | 11.68 ± 0.58 a | 599.3 ± 8.19 c | 350.33 ± 0.44 a | 8828.81 ± 38.12 c | |
| Chang7-2 | CK | 6.75 ± 0.55 c | 608.0 ± 10.18 a | 340.72 ± 0.68 a | 8838.81 ± 20.87 a |
| S1 | 8.37 ± 0.71 b | 587.6 ± 4.62 b | 340.56 ± 0.33 a | 8773.83 ± 40.94 b | |
| S2 | 10.97 ± 1.26 a | 579.3 ± 2.31 c | 340.27 ± 0.41 a | 8686.50 ± 32.65 c | |
| PH6WC | CK | 7.31 ± 0.49 c | 617.0 ± 2.86 a | 350.72 ± 0.43 a | 9082.96 ± 40.26 a |
| S1 | 9.44 ± 1.47 b | 610.3 ± 5.23 ab | 350.54 ± 0.29 a | 8974.36 ± 38.79 b | |
| S2 | 12.45 ± 1.38 a | 602.3 ± 3.01 b | 350.42 ± 0.14 a | 8869.11 ± 30.64 c | |
| PH4CV | CK | 6.21 ± 0.64 c | 605.0 ± 5.70 a | 340.34 ± 0.26 a | 8863.02 ± 42.06 a |
| S1 | 8.21 ± 1.22 b | 599.6 ± 9.48 ab | 340.27 ± 0.15 a | 8780.33 ± 30.17 b | |
| S2 | 11.43 ± 0.86 a | 582.3 ± 8.95 b | 340.28 ± 0.27 a | 8650.49 ± 60.35 c |
| Treatment | Indicators | Plant Height | Ear Height | Height of Gravity Center | Leaf Angle | Leaf Area |
|---|---|---|---|---|---|---|
| CK | LRI | −0.84 * | −0.93 ** | −0.76 * | −0.93 ** | −0.81 * |
| Yield | −0.64 ns | −0.78 * | −0.69 ns | −0.89 ** | −0.98 ** | |
| S1 | LRI | −0.79 * | −0.98 ** | −0.88 * | −0.91 ** | −0.76 * |
| Yield | −0.71 ns | −0.76 * | −0.66 | −0.92 ** | −0.94 ** | |
| S2 | LRI | −0.80 * | −0.96 ** | −0.85 * | −0.89 ** | −0.83 * |
| Yield | −0.73 ns | −0.71 ns | −0.57 ns | −0.95 ** | −0.95 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Guo, F.; Chai, M.; Gu, S.; Wang, D.; Wang, Z.; Chen, Y.; Xie, T.; Yang, D.; Zhang, Q. Effect of Ethephon on Sensitivity Difference of Lodging Resistance in Different Maize Inbred Lines. Agronomy 2025, 15, 1248. https://doi.org/10.3390/agronomy15051248
Liu S, Guo F, Chai M, Gu S, Wang D, Wang Z, Chen Y, Xie T, Yang D, Zhang Q. Effect of Ethephon on Sensitivity Difference of Lodging Resistance in Different Maize Inbred Lines. Agronomy. 2025; 15(5):1248. https://doi.org/10.3390/agronomy15051248
Chicago/Turabian StyleLiu, Siyao, Feng Guo, Mengzhu Chai, Shiwei Gu, Dacheng Wang, Zihao Wang, Yidan Chen, Tenglong Xie, Deguang Yang, and Qian Zhang. 2025. "Effect of Ethephon on Sensitivity Difference of Lodging Resistance in Different Maize Inbred Lines" Agronomy 15, no. 5: 1248. https://doi.org/10.3390/agronomy15051248
APA StyleLiu, S., Guo, F., Chai, M., Gu, S., Wang, D., Wang, Z., Chen, Y., Xie, T., Yang, D., & Zhang, Q. (2025). Effect of Ethephon on Sensitivity Difference of Lodging Resistance in Different Maize Inbred Lines. Agronomy, 15(5), 1248. https://doi.org/10.3390/agronomy15051248