

Examining Nicotinamide Application Methods in Alleviating Lead-Induced Stress in Spring Barley

Abstract
1. Introduction
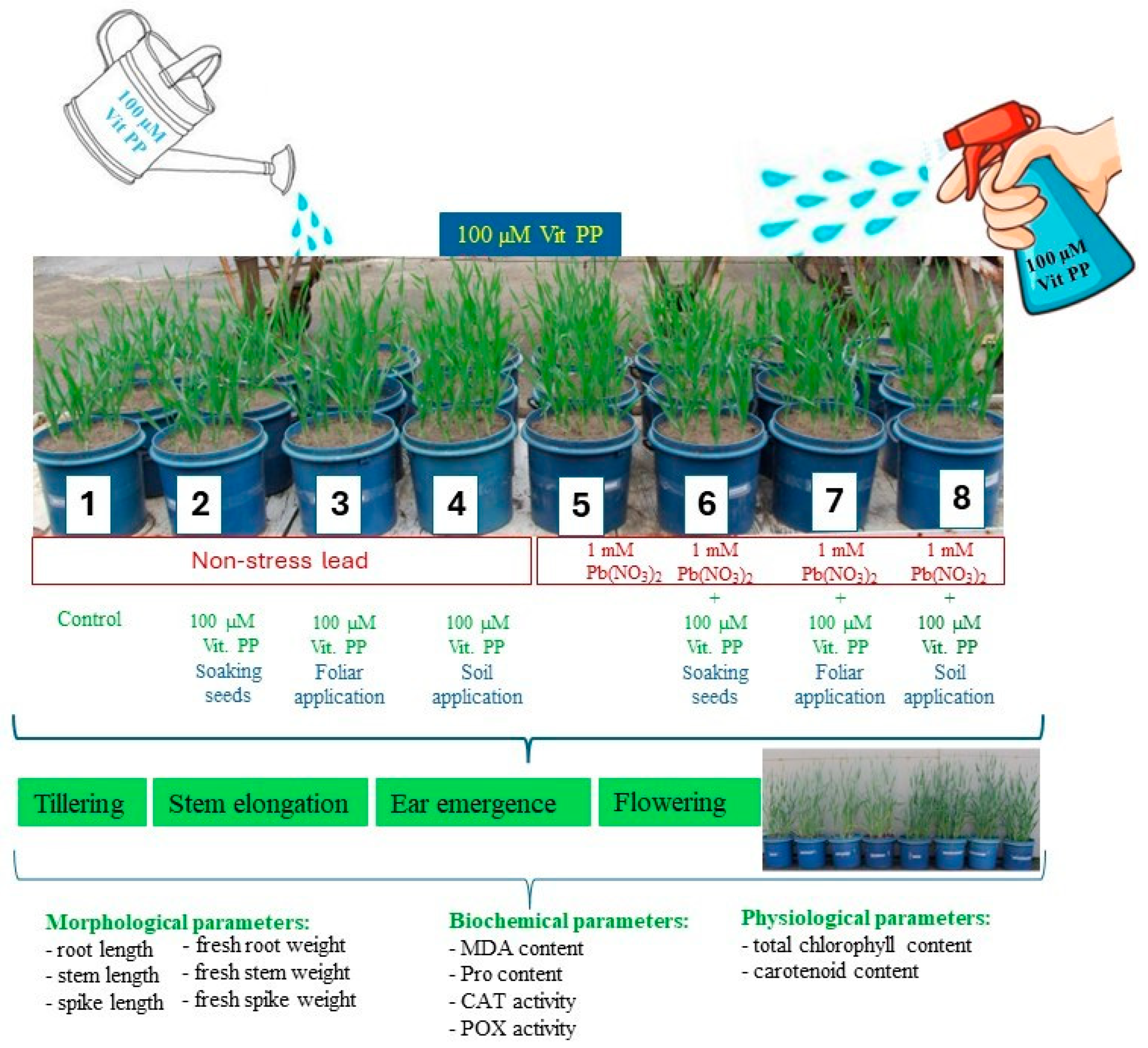
2. Materials and Methods
2.1. Experiment Site and Conditions
2.2. Determination of the Morphological Parameters
2.3. Determination of Biochemical and Physiological Parameters
2.4. Statistical Analysis of Results
3. Results
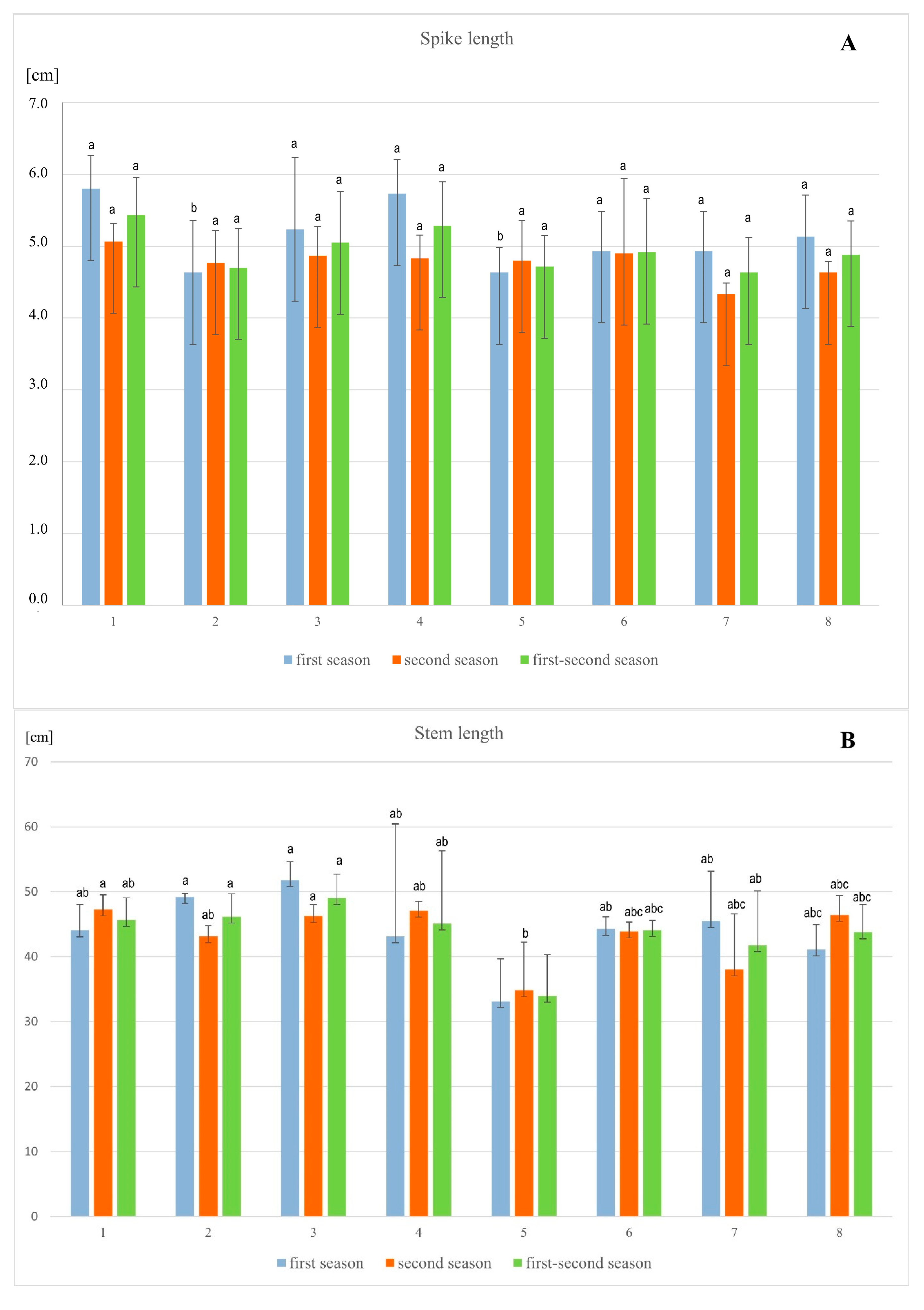
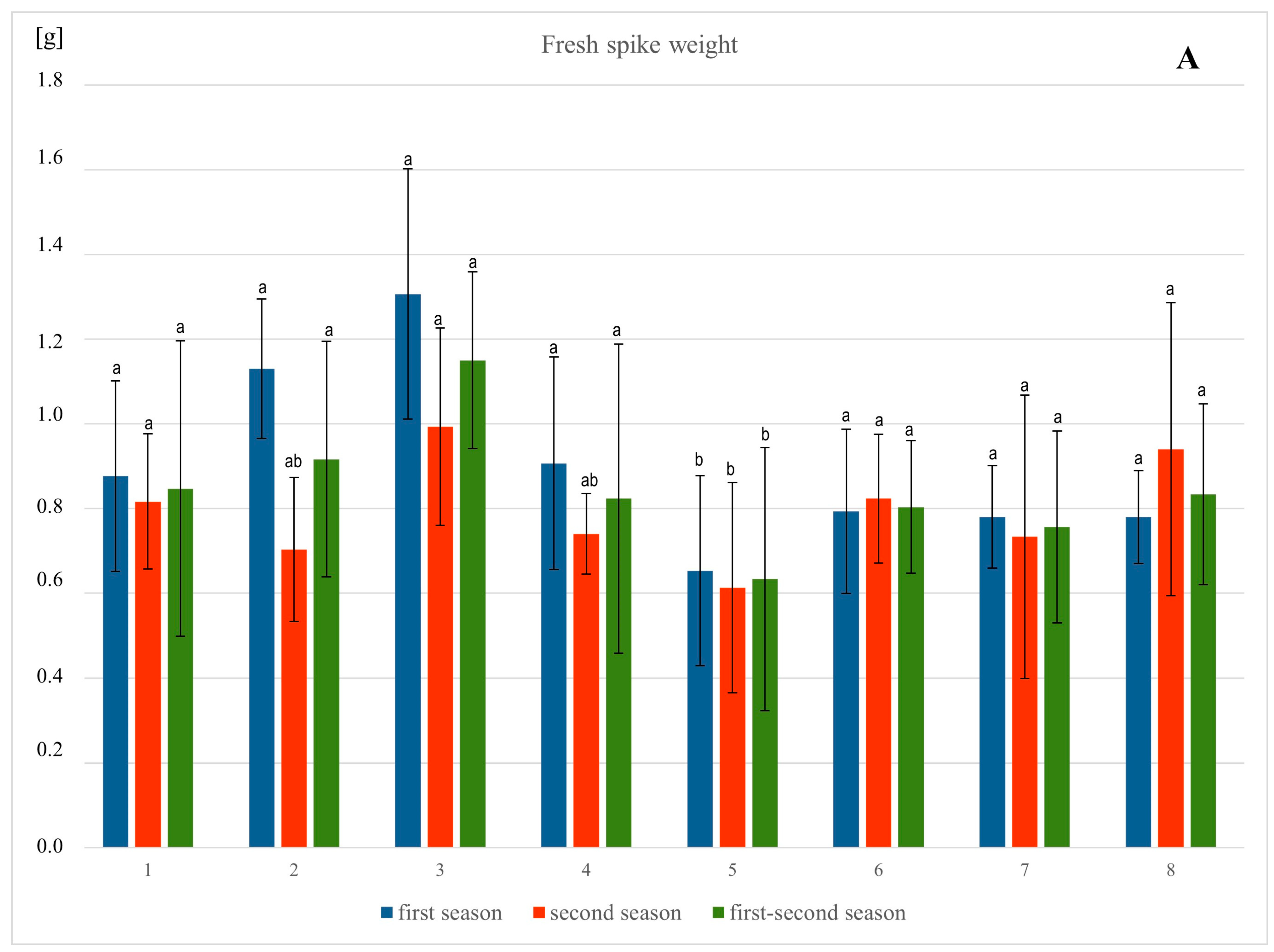
3.1. Plant Growth and Biomass
3.2. Antioxidant Enzyme Activities, Malondialdehyde, and Proline Contents
3.3. Total Chlorophyll and Carotenoid Contents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catalase [μmol∙H2O2∙g−1 FW∙min−1] (% Control) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 104.8 ± 5.49 c (100) | 100.4 ± 5.30 d (100) | 84.7 ± 2.74 c (100) | 60.7 ± 0.12 d (100) | 115.4 ± 10.26 b (100) | 110.6 ± 8.83 d (100) | 63.36 ± 5.28 c (100) | 48.9 ± 11.45 c (100) | 110.1 ± 8.71 c (100) | 105.5 ± 8.59 d (100) | 74.1 ± 12.30 c (100) | 54.8 ± 9.69 c (100) |
| 2 | 119.3 ± 5.14 bc (113.9) | 107.9 ± 7.53 cd (107.5) | 92.6 ± 4.54 c (109.3) | 67.7 ± 1.87 c (111.5) | 121.5 ± 3.12 b (105.30) | 114.4 ± 1.77 cd (103.34) | 68.9 ± 1.69 c (108.9) | 52.5 ± 4.17 c (107.3) | 120.4 ± 3.98 c (109.3) | 111.2 ± 6.02 d (105.4) | 80.8 ± 13.27 c (109.0) | 60.1 ± 8.80 c (109.7) |
| 3 | 108.1 ± 9.93 c (103.2) | 102.4 ± 1.84 d (102.02) | 85.0 ± 3.93 c (100.35) | 60.7 ± 1.97 d (100.1) | 113.2 ± 4.87 b (98.1) | 110.2 ± 5.67 d (99.6) | 61.9 ± 2.76 c (97.7) | 51.7 ± 5.39 c (105.56) | 110.6 ± 7.53 c (100.5) | 106.3 ± 5.70 d (100.7) | 73.5 ± 13.04 c (99.2) | 56.1 ± 6.15 c (102.4) |
| 4 | 109.8 ± 4.93 c (104.8) | 104.9 ± 4.27 d (104.6) | 88.8 ± 6.66 c (104.8) | 66.4 ± 1.24 cd (109.4) | 119.2 ± 7.76 b (103.3) | 121.2 ± 4.43 cd (109.5) | 67.7 ± 5.69 c (106.9) | 51.1 ± 1.88 c (104.4) | 114.5 ± 7.77 c (103.9) | 113.1 ± 9.69 d (107.2) | 78.3 ± 12.30 c (105.7) | 58.7 ± 8.52 c (107.1) |
| 5 | 150.2 ± 20.80 a (143.3) | 185.6 ± 2.34 a (184.8) | 178.4 ± 8.21 a (210.5) | 122.5 ± 2.38 a (201.8) | 170.63 ± 3.28 a (147.91 | 167.2 ± 2.04 a (151.1) | 121.6 ± 3.51 a (191.9) | 102.3 ± 2.43 a (209.1) | 160.4 ± 17.39 a (145.7) | 176.4 ± 10.14 a (167.2) | 150.0 ± 31.59 a (202.4) | 112.4 ± 11.25 a (205.1) |
| 6 | 143.6 ± 1.79 ab (137.0) | 132.9 ± 4.16 b (132.4) | 143.9 ± 6.39 b (169.9) | 90.6 ± 3.68 b (149.3) | 159.0 ± 4.95 ab (137.8) | 143.7 ± 3.73 b (129.9) | 91.4 ± 8.72 b (144.2) | 77.3 ± 7.44 b (157.9) | 151.3 ± 9.08 ab (137.4) | 138.3 ± 6.87 b (131.1) | 117.7 ± 29.61 b (158.8) | 83.9 ± 8.98 b (153.1) |
| 7 | 126.9 ± 2.17 abc (121.13) | 121.0 ± 4.48 bc (120.5) | 139.1 ± 5.75 b (164.1) | 91.2 ± 1.81 b (150.2) | 155.5 ± 6.56 ab (134.8) | 124.4 ± 8.78 c (112.4) | 83.3 ± 3.13 b (131.5) | 76.1 ± 5.3 b(155.6) | 141.2 ± 16.26 b (128.2) | 122.7 ± 3.41 c (116.3) | 111.2 ± 30.82 b (150.1) | 83.6 ± 8.96 b (152.5) |
| 8 | 122.5 ± 5.11 bc (116.9) | 126.0 ± 6.23 b (125.5) | 135.4 ± 5.52 b (159.7) | 87.9 ± 4.01 b (144.8) | 157.7 ± 4.23 ab (136.7) | 138.4 ± 11.02 b (125.1) | 94.5 ± 4.79 b (149.1) | 74.1 ± 2.09 b (151.4) | 140.1 ± 19.71 b (127.2) | 132.2 ± 7.86 b (125.3) | 114.9 ± 22.86 b (155.1) | 80.9 ± 8.07 b- (147.6) |
| Peroxidase [μM Purpurogallin∙g−1 FW∙min−1] (% Control) | ||||||||||||
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 3.7 ± 0.84 b (100) | 6.0 ± 0.51 c (100) | 8.0 ± 0.32 b (100) | 7.9 ± 1.08 c (100) | 3.6 ± 0.22 c (100) | 6.4 ± 0.31 b (100) | 7.4 ± 0.12 c (100) | 8.1 ± 1.32 b (100) | 3.7 ± 0.53 d (100) | 6.2 ± 0.43 b (100) | 7.7 ± 0.39 d (100) | 8.0 ± 0.84 d (100) |
| 2 | 4.1 ± 0.46 ab (110.4) | 6.2 ± 1.36 bc (103.7) | 8.4 ± 0.45 b (105.6) | 8.3 ± 0.61 bc (105.1) | 3.9 ± 0.22 bc (107.9) | 6.6 ± 1.24 b (102.8) | 7.6 ± 0.22 bc (103.5) | 8.3 ± 0.69 b (102.6) | 4.0 ± 0.32 cd (108.1) | 6.4 ± 0.84 b (103.2) | 8.0 ± 0.53 cd (103.9) | 8.3 ± 0.58 cd (103.7) |
| 3 | 4.1 ± 0.15 ab (109.6) | 6.2 ± 0.12 bc (102.6) | 7.9 ± 0.71 b (99.4) | 8.1 ± 0.68 bc (101.4) | 3.91 ± 0.19 bc (107.7) | 6.3 ± 0.74 b (98.2) | 7.5 ± 0.72 bc (102.1) | 8.1 ± 0.23 b (100.3) | 4.0 ± 0.15 cd (108.1) | 6.2 ± 0.48 b (100.1) | 7.7 ± 0.67 d (100.1) | 8.1 ± 0.45 d (101.2) |
| 4 | 4.1 ± 0.45 ab (108.9) | 6.1 ± 0.53 bc (102.3) | 7.9 ± 0.45 b (99.4) | 8.0 ± 0.38 c (101.6) | 3.8 ± 0.13 bc (105.2) | 6.5 ± 0.21 b (101.2) | 7.5 ± 0.71 bc (101.1) | 8.2 ± 0.44 b (100.8) | 3.9 ± 0.33 cd (105.4) | 6.3 ± 0.40 b (101.6) | 7.7 ± 0.59 d (100.1) | 8.1 ± 0.37 d (101.2) |
| 5 | 5.4 ± 0.49 a (144.8) | 8.4 ± 0.54 a (140.4) | 11.1 ± 1.03 a (138.7) | 10.9 ± 0.96 a (137.9) | 5.3 ± 0.19 a (146.38) | 8.8 ± 0.85 a (136.9) | 10.3 ± 0.82 a (139.6) | 12.1 ± 1.50 a (149.7) | 5.4 ± 0.34 a (145.9) | 8.6 ± 0.66 a (138.7) | 10.7 ± 0.95 a (138.9) | 11.5 ± 1.16 a (143.7) |
| 6 | 5.1 ± 0.49 ab (135.3) | 7.8 ± 0.29 a (130.8) | 9.6 ± 0.33 ab (120.1) | 9.9 ± 0.89 ab (125.8) | 4.6 ± 0.52 ab (126.1) | 8.1 ± 0.39 ab (125.8) | 9.1 ± 0.32 ab (123.1) | 9.9 ± 0.72 ab (122.0) | 4.8 ± 0.52 ab (129.7) | 7.9 ± 0.32 a (127.4) | 9.3 ± 0.39 b (120.8) | 9.9 ± 0.68 b (123.7) |
| 7 | 4.5 ± 0.39 ab (120.4) | 7.3 ± 0.99 abc (121.8) | 9.5 ± 1.24 ab (119.54) | 9.7 ± 0.82 abc (122.1) | 4.5 ± 0.43 b (123.2) | 7.9 ± 0.74 ab (123.5) | 8.8 ± 0.88 abc (118.8) | 9.7 ± 0.65 b (119.7) | 4.5 ± 0.28 bc (121.6) | 7.6 ± 0.85 a (122.5) | 9.2 ± 1.05 bc (119.5) | 9.7 ± 0.66 bc (121.2) |
| 8 | 4.7 ± 0.58 ab (126.6) | 7.58 ± 0.36 ab (126.3) | 9.1 ± 0.54 ab (114.0) | 9.57 ± 1.15 abc (120.5) | 4.3 ± 0.39 bc (119.9) | 7.8 ± 0.21 ab (122.7) | 8.7 ± 0.25 bc (117.5) | 9.4 ± 0.42 b (115.3) | 4.5 ± 0.49 bc (121.6) | 7.7 ± 0.29 a (124..2) | 8.9 ± 0.44 bcd (115.6) | 9.5 ± 0.78 bcd (118.7) |
| Proline [μmol∙g−1∙FW] (% Control) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 2.5 ± 0.20 a (100) | 2.6 ± 0.79 a (100) | 3.4 ± 1.44 d (100) | 3.4 ± 0.42 b (100) | 2.1 ± 0.68 a (100) | 2.3 ± 0.27 a (100) | 2.9 ± 0.13 d (100) | 2.6 ± 0.21 bc (100) | 2.3 ± 0.49 b (100) | 2.5 ± 0.57 cd (100) | 3.2 ± 0.95 c (100) | 3.0 ± 0.51 c (100) |
| 2 | 2.5 ± 0.11 a (98.7) | 2.7 ± 0.49 a (102.7) | 3.7 ± 0.18 cd (107.7) | 3.6 ± 0.38 b (106.3) | 2.1 ± 0.21 a (98.7) | 2.5 ± 0.54 a (107.2) | 3.1 ± 0.37 bcd (105.0) | 2.8 ± 0.16 b (105.5) | 2.3 ± 0.25 b (99.1) | 2.6 ± 0.49 bcd (104.8) | 3.4 ± 0.39 bc (106.3) | 3.2 ± 0.52 c (105.6) |
| 3 | 2.4 ± 0.64 a (96.93) | 2.6 ± 0.11 a (98.0) | 3.6 ± 1.13 cd (104.5) | 3.5 ± 0.36 b (102.9) | 2.0 ± 0.11 a (94.4) | 2.3 ± 0.47 a (101.4) | 3.0 ± 0.12 cd (102.4) | 2.7 ± 0.43 b (102.9) | 2.2 ± 0.47 b (96.1) | 2.5 ± 0.34 d (99.6) | 3.3 ± 0.78 bc (103.4) | 3.1 ± 0.55 c (102.6) |
| 4 | 2.5 ± 0.24 a (99.6) | 2.6 ± 0.16 a (99.9) | 3.7 ± 0.32 cd (107.0) | 3.4 ± 0.25 b (100.6) | 2.1 ± 0.17 a (96.9) | 2.3 ± 0.18 a (101.5) | 2.9 ± 0.15 d (99.6) | 2.7 ± 0.22 b (102.0) | 2.3 ± 0.30 b (98.7) | 2.5 ± 0.23 cd (100.8) | 3.3 ± 0.45 bc (103.4) | 3.0 ± 0.45 c (100.7) |
| 5 | 3.2 ± 0.89 a (130.3) | 3.9 ± 0.37 a (147.9) | 5.1 ± 0.33 a (149.5) | 5.5 ± 0.59 a (163.2) | 2.8 ± 0.15 a (135.5) | 3.3 ± 0.35 a (143.4) | 4.2 ± 0.38 a (142.6) | 3.8 ± 0.38 a (145.5) | 3.1 ± 0.60 a (133.0) | 3.6 ± 0.48 a (146.1) | 4.6 ± 0.60 a (146.2) | 4.7 ± 0.96 a (154.8) |
| 6 | 2.9 ± 0.52 a (119.9) | 3.7 ± 0.88 a (140.4) | 4.7 ± 0.14 b (138.0) | 4.9 ± 0.76 ab (145.4) | 2.8 ± 0.42 a (130.6) | 3.1 ± 0.31 a (133.9) | 3.8 ± 0.29 ab (130.2) | 3.5 ± 0.29 ab (130.2) | 2.9 ± 0.44 ab (125.2) | 3.4 ± 0.69 ab (137.6) | 4.3 ± 0.50 ab (134.3) | 4.2 ± 0.96 ab (139.2) |
| 7 | 2.7 ± 0.15 a (107.1) | 3.8 ± 0.36 a (142.2) | 4.4 ± 0.27 b (127.7) | 4.8 ± 0.33 ab (141.1) | 2.7 ± 0.21 a (129.2) | 2.9 ± 0.38 a (129.5) | 3.7 ± 0.21 ab (126.0) | 3.2 ± 0.46 ab (120.1) | 2.7 ± 0.17 ab (117.8) | 3.4 ± 0.55 ab (136.4) | 4.1 ± 0.43 abc (126.7) | 3.9 ± 0.96 b (131.5) |
| 8 | 2.8 ± 0.14 a (113.6) | 3.7 ± 0.25 a (138.5) | 4.6 ± 0.79 b (133.5) | 4.7 ± 0.18 ab (140.3) | 2.7 ± 0.22 a (127.1) | 2.9 ± 0.52 a (128.4) | 3.6 ± 0.32 abc (124.0) | 3.0 ± 0.34 ab (130.8) | 2.8 ± 0.18 ab (120.4) | 3.3 ± 0.54 abc (134.0) | 4.1 ± 0.74 abc (128.9) | 3.9 ± 0.96 b (129.2) |
| MDA [μmol∙g−1 FW] (% Control) | ||||||||||||
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 18.3 ± 1.11 b (100) | 25.3 ± 1.54 c (100) | 28.2 ± 3.68 b (100) | 30.1 ± 2.52 c (100) | 11.1 ± 1.29 c (100.0) | 24.9 ± 1.79 c (100) | 33.3 ± 1.07 b (100) | 36.5 ± 2.11 c (100) | 14.7 ± 4.08 d (100) | 25.1 ± 1.51 c (100) | 30.8 ± 3.71 b (100) | 33.3 ± 4.07 d (100) |
| 2 | 19.3 ± 1.15 b (105.2) | 26.2 ± 0.25 c (103.4) | 29.2 ± 0.69 b (103.6) | 32.1 ± 2.51 c (106.5) | 11.9 ± 1.58 bc (106.6) | 25.1 ± 2.23 c (96.5) | 36.8 ± 3.79 b (110.5) | 38.5 ± 4.09 c (105.4) | 15.5 ± 4.12 cd (105.7) | 25.6 ± 1.55 c (101.9) | 33.0 ± 4.82 b (107.3) | 35.3 ± 4.64 d (105.9) |
| 3 | 18.3 ± 0.56 b (100.1) | 25.2 ± 1.16 c (99.7) | 28.9 ± 1.69 b (102.4) | 31.1 ± 0.75 c (103.2) | 11.8 ± 1.48 bc (105.8) | 25.1 ± 2.62 c (96.5) | 34.8 ± 1.89 b (104.4) | 36.9 ± 4.23 c (101.2) | 15.1 ± 3.73 d (102.3) | 25.1 ± 1.82 c (100.1) | 31.8 ± 3.61 b (103.5) | 33.9 ± 4.21 d (102.1) |
| 4 | 18.5 ± 1.25 b (100.9) | 25.7 ± 0.92 c (101.6) | 28.5 ± 3.14 b (101.2) | 31.2 ± 1.55 c (103.8) | 11.1 ± 1.44 c (99.4) | 25.6 ± 0.63 c (98.5) | 35.2 ± 6.48 b (105.6) | 36.8 ± 3.37 c (100.8) | 14.7 ± 4.24 d (100.3) | 25.6 ± 0.71 c (101.9) | 31.9 ± 5.83 b (103.6) | 34.0 ± 3.83 d (102.2) |
| 5 | 28.9 ± 2.9 a (158.3) | 41.4 ± 1.21 a (163.4) | 45.4 ± 5.4 a (160.1) | 51.7 ± 5.12 a (171.6) | 18.6 ± 3.01 a (167.1) | 46.9 ± 4.07 a (180.8) | 62.9 ± 5.39 a (188.9) | 67.8 ± 2.19 a (185.8) | 23.8 ± 6.27 a (161.9) | 44.2 ± 4.05 a (175.6) | 54.2 ± 10.78 a (176.0) | 59.7 ± 9.52 a (179.4) |
| 6 | 26.9 ± 3.11 a (146.8) | 37.3 ± 2.32 ab (147.2) | 42.6 ± 2.62 a (150.9) | 46.6 ± 2.89 ab (154.7) | 16.9 ± 1.85 ab (151.5) | 41.6 ± 4.26 ab (160.4) | 57.5 ± 2.21 a (172.6) | 61.4 ± 5.88 ab (168.4) | 21.9 ± 5.95 ab (148.6) | 39.4 ± 3.88 b (156.9) | 50.0 ± 8.47 a (162.6) | 54.0 ± 9.13 ab (162.2) |
| 7 | 26.1 ± 3.71 a (142.6) | 33.9 ± 4.20 b (134.1) | 40.3 ± 4.51 ab (142.9) | 45.1 ± 2.52 ab (149.8) | 16.4 ± 2.62 ab (147.3) | 38.6 ± 3.34 b (148.8) | 56.2 ± 3.95 ab (168.8) | 53.7 ± 1.01 b (147.1) | 21.3 ± 6.05 ab (144.4) | 36.3 ± 4.25 b (144.3) | 48.3 ± 9.52 ab (156.9) | 49.4 ± 5.01 bc (148.3) |
| 8 | 23.2 ± 2.19 ab (126.76) | 32.7 ± 0.92 b (129.3) | 40.6 ± 5.82 ab (143.9) | 42.4 ± 4.85 b (141.0) | 15.2 ± 1.23 abc (137.1) | 38.3 ± 2.07 b (147.7) | 56.5 ± 5.12 ab (169.6) | 52.5 ± 2.21 b (143.9) | 19.2 ± 4.64 bc (130.7) | 35.5 ± 3.34 b (141.3) | 48.6 ± 10.01 ab (157.9) | 47.5 ± 6.47 c (142.6) |
| Total Chlorophyll [μg·g−1 FW] (% Control) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 122.7 ± 5.42 ab (100) | 119.5 ± 12.69 ab (100) | 130.4 ± 15.65 ab (100) | 107.2 ± 3.36 a (100) | 120.4 ± 2.46 ab (100) | 108.8 ± 13.95 abc (100) | 130.2 ± 6.64 a (100) | 104.9 ± 10.84 ab (100) | 121.6 ± 3.95 ab (100) | 114.2 ± 10.73 ab (100) | 130.3 ± 10.75 a (100) | 106.1 ± 7.28 a (100) |
| 2 | 131.6 ± 4.67 a (107.3) | 125.4 ± 2.73 ab (104.9) | 132.8 ± 3.54 ab (101.9) | 111.8 ± 2.93 a (104.3) | 123.2 ±14.96 ab (102.3) | 112.5 ± 7.38 ab (103.4) | 133.5 ± 4.06 a (102.5) | 106.7 ± 1.66 ab (101.6) | 127.4 ± 10.93 a (104.10 | 118.9 ± 8.64 a (104.1) | 133.1 ± 3.43 a (102.1) | 109.2 ± 3.52 a (102.9) |
| 3 | 133.8 ± 3.45 a (109.1) | 125.9 ± 23.28 ab (105.3) | 135.8 ± 5.84 ab (104.2) | 108.6 ± 9.8 a (101.3) | 128.8 ± 1.69 a (106.9) | 120.7 ± 9.31 a (110.9) | 146.9 ± 17.28 a (112.9) | 112.1 ± 16.02 ab (106.8) | 131.3 ± 8.17 a (107.9) | 123.3 ± 16.11 a (107.9 | 141.4 ± 13.04 a (108.5) | 110.3 ± 12.05 a (103.9) |
| 4 | 135.3 ± 3.78 a (110.3) | 129.7 ± 5.26 a (108.5) | 140.8 ± 5.31 a (108.0) | 115.8 ± 2.55 a (107.9 | 130.0 ± 6.83 a (107.9) | 120.6 ± 1.44 a (110.9) | 143.1 ± 7.46 a (109.9) | 115.8 ± 5.72 a (110.3) | 132.7 ± 5.72 a (109.1) | 125.2 ± 6.03 a (109.6) | 141.9 ± 5.92 a (108.9) | 115.8 ± 3.96 a (109.1) |
| 5 | 99.2 ± 4.18 c (80.8) | 86.7 ± 0.97 c (72.59) | 92.9 ± 4.92 c (71.2) | 71.2 ± 6.16 c (66.4) | 94.5 ± 4.56 c (78.4) | 76.9 ± 3.68 d (70.73) | 88.0 ± 10.42 b (67.6) | 66.2 ± 8.99 c (63.1) | 96.8 ± 4.67 c (79.7) | 81.8 ± 5.87 d (71.6) | 90.4 ± 7.76 b (69.4) | 68.7 ± 7.43 c (64.7) |
| 6 | 104.9 ± 2.77 c (85.5) | 97.6 ± 10.76 bc (81.7) | 104.6 ± 13.34 bc (80.27) | 72.1 ± 1.24 c (67.2) | 102.1 ± 5.51 bc (84.7) | 85.5 ± 1.78 d (78.6) | 91.4 ± 6.83 b (70.2) | 82.9 ± 5.63 bc (78.9) | 103.5 ± 4.19 c (85.1) | 91.6 ± 9.56 cd (80.2) | 98.0 ± 11.92 b (75.2) | 77.5 ± 6.96 bc (73.0) |
| 7 | 107.4 ± 8.88 c (87.5) | 98.2 ± 3.49 bc (82.1) | 114.9 ± 11.60 abc (88.2) | 76.6 ± 3.37 bc (71.4) | 105.0 ± 13.67 abc (87.2) | 91.4 ± 6.45 cd (83.9) | 96.6 ± 14.43 b (74.2) | 84.7 ± 12.76 bc (80.7) | 106.2 ± 10.39 c (87.3) | 94.8 ± 5.94 cd (83.0) | 105.7 ± 15.42 b (81.1) | 80.7 ± 9.45 bc (76.1) |
| 8 | 111.1 ± 3.37 bc (90.6) | 101.6 ± 11.23 abc (85.0) | 116.1 ± 20.93 abc (89.1) | 81.1 ± 1.48 b (75.6) | 108.4 ± 3.02 abc (89.9) | 95.4 ±3.36 bcd (87.7) | 97.1 ± 5.21 b (74.6) | 90.0 ± 16.27 abc (85.8) | 109.7 ± 3.23 bc (89.6) | 98.5 ± 8.16 bc (86.2 | 106.6 ± 17.13 b (81.8) | 85.5 ± 11.44 b (80.6) |
| Carotenoids [μg·g−1FW] (% Control) | ||||||||||||
| Combination | First Season | Second Season | Synthesis First—Second Season | |||||||||
| Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | Tillering | Stem Elongation | Ear Emergence | Flowering | |
| 1 | 44.3 ± 3.93 ab (100) | 42.8 ± 3.56 a (100) | 47.7 ± 4.37 ab (100) | 37.4 ± 2.24 ab (100) | 40.6 ± 2.48 ab (100) | 39.8 ± 8.61 a (100) | 36.8 ± 3.94 a (100) | 30.9 ± 6.47 ab (100) | 42.5 ± 3.57 abc (100) | 41.3 ± 6.13 a (100) | 42.2 ± 7.03 ab (100) | 34.2 ± 5.45 ab (100) |
| 2 | 46.3 ± 1.90 ab (104.6) | 45.5 ± 1.65 a (106.3) | 49.9 ± 1.62 a (104.6) | 39.6 ± 5.57 a (105.6) | 42.3 ± 6.49 ab (104.2) | 40.1 ± 2.49 a (100.7) | 38.0 ± 2.32 a (103.3) | 32.4 ± 5.01 ab (104.8) | 44.3 ±4.81 abc (104.2) | 42.8 ± 3.53 a (103.6) | 43.9 ± 6.75 a (104.0) | 35.9 ± 5.62 a (104.9) |
| 3 | 47.9 ± 1.83 a (108.0) | 44.1 ± 1.96 a (102.9) | 37.9 ± 4.01 ab (101.3) | 37.4 ± 9.11 ab (101.6) | 43.6 ± 3.23 ab (107.4) | 40.6 ± 8.91 a (101.9) | 38.3 ± 2.53 a (104.1) | 31.6 ± 3.91 ab (102.4) | 45.7 ± 3.30 ab (107.5) | 42.3 ± 6.07 a (102.4) | 43.4 ± 8.15 ab (102.8) | 34.8 ± 6.18 ab (101.7) |
| 4 | 47.6 ± 1.12 a (107.4) | 44.4 ± 3.35 a (103.6) | 48.5 ± 2.32 ab (101.7) | 38.4 ± 3.39 a (102.7) | 44.8 ± 4.26 a (110.3) | 41.8 ± 5.24 a (105.1) | 38.0 ± 2.20 a (103.4) | 34.2 ± 2.63 a (110.7) | 46.2 ± 3.18 a (108.7) | 43.1 ± 4.17 a (104.3) | 43.3 ± 6.07 ab (102.6) | 36.3 ± 4.94 a (106.1) |
| 5 | 34.9 ± 2.48 c (78.9) | 29.5 ± 3.03 bc (68.9) | 33.8 ± 7.94 b (70.9) | 25.8 ± 5.74 c (68.8) | 31.1 ± 4.09 b (76.5) | 29.1 ± 2.51 b (73.2) | 26.3 ± 3.06 b (71.5) | 22.9 ± 1.57 b (74.1) | 33.0 ± 3.70 e (77.6) | 29.3 ± 2.49 b (70.2) | 30.1 ± 6.07 c (71.3) | 24.4 ± 3.57 c (71.3) |
| 6 | 38.8 ± 3.02 bc (87.6) | 30.1 ± 5.21 bc (70.3) | 37.7 ± 2.98 ab (79.1) | 26.8 ± 2.71 c (71.6) | 33.6 ± 7.21 ab (82.8) | 34.6 ± 6.62 ab (86.9) | 31.9 ± 4.46 ab (86.8) | 25.9 ± 3.65 ab (83.9) | 36.2 ± 5.71 de (85.1) | 32.4 ± 5.87 b (78.4) | 34.9 ± 4.62 bc (82.7) | 26.4 ± 3.09 c (77.2) |
| 7 | 40.2 ± 4.28 abc (90.7) | 32.7 ± 2.49 b (76.4) | 42.1 ± 5.52 ab (88.2) | 29.3 ± 1.89 abc (78.3) | 35.2 ± 2.98 ab (86.6) | 36.0 ± 4.02 ab (90.6) | 34.7 ± 2.43 ab (94.3) | 27.7 ± 3.45 ab (89.6) | 37.7 ± 4.29 cde (88.7) | 34.4 ± 3.51 ab (83.3) | 38.3 ± 5.54 abc (90.7 | 28.5 ± 2.61 bc (83.3) |
| 8 | 40.6 ± 2.87 abc (91.6) | 33.5 ± 2.79 b (78.2) | 40.4 ± 5.45 ab (84.8) | 27.8 ± 2.06 bc (74.4) | 37.2 ± 3.09 ab (91.7) | 36.4 ± 4.87 ab (91.5) | 34.6 ± 5.79 ab (94.1) | 27.5 ± 2.21 ab (89.2) | 38.9 ± 3.24 cde (91.5) | 34.9 ± 3.89 ab (84.5) | 37.5 ± 5.96 abc (88.9) | 27.7 ± 1.91 bc (81.0) |
4. Discussion
4.1. Morphological Parameters
4.2. Biochemical Parameters
4.3. Physiological Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abhilash, P.C.; Tripathi, V.; Edrisi, S.A.; Dubey, R.K.; Bakshi, M.; Dubey, P.K.; Singh, H.B.; Ebbs, S.D. Sustainability of crop production from polluted lands. Energ. Ecol. Environ. 2016, 1, 54–65. [Google Scholar] [CrossRef]
- Fróna, D.; Szenderák, J.; Harangi-Rákos, M. The Challenge of Feeding the World. Sustainability 2019, 11, 5816. [Google Scholar] [CrossRef]
- Lambin, E.F.; Meyfroidt, P. Global land use change, economic globalization, and the looming land scarcity. Proc. Natl. Acad. Sci. USA 2011, 108, 3465–3472. [Google Scholar] [CrossRef]
- Vendruscolo, E.P.; Seleguin, A. Effects of vitamin pre-sowing treatment on sweet maize seedlings irrigated with saline water. Acta Agronómica 2020, 69, 20–25. [Google Scholar] [CrossRef]
- Vasilachi, I.C.; Stoleru, V.; Gavrilescu, M. Analysis of Heavy Metal Impacts on Cereal Crop Growth and Development in Contaminated Soils. Agriculture 2023, 13, 1983. [Google Scholar] [CrossRef]
- Angon, P.B.; Islam, M.S.; Shreejana, K.C.; Das, A.; Anjum, N.; Poudel, A.; Suchi, A.K. Sources, effects and present perspectives of heavy metals contamination: Soil, plants and human food chain. Heliyon 2024, 10, e28357. [Google Scholar] [CrossRef]
- Uchimiya, M.; Bannon, D.; Nakanishi, H.; McBride, M.B.; Williams, M.A.; Yoshihara, T. Chemical speciation, plant uptake, and toxicity of heavy metals in agricultural soils. J. Agric. Food Chem. 2020, 68, 12856–12869. [Google Scholar] [CrossRef]
- Yan, X.; Gao, D.; Zhang, F.; Zeng, C.; Xiang, W.; Zhang, M. Relationships between Heavy Metal Concentrations in Roadside Topsoil and Distance to Road Edge Based on Field Observations in the Qinghai-Tibet Plateau, China. Int. J. Environ. Res. Public Health 2013, 10, 762–775. [Google Scholar] [CrossRef]
- Nicholson, F.A.; Smith, S.R.; Alloway Carlton-Smithd, C.; Chambers, B.J. An Inventory of Heavy Metals Inputs to Agricultural Soils in England and Wales. Sci. Total Environ. 2003, 311, 205–219. [Google Scholar] [CrossRef]
- Gaya, U.; Ikechukwu, S. Heavy metal contamination of selected spices obtained from Nigeria. J. Appl. Sci. Environ. Manag. 2016, 20, 681–688. [Google Scholar] [CrossRef]
- Sharma, R.; Dubey, R. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Tiwari, S.; Tripathi, I.P.; Tiwari, H.L. Effects of Lead on Environment. Int. J. Emerg. Res. Manag. Technol. 2013, 2, 23–45. [Google Scholar]
- Khan, M.; Rolly, N.K.; Al Azzawi, T.N.I.; Imran, M.; Mun, B.-G.; Lee, I.-J.; Yun, B.-W. Lead (Pb)-Induced Oxidative Stress Alters the Morphological and Physio-Biochemical Properties of Rice (Oryza sativa L.). Agronomy 2021, 11, 409. [Google Scholar] [CrossRef]
- Aslam, M.; Aslam, A.; Sheraz, M.; Ali, B.; Ulhassan, Z.; Najeeb, U.; Zhou, W.; Gill, R.A. Lead Toxicity in Cereals: Mechanistic Insight Into Toxicity, Mode of Action, and Management. Front. Plant Sci. 2021, 11, 587785. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Ali, A.; Kour, N.; Bornhorst, J.; AlHarbi, K.; Rinklebe, J.; Abd El Moneim, D.; Ahmad, P.; Chung, Y.S. Heavy Metal Induced Oxidative Stress Mitigation and ROS Scavenging in Plants. Plants 2023, 12, 3003. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, U.; Khan, S.M.; Khalid, N.; Ahmad, Z.; Jehangir, S.; Fatima Rizvi, Z.; Lho, L.H.; Han, H.; Raposo, A. Detoxifying the heavy metals: A multipronged study of tolerance strategies against heavy metals toxicity in plants. Front. Plant Sci. 2023, 12, 1154571. [Google Scholar] [CrossRef]
- Jorjani, S.; Pehlivan Karakaş, F. Physiological and Biochemical Responses to Heavy Metals Stress in Plants. Int. J. Second. Metab. 2024, 11, 169–190. [Google Scholar] [CrossRef]
- Sadak, M.S. Physiological role of yeast extract and nicotinamide on Pisum sativum L. plants under heat stress. Int. J. Pharm. Tech. Res. 2016, 9, 170–178. [Google Scholar]
- El-Bassiouny, H.M.S.; Abd El-Monem, A.A.; Abdallah, M.M.S.; Soliman, K.M. Role of arbuscular mycorrhiza, α-tocopherol and nicotinamide on the nitrogen containing compounds and adaptation of sunflower plant to water stress. Biosci. Res. 2018, 15, 2068–2088. [Google Scholar]
- Feng, D.; Wang, R.; Sun, X.; Liu, L.; Liu, P.; Tang, J.; Zhang, C.; Liu, H. Heavy metal stress in plants: Ways to alleviate with exogenous substances. Sci. Total Environ. 2023, 897, 165397. [Google Scholar] [CrossRef] [PubMed]
- Soares, T.F.S.N.; Dias, D.C.F.D.S.; Oliveira, A.M.S.; Ribeiro, D.M.; Dias, L.A.D.S. Exogenous brassinosteroids increase lead stress tolerance in seed germination and seedling growth of Brassica juncea L. Ecotoxicol. Environ. 2020, 193, 110296. [Google Scholar] [CrossRef] [PubMed]
- Guedes, F.; Maia, C.F.; Silva, B.; Batista, B.L.; Alyemeni, M.N.; Ahmad, P.; Lobato, A. Exogenous 24-epibrassinolide stimulates root protection, and leaf antioxidant enzymes in lead stressed rice plants: Central roles to minimize Pb content and oxidative stress. Environ. Pollut. 2021, 280, 116992. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Ahmed, S.; Yasin, N.A.; Sardar, R.; Hussaan, M.; Gaafar, A.-R.Z.; Haider, F.U. 28-Homobrassinolide Primed Seed Improved Lead Stress Tolerance in Brassica rapa L. through Modulation of Physio-Biochemical Attributes and Nutrient Uptake. Plants 2023, 12, 3528. [Google Scholar] [CrossRef]
- Emamverdian, A.; Khalofah, A.; Pehlivan, N.; Zia-Ur-Rehman, M.; Li, Y.; Zargar, M. Exogenous application of jasmonates and brassinosteroids alleviates lead toxicity in bamboo by altering biochemical and physiological attributes. Environ. Sci. Pollut. Res. Int. 2024, 31, 7008–7026. [Google Scholar] [CrossRef]
- Piotrowska-Niczyporuk, A.; Bajguz, A.; Kotowska, U.; Zambrzycka-Szelewa, E.; Sienkiewicz, A. Auxins and cytokinins regulate phytohormone homeostasis and thiol-mediated detoxification in the green alga Acutodesmus obliquus exposed to lead stress. Sci. Rep. 2020, 10, 10193. [Google Scholar] [CrossRef]
- Arshad, T.; Maqbool, N.; Javed, F.; Wahid, A.; Arshad, M.U. Enhancing the defensive mechanism of lead affected barley (Hordeum vulgare L.) genotypes by exogenously applied salicylic acid. J. Agric. Sci. 2017, 9, 139–146. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Matin, M.A.; Fardus, J.; Hasanuzzaman, M.; Hossain, M.S.; Parvin, K. Foliar application of salicylic acid improves growth and yield attributes by upregulating the antioxidant defense system in Brassica campestris plants grown in lead-amended soils. Acta Agrobot. 2019, 72, 1765. [Google Scholar] [CrossRef]
- Bali, S.; Kaur, P.; Kohli, S.K.; Ohri, P.; Thukral, A.K.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Jasmonic acid induced changes in physio-biochemical attributes and ascorbate-glutathione pathway in Lycopersicon esculentum under lead stress at different growth stages. Sci. Total Environ. 2018, 645, 1344–1360. [Google Scholar] [CrossRef]
- Khan, I.; Iqbal, M.; Ashraf, M.Y.; Ashraf, M.A.; Ali, S. Organic chelants-mediated enhanced lead (Pb) uptake and accumulation is associated with higher activity of enzymatic antioxidants in spinach (Spinacea oleracea L.). J. Hazard. Mater. 2016, 317, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Saman, R.U.; Shahbaz, M.; Maqsood, M.F.; Lili, N.; Zulfiqar, U.; Haider, F.U.; Naz, N.; Shahzad, B. Foliar Application of Ethylenediamine Tetraacetic Acid (EDTA) Improves the Growth and Yield of Brown Mustard (Brassica juncea) by Modulating Photosynthetic Pigments, Antioxidant Defense, and Osmolyte Production under Lead (Pb) Stress. Plants 2023, 12, 115. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Ahmed, S.; Yasin, N.A.; Wahid, A.; Sardar, R. Exogenous application of glutathione enhanced growth, nutritional orchestration and physiochemical characteristics of Brassica oleracea L. under lead stress. Physiol. Mol. Biol. Plants 2023, 29, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Alamri, S.A.; Siddiqui, M.H.; Al-Khaishany, M.Y.Y.; Khan, M.; Ali, H.M.; Alaraidh, I.A.; Alsahli, A.A.; Al-Rabiah, H.; Mateen, M. Ascorbic acid improves the tolerance of wheat plants to lead toxicity. J. Plant Interact. 2018, 13, 409–419. [Google Scholar] [CrossRef]
- Sędzik-Wójcikowska, M.; Smolik, B.; Krupa-Małkiewicz, M. Effect of nicotinamide in alleviating stress caused by lead in spring barley seedling. J. Elem. 2019, 24, 281–291. [Google Scholar] [CrossRef]
- Sędzik-Wójcikowska, M.; Krupa-Małkiewicz, M.; Smolik, B. The Effect of Use of the Biologically Active Substances in Alleviating the Stress Caused by Lead in Barley Seedling on the Basis of Biochemical and Physiological Parameters. J. Ecol. Eng. 2023, 24, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.I.; Lee, T.G.; Lee, J.; Chae, M.J.; Lee, E.J.; Kim, M.S.; Jung, G.B.; Emmanuel, A.; Jeon, S.; Lee, B.R. Foliar-Applied Glutathione Mitigates Cadmium-Induced Oxidative Stress by Modulating Antioxidant-Scavenging, Redox-Regulating, and Hormone-Balancing Systems in Brassica napus. Front. Plant Sci. 2021, 12, 700413. [Google Scholar] [CrossRef] [PubMed]
- Yıldız, M.; Terzi, H. Exogenous cysteine alleviates chromium stress via reducing its uptake and regulating proteome in roots of Brassica napus L. seedlings. S. Afr. J. Bot. 2021, 139, 114–121. [Google Scholar] [CrossRef]
- Vendruscolo, E.P.; Rodrigues, A.H.A.; Oliveira, P.R.; Leitão, R.A.; Campos, L.F.C.; Seleguini, A.; Lima, S.F. Exogenous application of vitamins in upland rice. Rev. Agric. Neotrop. 2019, 6, 1–6. [Google Scholar] [CrossRef]
- Gakière, B.; Hao, J.; de Bont, L.; Pétriacq, P.; Nunes-Nesi, A.; Fernie, A.R. NAD+ Biosynthesis and Signaling in Plants. Crit. Rev. Plant Sci. 2018, 37, 259–307. [Google Scholar] [CrossRef]
- Farooq, T.H.; Bukhari, M.A.; Irfan, M.S.; Rafay, M.; Shakoor, A.; Rashid, M.H.U.; Lin, Y.; Saqib, M.; Malik, Z.; Khurshid, N. Effect of Exogenous Application of Nicotinic Acid on Morpho-Physiological Characteristics of Hordeum vulgare L. under Water Stress. Plants 2022, 11, 2443. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, S.; Chen, X.; Liu, R.; Zhu, J.; Shi, L.; Ren, A.; Zhao, M.W. NAD+-dependent Glsirt1 has a key role on secondary metabolism in Ganoderma lucidum. Microbiol. Res. 2022, 258, 126992. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant Secondary Metabolites as Defense Tools against Herbivores for Sustainable Crop Protection. Int. J. Mol. Sci. 2022, 28, 2690. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.A.; Javed, F.; Wahid, A.; Sadia, B. Alleviating effect of exogenous application of ascorbic acid on growth and mineral nutrients in cadmium stressed barley (Hordeum vulgare) seedlings. Int. J. Agric. Biol. 2016, 18, 73–79. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, A.; da Silva, J.A.T.; Fujita, M. Molecular Mechanism of Heavy Metal Toxicity and Tolerance in Plants: Central Role of Glutathione in Detoxification of Reactive Oxygen Species and Methylglyoxal and in Heavy Metal Chelation. J. Bot. 2012, 2012, 872875. [Google Scholar] [CrossRef]
- Lück, H. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: New York, NY, USA, 1963. [Google Scholar]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Bates, L.S. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Sudhakar, C.; Lakshim, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl Salinity. Plant Sci. 2001, 161, 613–619. [Google Scholar] [CrossRef]
- Arnon, D.J.; Allen, M.B.; Whatley, F. Photosynthesis by isolated chloroplast. Biochim. Biophys. Acta 1956, 20, 449–461. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and bof leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Hager, A.; Mayer-Berthenrath, T. Isolation and quantitative determination of carotenoids and chlorophylls of leaves, algaeand isolated chloroplasts using thin-layer chromatographic methods. Planta 1966, 69, 198–217. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-Q. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Kumar, V.; Tripathi, S.; Sharma, P. Heavy metal phytoextraction potential of native weeds and grasses from endocrine-disrupting chemicals rich complex distillery sludge and their histological observations during in-situ phytoremediation. Ecol. Eng. 2018, 111, 143–156. [Google Scholar] [CrossRef]
- Verma, S.; Dubey, R.S. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2023, 164, 645–655. [Google Scholar] [CrossRef]
- Hashem, H.A.; El-Sherif, N.A. Exogenous Jasmonic Acid Induces Lead Stress Tolerance in Kidney Bean (Phaseolus vulgaris L.) by Changing Amino Acid Profile and Stimulating Antioxidant Defense System. Jordan J. Biol. Sci. 2019, 12, 345–353. [Google Scholar]
- Afzaal, Z.; Hussain, I.; Ashraf, M.A.; Rasheed, R.; Javed, M.T.; Ansari, M.; Anwer, S.; Iqbal, M. Lead induced modulation in growth, chlorophyll pigment, nutrient uptake, antioxidant enzyme regulation, gene expression and fruit quality in two tomato cultivars. Int. J. Agric. Biol. 2020, 24, 1732–1744. [Google Scholar] [CrossRef]
- Hossain, B.; Akhtar, M. Growth and yield of barley (Hordeum vulgare L.) as affected by irrigation, sowing method and phosphorus level. Acad. J. Agric. Res. 2014, 2, 30–35. [Google Scholar]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Roumet, C.; Comas, L.H.; Weemstra, M.; Bengough, A.G.; Rewald, B.; Bardgett, R.D.; De Deyn, G.B.; Johnson, D.; Klimešová, J.; et al. Root traits as drivers of plant and ecosystem functioning: Current understanding, pitfalls and future research needs. New Phytol. 2021, 232, 1123–1158. [Google Scholar] [CrossRef]
- Dong, D.; Zhao, X.; Hua, X.; Liu, J.; Gao, M. Investigation of the potential mobility of Pb, Cd and Cr (VI) from moderately contaminated farmland soil to groundwater in Northeast, China. J. Hazard. Mater. 2009, 162, 1261–1268. [Google Scholar] [CrossRef]
- de Lima, S.F.; Vendruscolo, E.P.; Alves, V.C.D.; Arguelho, J.C.; de Abreu Pião, J.; de Castro Seron, C.; Martins, M.B.; Witt, T.W.; Serafim, G.M.; Merquides, L.M. Nicotinamide as a biostimulant improves soybean growth and yield. Open Agric. 2024, 9, 20220259. [Google Scholar] [CrossRef]
- Bassuony, F.M.; Hassanein, R.A.; Baraka, D.M.; Khalil, R.R. Physiological Effects of Nicotinamide and Ascorbic Acid on Zea mays Plant Grown Under Salinity Stress II-Changes in Nitrogen Constituents, Protein Profiles, Protease Enzyme and Certain Inorganic Cations. Aust. J. Basic Appl. Sci. 2008, 2, 350–359. [Google Scholar]
- Mohamed, M.H.; Badr, E.A.; Sadak, M.S.; Khedr, H.H. Effect of garlic extract, ascorbic acid and nicotinamide on growth, some biochemical aspects, yield and its components of three faba bean (Vicia faba L.) cultivars under sandy soil conditions. Bull. Natl. Res. Cent. 2020, 44, 100. [Google Scholar] [CrossRef]
- Sadak, M.S.; Rady, M.; Badr, N.M.; Gaballah, M.S. Increasing sunflower salt tolerance using nicotinamide and α-tocopherol. Int. J. Acad. Res. 2010, 2, 263–270. [Google Scholar]
- Kirkland, J.B.; Meyer-Ficca, M.L. Niacin. Adv. Food Nutr. Res. 2018, 83, 83–149. [Google Scholar] [CrossRef] [PubMed]
- Hathout, T.A. Diverse effects of uniconazole and nicotinamide on germination, growth, endogenous hormones and some enzymic activities of peas. Egypt. J. Physiol. Sci. 1995, 19, 77–95. [Google Scholar]
- Ali, R.M. Effect of Nicotinic Acid and Nicotinamide Adenine Dinucleotide on Growth and Content of Oil, Glycerol and Ricinine Alkaloids of Salinity stressed Bicinus communis L. Phyton 2002, 42, 269–277. [Google Scholar]
- Abdelhamid, M.A.; Sadak Mervat, S.H.; Schmidhalter, U.; El-Saady, A.M. Interactive effects of salinity stress and nicotinamide on physiological and biochemical parameters of faba bean plant. Acta Biol. Colomb. 2013, 18, 499–510. [Google Scholar]
- Khurshid, N.; Bukhari, M.A.; Ahmad, T.; Ahmad, Z.; Jatoi, W.N.; Abbas, S.M.; Latif, A.; Raza, A.; Aurangzaib, M.; Hashem, A.; et al. Exogenously applied nicotinic acid alleviates drought stress by enhancing morpho-physiological traits and antioxidant defense mechanisms in wheat. Ecotoxicol. Environ. Saf. 2023, 263, 115350. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Rachoski, M.; Gazquez, A.; Calzadilla, P.; Bezus, R.; Rodriguez, A.; Ruiz, O.; Menendez, A.; Maiale, S. Chlorophyll fluorescence and lipid peroxidation changes in rice somaclonal lines subjected to salt stress. Acta Physiol. Plant. 2015, 37, 117. [Google Scholar] [CrossRef]
- Navabpour, S.; Yamchi, A.; Bagherikia, S.; Kafi, H. Lead-induced oxidative stress and role of antioxidant defense in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2020, 26, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.K.; Dey, J.; Patra, S.; Pothal, D. Changes in the antioxidative enzyme activities and lipid peroxidation in wheat seedlings exposed to cadmium and lead stress. Braz. J. Plant Physiol. 2007, 19, 53–60. [Google Scholar] [CrossRef]
- Jiang, N.; Luo, X.; Zeng, J.; Yang, Z.R.; Zheng, L.Y.; Wang, S. Lead toxicity induced growth and antioxidant responses in Luffa cylindrica seedlings. Int. J. Agric. Biol. 2010, 12, 205–210. [Google Scholar]
- Yang, Y.; Zhang, Y.; Wei, X.; You, J.; Wang, W.; Lu, J.; Shi, R. Comparative antioxidative responses and proline metabolism in two wheat cultivars under short term lead stress. Ecotoxicol. Environ. Saf. 2011, 74, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Cândido, G.S.; Lima, F.R.; Vasques, I.C.; Souza, K.R.; Martins, G.C.; Pereira, P.; Engelhardt, M.M.; Reis, R.H.C.; Marques, J.J. Lead effects on sorghum and soybean physiology in oxisols. Arch. Agron. Soil Sci. 2020, 67, 260–274. [Google Scholar] [CrossRef]
- Pirzadah, T.B.; Malik, B.; Tahir, I.; Hakeem, K.R.; Alharby, H.F.; Rehman, R.U. Lead toxicity alters the antioxidant defense machinery and modulate the biomarkers in Tartary buckwheat plants. Int. Biodeterior. Biodegrad. 2020, 151, 104992. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.L.; Zhang, J.; Zheng, L.; Chen, D.; Wu, Z.; Shaheen, S.M.; Rinklebe, J.; Ok, Y.S.; Wang, H.; et al. Coconut-fiber biochar reduced the bioavailability of lead but increased its translocation rate in rice plants: Elucidation of immobilization mechanisms and significance of iron plaque barrier on roots using spectroscopic techniques. J. Hazard. Mater. 2020, 389, 122117. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhao, N. Oxidative stress and change in plant metabolism of maize (Zea mays L.) growing in contaminated soil with elemental sulfur and toxic effect of zinc. Plant Soil Environ. 2011, 57, 34–39. [Google Scholar] [CrossRef]
- Woźny, A.; Przybył, K. Komórki Roślinne w Warunkach Stresu; Wydawnictwo Naukowe Uniwersytetu im. Adama Mickiewicza w Poznaniu: Poznań, Poland, 2004. [Google Scholar]
- Özturk, L.; Demir, Y. In vivo and vitro protective role of proline. Plant Growth Regul. 2002, 38, 259–264. [Google Scholar] [CrossRef]
- Matysik, J.; Alia Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 82, 525–532. [Google Scholar]
- Rehman, M.Z.; Rizwan, M.; Ali, S.; Sabir, M.; Sohail, M.I. Contrasting effects of organic and inorganic amendments on reducing lead toxicity in wheat. Bull. Environ. Contam. Toxicol. 2017, 99, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Terry, N.; Huston, R.P. Limiting Factors in Photosynthesis. Plant Physiol. 1982, 10, 1541–1543. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smolik, B.; Sędzik-Wójcikowska, M. Examining Nicotinamide Application Methods in Alleviating Lead-Induced Stress in Spring Barley. Agronomy 2024, 14, 1314. https://doi.org/10.3390/agronomy14061314
Smolik B, Sędzik-Wójcikowska M. Examining Nicotinamide Application Methods in Alleviating Lead-Induced Stress in Spring Barley. Agronomy. 2024; 14(6):1314. https://doi.org/10.3390/agronomy14061314
Chicago/Turabian StyleSmolik, Beata, and Maja Sędzik-Wójcikowska. 2024. "Examining Nicotinamide Application Methods in Alleviating Lead-Induced Stress in Spring Barley" Agronomy 14, no. 6: 1314. https://doi.org/10.3390/agronomy14061314
APA StyleSmolik, B., & Sędzik-Wójcikowska, M. (2024). Examining Nicotinamide Application Methods in Alleviating Lead-Induced Stress in Spring Barley. Agronomy, 14(6), 1314. https://doi.org/10.3390/agronomy14061314