Evaluation of the Aluminum (Al3+) Tolerance of Camelina and the Potential for Using Melatonin or Nano-Selenium to Alleviate Al3+-Induced Stress in Camelina

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Source and Reagents
2.2. Study I: Evaluation of the Al3+ Tolerance of Camelina Genotypes
2.3. Study II: Effect of Melatonin or Nano-Se on Alleviation of the Al3+-Induced Stress in Camelina
2.3.1. Petri Dishes Study
2.3.2. Whole-Plant Study
2.4. Statistical Analysis
3. Results
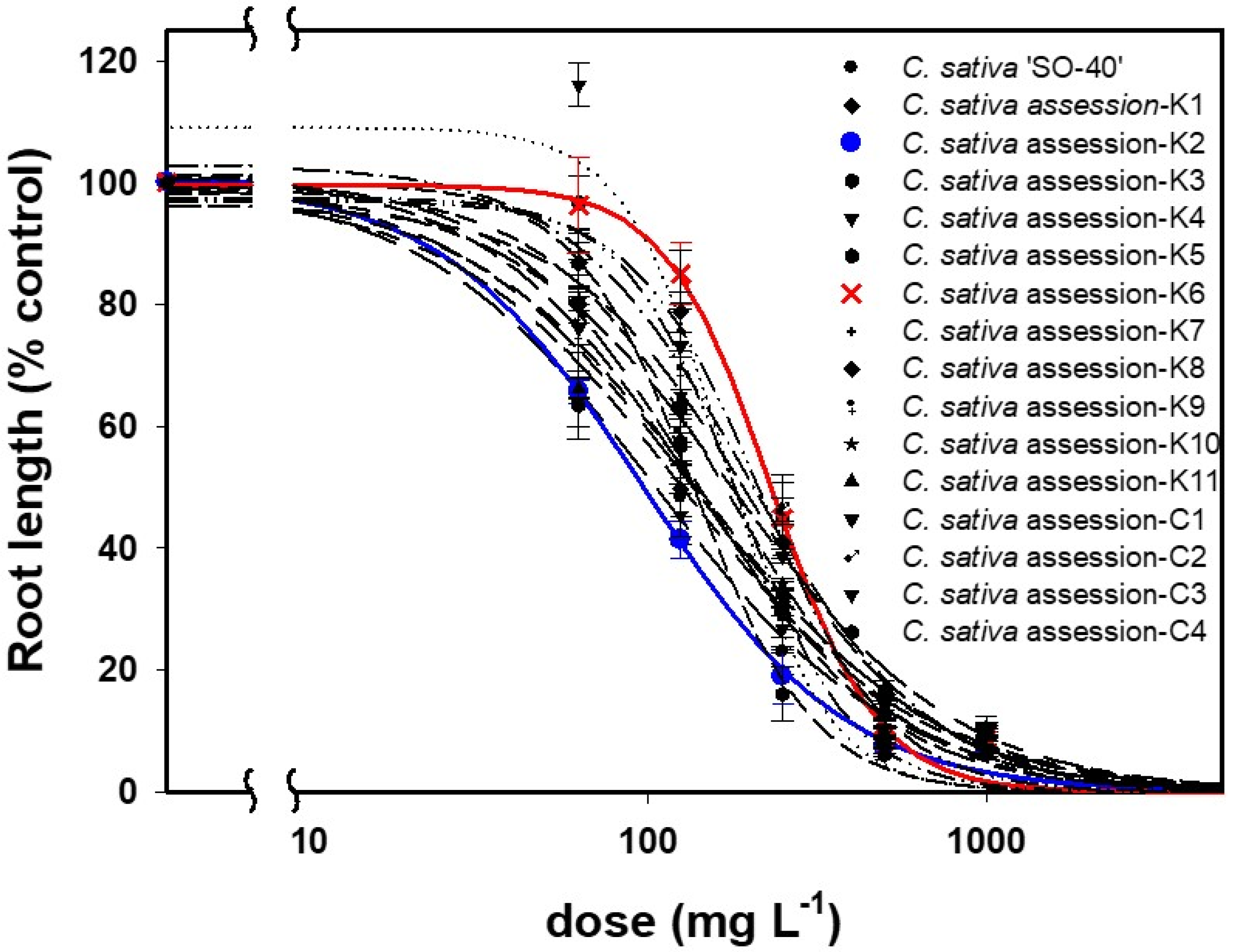
3.1. Dose–Responses and Al3+ Tolerance of Tested Camelina Genotypes
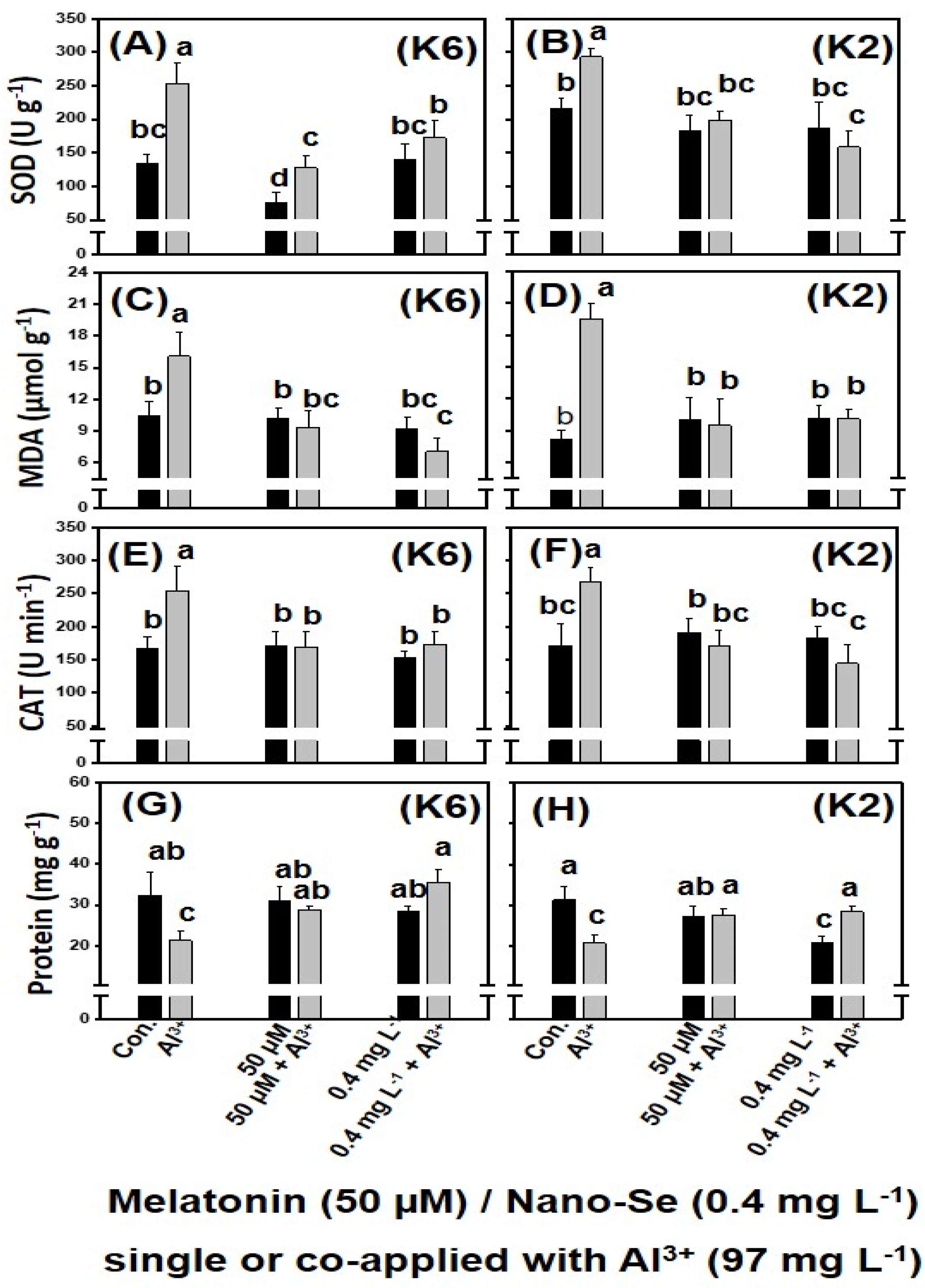
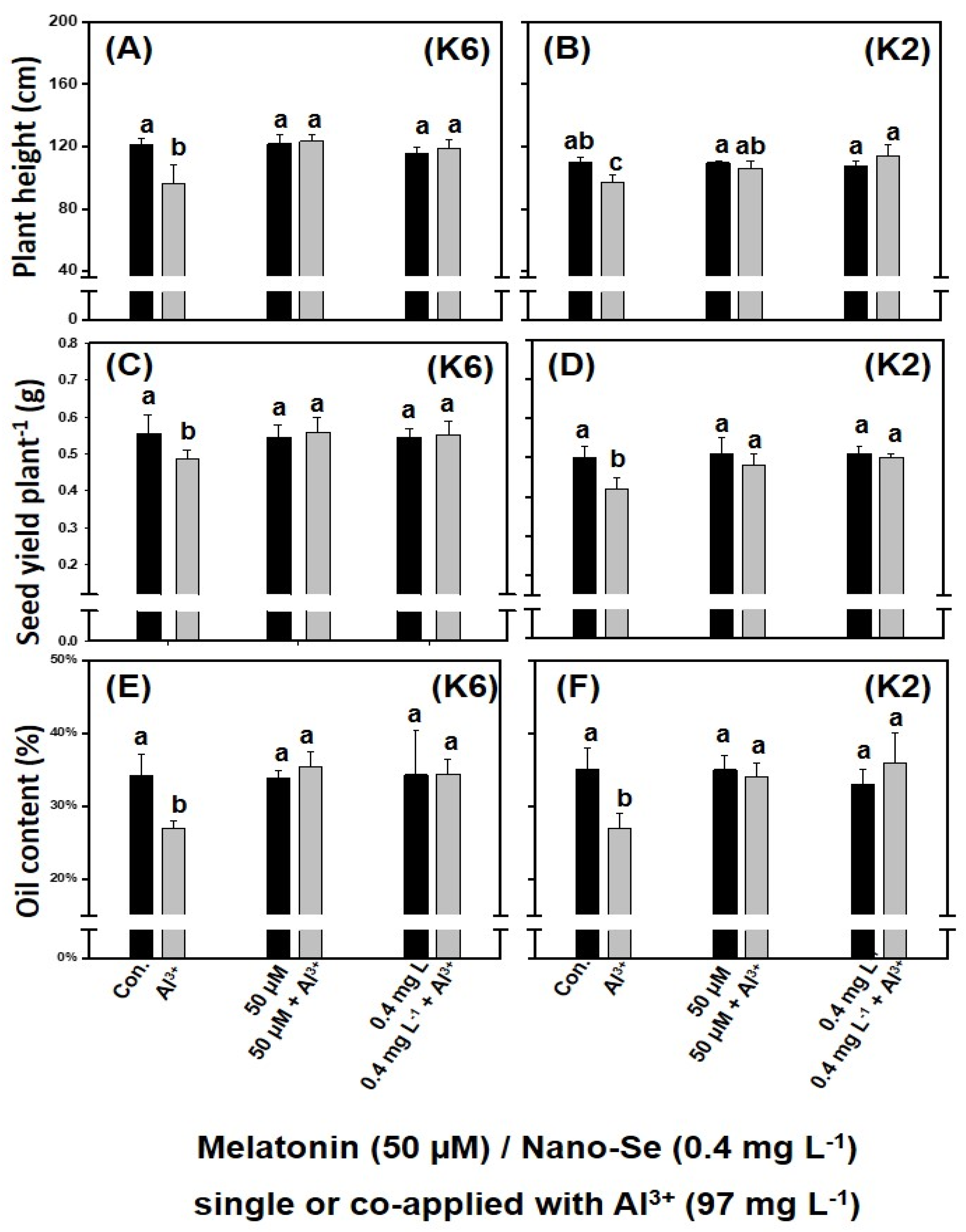
3.2. Alleviation of Al3+-Induced Stress Using Melatonin and Nano-Se
3.2.1. Petri Dish Study
3.2.2. Whole-Plant Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rahman, R.; Upadhyaya, H. Aluminium toxicity its tolerance in plant: A review. J. Plant Biol. 2021, 64, 101–121. [Google Scholar] [CrossRef]
- Shetty, R.; Vidya, C.S.; Nagabovanalli, P.; Lux, A.; Vaculík, M. Aluminum toxicity in plants and its possible mitigation in acid soils by biochar: A review. Sci. Total Environ. 2021, 765, 142744. [Google Scholar] [CrossRef]
- Zeng, H.; Li, Y.R.; Chen, W.J.; Yan, J.W.; Wu, J.S.; Lou, H.Q. Melatonin alleviates aluminum toxicity by regulating aluminum-responsive nonresponsive pathways in hickory. J. Hazard. Mater. 2023, 460, 132274. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.J. Crop production on acidic soils: Overcoming aluminum toxicity and phosphorus deficiency. Ann. Bot. 2010, 106, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V.; Hoekenga, O.A.; Pineros, M.A. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef]
- Kochian, L.V.; Pineros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crop Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Li, X.; Mupondwa, E. Life cycle assessment of camelina oil derived biodiesel and jet fuel in the Canadian Prairies. Sci. Total Environ. 2014, 481, 17–26. [Google Scholar] [CrossRef]
- Zhang, C.J.; Gao, Y.; Jiang, C.; Liu, L.; Wang, Y.; Kim, D.S.; Yu, J.; Yu, L.; Li, F.; Fan, Y. Camelina seed yield and quality in different growing environments in northern China. Ind. Crop Prod. 2021, 172, 114071. [Google Scholar] [CrossRef]
- Aziza, A.E.; Quezada, N.; Cherian, G. Antioxidative effect of dietary camelina meal in fresh, stored, or cooked broiler chicken meat. Poultry Sci. 2010, 89, 2711–2718. [Google Scholar] [CrossRef]
- Betancor, M.B.; Sprague, M.; Usher, S.; Sayanova, O.; Campbell, P.J.; Napier, J.A.; Tocher, D.R. A nutritionally-enhanced oil from transgenic Camelina sativa effectively replaces fish oil as a source of eicosapentaenoic acid for fish. Sci. Rep. 2015, 5, 8104. [Google Scholar] [CrossRef]
- Sindelar, A.; Schmer, M.; Gesch, R.W.; Forcella, F.; Eberle, C.A.; Thom, M.; Archer, D.W. Winter oilseed production for biofuel in the, U.S. Corn Belt: Opportunities and limitations. GCB Bioenergy 2017, 9, 508–524. [Google Scholar] [CrossRef]
- George, N.; Levers, L.; Thompson, S.; Hollingsworth, J.; Kafika, S. Modeling identifies optimal fall planting times and irrigation requirements for canola and camelina at locations across California. Calif. Agric. 2017, 71, 214–220. [Google Scholar] [CrossRef]
- Schillinger, W.F. Camelina: Long-term cropping systems research in a dry Mediterranean climate. Field Crops Res. 2019, 235, 87–94. [Google Scholar] [CrossRef]
- Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Ali, D.; Nadaliet, B. Genetic analysis of freezing tolerance in camelina [Camelina sativa (L.) Crantz] by diallel cross of winter and spring biotypes. Planta 2021, 253, 9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Auer, C. Over wintering assessment of camelina (Camelina sativa) cultivars and congeneric species in the northeastern US. Ind. Crops Prod. 2019, 139, 111532. [Google Scholar] [CrossRef]
- Séguin-Swartz, G.; Eynck, C.; Gugel, R.K.; Strelkov, S.E.; Olivier, C.Y.; Li, J.L.; Falk, K.C. Diseases of Camelina sativa (false flax). Can. J. Plant Pathol. 2009, 31, 375–386. [Google Scholar] [CrossRef]
- Soroka, J.; Olivier, C.; Grenkow, L.; Sguin-Swartz, G. Interactions between Camelina sativa (Brassicaceae) and insect pests of canola. Can. Entomol. 2015, 147, 193–214. [Google Scholar] [CrossRef]
- Stasnik, P.; Großkinsky, D.K.; Jonak, C. Physiological and phenotypic characterization of diverse Camelina sativa lines in response to waterlogging. Plant Physiol. Biochem. 2022, 183, 120–127. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Tanveer, A.; Anwar-U-Hag, M. Foliar applied thiourea improved physiological traits yield of camelina canola under normal heat stress conditions, J. Soil Sci. Plant Nutr. 2021, 21, 1666–1678. [Google Scholar] [CrossRef]
- Wu, H.Z.; Gao, Y.; Zhang, Y.X.; Yu, J.L.; Kim, D.S.; Chen, M.; Wang, Y.W.; Fan, Y.; Zhang, H.X.; Yan, X.B.; et al. Exogenous application of multi-walled carbon nanotubes (MWCNTs) and nano-Selenium (Nano-Se) alleviated the PEG-induced water deficit stress and improved the crop performance of camelina. Agronomy 2023, 13, 979. [Google Scholar] [CrossRef]
- Zuo, Y.T.; Zeng, W.Z.; Ao, C.; Chen, H.R.; Huang, J.S. Effects of multiwalled carbon nanotube and Bacillus atrophaeus application on crop root zone thermal characteristics of saline farmland. Heliyon 2023, 9, e13510. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.W.; Feng, N.J.; Zheng, D.F.; Zhou, H.; Liu, L.; Chen, G.J.; Mu, B.M. Physiological mechanism of exogenous brassinolide alleviating salt stress injury in rice seedlings. Sci. Rep. 2022, 12, 20439. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.R.; Li, D.; Shi, X.L.; Zhang, J.B.; An, O.S.; Wu, Y.L.; Kang, L.; Li, J.O.; Pan, C.P. Nanoselenium enhanced wheat resistance to aphids by regulating biosynthesis of, D.M.B.O.A.; volatile components. J. Agric. Food Chem. 2021, 69, 14103–14114. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Zhan, H.S.; Nie, X.J.; Zhang, T.; Li, S.; Wang, X.Y.; Du, X.H.; Tong, W.; Song, W.N. Melatonin: A small molecule but important for salt stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 709. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, X.; Ma, C.; Wang, Y.; Zhao, J. Melatonin enhances drought stress tolerance in maize through coordinated regulation of carbon and nitrogen assimilation. Plant Physiol. Biochem. 2021, 167, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The role of melatonin in salt stress responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef] [PubMed]
- Sami, A.; Shah, F.A.; Abdullah, M.; Yu, Z.X.; Jin, Z.K. Melatonin mitigates cadmium and aluminium toxicity through modulation of antioxidant potential in Brassica napus L. Plant Biol. 2020, 4, 679–690. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Wang, L.; Li, B.; Jin, C.; Lin, X. Melatonin: A master regulator of plant development and stress responses. J. Integr. Plant Biol. 2020, 63, 126–145. [Google Scholar] [CrossRef]
- Bukhari, M.A.; Yousaf, M.; Ahmad, Z.; Rafay, M.; Shah, A.N.; Abbas, A.; Shah, A.A.; Javed, T.; Afzal, M.; Ali, S.; et al. Enhancing drought stress tolerance in camelina (Camelina sativa L.) through exogenous application of potassium. Physiol. Plantarum. 2022, 174, e13779. [Google Scholar] [CrossRef]
- Naderi, R.; Afranjeh, E.; Heidari, B.; Emam, Y.; Egan, T.P. Salicylic acid and superabsorbent polymers could alleviate water deficit stress in camelina (Camelina Sativa L.). Commun. Soil Sci. Plant Anal. 2023, 54, 2863–2873. [Google Scholar] [CrossRef]
- Aghdasi, S.; AghaAlikhani, M.; Modarres-Sanavy, S.A.M.; Kahrizi, D. Exogenously used boron and 24-epibrassinolide improved oil quality and mitigate late-season water deficit stress in camelina. Ind. Crop. Prod. 2021, 171, 113885. [Google Scholar] [CrossRef]
- Bakyani, M.R.F.; Alinia, M.; Kazemeini, S.A.; Abadía, J.; Dadkhodaie, A. Foliar application of melatonin improves the salt tolerance on and redox homeostasis and seed oil fatty acid profile in Camelina sativa. Plants 2022, 11, 3113. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, J.; Gao, Y.; Li, Z.W.; Kim, D.-S.; Chen, M.; Fan, Y.; Zhang, H.; Yan, X.; Zhang, C.-J. Agronomic evaluation of shade tolerance of 16 spring Camelina sativa (L.) Crantz genotypes under different artificial shade levels using a modified membership function. Front. Plant Sci. 2022, 13, 978932. [Google Scholar] [CrossRef] [PubMed]
- Makeen, K.; Babu, G.S.; Lavanya, G.R.; Abraham, G. Studies of chlorophyll content by different methods in Black Gram (Vigna mungo L.). Int. J. Agric. Res. 2007, 2, 651–655. [Google Scholar]
- Martinelli, T.; Galasso, I. Phenological growth stages of Camelina sativa according to the extended BBCH scale. Ann. Appl. Biol. 2011, 158, 87–94. [Google Scholar] [CrossRef]
- Zou, Q. Plant physiology experimental guidance; Agriculture Press Publishing: Beijing, China, 2000. [Google Scholar]
- Knezevic, S.Z.; Streibig, J.C.; Ritz, C. Utilizing R software package for dose-response studies: The concept and data analysis. Weed Technol. 2007, 21, 840–848. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 3 January 2024).
- Dai, C.; Oiu, L.; Guo, L.; Jing, S.; Chen, X.; Cui, X.; Yang, Y. Salicylic acid alleviates aluminum-induced inhibition of biomass by enhancing photosynthesis and carbohydrate metabolism in Panax notoginseng. Plant Soil 2019, 445, 183–198. [Google Scholar] [CrossRef]
- Wang, L.; Fan, X.W.; Pan, J.L.; Huang, Z.B.; Li, Y.Z. Physiological characterization of maize tolerance to low dose of aluminum, highlighted by promoted by promoted leaf growth. Planta 2015, 242, 1391–1403. [Google Scholar] [CrossRef]
- Yan, L.; Riaz, M.; Liu, J.; Yu, M.; Cuncang, J. The aluminum tolerance and detoxification mechanisms in plants; recent advances and prospects. Crit Rev. Env. Sci. Tec. 2022, 52, 1491–1527. [Google Scholar] [CrossRef]
- Gao, T.; Liu, X.; Tan, K.; Zhang, D.; Zhu, B.; Ma, F.W.; Li, C. Introducing melatonin to the horticultural industry: Physiological roles, potential applications, and challenges. Hortic. Res. 2022, 22, uhac094. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Wu, Y.L.; Zhang, J.B.; An, Q.S.; Zhou, C.R.; Li, D.; Pan, C.P. Nano-selenium enhances the antioxidant capacity, organic acids and cucurbitacin B in melon (Cucumis melo L.) plants. Ecotoxicol. Environ. Saf. 2022, 241, 113777. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Pandey, P.; Upadhyay, T.K. Applications of nanotechnology-based agrochemicals in food security and sustainable agriculture: An overview. Agriculture 2022, 12, 1672. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants-focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene. 2019, 19, 100182. [Google Scholar] [CrossRef]
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semidaet, W.M. Selenium application in two methods promotes drought tolerance in Solanum lycopersicum plant by inducing the antioxidant defense system. Sci. Hortic. 2020, 266, 109290. [Google Scholar] [CrossRef]
- Efeoglu, B.; Ekmekci, Y.; Cicek, N. Physiological responses of three maize cultivars to drought stress recovery. S. Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef]
- Ahmed, Z.; Waraich, E.; Ahmad, R.; Shahbaz, M. Morpho-physiological and biochemical responses of camelina (Camelina sativa crantz) genotypes under drought stress. Int. J. Agric. Biol. 2017, 19, 1–7. [Google Scholar] [CrossRef]
- Nayyar, H.; Walia, D.P. Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Biol. Plantarum. 2003, 46, 275–279. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhu, X.F.; Peng, Y.X.; Zheng, C.; Li, G.X.; Liu, Y.; Shi, Y.Z.; Zheng, S.J. Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis. Plant Physiol. 2011, 155, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.O.; Fan, W.; Jin, J.F.; Xu, J.M.; Chen, W.W.; Yang, J.L.; Zheng, S.J. A NAC-type transcription factor confers aluminium resistance by regulating cell wall-associated receptor kinase 1 and cell wall pectin. Plant Cell Environ. 2020, 43, 463–478. [Google Scholar] [CrossRef] [PubMed]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohyd. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Borzoo, S.; Mohsenzadeh, S.; Kahrizi, D. Water-deficit stress and genotype variation induced alteration in seed characteristics of Camelina sativa. Rhizosphere 2021, 20, 100427. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Petri Dish Study | Whole-Plant Level Study |
|---|---|---|
| Various Solutions Treatment | ||
| 1 | Untreated control (only distilled water) | Untreated control (only distilled water) |
| 2 | 97 mg L−1 Al3+ | 97 mg L−1 Al3+ |
| 3 | 50 μM melatonin | 50 μM melatonin |
| 4 | 100 μM melatonin | 0.4 mg L−1 nano-Se |
| 5 | 200 μM melatonin | 97 mg L−1 Al3+ + 50 μM melatonin |
| 6 | 50 μM melatonin + 97 mg L−1 Al3+ | 97 mg L−1 Al3+ + 0.4 mg L−1 nano-Se |
| 7 | 100 μM melatonin + 97 mg L−1 Al3+ | - |
| 8 | 200 μM melatonin + 97 mg L−1 Al3+ | - |
| 9 | 0.4 mg L−1 nano-Se | - |
| 10 | 2 mg L−1 nano-Se | - |
| 11 | 10 mg L−1 nano-Se | - |
| 12 | 0.4 mg L−1 nano-Se + 97 mg L−1 Al3+ | - |
| 13 | 2 mg L−1 nano-Se + 97 mg L−1 Al3+ | - |
| 14 | 1 mg L−1 nano-Se + 97 mg L−1 Al3+ | - |
| Variables | DF | Percent Germination | Root Length | Shoot Length | Seedling Length | Chlorophyll Content | Fresh Weight | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | MS | F | p | ||
| Concentration (C) | 5 | 8206 | 131 | *** | 68,770 | 594 | *** | 73,238 | 351 | *** | 64,221 | 728 | *** | 80,059 | 170 | *** | 31,305 | 67 | *** |
| Genotype (G) | 15 | 365 | 5.9 | *** | 438 | 3.7 | *** | 877 | 4.2 | *** | 318 | 3.6 | *** | 2682 | 5.7 | *** | 2227 | 4.7 | *** |
| C × G | 75 | 233 | 3.7 | *** | 189 | 1.6 | ** | 702 | 3.4 | *** | 167 | 1.9 | *** | 729 | 1.6 | ** | 490 | 1.0 | NS a |
| Residual | 192 | 12,051 | 63 | - | 22,215 | 116 | - | 40,105 | 209 | - | 16,941 | 88 | - | 90,594 | 472 | - | 90,182 | 470 | - |
| Accessions No. | Root Length | Shoot Length | Seedling Length | Fresh Weight | Chlorophyll Content | Percent Germination | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B | D | GR50 | B | D | GR50 | B | D | GR50 | B | D | GR50 | B | D | GR50 | B | D | GR50 | |
| ‘SO-40’ | 1.3 (0.2) b | 99.8 (6.2) | 106.0 (18.3) | 4.47 (3.2) | 93.8 (5.4) | 549.2 (52.3) | 1.19 (0.2) | 99.5 (5.4) | 147.7 (23.0) | 1.35 (0.7) | 97.1 (11.0) | 643.5 (204.5) | 2.89 (1.2) | 94.8 (8.8) | 404.0 (82.9) | 2.88 (0.5) | 104.5 (2.5) | 848.0 (50.2) |
| CamK1 | 1.4 (0.2) | 101.2 (6.0) | 136.0 (19.8) | 28.20 (191.8) | 104.7 (4.5) | 517.0 (117.4) | 1.28 (0.2) | 100.5 (5.3) | 196.2 (28.2) | 0.81 (0.7) | 96.6 (14.8) | 1147.1 (711.0) | 4.60 (1.7) | 124.6 (6.9) | 636.3 (77.9) | 4.25 (2.4) | 96.9 (2.4) | 1091.1 (73.8) |
| CamK2 | 1.5 (0.3) | 100.1 (6.2) | 97.0 (14.7) | 19.47 (112.6) | 123.2 (4.5) | 463.4 (203.8) | 1.46 (0.2) | 98.6 (5.5) | 144.8 (21.7) | 2.35 (1.5) | 87.3 (8.2) | 686.7 (152.0) | 3.53 (1.2) | 146.6 (7.7) | 481.3 (50.7) | 1.23 (0.4) | 101.0 (3.6) | 1483.7 (311.1) |
| CamK3 | 2.5 (0.5) | 99.8 (5.9) | 137.0 (14.1) | 3.48 (1.3) | 105.3 (5.0) | 490.1 (49.7) | 1.82 (0.3) | 100.9 (5.1) | 177.9 (19.3) | 1.16 (0.8) | 97.0 (11.8) | 879.3 (343.9) | 2.86 (0.9) | 117.4 (8.2) | 412.3 (65.2) | 1.64 (0.5) | 94.5 (3.9) | 872.7 (90.0) |
| CamK4 | 2.94 (0.5) | 109.0 (4.94) | 169.8 (13.9) | 29.95 (327.9) | 114.0 (4.5) | 463.6 (384.1) | 2.60 (0.4) | 107.9 (4.3) | 208.9 (16.2) | 1.89 (0.5) | 131.2 (7.8) | 636.2 (112.8) | 6.17 (2.5) | 117.0 (7.7) | 347.6 (53.2) | 3.31 (1.4) | 97.5 (2.5) | 1126.0 (91.4) |
| CamK5 | 1.64 (0.3) | 102.8 (5.6) | 198.9 (25.8) | 16.13 (98.5) | 118.8 (4.5) | 527.3 (171.0) | 1.7 (0.3) | 102.1 (4.8) | 270.9 (30.7) | 2.17 (0.9) | 108.8 (7.2) | 910.2 (184.3) | 4.09 (2.8) | 99.3 (7.7) | 507.3 (71.3) | 3.76 (0.8) | 105.0 (2.3) | 923.4 (44.2) |
| CamK6 | 2.78 (0.7) | 99.6 (5.0) | 232.0 (21.2) | 20.88 (452.1) | 103.1 (4.5) | 896.0 (85.7) | 2.74 (0.6) | 98.8 (4.0) | 286.2 (21.9) | 1.24 (0.4) | 102.6 (11.3) | 1314.3 (101.7) | 4.48 (2.0) | 126.3 (7.7) | 418.1 (48.2) | 4.67 (4.2) | 101.8 (2.2) | 1251.8 (259.4) |
| CamK7 | 2.60 (0.7) | 97.5 (5.7) | 184.3 (18.7) | 19.97 (152.9) | 108.6 (5.2) | 472.7 (181.1) | 2.83 (0.6) | 97.4 (4.5) | 214.2 (17.8) | 1.45 (0.5) | 104.8 (10.5) | 331.0 (89.5) | 4.37 (2.1) | 108.2 (8.3) | 319.5 (45.0) | 1.57 (0.9) | 98.0 (4.4) | 1396 (342.8) |
| CamK8 | 2.33 (0.6) | 97.0 (5.6) | 218.0 (24.3) | 16.43 (233.8) | 112.5 (4.5) | 498.7 (21.6) | 2.42 (0.5) | 97.4 (4.3) | 273.3 (24.5) | 2.51 (0.9) | 110.2 (7.2) | 595.8 (99.5) | 2.87 (1.2) | 104.8 (9.0) | 386.6 (71.2) | 3.98 (1.0) | 99.9 (2.4) | 908 (44.8) |
| CamK9 | 1.62 (0.4) | 96.1 (6.5) | 201.4 (32.8) | 23.85 (897.4) | 103.3 (4.5) | 492.5 (278.5) | 2.15 (0.6) | 91.7 (5.6) | 283.6 (35.9) | 1.45 (0.9) | 95.6 (11.1) | 641.4 (195.3) | 3.76 (1.9) | 104.1 (8.0) | 428.7 (65.0) | 2.01 (0.6) | 98.6 (2.9) | 1213.5 (152.7) |
| CamK10 | 1.66 (0.3) | 99.1 (6.1) | 170.6 (23.8) | 1.69 (0.6) | 101.7 (6.0) | 862.8 (163.6) | 1.40 (0.2) | 100.1 (5.2) | 214.7 (28.8) | 0.82 (0.3) | 102.0 (11.6) | 509.3 (239.8) | 2.10 (0.9) | 96.3 (8.3) | 589.6 (138.9) | 0.54 (0.3) | 100.3 (4.2) | 20,661 (35,101) |
| CamK11 | 1.30 (0.2) | 98.3 (6.3) | 144.7 (25.4) | 2.70 (0.7) | 109.2 (4.9) | 801.0 (93.8) | 1.20 (0.2) | 98.2 (5.5) | 211.6 (33.9) | 0.10 (0.3) | 105.7 (11.3) | 311.8 (114.9) | 2.10 (0.9) | 106.2 (7.7) | 707.9 (120.1) | 0.59 (0.4) | 99.7 (4.4) | 18,447 (36,418) |
| CamC1 | 1.56 (0.3) | 100.7 (6.0) | 140.8 (19.5) | 2.06 (0.5) | 104.3 (5.9) | 488.9 (71.3) | 1.42 (0.2) | 100.4 (5.2) | 182.6 (24.0) | 0.62 (0.3) | 102.0 (12.0) | 316.5 (180.5) | 2.41 (1.1) | 101.6 (9.4) | 335.0 (67.4) | 1.15 (0.7) | 99.7 (4.0) | 2795 (1963) |
| CamC2 | 1.33 (0.2) | 98.9 (6.2) | 180.2 (30.2) | 3.09 (0.7) | 109.4 (4.8) | 664.5 (72.3) | 1.35 (0.2) | 97.6 (5.5) | 265.4 (40.1) | 0.75 (0.3) | 103.4 (11.5) | 465.7 (234.0) | 2.19 (0.8) | 111.8 (8.1) | 564.4 (112.2) | 0.90 (1.0) | 100.9 (3.7) | 11,005 (31,896) |
| CamC3 | 1.24 (0.2) | 100.5 (6.1) | 135.1 (22.4) | 2.48 (1.0) | 101.0 (5.0) | 812.2 (149.5) | 1.04 (0.2) | 100.3 (5.4) | 194.4 (32.7) | 1.39 (0.4) | 110.4 (9.9) | 361.8 (95.6) | 2.45 (1.0) | 102.5 (8.1) | 552.5 (108.0) | 1.25 (2.0) | 98.6 (4.0) | 6654 (18,916) |
| CamC4 | 1.26 (0.2) | 98.5 (6.3) | 129.3 (23.2) | 2.15 (0.5) | 106.1 (5.6) | 569.5 (79.2) | 1.25 (0.2) | 97.9 (5.6) | 177.4 (28.4) | 0.58 (0.3) | 101.2 (12.1) | 466.1 (304.4) | 2.17 (1.1) | 97.1 (9.7) | 425.6 (97.7) | 0.63 (0.3) | 100.1 (4.3) | 9663 (11,895) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Z.-C.; Chen, M.; Tang, Y.; Gao, Y.; Wu, H.-Z.; Min, X.; Kim, D.-S.; Yan, X.; Yu, J.; Zhang, C.-J. Evaluation of the Aluminum (Al3+) Tolerance of Camelina and the Potential for Using Melatonin or Nano-Selenium to Alleviate Al3+-Induced Stress in Camelina. Agronomy 2024, 14, 401. https://doi.org/10.3390/agronomy14030401
Lv Z-C, Chen M, Tang Y, Gao Y, Wu H-Z, Min X, Kim D-S, Yan X, Yu J, Zhang C-J. Evaluation of the Aluminum (Al3+) Tolerance of Camelina and the Potential for Using Melatonin or Nano-Selenium to Alleviate Al3+-Induced Stress in Camelina. Agronomy. 2024; 14(3):401. https://doi.org/10.3390/agronomy14030401
Chicago/Turabian StyleLv, Zhao-Chen, Min Chen, Yi Tang, Yang Gao, Hui-Zhen Wu, Xueyang Min, Do-Soon Kim, Xuebing Yan, Jialin Yu, and Chuan-Jie Zhang. 2024. "Evaluation of the Aluminum (Al3+) Tolerance of Camelina and the Potential for Using Melatonin or Nano-Selenium to Alleviate Al3+-Induced Stress in Camelina" Agronomy 14, no. 3: 401. https://doi.org/10.3390/agronomy14030401
APA StyleLv, Z.-C., Chen, M., Tang, Y., Gao, Y., Wu, H.-Z., Min, X., Kim, D.-S., Yan, X., Yu, J., & Zhang, C.-J. (2024). Evaluation of the Aluminum (Al3+) Tolerance of Camelina and the Potential for Using Melatonin or Nano-Selenium to Alleviate Al3+-Induced Stress in Camelina. Agronomy, 14(3), 401. https://doi.org/10.3390/agronomy14030401