


Effects of Cd Stress on Morphological and Physiological Characteristics of Maize Seedlings

Abstract
1. Introduction
2. Materials and Methods
2.1. Maize Material Culture Conditions and Cd Treatment
2.2. Measurement Items and Methods
2.2.1. Growth Indicators
2.2.2. Measurement of Physiological and Biochemical Indicators
2.2.3. Determination of Cd Content in Maize above Ground and below Ground under Different Treatments
2.2.4. Root Scanning
2.3. Statistical Analysis
3. Results
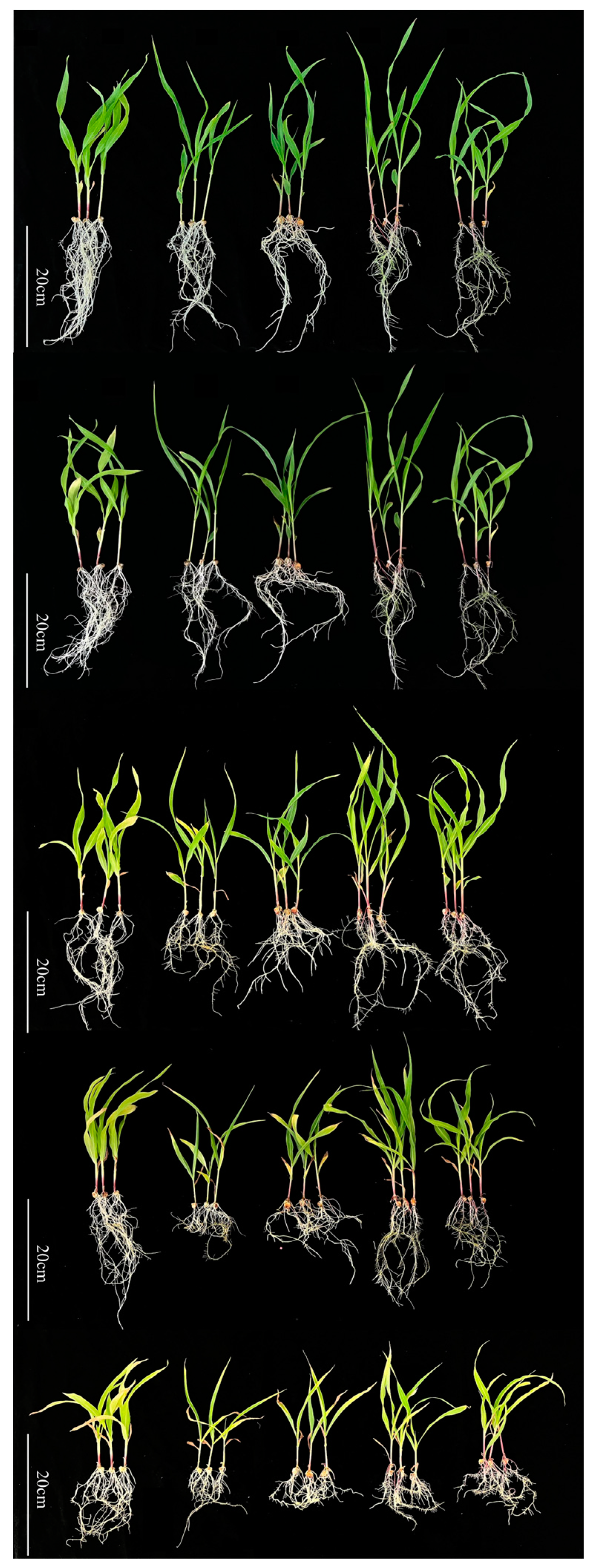
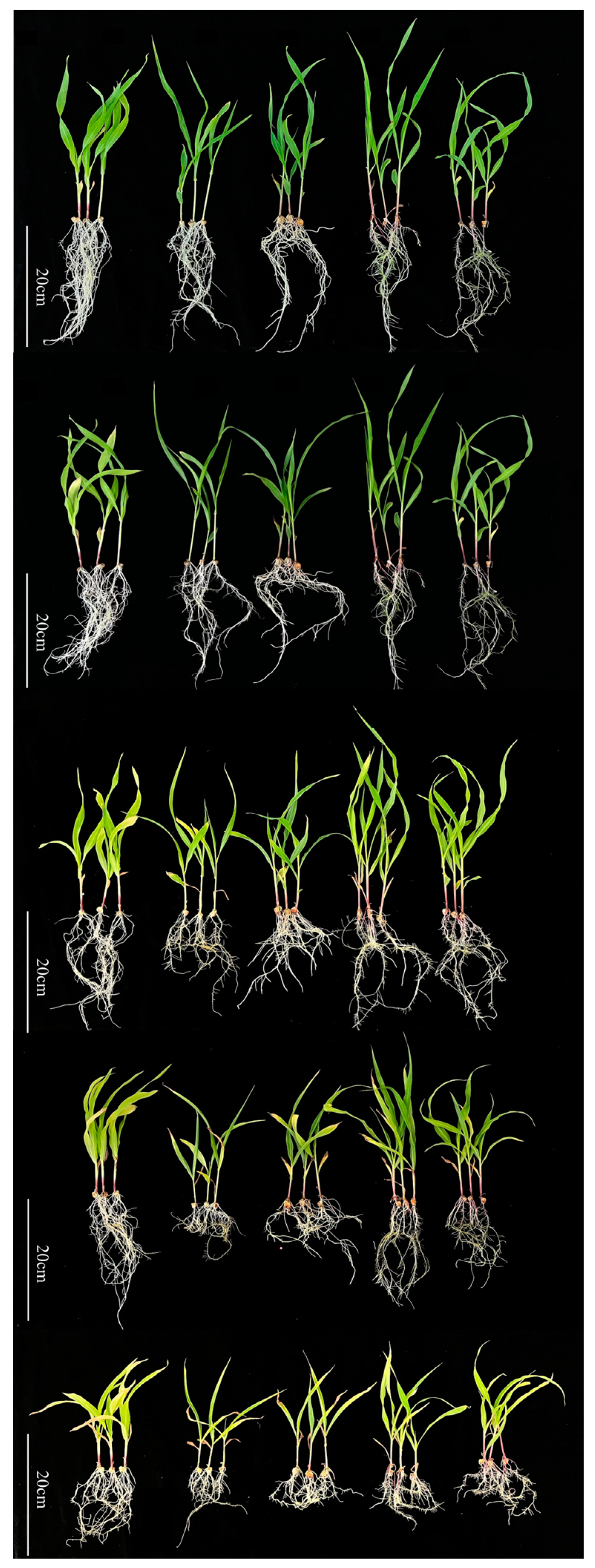
3.1. Impact of Cd Stress on Maize Seedling Growth
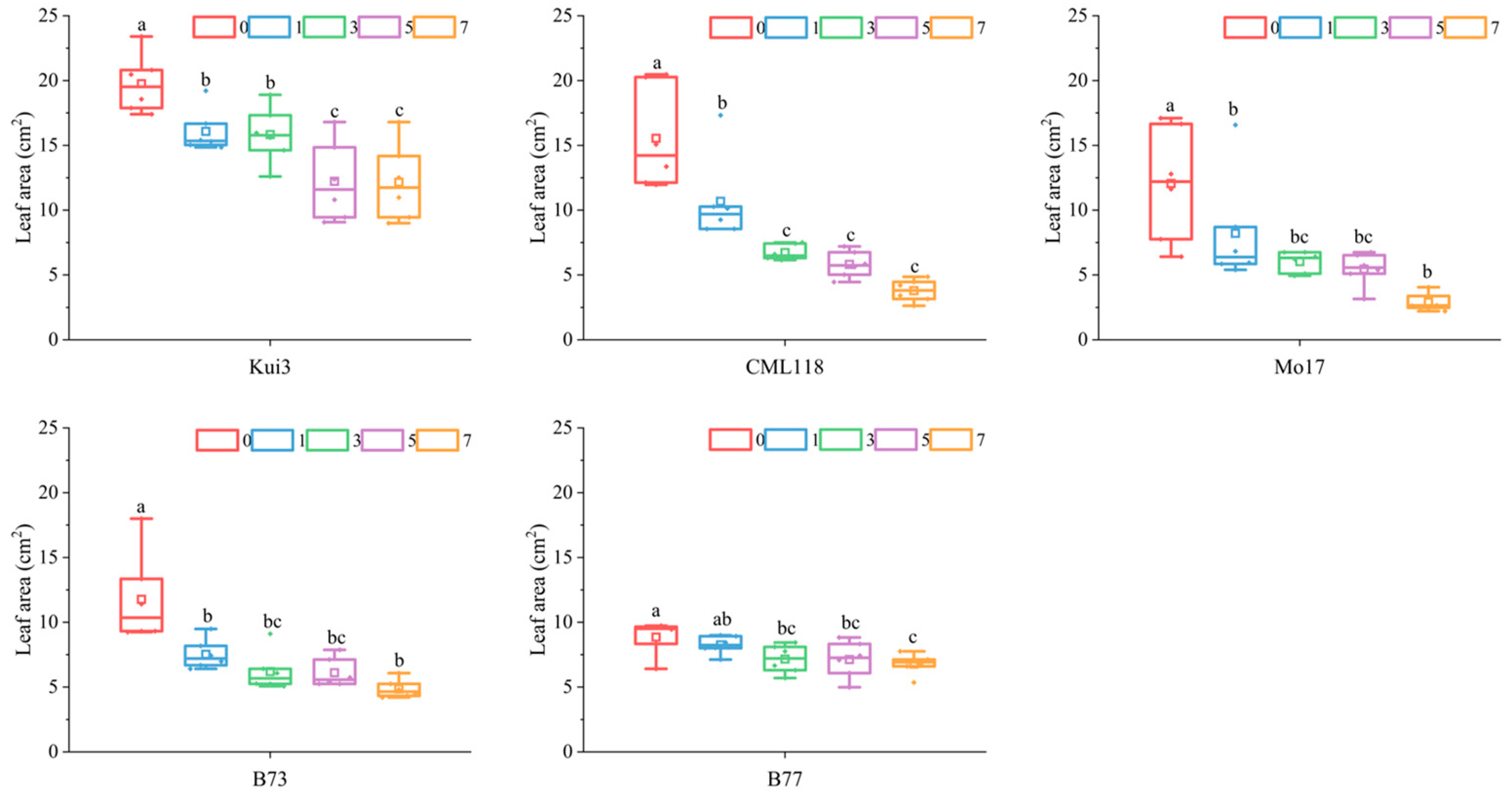
3.2. Impact of Cd Stress on Maize Blades
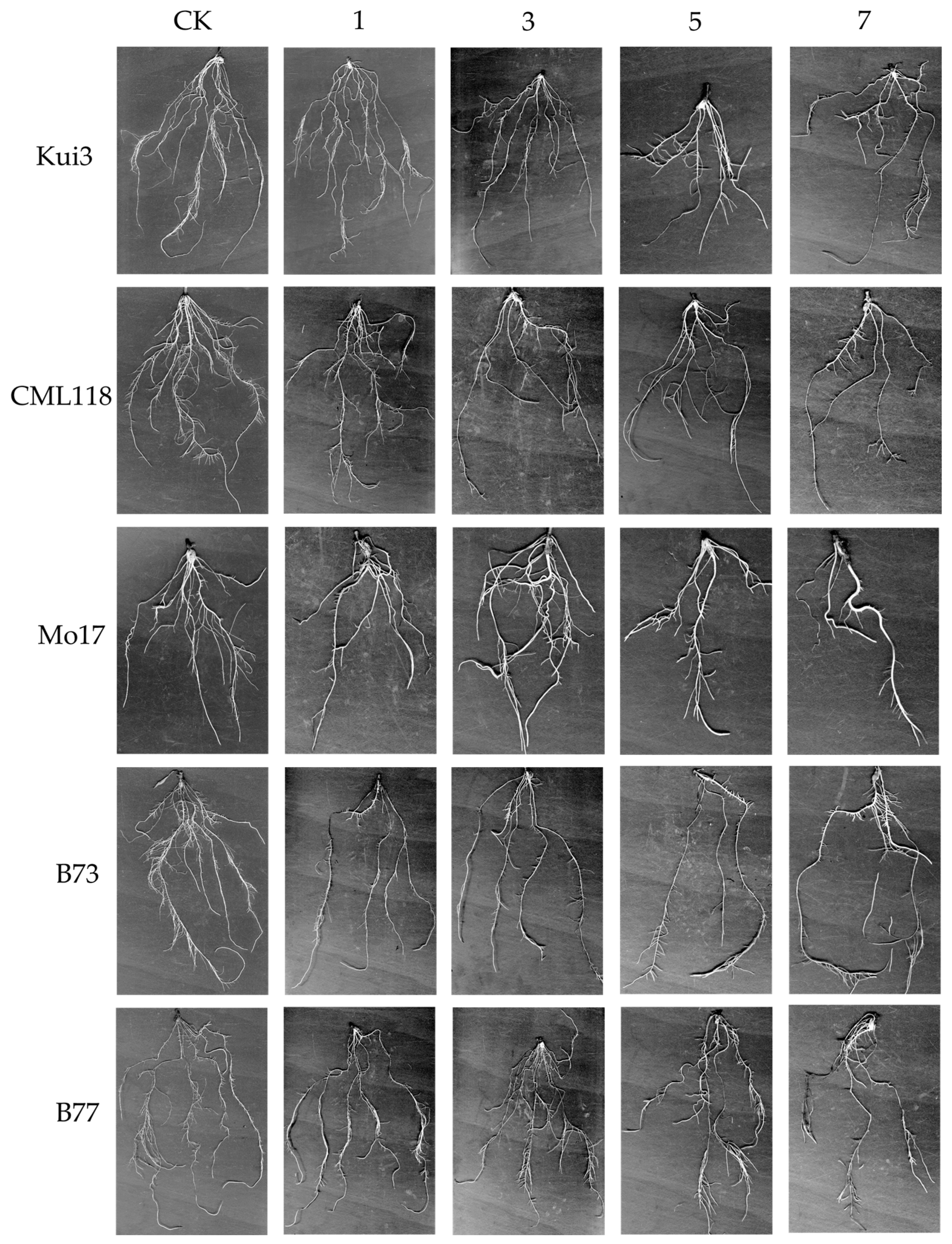
3.3. Effect of Cd on Root Morphological Traits
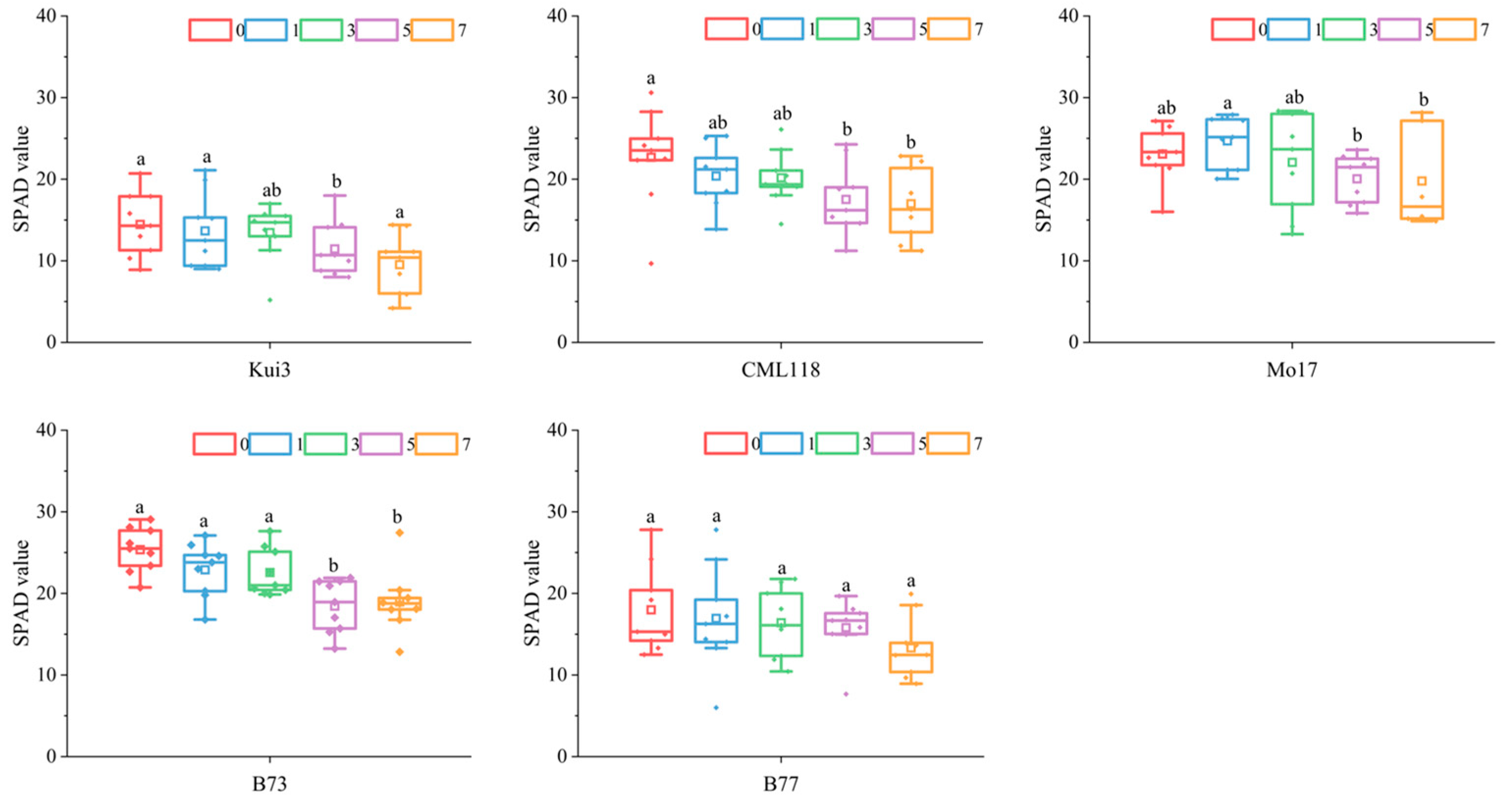
3.4. Effect of Cd Stress on SPAD Value
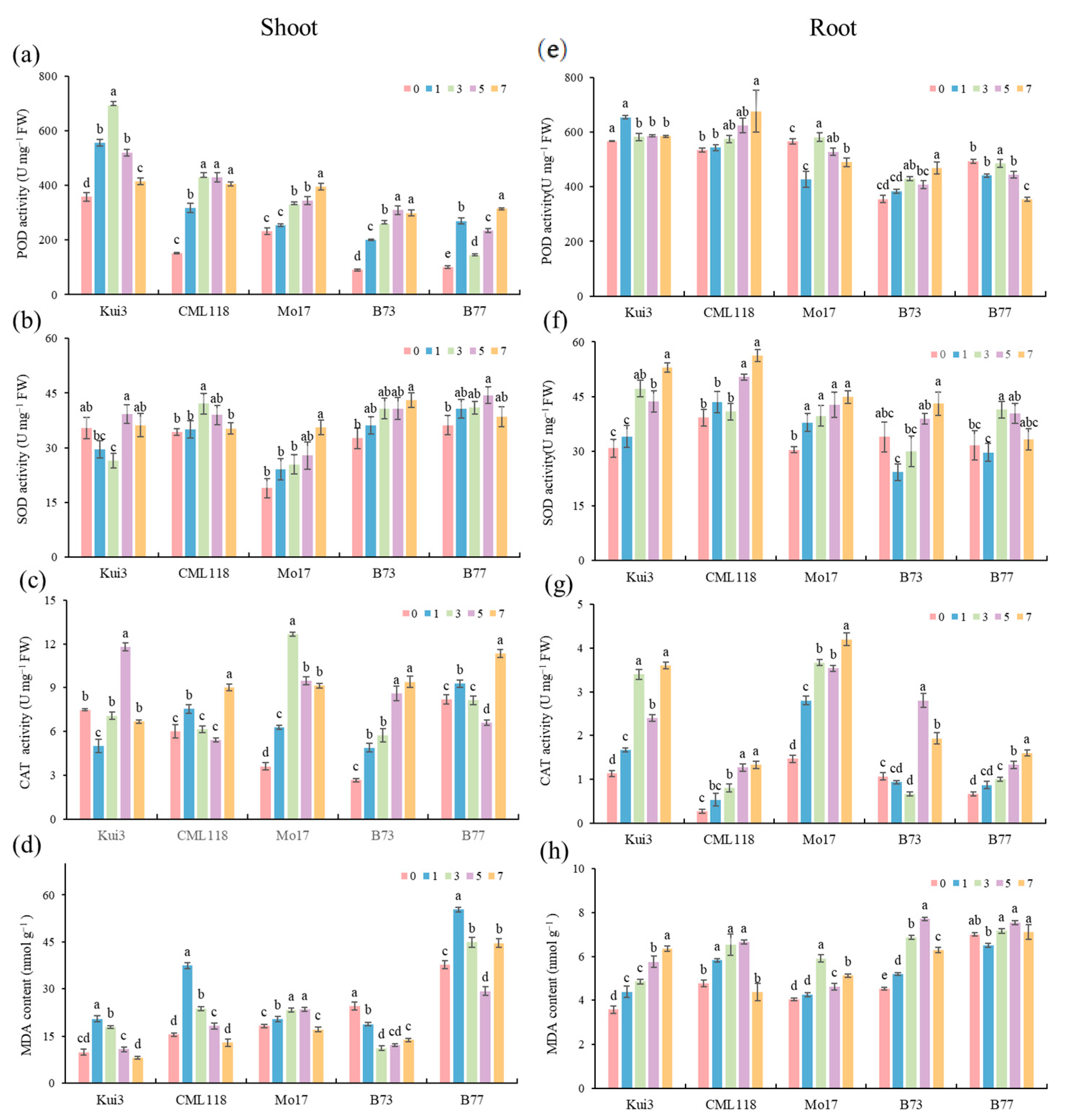
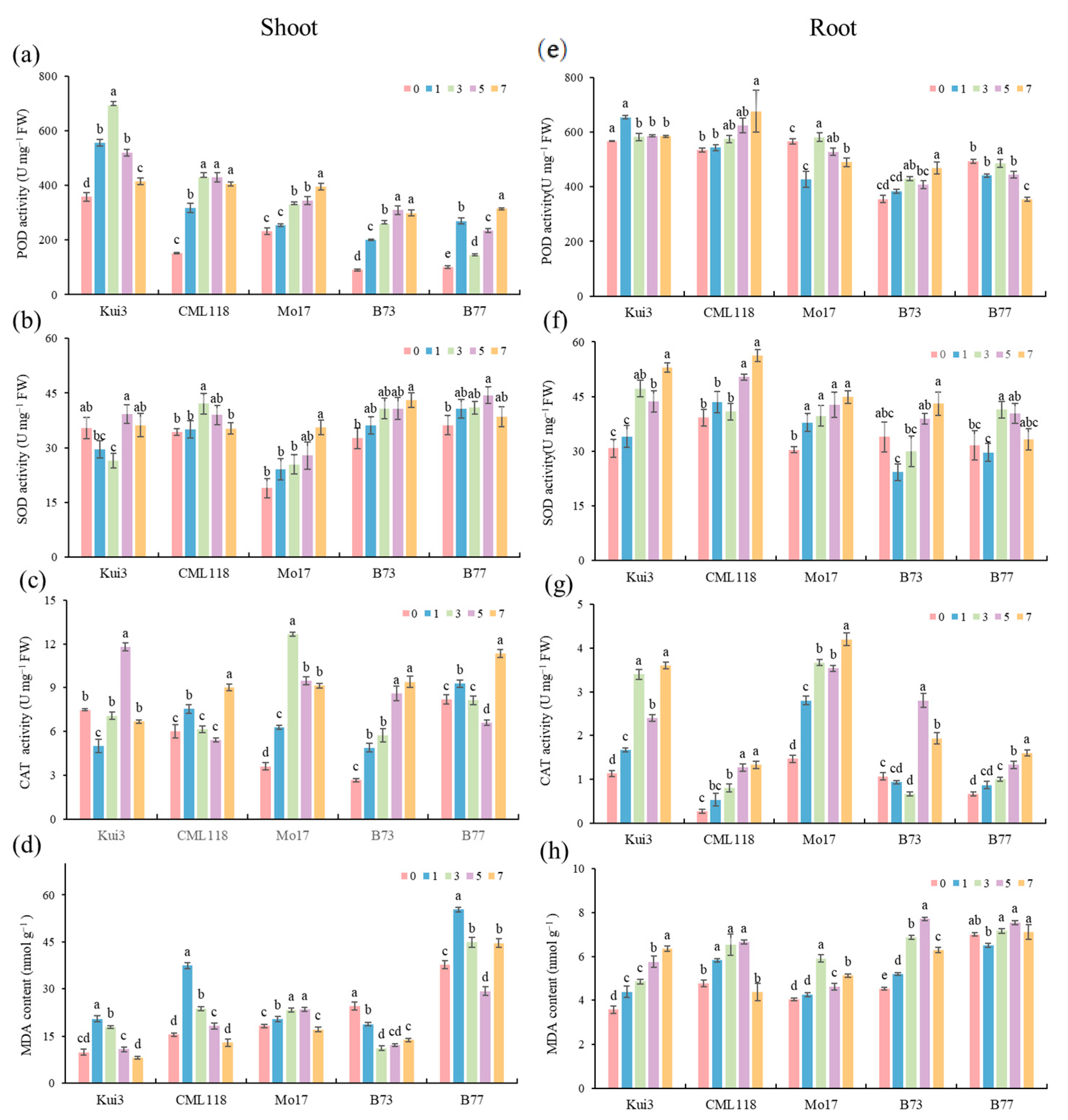
3.5. Effects of the Protective Enzyme System and MDA Content under Cd Stress
3.6. Effects of Osmoregulatory Capacity under Cd Stress
3.7. Cd Uptake and Transport in Different Maize Varieties
4. Discussion
4.1. Changes in Biomass under Cd Treatment
4.2. Cd Affects Relative Leaf Water Content and SPAD Values of Seedlings
4.3. Antioxidant Defense System Stress Response
4.4. Cd Treatment Triggers Oxidative Damage to Lipid Membranes
4.5. Changes in Soluble Protein Content
4.6. Regulation of Soluble Sugars under Cd Stress
4.7. Cd Stress Induced High Accumulation of Proline
4.8. Cd Accumulation and Transport
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. Biotechnol. Adv. 2009, 27, 195–226. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, H.; Tao, S. The Challenges and Solutions for Cadmium-contaminated Rice in China: A Critical Review. Environ Int. 2016, 92, 515–532. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Environmental Health Criteria 134: Cadmium; WHO: Geneva, Switzerland, 1992. [Google Scholar]
- Qin, P.; Wang, L.; Liu, K.; Mao, S.; Li, Z.; Gao, S.; Shi, H.; Liu, Y. Genomewide association study of Aegilops tauschii traits under seedling-stage cadmium stress. Crop J. 2015, 3, 405–415. [Google Scholar] [CrossRef]
- Sandalio, L.; Dalurzo, H.; Gómez, M.; Romero-Puertas, M.; del Río, L. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef]
- Liu, H.; Hussain, S.; Peng, S.; Huang, J.; Cui, K.; Nie, L. Potentially toxic elements concentration in milled rice differ among various planting patterns. Field Crops Res. 2014, 168, 19–26. [Google Scholar] [CrossRef]
- Wang, X.K.; Gong, X.; Cao, F.; Wang, Y.; Zhang, G.; Wu, F. HvPAA1 encodes a P-Type ATPase, a novel gene for cadmium accumulation and tolerance in barley (Hordeum vulgare L.). Int. J. Mol. Sci. 2019, 20, 1732. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Peng, G.; Huang, L. Trophic transfer, bioaccumulation, and potential health risk of trace elements in water and aquatic organisms of Yundang Lagoon at Xiamen in China. Toxin Rev. 2023, 42, 242–256. [Google Scholar] [CrossRef]
- Johri, N.; Jacquillet, G.; Unwin, R. Heavy metal poisoning: The effects of cadmium on the kidney. Biometals 2010, 23, 783–792. [Google Scholar] [CrossRef]
- Godt, J.; Scheidig, F.; Grosse-Siestrup, C.; Esche, V.; Brandenburg, P.; Reich, A.; Groneberg, D.A. The toxicity of cadmium and resulting hazards for human health. J. Occup. Med. Toxicol. 2006, 1, 22. [Google Scholar] [CrossRef]
- Mezynska, M.; Brzóska, M.M. Environmental exposure to cadmium—A risk for health of the general population in industrialized countries and preventive strategies. Environ. Sci. Pollut. Res. 2018, 25, 3211–3232. [Google Scholar] [CrossRef]
- Sarma, S.N.; Saleem, A.; Lee, J.-Y.; Tokumoto, M.; Hwang, G.-W.; Chan, H.M.; Satoh, M. Effects of long-term cadmium exposure on urinary metabolite profiles in mice. J. Toxicol. Sci. 2018, 43, 89–100. [Google Scholar] [CrossRef]
- Shah, K.; Kumar, R.G.; Verma, S.; Dubey, R. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci. 2001, 161, 1135–1144. [Google Scholar] [CrossRef]
- Tai, Z.; Yin, X.; Fang, Z.; Shi, G.; Lou, L.; Cai, Q. Exogenous GR24 alleviates cadmium toxicity by reducing cadmium uptake in switchgrass (Panicum virgatum) seedlings. Int. J. Environ. Res. Public Health 2017, 14, 852. [Google Scholar] [CrossRef] [PubMed]
- Zoufan, P.; Jalali, R.; Hassibi, P.; Neisi, E.; Rastegarzadeh, S. Evaluation of antioxidant bioindicators and growth responses in Malva parviflora L. exposed to cadmium. Physiol. Mol. Biol. Plants 2018, 24, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Amri, B.; Khamassi, K.; Ali, M.B.; da Silva, J.A.T.; Ben Kaab, L.B. Effects of gibberellic acid on the process of organic reserve mobilization in barley grains germinated in the presence of cadmium and molybdenum. S. Afr. J. Bot. 2016, 106, 35–40. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotox. Environ. Safe 2017, 137, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.L.; Shang, S.H.; Zhang, H.T.; Jabeen, Z.; Zhang, G.P. Sodium chloride enhances cadmium tolerance through reducing cadmium accumulation and increasing anti-oxidative enzyme activity in tobacco. Environ. Toxicol. Chem. 2013, 32, 1420–1425. [Google Scholar] [CrossRef]
- Wei, T.; Liu, X.; Dong, M.; Lv, X.; Hua, L.; Jia, H.; Ren, X.; Yu, S.; Guo, J.; Li, Y. Rhizosphere iron and manganese-oxidizing bacteria stimulate root iron plaque formation and regulate Cd uptake of rice plants (Oryza sativa L.). J. Environ. Manag. 2021, 278, 111533. [Google Scholar] [CrossRef]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Arora, P.; Kumar, S.; Chaudhury, A. Perspectives for genetic engineering of poplars for enhanced phytoremediation abilities. Ecotoxicology 2010, 19, 1574–1588. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shi, C.; Liu, L.; Wang, C.; Qiao, W.; Gu, Z.; Wang, X. Lanthanum element induced imbalance of mineral nutrients, HSP 70 production and DNA-protein crosslink, Leading to hormetic response of cell cycle progression in root tips of Vicia faba L. seedlings. Dose-Response 2012, 10, 11–041. [Google Scholar] [CrossRef]
- Wu, Z.; Zhao, X.; Sun, X.; Tan, Q.; Tang, Y.; Nie, Z.; Qu, C.; Chen, Z.; Hu, C. Antioxidant enzyme systems and the ascorbate–glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 2015, 138, 526–536. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, X.; Hou, L.; Ji, S.; Zhang, Y.; Fan, W.; Li, T.; Zhang, L.; Liu, P.; Yang, L. Effects of cadmium on transcription, physiology, and ultrastructure of two tobacco cultivars. Sci. Total Environ. 2023, 869, 161751. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, R.H.; Nowroz, F.; Nahar, K. Insight into the physiological and biochemical mechanisms of biostimulating effect of Ascophyllum nodosum and Moringa oleifera extracts to minimize cadmium-induced oxidative stress in rice. Environ. Sci. Pollut. Res. 2023, 30, 55298–55313. [Google Scholar] [CrossRef]
- Wang, M.; Zou, J.; Duan, X.; Jiang, W.; Liu, D. Cadmium accumulation and its effects on metal uptake in maize (Zea mays L.). Bioresour. Technol. 2007, 98, 82–88. [Google Scholar] [CrossRef]
- Zhang, X.Z. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. Res. Methodol. Crop Physiol. 1992, 208–211. [Google Scholar]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 11, 764–775. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jin, C.W.; Liu, Y.; Mao, Q.Q.; Wang, Q.; Du, S.T. Mild Fe-deficiency improves biomass production and quality of hydroponic-cultivated spinach plants (Spinacia oleracea L.). Food Chem. 2013, 138, 2188–2194. [Google Scholar] [CrossRef] [PubMed]
- Groppa; Rosales, E.; Iannone, M.; Benavides, M. Nitric oxide, polyamines and Cd-induced phytotoxicity in wheat roots. Phytochemistry 2008, 69, 2609–2615. [Google Scholar] [CrossRef] [PubMed]
- Daud, M.K.; Sun, Y.; Dawood, M.; Hayat, Y.; Variath, M.T.; Wu, Y.X.; Mishkat, U.; Najeeb, U.; Zhu, S. Cadmium-induced functional and ultrastructural alterations in roots of two transgenic cotton cultivars. J. Hazard. Mater. 2009, 161, 463–473. [Google Scholar] [CrossRef]
- Bauddh, K.; Singh, R.P. Differential toxicity of cadmium to mustard (Brassica juncia L.) genotypes under higher metal levels. J. Environ. Biol. 2011, 32, 355. [Google Scholar]
- Hernandez, L.E.; Cooke, D.T. Modification of the root plasma membrane lipid composition of cadmium-treated. Pisum. Sativum. J. Exp. Bot. 1997, 48, 1375–1381. [Google Scholar] [CrossRef]
- Borges, K.L.R.; Salvato, F.; Alcântara, B.K.; Nalin, R.S.; Piotto, F.; Azevedo, R.A. Temporal dynamic responses of roots in contrasting tomato genotypes to cadmium tolerance. Ecotoxicology 2018, 27, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Xin, J.; Dai, H.; Liu, A.; Zhou, W.; Yi, Y.; Liao, K. Root morphological responses of three hot pepper cultivars to Cd exposure and their correlations with Cd accumulation. Environ. Sci. Pollut. Res. 2015, 22, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Bahn, M.; Lattanzi, F.A.; Hasibeder, R.; Wild, B.; Koranda, M.; Danese, V.; Brüggemann, N.; Schmitt, M.; Siegwolf, R.; Richter, A. Responses of belowground carbon allocation dynamics to extended shading in mountain grassland. New Phytol. 2013, 198, 116–126. [Google Scholar] [CrossRef]
- Muradoglu, F.; Gundogdu, M.; Ercisli, S.; Encu, T.; Balta, F.; Jaafar, H.Z.; Zia-Ul-Haq, M. Cadmium toxicity affects chlorophyll a and b content, antioxidant enzyme activities and mineral nutrient accumulation in strawberry. Biol. Res. 2015, 48, 11. [Google Scholar] [CrossRef]
- Lagriffoul, A.; Mocquot, B.; Mench, M.; Vangronsveld, J. Cadmium toxicity effects on growth, mineral and chlorophyll contents, and activities of stress related enzymes in young maize plants (Zea mays L.). Plant Soil. 1998, 200, 241–250. [Google Scholar] [CrossRef]
- Shackira, A.M.; Puthur, J.T. Enhanced phytostabilization of cadmium by a halophyte—Acanthus ilicifolius L. Int. J. Phytorem. 2017, 19, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Bankaji, I.; Caçador, I.; Sleimi, N. Physiological and biochemical responses of Suaeda fruticosa to cadmium and copper stresses: Growth, nutrient uptake, antioxidant enzymes, phytochelatin, and glutathione levels. Environ. Sci. Pollut. Res. 2015, 22, 13058–13069. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.T.; Akram, M.S.; Tanwir, K.; Chaudhary, H.J.; Ali, Q.; Stoltz, E.; Lindberg, S. Cadmium spiked soil modulates root organic acids exudation and ionic contents of two differentially Cd tolerant maize (Zea mays L.) cultivars. Ecotox. Environ. Safe 2017, 141, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, Q.; Diao, C. Effects of cadmium and arsenic on growth and metal accumulation of Cd-hyperaccumulator Solanum nigrum L. Bioresour. Technol. 2008, 99, 1103–1110. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Tanyolac, D.; Ayhan, B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of cadmium stress on growth and physiological characteristics of sassafras seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef]
- Grill, E.; Löffler, S.; Winnacker, E.L.; Zenk, M.H. Phytochelatins, the heavy-metal-binding peptides of plants, are synthesized from glutathione by a specific γ-glutamylcysteine dipeptidyl transpeptidase (phytochelatin synthase). Proc. Natl. Acad. Sci. USA 1989, 86, 6838–6842. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Chen, L. Growth and physiological responses of Pennisetum sp. to cadmium stress under three different soils. Environ. Sci. Pollut. Res. 2021, 28, 14867–14881. [Google Scholar] [CrossRef]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ. 2008, 54, 262. [Google Scholar] [CrossRef]
- Nedjimi, B.; Daoud, Y. Cadmium accumulation in Atriplex halimus subsp. schweinfurthii and its influence on growth, proline, root hydraulic conductivity and nutrient uptake. Flora 2009, 204, 316–324. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef]
- Wang, P.; Deng, X.; Huang, Y.; Fang, X.; Zhang, J.; Wan, H.; Yang, C. Root morphological responses of five soybean [Glycine max (L.) Merr] cultivars to cadmium stress at young seedlings. Environ. Sci. Pollut. Res. 2016, 23, 1860–1872. [Google Scholar] [CrossRef] [PubMed]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef]
- Haque, A.M.; Tasnim, J.; El-Shehawi, A.M.; Rahman, M.A.; Parvez, M.S.; Ahmed, M.B.; Kabir, A.H. The Cd-induced morphological and photosynthetic disruption is related to the reduced Fe status and increased oxidative injuries in sugar beet. Plant Physiol. Biochem. 2021, 166, 448–458. [Google Scholar] [CrossRef]
- Biswal, B.; Joshi, P.N.; Raval, M.K.; Biswal, U.C. Photosynthesis, a global sensor of environmental stress in green plants: Stress signalling and adaptation. Curr. Sci. India 2011, 47–56. [Google Scholar]
- Matysik, J.; Alia Bhalu, B.; Mohanty, P. Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr. Sci. 2002, 525–532. [Google Scholar]
- Gallardo, M.; Turner, N.C.; Ludwig, C. Water relations, gas exchange and abscisic acid content of Lupinus cosentinii leaves in response to drying different proportions of the root system. J. Exp. Bot. 1994, 45, 909–918. [Google Scholar] [CrossRef]
- Lin , R.; Wang, X.; Luo, Y.; Du, W.; Guo, H.; Yin, D. Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings (Triticum aestivum L.). Chemosphere 2007, 69, 89–98. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Treatment (mg L−1) | Shoot Fresh Weight (mg) | Root Fresh Weight (mg) | Shoot Dry Weight (mg) | Root Dry Weight (mg) | Root/Shoot |
|---|---|---|---|---|---|---|
| Kui3 | 0 | 1362.80 ± 69.96 a | 775.40 ± 42.56 a | 121.70 ± 8.56 a | 76.80 ± 5.62 a | 636.78 ± 41.03 b |
| 1 | 1094.60 ± 60.44 b | 740.30 ± 40.67 ab | 91.60 ± 4.45 b | 61.40 ± 3.46 b | 672.97 ± 30.79 b | |
| 3 | 944.70 ± 46.23 bc | 605.50 ± 12.87 bc | 88.90 ± 2.89 b | 65.70 ± 1.39 b | 740.23 ± 11.82 ab | |
| 5 | 835.30 ± 77.35c | 492.00 ± 87.38 c | 81.20 ± 7.98 b | 64.60 ± 0.70 b | 834.16 ± 81.29 a | |
| 7 | 808.00 ± 18.41 c | 563.40 ± 35.40 c | 87.70 ± 1.23 b | 58.20 ± 4.58 b | 666.03 ± 56.09 b | |
| CML118 | 0 | 979.20 ± 131.74 a | 545.00 ± 34.13 a | 95.10 ± 11.12 a | 53.00 ± 2.55 a | 587.93 ± 63.75 b |
| 1 | 701.30 ± 62.84 b | 489.70 ± 47.80 a | 70.90 ± 6.19 b | 52.10 ± 4.68 b | 745.37 ± 63.11 ab | |
| 3 | 548.10 ± 59.18 bc | 363.80 ± 18.40 b | 61.80 ± 4.72 bc | 44.20 ± 2.76 ab | 733.39 ± 68.16 ab | |
| 5 | 420.70 ± 55.01 c | 316.10 ± 33.07 b | 47.60 ± 5.96 cd | 37.80 ± 3.05 b | 875.45 ± 153.85 ab | |
| 7 | 321.90 ± 14.14 c | 292.60 ± 17.75 b | 40.30 ± 2.53 d | 36.60 ± 3.85 b | 932.00 ± 120.75 a | |
| Mo17 | 0 | 857.40 ± 48.39 a | 807.10 ± 64.82 a | 91.40 ± 3.97 a | 70.30 ± 7.41 a | 761.91 ± 58.98 b |
| 1 | 686.10 ± 73.92 b | 595.60 ± 39.18 b | 61.60 ± 7.84 b | 58.80 ± 3.93 ab | 1000.76 ± 94.81 a | |
| 3 | 511.10 ± 15.54 c | 564.90 ± 55.37 b | 54.60 ± 2.87 b | 56.80 ± 4.41 b | 1047.54 ± 76.77 a | |
| 5 | 475.80 ± 34.8 c | 519.90 ± 42.87 b | 51.50 ± 2.90 b | 55.20 ± 2.12 b | 1083.08 ± 60.38 a | |
| 7 | 474.70 ± 34.34 c | 513.60 ± 26.96 b | 47.10 ± 3.60 b | 50.70 ± 1.39 b | 1111.54 ± 90.50 a | |
| B73 | 0 | 1346.00 ± 84.12 a | 714.80 ± 56.21 a | 123.50 ± 7.32 a | 68.90 ± 4.77 a | 575.85 ± 69.01 b |
| 1 | 1229.10 ± 83.55 a | 697.30 ± 87.81 ab | 111.70 ± 7.45 a | 65.50 ± 7.91 ab | 578.32 ± 40.15 b | |
| 3 | 937.90 ± 33.10 b | 604.10 ± 58.95 ab | 94.50 ± 4.08 b | 60.10 ± 5.54 ab | 648.17 ± 73.52 b | |
| 5 | 934.10 ± 63.02 b | 604.00 ± 49.47 ab | 84.90 ± 5.58 b | 59.50 ± 5.48 ab | 703.95 ± 57.78 ab | |
| 7 | 565.10 ± 33.07 c | 513.40 ± 16.20 b | 60.50 ± 2.55 c | 51.30 ± 0.44 b | 855.05 ± 31.21 a | |
| B77 | 0 | 1223.20 ± 111.05 a | 640.90 ± 29.69 a | 95.40 ± 4.70 a | 65.00 ± 5.15 a | 697.33 ± 81.21 a |
| 1 | 1164.30 ± 94.68 a | 591.60 ± 29.83 a | 95.00 ± 3.54 a | 60.60 ± 2.46 a | 640.19 ± 30.61 a | |
| 3 | 1136.10 ± 53.40 a | 489.20 ± 42.19 b | 88.80 ± 4.98 ab | 55.00 ± 4.40 ab | 636.76 ± 79.22 a | |
| 5 | 807.40 ± 47.94 b | 431.50 ± 25.79 b | 76.50 ± 3.45 b | 49.10 ± 2.34 bc | 649.33 ± 44.16 a | |
| 7 | 567.50 ± 48.19 c | 381.40 ± 16.91 c | 58.70 ± 5.50 c | 43.10 ± 2.61 c | 778.86 ± 100.52 a |
| Lines | Treatment | RL | RD | SA | RV |
|---|---|---|---|---|---|
| (mg L−1) | (cm) | (cm) | (cm2) | (cm3) | |
| Kui3 | 0 | 759.17 ± 87.22 a | 1.29 ± 0.22 a | 30,601.08 ± 6048.20 a | 267,947.13 ± 95,851.08 ab |
| 1 | 780.29 ± 122.77 a | 1.35 ± 0.14 a | 32,998.30 ± 5986.54 a | 314,624.65 ± 101,084.07 a | |
| 3 | 457.49 ± 53.59 a | 1.45 ± 0.09 a | 20,381.30 ± 2646.48 ab | 171,209.33 ± 29,402.98 ab | |
| 5 | 271.75 ± 35.87 a | 1.28 ± 0.09 a | 10,699.13 ± 1752.38 b | 77,263.45 ± 16,943.58 b | |
| 7 | 420.69 ± 10.31 a | 1.52 ± 0.27 a | 19,858.77 ± 3538.09 ab | 195,720.43 ± 73,297.12 ab | |
| CML118 | 0 | 640.91 ± 92.25 b | 1.01 ± 0.13 b | 19,355.30 ± 2672.21 a | 130,650.78 ± 25,961.96 a |
| 1 | 442.76 ± 34.11 ab | 1.29 ± 0.09 ab | 17,637.38 ± 1719.65 ab | 141,580.28 ± 20,126.45 a | |
| 3 | 409.70 ± 80.53 ab | 1.26 ± 0.06 ab | 16,410.11 ± 3664.43 ab | 130,575.44 ± 39,205.36 a | |
| 5 | 214.18 ± 41.4 ab | 1.54 ± 0.17 ab | 10,875.39 ± 3046.29 b | 96,855.92 ± 38,513.72 a | |
| 7 | 184.15 ± 25.97 a | 1.91 ± 0.50 a | 9761.01 ± 1461.29 b | 92,493.01 ± 33,422.47 a | |
| Mo17 | 0 | 320.86 ± 30.28 b | 1.19 ± 0.09 b | 11,793.15 ± 1278.11 a | 82,311.92 ± 14,694.72 ab |
| 1 | 253.84 ± 26.92 a | 1.62 ± 0.05 a | 12,565.18 ± 1356.32 a | 108,211.69 ± 15,877.74 a | |
| 3 | 187.25 ± 10.18 ab | 1.33 ± 0.10 ab | 7707.00 ± 815.80 b | 52,205.94 ± 8313.46 bc | |
| 5 | 164.82 ± 15.03 b | 1.26 ± 0.13 b | 6598.41 ± 1260.84 b | 43,316.70 ± 13,194.67 c | |
| 7 | 122.11 ± 16.53 b | 1.28 ± 0.10 b | 4904.98 ± 911.39 b | 30,046.59 ± 7463.56 c | |
| B73 | 0 | 635.53 ± 73.71 b | 0.95 ± 0.07 b | 18,654.92 ± 2104.86 b | 11,8703.15 ± 21,136.51 b |
| 1 | 552.99 ± 42.97 a | 1.61 ± 0.05 a | 27,930.21 ± 2435.96 a | 266,296.66 ± 33,287.42 a | |
| 3 | 405.96 ± 42.92 a | 1.45 ± 0.11 a | 18,969.81 ± 3340.87 b | 152,477.86 ± 38,490.31 b | |
| 5 | 362.09 ± 24.09 ab | 1.29 ± 0.15 ab | 14,398.89 ± 2023.66 b | 105,023.35 ± 30,631.84 b | |
| 7 | 307.59 ± 30.31 a | 1.43 ± 0.21 a | 14,141.16 ± 3241.42 b | 118,651.97 ± 48,260.56 b | |
| B77 | 0 | 635.72 ± 92.05 a | 1.39 ± 0.17 a | 25,700.37 ± 2733.05 a | 216,865.12 ± 28,226.93 a |
| 1 | 557.89 ± 66.87 a | 1.22 ± 0.05 a | 20,992.72 ± 2428.58 ab | 160,270.98 ± 33,610.14 a | |
| 3 | 441.92 ± 55.45 a | 1.25 ± 0.07 a | 17,126.24 ± 2651.07 b | 138,125.06 ± 40,348.61 ab | |
| 5 | 368.53 ± 32.91 a | 1.30 ± 0.15 a | 15,556.21 ± 2719.56 b | 125,692.55 ± 33,952.36 ab | |
| 7 | 235.84 ± 34.53 a | 1.08 ± 0.05 a | 7861.72 ± 1162.25 c | 46,404.89 ± 7928.91 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, S.; Wu, Y.; Zeng, Q.; Zhang, A.; Duan, M.; Deng, M. Effects of Cd Stress on Morphological and Physiological Characteristics of Maize Seedlings. Agronomy 2024, 14, 379. https://doi.org/10.3390/agronomy14020379
Deng S, Wu Y, Zeng Q, Zhang A, Duan M, Deng M. Effects of Cd Stress on Morphological and Physiological Characteristics of Maize Seedlings. Agronomy. 2024; 14(2):379. https://doi.org/10.3390/agronomy14020379
Chicago/Turabian StyleDeng, Siqing, Yueying Wu, Qingping Zeng, Aoni Zhang, Meijuan Duan, and Min Deng. 2024. "Effects of Cd Stress on Morphological and Physiological Characteristics of Maize Seedlings" Agronomy 14, no. 2: 379. https://doi.org/10.3390/agronomy14020379
APA StyleDeng, S., Wu, Y., Zeng, Q., Zhang, A., Duan, M., & Deng, M. (2024). Effects of Cd Stress on Morphological and Physiological Characteristics of Maize Seedlings. Agronomy, 14(2), 379. https://doi.org/10.3390/agronomy14020379