
Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry


 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Trait Investigation and Phenotyping
2.3. Statistical Analysis
3. Results
3.1. Phenotypic Variation of Agronomic Traits
3.1.1. Fruit-Related Traits
Fruit Size
Fruit Color
Fruit Flavor
Fruit Shape
Fruit Stalk
3.1.2. Phenological Traits and Flower-Related Traits
3.2. Inheritance Trend of Agronomic Traits
3.2.1. Fruit-Related Traits
3.2.2. Phenological Traits and Flower-Related Traits
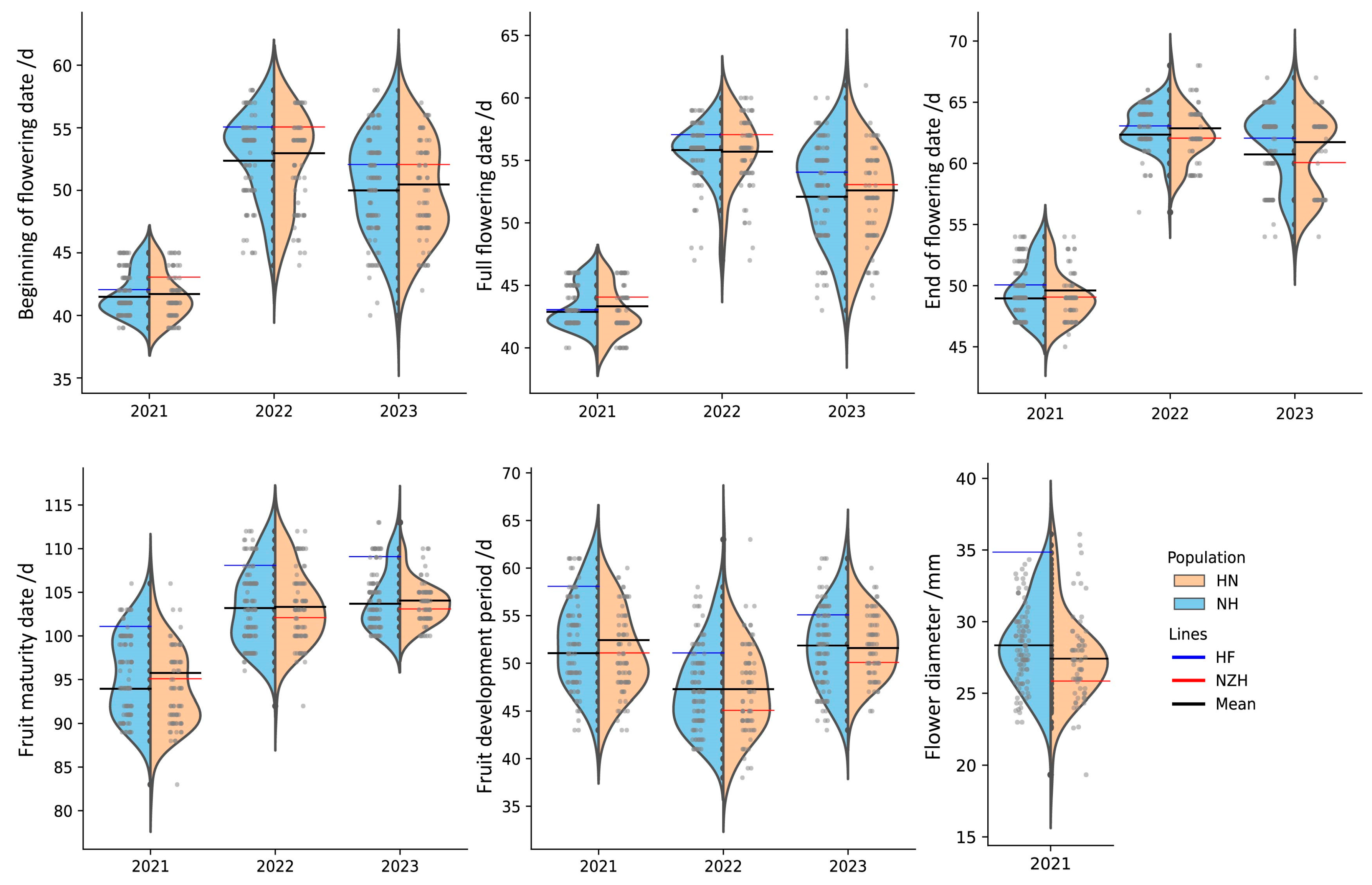
3.3. Phenotypic Difference Between NH and HN Populations
3.4. Trait Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yü, D.J.; Lu, L.T.; Gu, C.Z.; Guan, K.J.; Li, C.L.; Chen, S.H. Flora Reipublicae Popularis Sinicae, 38; Science Press: Beijing, China, 1986. [Google Scholar]
- Cao, J.; Jiang, Q.; Lin, J.; Li, X.; Sun, C.; Chen, K. Physicochemical characterisation of four cherry species (Prunus spp.) grown in China. Food Chem. 2015, 173, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.S.; Wang, H.; Zhang, J.; Chen, Q.; He, W.; Zhang, Y.; Luo, Y.; Tang, H.R.; Wang, Y.; Wang, X.R. Comparative metabolomics profiling highlights unique color variation and bitter taste formation of Chinese cherry fruits. Food Chem. 2024, 439, 138072. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.S.; Bernard, A.; Wang, Y.; Dirlewanger, E.; Wang, X. Genomes and integrative genomic insights into the genetic architecture of main agronomic traits in the edible cherries. Hortic. Res. 2024, e269. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Zhang, J.; Liu, Z.S.; Chen, Q.; He, W.; Yang, S.F.; Lin, Y.X.; Zhang, Y.T.; Li, M.Y.; et al. Survey on intra-specific crossing and F1 seedling cultivation in seven combinations of Chinese cherry. Hortic. J. 2022, 91, 267–275. [Google Scholar] [CrossRef]
- Fortuny, A.P.; Bueno, R.A.; Pereira Da Costa, J.H.; Zanor, M.I.; Rodríguez, G.R.; Gibon, Y. Tomato fruit quality traits and metabolite content are affected by reciprocal crosses and heterosis. J. Exp. Bot. 2021, 72, 5407–5425. [Google Scholar] [CrossRef]
- Gonzalo, M.; Vyn, T.J.; Holland, J.B.; Mcintyre, L.M. Mapping reciprocal effects and interactions with plant density stress in Zea mays L. Heredity 2007, 99, 14–30. [Google Scholar] [CrossRef]
- Rapoport, H.F.; Moreno-Alias, I.; de la Rosa-Peinazo, M.A.; Frija, A.; de la Rosa, R.; Leon, L. Floral quality characterization in Olive progenies from reciprocal crosses. Plants 2022, 11, 1285. [Google Scholar] [CrossRef]
- Naves, E.R.; Scossa, F.; Araújo, W.L.; Nunes-Nesi, A.; Fernie, A.R.; Zsögön, A. Heterosis and reciprocal effects for agronomic and fruit traits in Capsicum pepper hybrids. Sci. Hortic. 2022, 295, 110821. [Google Scholar] [CrossRef]
- Zheng, X.; Yue, C.; Gallardo, K.; Mccracken, V.; Luby, J.; Mcferson, J. What attributes are consumers looking for in sweet cherries? evidence from choice experiments. Agric. Resour. Econ. Rev. 2016, 45, 124–142. [Google Scholar] [CrossRef]
- Mackay, T.F.C.; Stone, E.A.; Ayroles, J.F. The genetics of quantitative traits: Challenges and prospects. Nat. Rev. Genet. 2009, 10, 565–577. [Google Scholar] [CrossRef]
- Paril, J.; Reif, J.; Fournier Level, A.; Pourkheirandish, M. Heterosis in crop improvement. Plant J. 2024, 117, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Le Dantec, L.; Barreneche, T.; Dirlewanger, E.; Quero-García, J. New insights into fruit firmness and weight control in sweet cherry. Plant Mol. Biol. Rep. 2015, 33, 783–796. [Google Scholar] [CrossRef]
- Piaskowski, J.; Hardner, C.; Cai, L.; Zhao, Y.; Iezzoni, A.; Peace, C. Genomic heritability estimates in sweet cherry reveal non-additive genetic variance is relevant for industry-prioritized traits. BMC Genet. 2018, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Calle, A.; Wünsch, A. Multiple-population QTL mapping of maturity and fruit-quality traits reveals LG4 region as a breeding target in sweet cherry (Prunus avium L.). Hortic. Res. 2020, 7, 127. [Google Scholar] [CrossRef]
- .Calle, A.; Cai, L.; Iezzoni, A.; Wünsch, A. Genetic dissection of bloom time in low chilling sweet cherry (Prunus avium L.) using a multi-family QTL approach. Front. Plant Sci. 2020, 10, 1647. [Google Scholar] [CrossRef]
- Branchereau, C.; Hardner, C.; Dirlewanger, E.; Wenden, B.; Le Dantec, L.; Alletru, D.; Parmentier, J.; Ivančič, A.; Giovannini, D.; Brandi, F.; et al. Genotype-by-environment and QTL-by-environment interactions in sweet cherry (Prunus avium L.) for flowering date. Front. Plant Sci. 2023, 14, 1142974. [Google Scholar] [CrossRef]
- Szilágyi, S.; Horváth-Kupi, T.; Desiderio, F.; Bekefi, Z. Evaluation of sweet cherry (Prunus avium L.) cultivars for fruit size by FW_G2a QTL analysis and phenotypic characterization. Sci. Hortic. 2022, 292, 110656. [Google Scholar] [CrossRef]
- Calle, A.; Balas, F.; Cai, L.; Iezzoni, A.; López-Corrales, M.; Serradilla, M.J.; Wünsch, A. Fruit size and firmness QTL alleles of breeding interest identified in a sweet cherry ‘Ambrunés’ × ‘Sweetheart’ population. Mol. Breeding 2020, 40, 86. [Google Scholar] [CrossRef]
- Cai, L.; Quero-García, J.; Barreneche, T.; Dirlewanger, E.; Saski, C.; Iezzoni, A. A fruit firmness QTL identified on linkage group 4 in sweet cherry (Prunus avium L.) is associated with domesticated and bred germplasm. Sci. Rep. 2019, 9, 5008. [Google Scholar] [CrossRef]
- Hardner, C.M.; Hayes, B.J.; Kumar, S.; Vanderzande, S.; Cai, L.; Piaskowski, J.; Quero-Garcia, J.; Campoy, J.A.; Barreneche, T.; Giovannini, D.; et al. Prediction of genetic value for sweet cherry fruit maturity among environments using a 6K SNP array. Hortic. Res. 2019, 6, 6–15. [Google Scholar] [CrossRef]
- Mgbechi-Ezeri, J.; Porter, L.; Johnson, K.B.; Oraguzie, N. Assessment of sweet cherry (Prunus avium L.) genotypes for response to bacterial canker disease. Euphytica 2017, 213, 1. [Google Scholar] [CrossRef]
- Quero-García, J.; Letourmy, P.; Campoy, J.A.; Branchereau, C.; Malchev, S.; Barreneche, T.; Dirlewanger, E. Multi-year analyses on three populations reveal the first stable QTLs for tolerance to rain-induced fruit cracking in sweet cherry (Prunus avium L.). Hortic. Res. 2021, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.S.; Yang, X.Q.; Wang, Z.Y.; Ma, L.; Tu, H.X.; Ma, Y.; Zhou, J.T.; Zhang, J.; Wang, H.; et al. Inheritance analysis of fruit-related traits in Chinese cherry [Cerasus pseudocerasus (Lindl.) G.Don] breeding progenies. Sci. Hortic. 2023, 307, 111519. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, G.P.; Liu, Z.S.; Zhang, J.; Ma, L.; Tian, T.; Wang, H.; Chen, T.; Chen, Q.; He, W.; et al. Phenotyping in flower and main fruit traits of Chinese cherry [Cerasus pseudocerasus (Lindl.) G.Don]. Sci. Hortic. 2022, 296, 110920. [Google Scholar] [CrossRef]
- Karagiannis, E.; Sarrou, E.; Michailidis, M.; Tanou, G.; Ganopoulos, I.; Bazakos, C.; Kazantzis, K.; Martens, S.; Xanthopoulou, A.; Molassiotis, A. Fruit quality trait discovery and metabolic profiling in sweet cherry genebank collection in Greece. Food Chem. 2021, 342, 128315. [Google Scholar] [CrossRef]
- Hou, F.L. Plant Physiology Experiment; Science Press: Beijing, China, 2015. [Google Scholar]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower development in sweet cherry framed in the BBCH scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv Prepr. 2014, arXiv:1406.5823. [Google Scholar]
- Joseph, B.; Corwin, J.A.; Li, B.; Atwell, S.; Kliebenstein, D.J. Cytoplasmic genetic variation and extensive cytonuclear interactions influence natural variation in the metabolome. Elife 2013, 2, e776. [Google Scholar] [CrossRef]
- Asins, M.J.; Raga, V.; Bernet, G.P.; Carbonell, E.A. Genetic analysis of reproductive, vegetative and fruit quality traits to improve Citrus varieties. Tree Genet. Genomes 2015, 11, 117. [Google Scholar] [CrossRef]
- Zinelabidine, L.H.; Torres-Pérez, R.; Grimplet, J.; Baroja, E.; Ibáñez, S.; Carbonell-Bejerano, P.; Martínez-Zapater, J.M.; Ibáñez, J.; Tello, J. Genetic variation and association analyses identify genes linked to fruit set-related traits in grapevine. Plant Sci. 2021, 306, 110875. [Google Scholar] [CrossRef]
- Fortuny, A.P.; Mengarelli, D.A.; Pereira Da Costa, J.H.; Rodríguez, G.R.; Zanor, M.I. Reciprocal effect and heterosis for tomato fruit metabolites revealed by whole transcriptomic analysis of two cultivars and their reciprocal hybrids. Sci. Hortic. 2023, 308, 111583. [Google Scholar] [CrossRef]
- Liu, J.C.; Zhang, Q.P.; Niu, T.Q.; Liu, N.; Zhang, Y.P.; Xu, M.; Ma, X.X.; Zhang, Y.J.; Liu, S.; Liu, W.S. Analysis of inherited tendency of fruit characteristics in F1 group of reciprocal crossing between "Chuanzhihong" and "Saimaiti" in apricots. J. Fruit Sci. 2020, 37, 625–634. [Google Scholar]
- Lei, L.; Xu, G.H.; Wang, L.; Du, Q.H.; Li, Y.F.; Wang, H.X. Screening and inheritance of fruit storage-related traits based on reciprocal cross of Southern×Northern high bush blueberry (Vaccinium linn). Sci. Agric. Sinica 2020, 53, 4045. [Google Scholar] [CrossRef]
- Montanari, S.; Thomson, S.; Cordiner, S.; Günther, C.S.; Miller, P.; Deng, C.H.; Mcghie, T.; Knäbel, M.; Foster, T.; Turner, J.; et al. High-density linkage map construction in an autotetraploid blueberry population and detection of quantitative trait loci for anthocyanin content. Front. Plant Sci. 2022, 13, 965397. [Google Scholar] [CrossRef] [PubMed]
- Rawandoozi, Z.J.; Hartmann, T.P.; Carpenedo, S.; Gasic, K.; Da Silva Linge, C.; Cai, L.; Van de Weg, E.; Byrne, D.H. Identification and characterization of QTLs for fruit quality traits in peach through a multi-family approach. BMC Genomics 2020, 21, 522. [Google Scholar] [CrossRef]
- Valderrama-Soto, D.; Salazar, J.; Sepúlveda-González, A.; Silva-Andrade, C.; Gardana, C.; Morales, H.; Battistoni, B.; Jiménez-Muñoz, P.; González, M.; Peña-Neira, Á.; et al. Detection of quantitative trait loci controlling the content of phenolic compounds in an Asian plum (Prunus salicina L.) F1 population. Front. Plant Sci. 2021, 12, 679059. [Google Scholar] [CrossRef]
- Salazar, J.A.; Pacheco, I.; Shinya, P.; Zapata, P.; Silva, C.; Aradhya, M.; Velasco, D.; Ruiz, D.; Martínez-Gómez, P.; Infante, R. Genotyping by sequencing for SNP-based linkage analysis and identification of QTLs linked to fruit quality traits in Japanese plum (Prunus salicina Lindl.). Front. Plant Sci. 2017, 8, 476. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Liu, W.; Liu, N.; Zhang, Y.; Xu, M.; Liu, S.; Ma, X.; Zhang, Y. Construction of a high-density genetic map and identification of quantitative trait loci linked to fruit quality traits in apricots using specific-locus amplified fragment sequencing. Front. Plant Sci. 2022, 13, 798700. [Google Scholar] [CrossRef]
- Xuesen, C.; Nan, W.; Zongying, Z.; Zhiquan, M.; Chengmiao, Y. Understanding and thinking about some problems of fruit tree germplasm resources and genetic breeding. Sci. Agric. Sinica 2022, 55, 3395–3410. [Google Scholar] [CrossRef]
- Zhang, G.; Sebolt, A.M.; Sooriyapathirana, S.S.; Wang, D.; Bink, M.C.; Olmstead, J.W.; Iezzoni, A.F. Fruit size QTL analysis of an F1 population derived from a cross between a domesticated sweet cherry cultivar and a wild forest sweet cherry. Tree Genet. Genomes 2010, 6, 25–36. [Google Scholar] [CrossRef]
- Rosyara, U.R.; Bink, M.C.A.M.; van de Weg, E.; Zhang, G.; Wang, D.; Sebolt, A.; Dirlewanger, E.; Quero-Garcia, J.; Schuster, M.; Iezzoni, A.F. Fruit size QTL identification and the prediction of parental QTL genotypes and breeding values in multiple pedigreed populations of sweet cherry. Mol. Breeding 2013, 32, 875–887. [Google Scholar] [CrossRef]
- Sanchez-Perez, R.; Pavan, S.; Mazzeo, R.; Moldovan, C.; Aiese, C.R.; Del, C.J.; Ricciardi, F.; Lotti, C.; Ricciardi, L.; Dicenta, F.; et al. Mutation of a bHLH transcription factor allowed almond domestication. Science 2019, 364, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, W.; Pan, Y.; Ge, L.; Wu, C.; Wang, J.; Liu, M.; Yan, F. Comparison and genetic variation analysis of important fruit traits in jujube F1 hybrids by different male parents. Agronomy 2024, 14, 459. [Google Scholar] [CrossRef]
- Altan, H.; Bircan, M.; Caliskan, O. Inheritance of earliness and fruit quality characteristics in five apricot progenies. Erwerbsobstbau 2022, 64, 591–601. [Google Scholar] [CrossRef]
- Desnoues, E.; Baldazzi, V.; Génard, M.; Mauroux, J.; Lambert, P.; Confolent, C.; Quilot-Turion, B. Dynamic QTLs for sugars and enzyme activities provide an overview of genetic control of sugar metabolism during peach fruit development. J. Exp. Bot. 2016, 67, 3419–3431. [Google Scholar] [CrossRef]
- Castede, S.; Campoy, J.A.; Garcia, J.Q.; Le Dantec, L.; Lafargue, M.; Barreneche, T.; Wenden, B.; Dirlewanger, E. Genetic determinism of phenological traits highly affected by climate change in Prunus avium: Flowering date dissected into chilling and heat requirements. New Phytol. 2014, 202, 703–715. [Google Scholar] [CrossRef]
- Cai, L.; Stegmeir, T.; Sebolt, A.; Zheng, C.; Bink, M.C.A.M.; Iezzoni, A. Identification of bloom date QTLs and haplotype analysis in tetraploid sour cherry (Prunus cerasus). Tree Genet. Genomes 2018, 14, 22. [Google Scholar] [CrossRef]
- Dirlewanger, E.; Quero-Garcia, J.; Le Dantec, L.; Lambert, P.; Ruiz, D.; Dondini, L.; Illa, E.; Quilot-Turion, B.; Audergon, J.M.; Tartarini, S.; et al. Comparison of the genetic determinism of two key phenological traits, flowering and maturity dates, in three Prunus species: Peach, apricot and sweet cherry. Heredity 2012, 109, 280–292. [Google Scholar] [CrossRef]
- Salazar, J.A.; Ruiz, D.; Campoy, J.A.; Tartarini, S.; Dondini, L.; Martínez-Gómez, P. Inheritance of reproductive phenology traits and related QTL identification in apricot. Tree Genet. Genomes 2016, 12, 1. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J.; Rees, D.J.G.; Celton, J.M.; Martínez-Gómez, P. Inheritance of flowering time in apricot (Prunus armeniaca L.) and analysis of linked quantitative trait loci (QTLs) using simple sequence repeat (SSR) markers. Plant Mol. Biol. Rep. 2011, 29, 404–410. [Google Scholar] [CrossRef]
- Kitamura, Y.; Habu, T.; Yamane, H.; Nishiyama, S.; Kajita, K.; Sobue, T.; Kawai, T.; Numaguchi, K.; Nakazaki, T.; Kitajima, A.; et al. Identification of QTLs controlling chilling and heat requirements for dormancy release and bud break in Japanese apricot (Prunus mume). Tree Genet. Genomes 2018, 14, 33. [Google Scholar] [CrossRef]
- Ballester, J.; Socias, I. Company, R.; Arus, P.; De Vicente, M.C. Genetic mapping of a major gene delaying blooming time in almond. Plant Breeding 2001, 120, 268–270. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category/Trait | Abbreviation | Year | Cross | Sample Size | Mean | Max | Min | CV/% | K | SK |
|---|---|---|---|---|---|---|---|---|---|---|
| Fruit size | ||||||||||
| Fruit longitude diameter/mm | FLoD | 2021 | NH | 108 | 19.43 | 24.66 | 15.99 | 8.95 | 0.37 | 0.53 |
| HN | 77 | 18.99 | 23.82 | 16.15 | 7.79 | 1.18 | 0.76 | |||
| 2022 | NH | 114 | 17.43 | 20.52 | 13.92 | 8.44 | −0.64 | −0.06 | ||
| HN | 87 | 17.41 | 21.02 | 13.09 | 7.85 | 1.02 | −0.17 | |||
| 2023 | NH | 108 | 17.77 | 21.91 | 11.96 | 10.21 | 0.46 | −0.23 | ||
| HN | 68 | 18.23 | 22.00 | 13.23 | 7.89 | 1.42 | −0.39 | |||
| Fruit transverse diameter/mm | FTraD | 2021 | NH | 108 | 19.86 | 23.83 | 15.59 | 7.76 | 0.14 | 0.33 |
| HN | 77 | 18.36 | 22.38 | 14.13 | 8.98 | 0.11 | 0.02 | |||
| 2022 | NH | 114 | 17.87 | 21.67 | 14.39 | 6.91 | 0.27 | 0.18 | ||
| HN | 87 | 18.12 | 21.48 | 14.65 | 7.34 | 0.38 | −0.16 | |||
| 2023 | NH | 108 | 18.54 | 21.55 | 12.05 | 8.79 | 2.24 | −0.89 | ||
| HN | 68 | 19.20 | 21.99 | 16.12 | 6.80 | −0.07 | −0.24 | |||
| Fruit lateral diameter/mm | FLaD | 2021 | NH | 108 | 17.30 | 21.07 | 14.35 | 7.83 | 0.08 | 0.47 |
| HN | 77 | 16.86 | 20.44 | 14.12 | 8.20 | −0.03 | 0.36 | |||
| 2022 | NH | 114 | 15.70 | 18.66 | 12.87 | 7.16 | 0.01 | 0.30 | ||
| HN | 87 | 15.92 | 19.24 | 13.01 | 6.93 | 0.78 | −0.02 | |||
| 2023 | NH | 108 | 16.44 | 19.77 | 10.69 | 9.23 | 1.64 | −0.74 | ||
| HN | 68 | 17.04 | 19.53 | 13.31 | 7.71 | 0.35 | −0.54 | |||
| Fruit weight/g | FW | 2021 | NH | 107 | 4.35 | 7.46 | 2.59 | 21.68 | 0.83 | 0.86 |
| HN | 77 | 4.06 | 6.48 | 2.45 | 19.20 | 0.10 | 0.28 | |||
| 2022 | NH | 114 | 3.29 | 5.09 | 1.81 | 18.90 | 0.06 | 0.49 | ||
| HN | 87 | 3.35 | 5.23 | 1.85 | 19.42 | 0.40 | 0.19 | |||
| 2023 | NH | 109 | 3.73 | 6.36 | 1.80 | 21.47 | 0.44 | 0.31 | ||
| HN | 68 | 3.88 | 5.85 | 2.32 | 17.17 | 0.30 | 0.13 | |||
| Stone weight/g | FW (stone) | 2021 | NH | 102 | 0.20 | 0.31 | 0.10 | 23.00 | −0.44 | 0.26 |
| HN | 73 | 0.20 | 0.33 | 0.11 | 21.87 | 0.51 | 0.51 | |||
| Fruit color | ||||||||||
| Anthocyanin /mg·kg−1 | An | 2021 | NH | 66 | 24.42 | 112.51 | 3.12 | 86.63 | 5.75 | 2.23 |
| HN | 35 | 22.27 | 79.94 | 1.80 | 75.24 | 3.43 | 1.73 | |||
| a | a | 2022 | NH | 114 | 22.52 | 29.22 | 5.81 | 21.55 | 2.76 | −1.58 |
| HN | 87 | 22.89 | 29.40 | 8.66 | 16.67 | 1.60 | −1.04 | |||
| b | b | 2022 | NH | 114 | 10.41 | 17.85 | 0.67 | 33.91 | 0.66 | −0.69 |
| HN | 87 | 10.25 | 14.58 | 1.95 | 27.13 | 0.40 | −0.92 | |||
| L | L | 2022 | NH | 114 | 33.53 | 42.62 | 25.01 | 11.40 | −0.10 | 0.02 |
| HN | 87 | 33.62 | 40.87 | 26.61 | 9.72 | −0.57 | −0.17 | |||
| Fruit flavor | ||||||||||
| Total soluble solid/% | TSS | 2021 | NH | 107 | 14.47 | 19.65 | 8.13 | 14.83 | 0.06 | 0.13 |
| HN | 78 | 14.52 | 23.19 | 9.21 | 14.75 | 2.76 | 0.83 | |||
| 2022 | NH | 113 | 17.39 | 24.25 | 12.50 | 13.97 | 0.13 | 0.47 | ||
| HN | 87 | 17.29 | 22.15 | 13.22 | 11.49 | −0.27 | 0.41 | |||
| 2023 | NH | 109 | 16.35 | 22.50 | 12.38 | 14.66 | −0.34 | 0.54 | ||
| HN | 68 | 15.84 | 21.37 | 12.55 | 12.14 | 0.63 | 0.92 | |||
| Titratable acid/% | TA | 2021 | NH | 43 | 0.60 | 1.13 | 0.31 | 33.46 | 0.00 | 0.69 |
| HN | 27 | 0.59 | 0.96 | 0.25 | 25.46 | 0.70 | −0.07 | |||
| 2022 | NH | 113 | 0.54 | 1.01 | 0.30 | 21.27 | 1.91 | 0.92 | ||
| HN | 87 | 0.55 | 0.92 | 0.33 | 23.13 | −0.48 | 0.25 | |||
| 2023 | NH | 109 | 0.54 | 0.89 | 0.32 | 22.70 | 0.04 | 0.61 | ||
| HN | 68 | 0.49 | 0.77 | 0.26 | 19.03 | 0.46 | 0.27 | |||
| TSS/TA | TSS/TA | 2021 | NH | 43 | 26.36 | 45.59 | 9.55 | 37.64 | −0.87 | 0.34 |
| HN | 26 | 24.47 | 51.20 | 15.21 | 35.28 | 3.34 | 1.82 | |||
| 2022 | NH | 113 | 33.50 | 56.82 | 16.84 | 18.99 | 0.86 | 0.48 | ||
| HN | 87 | 33.05 | 49.82 | 21.02 | 20.98 | −0.27 | 0.50 | |||
| 2023 | NH | 109 | 31.78 | 59.69 | 16.82 | 22.81 | 1.33 | 0.78 | ||
| HN | 70 | 33.71 | 57.97 | 22.89 | 19.03 | 3.04 | 1.30 | |||
| pH | pH | 2021 | NH | 43 | 4.00 | 4.47 | 3.43 | 6.52 | −0.53 | −0.05 |
| HN | 27 | 4.00 | 4.71 | 3.76 | 5.62 | 2.89 | 1.60 | |||
| Soluble sugar /g·kg−1 | SS | 2021 | NH | 66 | 116.44 | 222.96 | 56.57 | 24.21 | 2.32 | 0.90 |
| HN | 35 | 123.12 | 199.76 | 71.91 | 26.52 | −0.46 | 0.26 | |||
| Fruit shape | ||||||||||
| Fruit shape index | FSh (index) | 2021 | NH | 108 | 0.98 | 1.12 | 0.87 | 5.69 | −0.35 | 0.17 |
| HN | 77 | 0.98 | 1.12 | 0.87 | 4.81 | 0.62 | 0.60 | |||
| 2022 | NH | 114 | 0.98 | 1.15 | 0.82 | 5.77 | 0.60 | 0.37 | ||
| HN | 87 | 0.96 | 1.09 | 0.78 | 4.99 | 1.98 | −0.23 | |||
| 2023 | NH | 108 | 0.96 | 1.10 | 0.85 | 5.22 | 0.09 | 0.49 | ||
| HN | 68 | 0.95 | 1.07 | 0.82 | 4.53 | 0.69 | 0.01 | |||
| Fruit stalk length/mm | FSL | 2021 | NH | 108 | 20.25 | 35.30 | 7.24 | 20.47 | 1.83 | 0.38 |
| HN | 76 | 19.38 | 30.01 | 9.93 | 21.26 | 0.15 | 0.40 | |||
| 2022 | NH | 114 | 22.62 | 32.18 | 13.30 | 15.96 | −0.13 | 0.33 | ||
| HN | 87 | 21.95 | 31.55 | 14.68 | 18.58 | −0.35 | 0.37 | |||
| 2023 | NH | 108 | 23.08 | 38.05 | 13.67 | 18.32 | 1.52 | 0.88 | ||
| HN | 68 | 22.84 | 35.69 | 14.28 | 18.64 | 0.61 | 0.73 | |||
| Phenology (flower and fruit) | ||||||||||
| Beginning of flowering date/d | FD (beginning) | 2021 | NH | 109 | 41.69 | 45 | 39 | 4.06 | −0.66 | 0.71 |
| HN | 77 | 41.47 | 45 | 39 | 4.30 | −0.52 | 0.58 | |||
| 2022 | NH | 97 | 52.94 | 58 | 45 | 6.13 | 0.04 | −0.76 | ||
| HN | 70 | 52.30 | 57 | 44 | 7.06 | −0.87 | −0.49 | |||
| 2023 | NH | 113 | 50.44 | 58 | 40 | 7.73 | −0.58 | −0.29 | ||
| HN | 76 | 50.03 | 57 | 42 | 7.47 | −1.00 | 0.08 | |||
| Full flowering date/d | FD (full) | 2021 | NH | 109 | 43.31 | 46 | 40 | 3.81 | −1.09 | 0.54 |
| HN | 76 | 42.86 | 46 | 40 | 4.24 | −0.64 | 0.43 | |||
| 2022 | NH | 102 | 55.68 | 59 | 47 | 4.01 | 3.60 | −1.48 | ||
| HN | 80 | 55.80 | 60 | 47 | 4.84 | 1.22 | −0.93 | |||
| 2023 | NH | 113 | 52.59 | 60 | 43 | 7.04 | −0.44 | −0.43 | ||
| HN | 74 | 52.07 | 61 | 44 | 6.69 | −0.34 | −0.06 | |||
| End of flowering date/d | FD (end) | 2021 | NH | 109 | 49.60 | 54 | 47 | 4.22 | −0.68 | 0.52 |
| HN | 76 | 48.93 | 54 | 45 | 3.94 | 0.57 | 0.89 | |||
| 2022 | NH | 103 | 62.88 | 66 | 56 | 2.69 | 1.81 | −0.83 | ||
| HN | 83 | 62.36 | 68 | 59 | 3.34 | 0.18 | 0.21 | |||
| 2023 | NH | 113 | 61.73 | 67 | 54 | 4.65 | −0.08 | −0.94 | ||
| HN | 67 | 60.49 | 67 | 54 | 5.29 | −1.55 | −0.18 | |||
| Fruit maturity date/d | MD | 2021 | NH | 108 | 95.72 | 106 | 89 | 4.76 | −1.24 | 0.14 |
| HN | 78 | 93.85 | 106 | 83 | 4.61 | −0.25 | 0.37 | |||
| 2022 | NH | 114 | 103.28 | 112 | 96 | 4.08 | −0.93 | 0.32 | ||
| HN | 87 | 103.16 | 112 | 92 | 3.96 | −0.28 | 0.23 | |||
| 2023 | NH | 109 | 104.00 | 113 | 100 | 3.22 | −0.25 | 0.83 | ||
| HN | 71 | 103.65 | 110 | 100 | 2.18 | 1.00 | 0.73 | |||
| Fruit development period/d | FDP | 2021 | NH | 108 | 52.41 | 61 | 43 | 8.59 | −0.72 | 0.15 |
| HN | 76 | 50.97 | 60 | 43 | 8.14 | −0.73 | 0.26 | |||
| 2022 | NH | 102 | 47.25 | 58 | 41 | 9.07 | −0.45 | 0.51 | ||
| HN | 80 | 47.25 | 63 | 38 | 9.70 | 0.54 | 0.48 | |||
| 2023 | NH | 109 | 51.56 | 61 | 43 | 8.02 | −0.58 | 0.11 | ||
| HN | 67 | 51.82 | 60 | 45 | 6.43 | −0.52 | 0.11 | |||
| Flower | ||||||||||
| Flower diameter /mm | FlD | 2021 | NH | 95 | 28.31 | 34.33 | 23.00 | 9.63 | −0.50 | 0.23 |
| HN | 66 | 27.34 | 36.10 | 19.33 | 10.48 | 1.78 | 0.62 | |||
| Trait | Abbr. | Year | Parent | Cross | Sample Size | Frequency Distribution of F1 Population (%) | Mean | H2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NZH | HF | MP | 1 | 2 | 3 | 4 | 5 | 6 | |||||||
| Fruit color, skin | FC (skin) | 2021 | 3 | 5 | 4 | NH | 106 | 1 (0.94) | 23 (21.70) | 62 (58.49) | 14 (13.21) | 6 (5.66) | 4.01 | NH = 0.93; HN = 0.89 | |
| HN | 74 | 14 (18.92) | 49 (66.22) | 10 (13.51) | 1 (1.35) | 3.97 | |||||||||
| 2022 | 3 | 5 | 4 | NH | 114 | 1 (0.88) | 15 (13.16) | 75 (65.79) | 17 (14.91) | 6 (5.26) | 4.11 | ||||
| HN | 87 | 10 (11.49) | 63 (72.41) | 13 (14.94) | 1 (1.15) | 4.06 | |||||||||
| 2023 | 3 | 5 | 4 | NH | 108 | 3 (2.78) | 25 (23.15) | 61 (56.48) | 15 (13.89) | 4 (3.70) | 3.93 | ||||
| HN | 68 | 13 (19.12) | 40 (58.82) | 15 (22.06) | 4.03 | ||||||||||
| Fruit color, flesh | FC (flesh) | 2021 | 1 | 3 | 2 | NH | 107 | 26 (24.30) | 61 (57.01) | 16 (14.95) | 4 (3.74) | 1.98 | NH = 0.88; HN = 0.67 | ||
| HN | 78 | 18 (23.08) | 52 (66.67) | 5 (6.41) | 3 (3.85) | 1.91 | |||||||||
| 2022 | 1 | 3 | 2 | NH | 114 | 37 (32.46) | 56 (49.12) | 18 (15.79) | 3 (2.63) | 1.89 | |||||
| HN | 86 | 31 (36.05) | 42 (48.84) | 11 (12.79) | 2 (2.33) | 1.81 | |||||||||
| 2023 | 1 | 3 | 2 | NH | 108 | 21 (19.44) | 64 (59.26) | 18 (16.67) | 3 (2.78) | 2 (1.85) | 2.08 | ||||
| HN | 68 | 15 (22.06) | 40 (58.82) | 11 (16.18) | 1 (1.47) | 1 (1.47) | 2.01 | ||||||||
| Color around the stone | FC (stone) | 2022 | 1 | 3 | 2 | NH | 98 | 26 (26.53) | 21 (21.43) | 28 (28.57) | 23 (23.47) | 2.49 | / | ||
| HN | 83 | 34 (40.96) | 16 (19.28) | 18 (21.69) | 15 (18.07) | 2.17 | |||||||||
| Fruit flavor (sour/sweet) | FF | 2021 | 3 | 2 | 2.5 | NH | 107 | 7 (6.54) | 47 (43.93) | 24 (22.43) | 29 (27.10) | 2.7 | / | ||
| HN | 78 | 3 (3.85) | 42 (53.85) | 20 (25.64) | 13 (16.67) | 2.55 | |||||||||
| 2022 | 3 | 2 | 2.5 | NH | 112 | 2 (1.79) | 73 (65.18) | 18 (16.07) | 19 (16.96) | 2.48 | |||||
| HN | 86 | 1 (1.16) | 42 (48.84) | 24 (27.91) | 19 (22.09) | 2.71 | |||||||||
| Bitter taste | Bitter | 2021 | 2 | 1 | 0.5 | NH | 97 | 72 (74.23) | 15 (15.46) | 10 (10.31) | 1.36 | NH = 0.98; HN = 0.95 | |||
| HN | 73 | 52 (71.23) | 16 (21.92) | 5 (6.85) | 1.36 | ||||||||||
| 2022 | 2 | 1 | 0.5 | NH | 112 | 76 (67.86) | 21 (18.75) | 15 (13.39) | 1.46 | ||||||
| HN | 85 | 53 (62.35) | 19 (22.35) | 13 (15.29) | 1.53 | ||||||||||
| 2023 | 2 | 1 | 0.5 | NH | 97 | 73 (75.26) | 15 (15.46) | 9 (9.28) | 1.34 | ||||||
| HN | 71 | 50 (70.42) | 16 (22.54) | 5 (7.04) | 1.37 | ||||||||||
| Fruit shape | FSh | 2021 | 1 | 5 | 3 | NH | 105 | 44 (41.90) | 4 (3.81) | 18 (17.14) | 16 (15.24) | 23 (21.90) | 2.71 | NH = 0.89; HN = 0.88 | |
| HN | 76 | 32 (42.11) | 2 (2.63) | 17 (22.37) | 5 (6.58) | 20 (26.32) | 2.72 | ||||||||
| 2022 | 1 | 5 | 3 | NH | 114 | 45 (39.47) | 8 (7.02) | 22 (19.30) | 15 (13.16) | 24 (21.05) | 2.69 | ||||
| HN | 87 | 44 (50.57) | 2 (2.30) | 16 (18.39) | 1 (1.15) | 24 (27.59) | 2.53 | ||||||||
| 2023 | 1 | 5 | 3 | NH | 108 | 31 (28.70) | 10 (9.26) | 29 (26.85) | 23 (21.30) | 15 (13.89) | 2.82 | ||||
| HN | 68 | 15 (22.06) | 3 (4.41) | 21 (30.88) | 16 (23.53) | 13 (19.12) | 3.13 | ||||||||
| Fruit top shape | FSh (top) | 2021 | 4 | 2 | 3 | NH | 106 | 3 (2.83) | 25 (23.58) | 43 (40.57) | 35 (33.02) | 3.04 | NH = 0.92; HN = 0.84 | ||
| HN | 76 | 3 (3.95) | 31 (40.79) | 29 (38.16) | 13 (17.11) | 2.68 | |||||||||
| 2022 | 4 | 2 | 3 | NH | 114 | 5 (4.39) | 23 (20.18) | 44 (38.60) | 42 (36.84) | 3.08 | |||||
| HN | 87 | 1 (1.15) | 30 (34.48) | 25 (28.74) | 31 (35.63) | 2.99 | |||||||||
| 2023 | 4 | 2 | 3 | NH | 108 | 4 (3.70) | 20 (18.52) | 56 (51.85) | 28 (25.93) | 3.00 | |||||
| HN | 68 | 3 (4.41) | 11 (16.18) | 32 (47.06) | 22 (32.35) | 3.07 | |||||||||
| Hairy stalk | HS | 2021 | 2 | 1 | 1.5 | NH | 107 | 7 (6.54) | 13 (12.15) | 87 (81.31) | 2.75 | NH = 0.84; HN = 0.94 | |||
| HN | 78 | 13 (16.67) | 13 (16.67) | 52 (66.67) | 2.5 | ||||||||||
| 2022 | 2 | 1 | 1.5 | NH | 114 | 4 (3.51) | 15 (13.16) | 95 (83.33) | 2.8 | ||||||
| HN | 87 | 12 (13.79) | 16 (18.39) | 59 (67.82) | 2.54 | ||||||||||
| 2023 | 2 | 1 | 1.5 | NH | 113 | 17 (15.04) | 25 (22.12) | 71 (62.83) | 2.48 | ||||||
| HN | 86 | 18 (20.93) | 17 (19.77) | 51 (59.30) | 2.38 | ||||||||||
| Flower bud color | FlC (bud) | 2021 | 2 | 2 | 2 | NH | 108 | 1 (0.93) | 39 (36.11) | 68 (62.96) | 2.62 | / | |||
| HN | 75 | 2 (2.67) | 37 (49.33) | 36 (48.00) | 2.45 | ||||||||||
| Flower petal color | FlC (petal) | 2021 | 1 | 2 | 1.5 | NH | 108 | 37 (34.26) | 49 (45.37) | 22 (20.37) | 1.86 | / | |||
| HN | 75 | 26 (34.67) | 37 (49.33) | 12 (16.00) | 1.81 | ||||||||||
| Category/Trait | Year | NZH | HF | MP | Cross | Mean | Hm | RHm/% | Hb | RHb/% | HH/% | HM/% | LL/% | H2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fruit size | ||||||||||||||
| FloD/mm | 2021 | 17.36 | 21.88 | 19.62 | NH | 19.43 | −0.19 | −0.97 | −2.45 | −11.20 | 10.19 | 48.15 | 12.96 | NH = 0.84; HN = 0.82 |
| HN | 18.99 | −0.63 | −3.21 | −2.89 | −13.20 | 3.90 | 28.57 | 11.69 | ||||||
| 2022 | 16.37 | 20.23 | 18.30 | NH | 17.43 | −0.87 | −4.73 | −2.80 | −13.82 | 2.63 | 30.70 | 22.81 | ||
| HN | 17.41 | −0.89 | −4.84 | −2.82 | −13.92 | 4.60 | 20.69 | 20.69 | ||||||
| 2023 | 17.12 | 20.39 | 18.76 | NH | 17.77 | −0.99 | −5.26 | −2.62 | −12.85 | 8.33 | 25.93 | 37.96 | ||
| HN | 18.23 | −0.52 | −2.78 | −2.16 | −10.58 | 7.35 | 35.29 | 17.65 | ||||||
| FTraD/mm | 2021 | 18.63 | 21.45 | 20.04 | NH | 19.86 | −0.18 | −0.91 | −1.59 | −7.42 | 14.81 | 47.22 | 21.30 | NH = 0.76; HN = 0.78 |
| HN | 18.36 | −1.68 | −8.37 | −3.09 | −14.40 | 7.79 | 36.36 | 25.97 | ||||||
| 2022 | 17.29 | 19.83 | 18.56 | NH | 17.87 | −0.69 | −3.73 | −1.96 | −9.90 | 7.02 | 24.56 | 30.70 | ||
| HN | 18.12 | −0.44 | −2.36 | −1.71 | −8.61 | 10.34 | 36.78 | 24.14 | ||||||
| 2023 | 18.57 | 20.60 | 19.59 | NH | 18.54 | −1.05 | −5.34 | −2.06 | −10.01 | 10.19 | 24.07 | 48.15 | ||
| HN | 19.20 | −0.39 | −1.98 | −1.40 | −6.81 | 10.29 | 39.71 | 26.47 | ||||||
| FLaD/mm | 2021 | 17.03 | 18.77 | 17.90 | NH | 17.30 | −0.60 | −3.35 | −1.47 | −7.83 | 12.96 | 30.56 | 44.44 | NH = 0.74; HN = 0.76 |
| HN | 16.86 | −1.04 | −5.79 | −1.91 | −10.16 | 10.39 | 29.87 | 46.75 | ||||||
| 2022 | 15.34 | 17.57 | 16.46 | NH | 15.70 | −0.75 | −4.58 | −1.87 | −10.64 | 7.02 | 14.91 | 38.60 | ||
| HN | 15.92 | −0.53 | −3.24 | −1.65 | −9.38 | 6.90 | 28.74 | 26.44 | ||||||
| 2023 | 16.67 | 18.23 | 17.45 | NH | 16.44 | −1.01 | −5.78 | −1.79 | −9.81 | 10.19 | 32.41 | 56.48 | ||
| HN | 17.04 | −0.41 | −2.36 | −1.19 | −6.54 | 13.24 | 39.71 | 33.82 | ||||||
| FW/g | 2021 | 3.81 | 5.35 | 4.58 | NH | 4.35 | −0.23 | −4.93 | −1.00 | −18.62 | 12.15 | 34.58 | 29.91 | NH = 0.80; HN = 0.76 |
| HN | 4.06 | −0.52 | −11.45 | −1.29 | −24.19 | 3.90 | 28.57 | 40.26 | ||||||
| 2022 | 3.2 | 4.25 | 3.73 | NH | 3.29 | −0.44 | −11.68 | −0.96 | −22.59 | 7.89 | 19.30 | 48.25 | ||
| HN | 3.35 | −0.38 | −10.13 | −0.90 | −21.23 | 8.05 | 24.14 | 40.23 | ||||||
| 2023 | 3.69 | 4.95 | 4.32 | NH | 3.73 | −0.59 | −13.67 | −1.22 | −24.65 | 7.34 | 23.85 | 26.61 | ||
| HN | 3.88 | −0.44 | −10.08 | −1.07 | −21.53 | 5.88 | 25.00 | 36.76 | ||||||
| FW (stone)/g | 2021 | 0.2 | 0.29 | 0.25 | NH | 0.20 | −0.05 | −19.37 | −0.09 | −31.88 | 0.98 | 11.76 | 52.94 | / |
| HN | 0.20 | −0.04 | −16.52 | −0.09 | −29.48 | 4.11 | 10.96 | 43.84 | ||||||
| Fruit color | ||||||||||||||
| An/mg·kg−1 | 2021 | 6.96 | 42.02 | 24.49 | NH | 24.42 | −0.07 | −0.30 | −17.60 | −41.89 | 13.64 | 31.82 | 10.61 | / |
| HN | 22.27 | −2.22 | −9.07 | −19.75 | −47.00 | 11.43 | 31.43 | 14.29 | ||||||
| a | 2022 | 25.51 | 19.74 | 22.63 | NH | 22.52 | −0.10 | −0.46 | −2.99 | −11.72 | 24.56 | 62.28 | 21.05 | / |
| HN | 22.89 | 0.27 | 1.19 | −2.62 | −10.26 | 22.99 | 62.07 | 14.94 | ||||||
| b | 2022 | 13.89 | 7.34 | 10.62 | NH | 10.41 | −0.21 | −1.93 | −3.48 | −25.05 | 12.28 | 52.63 | 18.42 | / |
| HN | 10.25 | −0.36 | −3.41 | −3.64 | −26.18 | 3.45 | 54.02 | 16.09 | ||||||
| L | 2022 | 40.43 | 30.74 | 35.59 | NH | 33.53 | −2.06 | −5.79 | −6.90 | −17.08 | 5.26 | 25.44 | 21.93 | / |
| HN | 33.62 | −1.96 | −5.51 | −6.81 | −16.83 | 2.30 | 31.03 | 20.69 | ||||||
| Fruit flavor | ||||||||||||||
| TSS/% | 2021 | 11.36 | 12.97 | 12.17 | NH | 14.47 | 2.31 | 18.97 | 1.50 | 11.59 | 76.64 | 89.72 | 6.54 | NH = 0.62; HN = 0.55 |
| HN | 14.52 | 2.35 | 19.35 | 1.55 | 11.94 | 76.92 | 96.15 | 3.85 | ||||||
| 2022 | 15.3 | 16.77 | 16.04 | NH | 17.39 | 1.36 | 8.47 | 0.62 | 3.72 | 54.87 | 73.45 | 17.70 | ||
| HN | 17.29 | 1.25 | 7.82 | 0.52 | 3.09 | 50.57 | 72.41 | 12.64 | ||||||
| 2023 | 13.65 | 15.95 | 14.80 | NH | 16.35 | 1.55 | 10.46 | 0.40 | 2.49 | 51.38 | 67.89 | 13.76 | ||
| HN | 15.84 | 1.04 | 7.00 | −0.11 | −0.71 | 38.24 | 67.65 | 11.76 | ||||||
| TA/% | 2021 | 0.67 | 0.58 | 0.63 | NH | 0.60 | −0.02 | −3.22 | −0.07 | −9.72 | 30.23 | 37.21 | 44.19 | / |
| HN | 0.59 | −0.03 | −4.89 | −0.08 | −11.28 | 18.52 | 48.15 | 37.04 | ||||||
| 2022 | 0.74 | 0.54 | 0.64 | NH | 0.54 | −0.10 | −16.08 | −0.20 | −27.42 | 5.31 | 15.04 | 53.10 | ||
| HN | 0.55 | −0.09 | −14.19 | −0.19 | −25.78 | 6.90 | 22.99 | 44.83 | ||||||
| 2023 | 0.64 | 0.45 | 0.55 | NH | 0.54 | −0.01 | −1.05 | −0.10 | −15.74 | 19.27 | 44.95 | 25.69 | ||
| HN | 0.49 | −0.06 | −10.55 | −0.15 | −23.83 | 4.41 | 22.06 | 32.35 | ||||||
| TSS/TA | 2021 | 16.96 | 22.36 | 19.66 | NH | 26.36 | 6.70 | 34.09 | 4.00 | 17.88 | 58.14 | 69.77 | 16.28 | / |
| HN | 24.47 | 4.81 | 24.47 | 2.11 | 9.43 | 53.85 | 76.92 | 15.38 | ||||||
| 2022 | 20.81 | 31.25 | 26.03 | NH | 33.50 | 7.47 | 28.69 | 2.25 | 7.20 | 61.95 | 91.15 | 1.77 | ||
| HN | 33.05 | 7.02 | 26.98 | 1.80 | 5.77 | 54.02 | 82.76 | 0.00 | ||||||
| 2023 | 21.18 | 35.42 | 28.30 | NH | 31.78 | 3.48 | 12.30 | −3.64 | −10.28 | 29.36 | 66.06 | 3.67 | ||
| HN | 33.71 | 5.41 | 19.13 | −1.71 | −4.82 | 28.57 | 78.57 | 0.00 | ||||||
| pH | 2021 | 4.01 | 3.94 | 3.98 | NH | 4.00 | 0.02 | 0.53 | −0.01 | −0.35 | 48.84 | 48.84 | 41.86 | / |
| HN | 4.00 | 0.02 | 0.60 | −0.01 | −0.28 | 29.63 | 29.63 | 48.15 | ||||||
| SS/g·kg− 1 | 2021 | 92.35 | 117.27 | 104.81 | NH | 116.44 | 11.63 | 11.09 | −0.83 | −0.71 | 43.94 | 65.15 | 13.64 | / |
| HN | 123.12 | 18.31 | 17.47 | 5.85 | 4.99 | 51.43 | 71.43 | 17.14 | ||||||
| Fruit shape | ||||||||||||||
| FSh (index) | 2021 | 0.87 | 1.02 | 0.95 | NH | 0.98 | 0.04 | 3.74 | −0.04 | −3.89 | 23.15 | 67.59 | 0.00 | NH = 0.82; HN = 0.72 |
| HN | 0.98 | 0.03 | 3.25 | −0.04 | −4.34 | 14.29 | 63.64 | 0.00 | ||||||
| 2022 | 0.85 | 1.02 | 0.94 | NH | 0.98 | 0.04 | 4.53 | −0.04 | −4.18 | 19.30 | 71.93 | 0.88 | ||
| HN | 0.96 | 0.03 | 2.88 | −0.06 | −5.69 | 9.20 | 65.52 | 1.15 | ||||||
| 2023 | 0.88 | 0.99 | 0.93 | NH | 0.96 | 0.03 | 2.75 | −0.03 | −3.17 | 24.07 | 66.67 | 3.70 | ||
| HN | 0.95 | 0.02 | 1.85 | −0.04 | −4.02 | 13.24 | 58.82 | 2.94 | ||||||
| FSL/mm | 2021 | 19.88 | 21.40 | 20.64 | NH | 20.25 | −0.39 | −1.90 | −1.15 | −5.38 | 34.26 | 44.44 | 46.30 | NH = 0.88; HN = 0.87 |
| HN | 19.38 | −1.26 | −6.10 | −2.02 | −9.43 | 30.26 | 32.89 | 59.21 | ||||||
| 2022 | 20.63 | 26.51 | 23.57 | NH | 22.62 | −0.95 | −4.05 | −3.89 | −14.69 | 15.79 | 36.84 | 35.09 | ||
| HN | 21.95 | −1.62 | −6.88 | −4.56 | −17.21 | 13.79 | 34.48 | 41.38 | ||||||
| 2023 | 24.04 | 24.94 | 24.49 | NH | 23.08 | −1.41 | −5.77 | −1.86 | −7.47 | 25.93 | 30.56 | 53.70 | ||
| HN | 22.84 | −1.65 | −6.75 | −2.10 | −8.43 | 27.94 | 30.88 | 63.24 | ||||||
| Phenology (flower and fruit) | ||||||||||||||
| FD (beginning) /d | 2021 | 43 | 42 | 42.5 | NH | 41.69 | −0.81 | −1.91 | −1.31 | −3.05 | 20.18 | 29.36 | 62.39 | NH = 0.76; HN = 0.78 |
| HN | 41.47 | −1.03 | −2.43 | −1.53 | −3.56 | 18.18 | 23.38 | 59.74 | ||||||
| 2022 | 55 | 55 | 55.0 | NH | 52.94 | −2.06 | −3.75 | −2.06 | −3.75 | 16.49 | 16.49 | 72.16 | ||
| HN | 52.30 | −2.70 | −4.91 | −2.70 | −4.91 | 18.57 | 18.57 | 74.29 | ||||||
| 2023 | 52 | 52 | 52.0 | NH | 50.44 | −1.56 | −3.00 | −1.56 | −3.00 | 38.05 | 38.05 | 56.64 | ||
| HN | 50.03 | −1.97 | −3.80 | −1.97 | −3.80 | 34.21 | 34.21 | 63.16 | ||||||
| FD (full)/d | 2021 | 44 | 43 | 43.5 | NH | 43.31 | −0.19 | −0.43 | −0.69 | −1.56 | 31.19 | 34.86 | 48.62 | NH = 0.77; HN = 0.79 |
| HN | 42.86 | −0.64 | −1.48 | −1.14 | −2.60 | 19.74 | 34.21 | 60.53 | ||||||
| 2022 | 57 | 57 | 57.0 | NH | 55.68 | −1.32 | −2.32 | −1.32 | −2.32 | 17.65 | 17.65 | 70.59 | ||
| HN | 55.80 | −1.20 | −2.11 | −1.20 | −2.11 | 27.50 | 27.50 | 61.25 | ||||||
| 2023 | 53 | 54 | 53.5 | NH | 52.59 | −0.91 | −1.70 | −1.41 | −2.61 | 41.59 | 45.13 | 44.25 | ||
| HN | 52.07 | −1.43 | −2.68 | −1.93 | −3.58 | 31.08 | 35.14 | 60.81 | ||||||
| FD (end)/d | 2021 | 49 | 50 | 49.5 | NH | 49.60 | 0.10 | 0.19 | −0.40 | −0.81 | 27.52 | 43.12 | 29.36 | NH = 0.71; HN = 0.68 |
| HN | 48.93 | −0.57 | −1.14 | −1.07 | −2.13 | 17.11 | 23.68 | 39.47 | ||||||
| 2022 | 62 | 63 | 62.5 | NH | 62.88 | 0.38 | 0.61 | −0.12 | −0.18 | 43.69 | 50.49 | 11.65 | ||
| HN | 62.36 | −0.14 | −0.22 | −0.64 | −1.01 | 34.94 | 34.94 | 21.69 | ||||||
| 2023 | 60 | 62 | 61.0 | NH | 61.73 | 0.73 | 1.20 | −0.27 | −0.43 | 66.37 | 67.26 | 32.74 | ||
| HN | 60.49 | −0.51 | −0.83 | −1.51 | −2.43 | 53.73 | 53.73 | 41.79 | ||||||
| MD/d | 2021 | 93 | 97 | 95.0 | NH | 95.72 | 0.72 | 0.76 | −1.28 | −1.32 | 37.04 | 50.00 | 34.26 | NH = 0.74; HN = 0.77 |
| HN | 93.85 | −1.15 | −1.21 | −3.15 | −3.25 | 20.51 | 39.74 | 50.00 | ||||||
| 2022 | 102 | 108 | 105.0 | NH | 103.28 | −1.72 | −1.64 | −4.72 | −4.37 | 13.16 | 37.72 | 43.86 | ||
| HN | 103.16 | −1.84 | −1.75 | −4.84 | −4.48 | 13.79 | 29.89 | 40.23 | ||||||
| 2023 | 103 | 109 | 106.0 | NH | 104.00 | −2.00 | −1.89 | −5.00 | −4.59 | 12.84 | 19.27 | 45.87 | ||
| HN | 103.65 | −2.35 | −2.22 | −5.35 | −4.91 | 4.23 | 8.45 | 36.62 | ||||||
| FDP/d | 2021 | 49 | 54 | 51.5 | NH | 52.41 | 0.91 | 1.76 | −1.59 | −2.95 | 31.48 | 57.41 | 22.22 | NH = 0.74; HN = 0.75 |
| HN | 50.97 | −0.53 | −1.02 | −3.03 | −5.60 | 23.68 | 42.11 | 34.21 | ||||||
| 2022 | 45 | 51 | 48.0 | NH | 47.25 | −0.75 | −1.55 | −3.75 | −7.34 | 18.63 | 37.25 | 34.31 | ||
| HN | 47.25 | −0.75 | −1.56 | −3.75 | −7.35 | 1.25 | 38.75 | 33.75 | ||||||
| 2023 | 50 | 55 | 52.5 | NH | 51.56 | −0.94 | −1.79 | −3.44 | −6.26 | 20.18 | 42.20 | 33.03 | ||
| HN | 51.82 | −0.68 | −1.29 | −3.18 | −5.78 | 14.93 | 44.78 | 25.37 | ||||||
| Flower | ||||||||||||||
| FlD/mm | 2021 | 25.80 | 34.42 | 30.11 | NH | 28.31 | −1.80 | −5.96 | −6.11 | −17.74 | 1.05 | 21.05 | 21.05 | / |
| HN | 27.34 | −2.77 | −9.21 | −7.08 | −20.58 | 3.03 | 12.12 | 25.76 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Yang, S.; Hao, L.; Wang, H.; Zhang, J.; He, W.; Li, M.; Lin, Y.; Zhang, Y.; Chen, Q.; et al. Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry. Agronomy 2024, 14, 2862. https://doi.org/10.3390/agronomy14122862
Liu Z, Yang S, Hao L, Wang H, Zhang J, He W, Li M, Lin Y, Zhang Y, Chen Q, et al. Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry. Agronomy. 2024; 14(12):2862. https://doi.org/10.3390/agronomy14122862
Chicago/Turabian StyleLiu, Zhenshan, Shuaiwei Yang, Lisu Hao, Hao Wang, Jing Zhang, Wen He, Mengyao Li, Yuanxiu Lin, Yunting Zhang, Qing Chen, and et al. 2024. "Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry" Agronomy 14, no. 12: 2862. https://doi.org/10.3390/agronomy14122862
APA StyleLiu, Z., Yang, S., Hao, L., Wang, H., Zhang, J., He, W., Li, M., Lin, Y., Zhang, Y., Chen, Q., Zhang, Y., Luo, Y., Tang, H., Wang, Y., & Wang, X. (2024). Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry. Agronomy, 14(12), 2862. https://doi.org/10.3390/agronomy14122862