Appropriate Planting Density Can Improve the Storage Root Yield and Commercial Features of Sweet Potato (Ipomoea batatas L.) by Optimizing the Photosynthetic Performance


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Sampling Method
2.4. Physiological Analysis
2.4.1. Functional Leaf SPAD Value and Leaf Nitrogen Content
2.4.2. Photosynthetic Characteristic
2.4.3. Chlorophyll Fluorescence Measurements
2.4.4. Canopy Photosynthetic Measurement
2.4.5. Agronomic Traits
2.4.6. Storage Root Yield, Yield Component, and Commercial Traits
2.4.7. Storage Root Carbohydrate Content
2.4.8. Real-Time Quantitative PCR Analysis
2.5. Statistical Analysis
3. Results
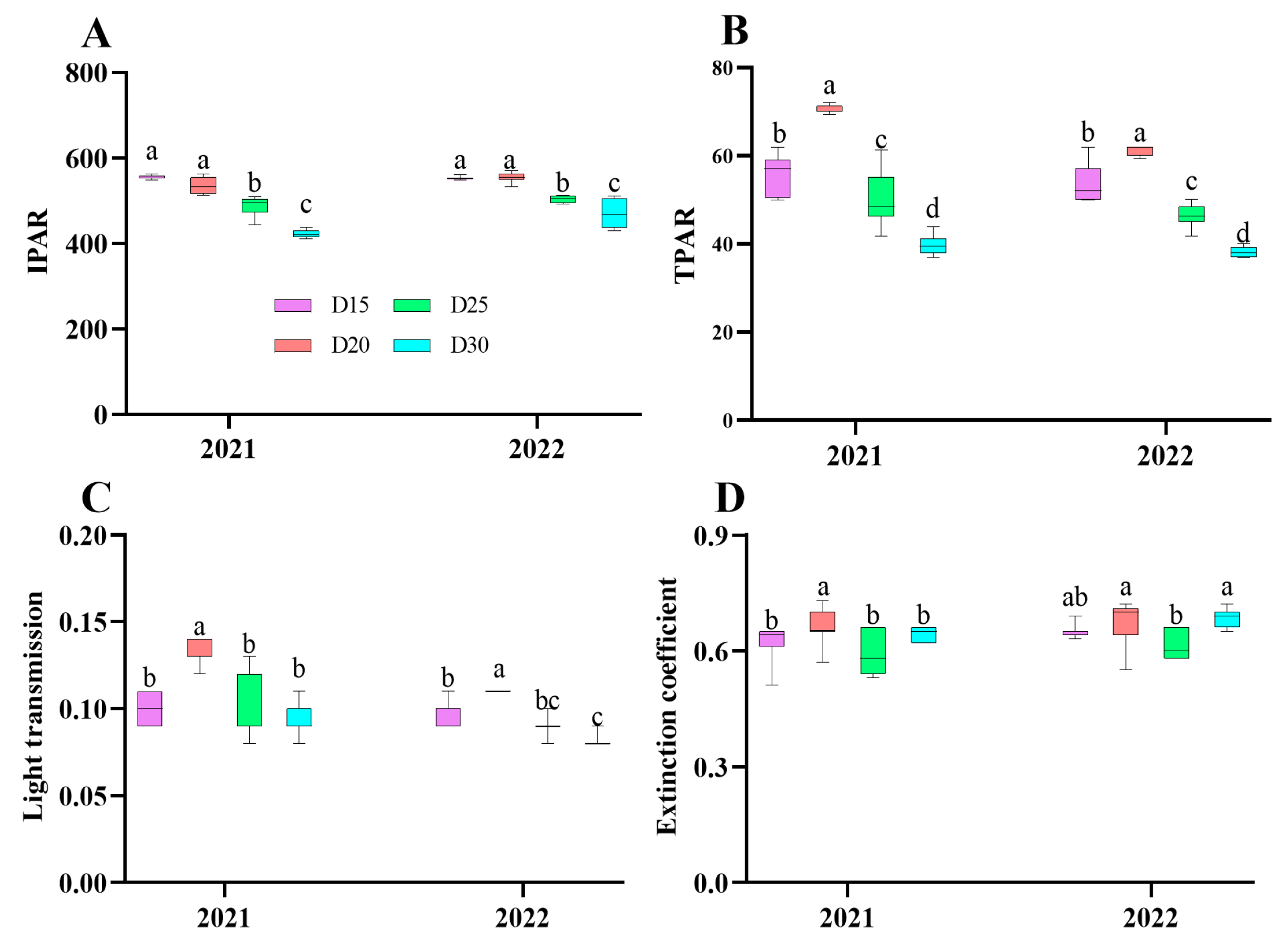
3.1. Canopy Spectrum Parameter
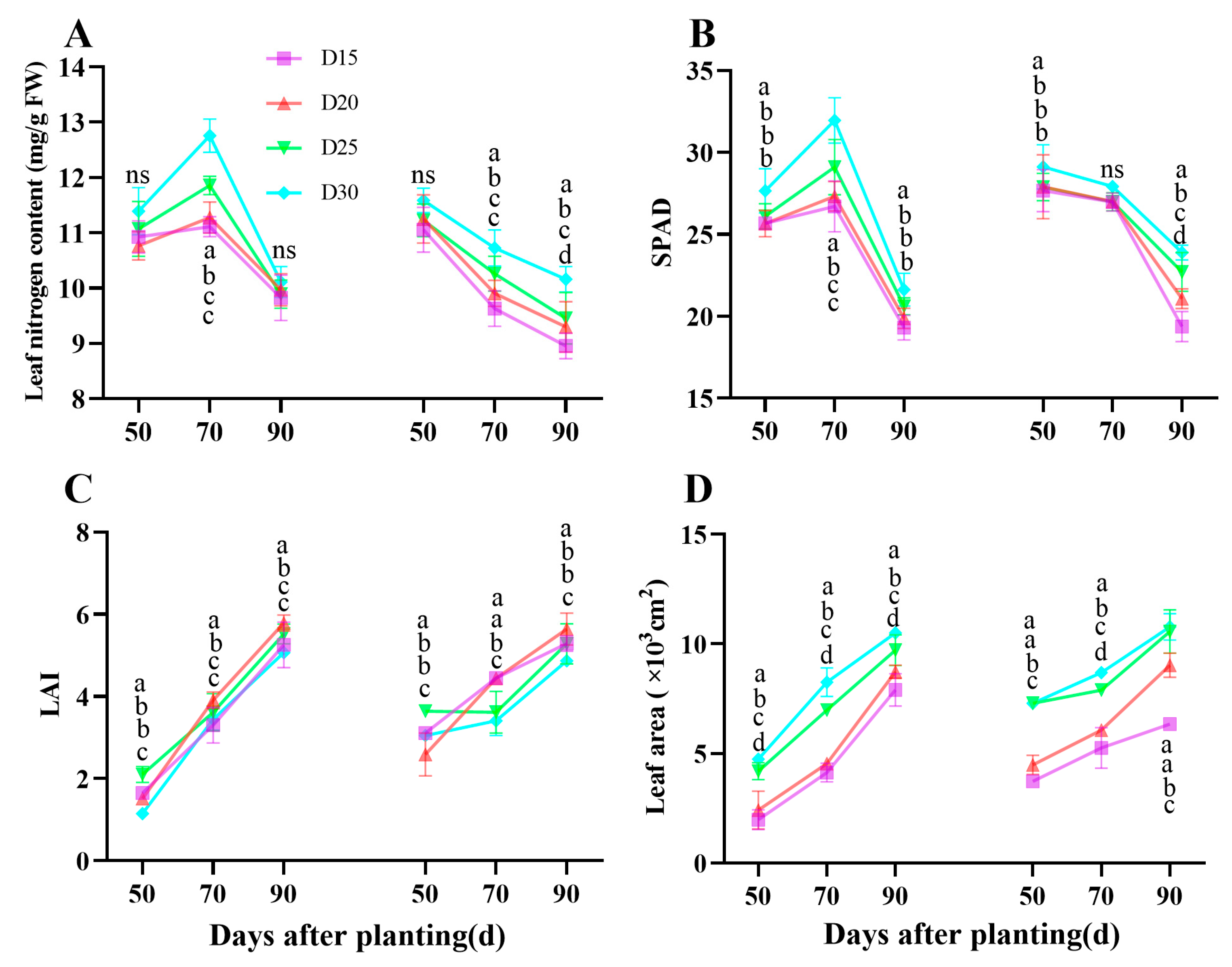
3.2. Leaf Nitrogen Content, Chlorophyll Content, LAI, and LA Characteristics During Bulking Period
3.3. Leaf Gas Exchange
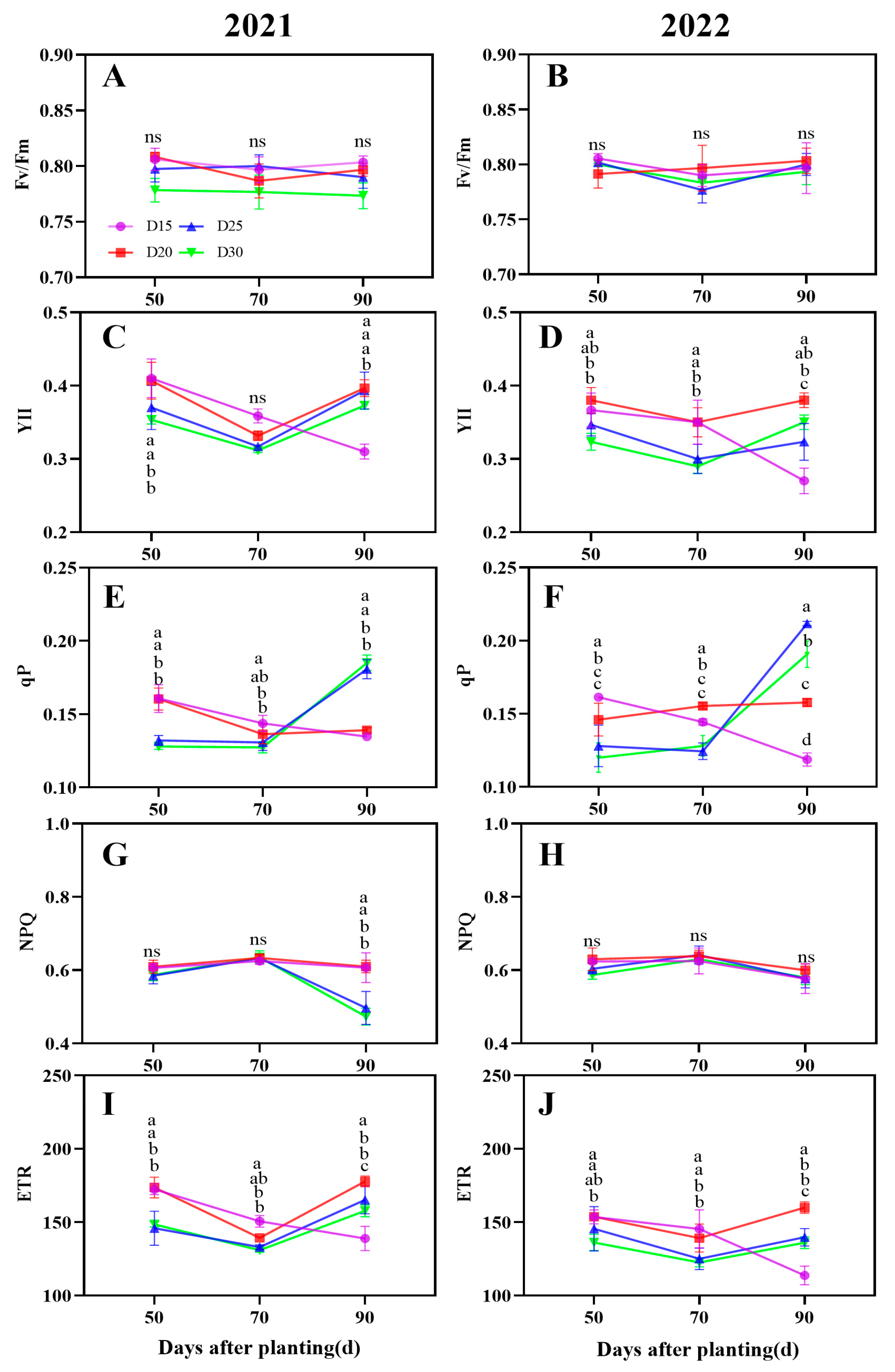
3.4. Chlorophyll Fluorescence Parameters
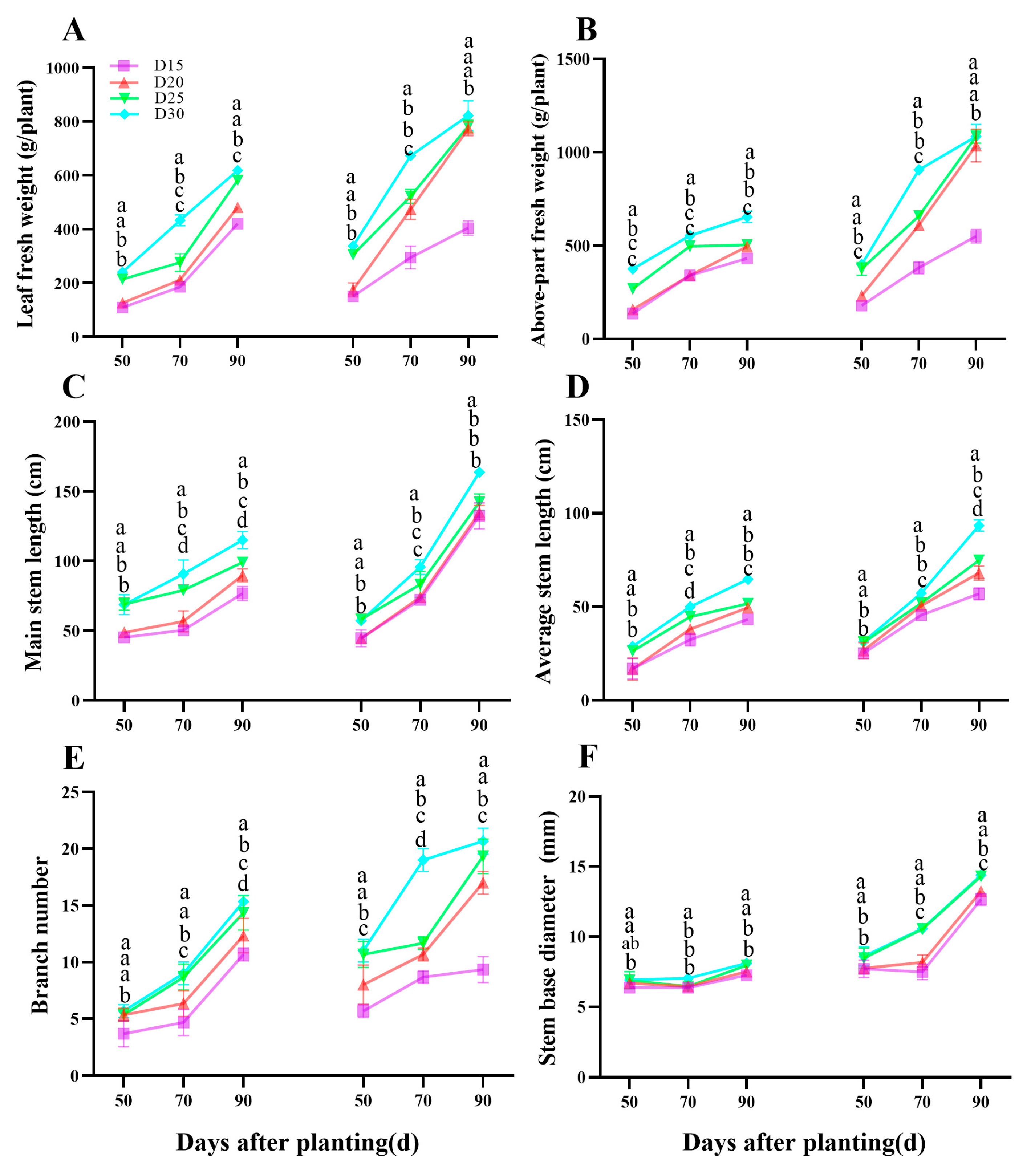
3.5. Above-Part Development
3.6. Dry Matter Accumulation and Allocation
3.7. Genes Expression Level on Regulating Carbohydrates Metabolism in Storage Root
3.8. Carbohydrate Content on Storage Root During Bulking Period
3.9. Storage Root Development Characteristics During Bulking Period
3.10. Storage Root Characteristics at Canopy Closure Period
3.11. Storage Root Yield, Yield Component, and Appearance Quality at Harvest Period
3.12. Storage Root Commercial Characteristics at Harvest Time
3.13. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kumar, S.; Wang, S.; Wang, M.; Zeb, S.; Khan, M.N.; Chen, Y.; Zhu, G.; Zhu, Z. Enhancement of sweetpotato tolerance to chromium stress through melatonin and glutathione: Insights into photosynthetic efficiency, oxidative defense, and growth parameters. Plant Physiol. Biochem. 2024, 208, 108509. [Google Scholar] [CrossRef] [PubMed]
- Buettner, D. The Blue Zones Solution: Eating and Living Like the World’s Healthiest People; National Geographic Books: Washington, DC, USA, 2005. [Google Scholar]
- Tairo, F.; Mukasa, S.B.; Jones, R.A.C.; Kullaya, A.; Rubaihayo, P.R.; Valkonen, J.P.T. Unravelling the genetic diversity of the three main viruses involved in Sweet Potato Virus Disease (SPVD), and its practical implications. Mol. Plant Pathol. 2005, 6, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Ebem, E.C.; Afuape, S.O.; Chukwu, S.C.; Ubi, B.E. Genotype × Environment Interaction and Stability Analysis for Root Yield in Sweet Potato [Ipomoea batatas (L.) Lam]. Front. Agron. 2021, 3, 665564. [Google Scholar] [CrossRef]
- Liang, Q.; Chen, H.; Chang, H.; Liu, Y.; Wang, Q.; Wu, J.; Liu, Y.; Kumar, S.; Chen, Y.; Chen, Y.; et al. Influence of Planting Density on Sweet Potato Storage Root Formation by Regulating Carbohydrate and Lignin Metabolism. Plants 2023, 12, 2039. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Fan, J.; Yang, R.; Xu, X.; Liu, L.; Li, S.; Zhang, F.; Li, Z. Interactive effects of plant density and nitrogen rate on grain yield, economic benefit, water productivity and nitrogen use efficiency of drip-fertigated maize in northwest China. Agric. Water Manag. 2022, 263, 107453. [Google Scholar] [CrossRef]
- Postma, J.A.; Hecht, V.L.; Hikosaka, K.; Nord, E.A.; Pons, T.L.; Poorter, H. Dividing the pie: A quantitative review on plant density responses. Plant Cell Environ. 2020, 44, 1072–1094. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Ma, L.; Lv, X.; Meng, Y.; Zhou, Z. Straw returning coupled with nitrogen fertilization increases canopy photosynthetic capacity, yield and nitrogen use efficiency in cotton. Eur. J. Agron. 2021, 126, 126267. [Google Scholar] [CrossRef]
- Cao, Y.-J.; Wang, L.-C.; Gu, W.-R.; Wang, Y.-J.; Zhang, J.-H. Increasing photosynthetic performance and post-silking N uptake by moderate decreasing leaf source of maize under high planting density. J. Integr. Agric. 2021, 20, 494–510. [Google Scholar] [CrossRef]
- Liu, T.N.; Wang, Z.L.; Cai, T. Canopy apparent photosynthetic characteristics and yield of two spike-type wheat cultivars in response to row spacing under high plant density. PLoS ONE 2016, 11, e0148582. [Google Scholar] [CrossRef]
- Mussgnug, J.H.; Thomas-Hall, S.; Rupprecht, J.; Foo, A.; Klassen, V.; McDowall, A.; Schenk, P.M.; Kruse, O.; Hankamer, B. Engineering photosynthetic light capture: Impacts on improved solar energy to biomass conversion. Plant Biotechnol. J. 2007, 5, 802–814. [Google Scholar] [CrossRef]
- Singh, S.K.; Badgujar, G.; Reddy, V.R.; Fleisher, D.H.; Bunce, J.A. Carbon dioxide diffusion across stomata and mesophyll and photo-biochemical processes as affected by growth CO2 and phosphorus nutrition in cotton. J. Plant Physiol. 2013, 170, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Rascher, U.; Bobich, E.G.; Lin, G.H.; Walter, A.; Morris, T.; Naumann, M.; Nichol, C.J.; Pierce, D.; Bil, K.; Kudeyarov, V.; et al. Functional diversity of photosynthesis during drought in a model tropical rainforest–the contributions of leaf area, photo-synthetic electron transport and stomatal conductance to reduction in net ecosystem carbon exchange. Plant Cell Environ. 2004, 27, 1239–1256. [Google Scholar] [CrossRef]
- Long, J.-R.; Ma, G.-H.; Wan, Y.-Z.; Song, C.-F.; Sun, J.; Qin, R.-J. Effects of Nitrogen Fertilizer Level on Chlorophyll Fluorescence Characteristics in Flag Leaf of Super Hybrid Rice at Late Growth Stage. Rice Sci. 2013, 20, 220–228. [Google Scholar] [CrossRef]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.; Sanglard, L.M.; Reis, J.V.; Detmann, E.; Rodrigues, F.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon nutrition increases grain yield, which, in turn, exerts a feed-forward stimulation of photosynthetic rates via enhanced mesophyll conductance and alters primary metabolism in rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef]
- Chen, M.; Liang, F.; Yan, Y.; Wang, Y.; Zhang, Y.; Tian, J.; Jiang, C.; Zhang, W. Boll-leaf system gas exchange and its application in the analysis of cotton photosynthetic function. Photosynth. Res. 2021, 150, 251–262. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Oleksyn, J.; Reich, P.B. Seedlings of five boreal tree species differ in acclimation of net photosynthesis to elevated CO2 and temperature. Tree Physiol. 1998, 18, 715–726. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. Forest growth and species distribution in a changing climate. Tree Physiol. 2000, 20, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Chakrabarti, S.; Makeshkumar, T.; Saravanan, R. Molecular regulation of storage root formation and development in sweet potato. Hortic. Rev. 2014, 42, 157–208. [Google Scholar]
- Liu, X.; Peng, Y.; Yang, Q.; Wang, X.; Cui, N. Determining optimal deficit irrigation and fertilization to increase mango yield, quality, and WUE in a dry hot environment based on TOPSIS. Agric. Water Manag. 2020, 245, 106650. [Google Scholar] [CrossRef]
- Rascher, U.; Liebig, M.; Lüttge, U. Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the field. Plant Cell Environ. 2000, 23, 1397–1405. [Google Scholar] [CrossRef]
- Dong, B.; Wang, Z.; Evers, J.B.; Stomph, T.J.; van der Putten, P.E.; Yin, X.; Wang, J.L.; Sprangers, T.; Hang, X.; van der Werf, W. Competition for light and nitrogen with an earlier-sown species negatively affects leaf traits and leaf photosynthetic capacity of maize in relay intercropping. Eur. J. Agron. 2024, 155, 127119. [Google Scholar] [CrossRef]
- Villordon, A.; LaBonte, D.; Firon, N.; Carey, E. Variation in Nitrogen Rate and Local Availability Alter Root Architecture Attributes at the Onset of Storage Root Initiation in ‘Beauregard’ Sweetpotato. HortScience 2013, 48, 808–815. [Google Scholar] [CrossRef]
- Si, C.-C.; Liang, Q.-G.; Liu, H.-J.; Wang, N.; Kumar, S.; Chen, Y.-L.; Zhu, G.-P. Response Mechanism of Endogenous Hormones of Potential Storage Root to Phosphorus and Its Relationship with Yield and Appearance Quality of Sweetpotato. Front. Plant Sci. 2022, 13, 872422. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, G.; Liu, G.; Wang, K.; Xie, R.; Hou, P.; Ming, B.; Wang, Z.; Li, S. Improving the yield potential in maize by constructing the ideal plant type and optimizing the maize canopy structure. Food Energy Secur. 2021, 10, e312. [Google Scholar] [CrossRef]
- Liu, T.; Gu, L.; Dong, S.; Zhang, J.; Liu, P.; Zhao, B. Optimum leaf removal increases canopy apparent photosynthesis, 13C-photosynthate distribution and grain yield of maize crops grown at high density. Field Crop. Res. 2015, 170, 32–39. [Google Scholar] [CrossRef]
- Yang, G.-Z.; Luo, X.-J.; Nie, Y.-C.; Zhang, X.-L. Effects of Plant Density on Yield and Canopy Micro Environment in Hybrid Cotton. J. Integr. Agric. 2014, 13, 2154–2163. [Google Scholar] [CrossRef]
- Fan, P.; Anten, N.P.; Evers, J.B.; Li, Y.; Li, S.; Ming, B.; Xie, R. Higher yields of modern maize cultivars are not associated with coordinated light and N distribution within the canopy. Field Crop. Res. 2023, 305, 109182. [Google Scholar] [CrossRef]
- Bashir, N.; Athar, H.U.R.; Kalaji, H.M.; Wróbel, J.; Mahmood, S.; Zafar, Z.U.; Ashraf, M. Is photoprotection of PSII one of the key mechanisms for drought tolerance in maize? Int. J. Mol. Sci. 2021, 22, 13490. [Google Scholar] [CrossRef]
- Peng, J.; Feng, Y.; Wang, X.; Li, J.; Xu, G.; Phonenasay, S.; Luo, Q.; Han, Z.; Lu, W. Effects of nitrogen application rate on the photosynthetic pigment, leaf fluorescence characteristics, and yield of indica hybrid rice and their interrelations. Sci. Rep. 2021, 11, 7485. [Google Scholar] [CrossRef]
- Perera-Castro, A.V.; Flexas, J. The ratio of electron transport to assimilation (ETR/AN): Underutilized but essential for assessing both equipment’s proper performance and plant status. Planta 2023, 257, 29. [Google Scholar] [CrossRef]
- Yang, H.; Chai, Q.; Yin, W.; Hu, F.; Qin, A.; Fan, Z.; Yu, A.; Zhao, C.; Fan, H. Yield photosynthesis and leaf anatomy of maize in inter- and mono-cropping systems at varying plant densities. Crop. J. 2021, 10, 893–903. [Google Scholar] [CrossRef]
- Samedani, B.; Juraimi, A.S.; Anwar, M.P.; Rafii, M.Y.; Sheikh Awadz, S.H.; Anuar, A.R. Competitive interaction of Axonopus compressus and Asystasia gangetica under contrasting sunlight intensity. Sci. World J. 2013, 2013, 308646. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Zheng, J.; Tan, D.K.Y.; Khan, A.; Akhtar, K.; Kong, X.; Munsif, F.; Iqbal, A.; Afridi, M.Z.; Ullah, A.; et al. Changes in Leaf Structural and Functional Characteristics when Changing Planting Density at Different Growth Stages Alters Cotton Lint Yield under a New Planting Model. Agronomy 2019, 9, 859. [Google Scholar] [CrossRef]
- Wu, B.; Zuo, W.; Yang, P.; Zhang, W. Optimal water and nitrogen management increases cotton yield through improving leaf number and canopy light environment. Field Crop. Res. 2023, 290, 108745. [Google Scholar] [CrossRef]
- Guo, Y.; Yin, W.; Fan, H.; Fan, Z.; Hu, F.; Yu, A.; Zhao, C.; Chai, Q.; Aziiba, E.A.; Zhang, X. Photosynthetic Physiological Characteristics of Water and Nitrogen Coupling for Enhanced High-Density Tolerance and Increased Yield of Maize in Arid Irrigation Regions. Front. Plant Sci. 2021, 12, 726568. [Google Scholar] [CrossRef]
- Zheng, B.; Li, Y.; Wu, Q.; Zhao, W.; Ren, T.; Zhang, X.; Li, G.; Ning, T.; Zhang, Z. Maize (Zea mays L.) planted at higher density utilizes dynamic light more efficiently. Plant Cell Environ. 2023, 46, 3305–3322. [Google Scholar] [CrossRef]
- Dai, Y.L.; Fan, J.L.; Liao, Z.Q.; Zhang, C.; Yu, J.; Feng, H.L.; Zhang, C.; Li, Z.J. Supplemental irrigation and modified plant density improved photosynthesis, grain yield and water productivity of winter wheat under ridge-furrow mulching. Agric. Water Manag. 2022, 274, 107985. [Google Scholar] [CrossRef]
- Liao, Z.Q.; Zeng, H.L.; Fan, J.L.; Lai, Z.L.; Zhang, C.; Zhang, F.C.; Wang, H.D.; Cheng, M.H.; Guo, J.J.; Li, Z.J.; et al. Effects of plant density, nitrogen rate and supplemental irrigation on photosynthesis, root growth, seed yield and water-nitrogen use efficiency of soybean under ridge-furrow plastic mulching. Agric. Water Manag. 2022, 268, 107688. [Google Scholar] [CrossRef]
- Xia, F.; Yang, Y.; Zhang, S.; Li, D.; Sun, W.; Xie, Y. Influencing factors of the supply-demand relationships of carbon sequestration and grain provision in China: Does land use matter the most? Sci. Total. Environ. 2022, 832, 154979. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Muñoz, F.J.; Montero, M.; Etxeberria, E.; Sesma, M.T.; Ovecka, M.; Bahaji, A.; Ezquer, I.; Li, J.; Prat, S.; et al. Enhancing Sucrose Synthase Activity in Transgenic Potato (Solanum tuberosum L.) Tubers Results in Increased Levels of Starch, ADPglucose and UDPglucose and Total Yield. Plant Cell Physiol. 2009, 50, 1651–1662. [Google Scholar] [CrossRef]
- Hashida, Y.; Hirose, T.; Okamura, M.; Hibara, K.-I.; Ohsugi, R.; Aoki, N. A reduction of sucrose phosphate synthase (SPS) activity affects sucrose/starch ratio in leaves but does not inhibit normal plant growth in rice. Plant Sci. 2016, 253, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, S.; Zhao, Y.; Li, B.; Zhang, J. Over-expression of AGPase genes enhances seed weight and starch content in transgenic maize. Planta 2011, 233, 241–250. [Google Scholar] [CrossRef]
- Kang, X.; Gao, W.; Cui, B.; El-Aty, A.A. Structure and genetic regulation of starch formation in sorghum (Sorghum bicolor (L.) Moench) endosperm: A review. Int. J. Biol. Macromol. 2023, 239, 124315. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Liu, H.; Yin, X.; Zhao, Q.; Shi, C. Potassium-mediated regulation of sucrose metabolism and storage root formation in sweet potato. Arch. Agron. Soil Sci. 2021, 67, 703–713. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, Z.H.; Xia, H.Q.; Sheng, M.F.; Liu Ming Pan, S.Y.; Li, Z.Y.; Liu, J.R. Potassium fertilization stimulates sucrose-to-starch conversion and root formation in sweet potato (Ipomoea batatas (L.) Lam.). Int. J. Mol. Sci. 2021, 22, 4826. [Google Scholar] [CrossRef] [PubMed]
- Si, C.; Shi, C.; Liu, H.; Zhan, X.; Liu, Y. Effects of nitrogen forms on carbohydrate metabolism and storage-root formation of sweet potato. J. Plant Nutr. Soil Sci. 2018, 181, 419–428. [Google Scholar] [CrossRef]
- Dumbuya, G.; Alemayehu, H.A.; Hasan, M.; Matsunami, M.; Shimono, H. Effect of soil temperature on growth and yield of sweet potato (Ipomoea batatas L.) under cool climate. J. Agric. Meteorol. 2021, 77, 118–127. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Treatment | Average SR Diameter (mm) | Average SR Fresh Weight (g) | Young SR Number (2 < Φ < 5 mm) | Developing SR Number (5 < Φ < 20 mm) | Mature SR Number (Φ > 20 mm) | SR Number |
|---|---|---|---|---|---|---|---|
| 2021 (Haikou) | D15 † | 5.14 c ‡ | 1.35 c | 2.33 a | 2.33 c | - | 4.67 b |
| D20 | 7.84 b | 2.60 b | 1.67 b | 4.00 a | - | 5.67 a | |
| D25 | 8.07 b | 4.87 a | 0.67 c | 3.67 ab | - | 4.33 bc | |
| D30 | 10.62 a | 4.70 a | 0.67 c | 3.00 b | - | 3.67 c | |
| 2022 (Sanya) | D15 | 18.99 b | 18.96 b | - | 3.33 a | 3.67 b | 7.00 a |
| D20 | 21.60 a | 26.74 a | - | 2.33 bc | 4.33 a | 6.67 a | |
| D25 | 21.11 a | 27.74 a | - | 2.00 c | 4.00 ab | 6.00 b | |
| D30 | 22.67 a | 28.87 a | - | 2.67 b | 4.00 ab | 6.67 a | |
| 2023 (Sanya) | D15 | 20.95 c | 31.14 c | - | 0.67 c | 4.67 b | 5.33 c |
| D20 | 20.80 c | 32.90 b | - | 2.33 a | 4.67 a | 7.00 a | |
| D25 | 23.27 b | 36.42 a | - | 1.67 b | 4.67 a | 6.33 b | |
| D30 | 24.66 a | 34.09 ab | - | 1.67 b | 3.67 b | 5.33 c |
| Years | Treatment | SR Diameter (mm) | SR Length (cm) | L/D Ratio | Average SR Weight (g) | CV (%) | Average SR Number Per Plant | Yield (kg/hm2) | Yield Increment (%) |
|---|---|---|---|---|---|---|---|---|---|
| 2021 (Haikou) | D15 † | 36.95 c ‡ | 9.98 bc | 2.7 | 104.56 b | 5.64 | 3.57 d | 29,763.09 b | 14.41 # |
| D20 | 38.75 b | 9.69 c | 2.5 | 111.13 b | 1.77 | 4.52 a | 32,497.89 a | 24.92 | |
| D25 | 39.39 a | 10.24 b | 2.6 | 127.26 a | 11.87 | 4.16 b | 26,013.56 c | - | |
| D30 | 39.15 a | 11.74 a | 3.0 | 133.97 a | 5.49 | 4.00 c | 24,154.95 d | −7.14 | |
| 2022 (Sanya) | D15 | 39.04 b | 10.15 a | 2.6 | 88.50 d | 7.69 | 5.26 c | 38,795.60 b | 3.46 |
| D20 | 39.81 b | 9.15 b | 2.3 | 110.81c | 2.90 | 6.19 a | 42,883.33 a | 14.13 | |
| D25 | 39.72 b | 9.53 ab | 2.4 | 131.66 b | 6.71 | 5.71 b | 37,494.90 b | - | |
| D30 | 42.71 a | 9.82 ab | 2.3 | 170.61a | 7.64 | 5.01 d | 35,592.04 c | −5.34 | |
| 2023 (Sanya) | D15 | 51.64 b | 14.31 b | 2.7 | 174.65 c | 5.55 | 5.44 b | 47,801.67 b | 3.69 |
| D20 | 50.19 b | 15.15 ab | 3.0 | 200.59 b | 3.65 | 6.41 a | 50,025.00 a | 8.47 | |
| D25 | 51.31 b | 15.92 a | 3.9 | 215.37 ab | 4.52 | 5.99 ab | 46,120.27 b | - | |
| D30 | 53.52 a | 15.87 a | 3.0 | 235.26 a | 6.33 | 5.63 b | 45,416.67 b | −1.51 |
| Years | Treatments † | Commercial SR Weight (g) | Commercial SR Number Per Plant | Large SR Number | Middle SR Number | Small SR Number | Commercial Rate (%) | Commercial SR Yield (kg hm−2) | Increment (%) |
|---|---|---|---|---|---|---|---|---|---|
| 2021 (Haikou) | D15 | 122.61 b ‡ | 2.33 c | 0.33 c | 1.67 b | 0.33 a | 65.71 | 19,558.23 b | −0.51 # |
| D20 | 129.91 b | 3.00 a | 1.00 b | 2.00 a | 0.00 b | 74.95 | 24,367.17 a | 23.35 | |
| D25 | 158.45 a | 2.33 c | 1.33 a | 1.00 d | 0.00 b | 75.45 | 19,697.51 b | - | |
| D30 | 162.41 a | 2.67 b | 1.33 a | 1.33 c | 0.00 b | 70.37 | 16,998.00 c | −16.16 | |
| 2022 (Sanya) | D15 | 98.89 d | 3.00 c | 1.00 b | 2.00 b | 0.00 c | 64.32 | 24,954.02 c | 29.60 |
| D20 | 117.89 c | 4.67 a | 1.00 b | 3.00 a | 0.67 a | 80.08 | 34,342.08 a | 33.46 | |
| D25 | 136.87 b | 4.00 b | 1.67 a | 2.00 b | 0.33 b | 68.48 | 25,676.50 b | - | |
| D30 | 201.50 a | 3.00 c | 1.67 a | 1.33 c | 0.00 c | 70.79 | 25,198.06 bc | −1.95 | |
| 2023 (Sanya) | D15 | 179.46 c | 4.33 c | 0.00 d | 4.33 a | 0.00 a | 81.78 | 39,092.21 b | 0.92 |
| D20 | 203.59 b | 5.33 a | 1.00 b | 4.33 a | 0.00 a | 84.39 | 42,216.09 a | 8.96 | |
| D25 | 232.03 a | 4.67 b | 1.67 a | 3.00 b | 0.00 a | 83.99 | 38,736.41 b | - | |
| D30 | 242.80 a | 4.33 c | 1.67 a | 2.67 c | 0.00 a | 79.37 | 36,047.21 c | −6.94 |
| SRN † | SRFW | Yield | IPAR | TPAR | Pn | Gs | |
|---|---|---|---|---|---|---|---|
| SRN | 1.00 | −0.751 *** | 0.793 *** | 0.791 *** | 0.593 ** | −0.596 ** | 0.018 ns |
| SRFW | −0.751 *** # | 1.00 | −0.682 *** | −0.938 *** | −0.618 ** | 0.540 * | 0.324 ns |
| Yield | 0.793 *** | −0.682 *** | 1.00 | 0.764 *** | 0.658 ** | −0.393 ns | 0.010 ns |
| Ci | Tr | Fv/Fm | qP | ΦPSII | ETR | NPQ | |
| SRN | 0.506 * | −0.233 ns | 0.389 ns | −0.546 * | −0.705 *** | −0.500 * | 0.571 ** |
| SRFW | −0.316 ns | 0.429 ns | −0.637 ** | 0.730 ** | 0.901 *** | 0.685 ** | −0.631 ** |
| Yield | 0.317 ns | −0.175 ns | 0.549 * | −0.648 ** | −0.793 *** | −0.676 ** | 0.578 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Q.; Chen, H.; Chen, Y.; Kumar, S.; Chang, H.; Wu, J.; Chen, Y.; Liu, Y.; Wang, Q.; Zhu, G. Appropriate Planting Density Can Improve the Storage Root Yield and Commercial Features of Sweet Potato (Ipomoea batatas L.) by Optimizing the Photosynthetic Performance. Agronomy 2024, 14, 2579. https://doi.org/10.3390/agronomy14112579
Liang Q, Chen H, Chen Y, Kumar S, Chang H, Wu J, Chen Y, Liu Y, Wang Q, Zhu G. Appropriate Planting Density Can Improve the Storage Root Yield and Commercial Features of Sweet Potato (Ipomoea batatas L.) by Optimizing the Photosynthetic Performance. Agronomy. 2024; 14(11):2579. https://doi.org/10.3390/agronomy14112579
Chicago/Turabian StyleLiang, Qinggan, Hongrong Chen, Yanli Chen, Sunjeet Kumar, Hailong Chang, Jiantao Wu, Yue Chen, Yonghua Liu, Qinnan Wang, and Guopeng Zhu. 2024. "Appropriate Planting Density Can Improve the Storage Root Yield and Commercial Features of Sweet Potato (Ipomoea batatas L.) by Optimizing the Photosynthetic Performance" Agronomy 14, no. 11: 2579. https://doi.org/10.3390/agronomy14112579
APA StyleLiang, Q., Chen, H., Chen, Y., Kumar, S., Chang, H., Wu, J., Chen, Y., Liu, Y., Wang, Q., & Zhu, G. (2024). Appropriate Planting Density Can Improve the Storage Root Yield and Commercial Features of Sweet Potato (Ipomoea batatas L.) by Optimizing the Photosynthetic Performance. Agronomy, 14(11), 2579. https://doi.org/10.3390/agronomy14112579