
Monitoring the Degradation of Semi-Natural Grassland Associations under Different Land-Use Patterns

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
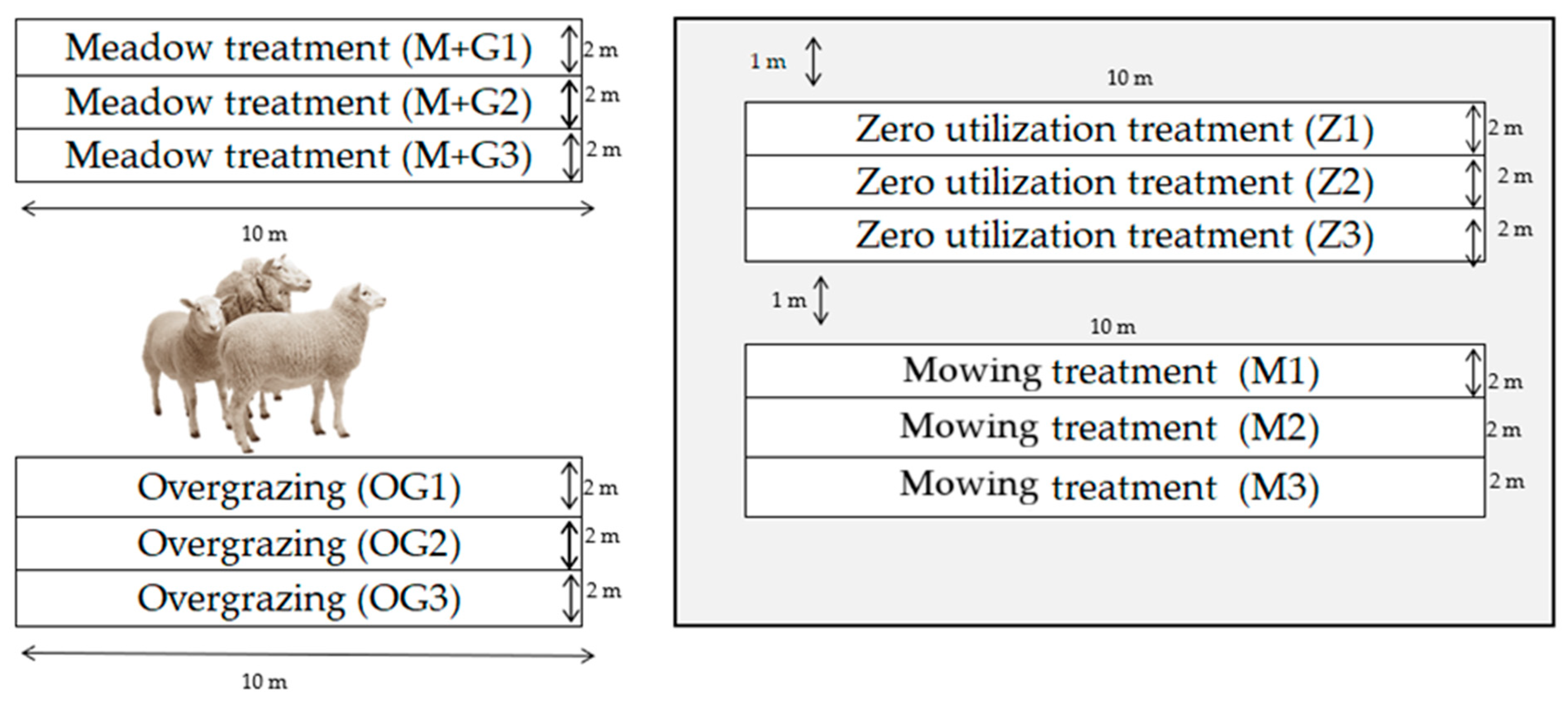
2.1. Description of the Experimental Site
- -
- Zero-utilisation treatment: the plot is not utilised (denoted Z);
- -
- Mowing treatment: removal of phytomass by mowing in the 3rd decade of May (designation: M);
- -
- Meadow treatment (mowing and grazing): removal of phytomass by mowing in the 3rd decade of May, followed by sheep grazing (4 sheep/ha) in August (designation: M + G).
- -
- Overgrazing management treatment: grazing 25 sheep/ha continuously (designation: OG). The whole flock passed through the overgrazed plots and grazed them. The flock consists of female sheep of mixed age (1–4 years), weight (50–90 kg), and breed (Magyar Merino, Blanc du Massif Central, Berrichon du Cher).
2.2. Meteorological Data
2.3. The Methods Used in the Study
- -
- Specialists (S): character species indicating variation in production area. Their absence indicates site disturbance; their reappearance indicates site rehabilitation.
- -
- Competitors (C): Dominant species of natural plant communities indicating community stability.
- -
- Generalists (G): Species of natural communities with a wide ecological tolerance, which play an important role in community stability and maintenance of diversity.
- -
- Natural pioneers (NP): They play an important role in the regeneration or rehabilitation of the community.
- -
- Disturbance-tolerant plant species (DT): Pioneer elements of incipient secondary succession.
- -
- Natural weed species (W): Plants of an area with a persistent anthropogenic influence.
- -
- Invasive alien species (I): Alien flora elements indicating that the site is/was used for persistent economic purposes.
- -
- Invasive species (A): Alien plants of the landscape and flora that have been introduced into the flora as a result of anthropogenic activity.
- -
- Ruderal competitors (RC): Type-forming or dominant weed species of the natural flora that are able to change the direction of succession.
- -
- Aggressive, alien invasive species (AC): Landscape and alien flora plants that have the potential to alter the progressive direction of succession, while creating an alien flora and threatening the survival and rehabilitation of communities.
2.4. Statistical Evaluation of the Experiment
3. Results
3.1. Results of the Botanical Survey
3.2. Evaluation of the SBT Classification of Plants
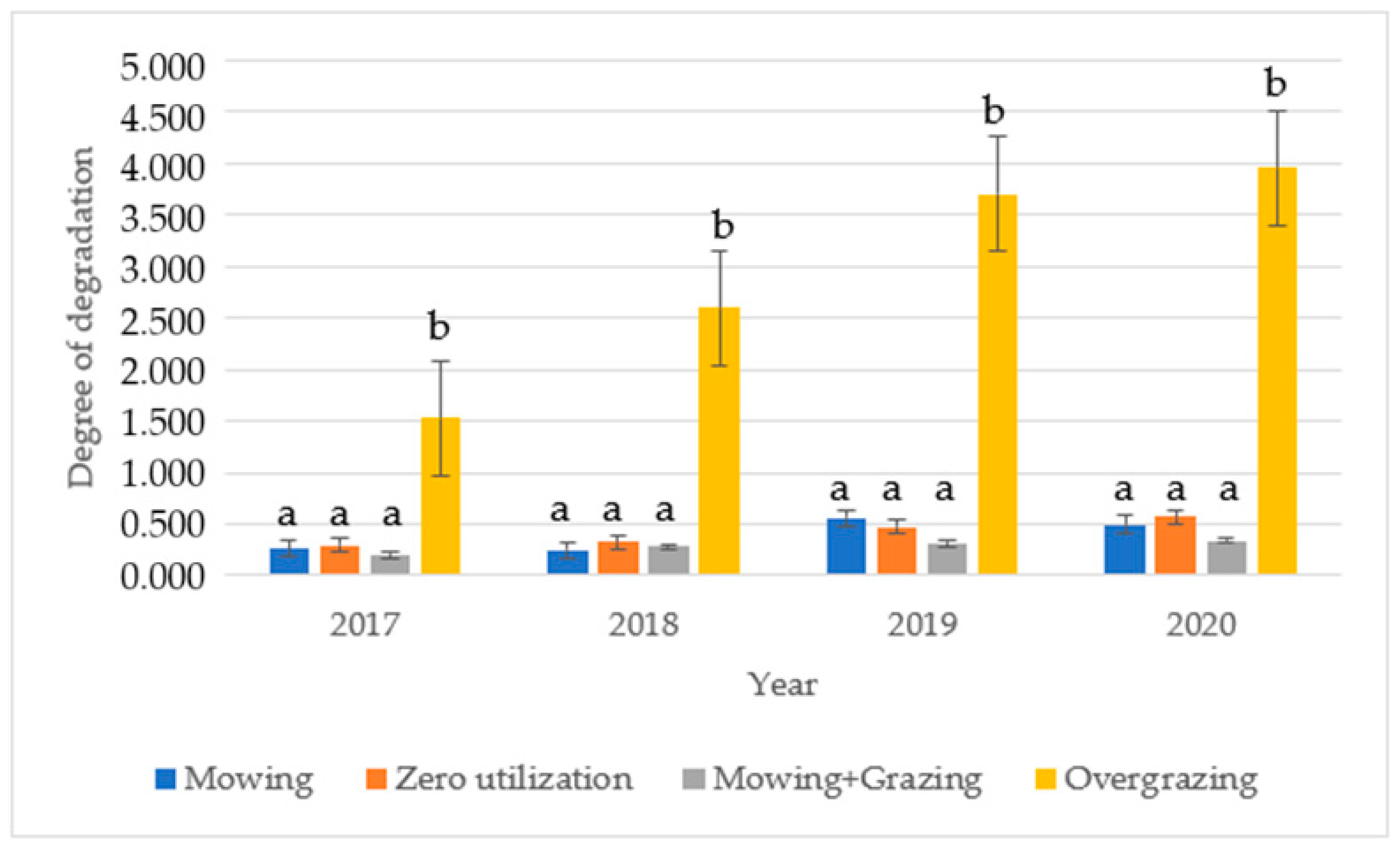
3.3. Results of the Degradation Degree Calculation
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, T.; Hou, G.; Sun, J.; Zong, N.; Shi, P. Degradation shifts plant communities from S- to R-strategy in an alpine meadow, Tibetan Plateau. Sci. Total Environ. 2021, 800, 149572. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Zhang, Y.; Ganjurjav, H.; Dong, S.; Gao, Q. Excessive plant compensatory growth: A potential endogenous driver of meadow degradation on the Qinghai-Tibetan Plateau. Ecosyst. Health Sustain. 2020, 6, 1816500. [Google Scholar] [CrossRef]
- Tiscornia, G.; Jaurena, M.; Baethgen, W. Drivers, Process, and Consequences of Native Grassland Degradation: Insights from a Literature Review and a Survey in Río de la Plata Grasslands. Agronomy 2019, 9, 239. [Google Scholar] [CrossRef]
- Wu, G.L.; Ren, G.H.; Dong, Q.M.; Shi, J.J.; Wang, Y.L. Above- and belowground response along degradation gradient in an alpine grassland of the Qinghai-Tibetan Plateau. Acta Hydrochim. Et Hydrobiol. 2014, 42, 319–323. [Google Scholar] [CrossRef]
- Lu, X.; Kelsey, K.C.; Yan, Y.; Sun, J.; Wang, X.; Cheng, G.; Neff, J.C. Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai-Tibetan Plateau: A synthesis. Ecosphere 2017, 8, e01656. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Salmon, J.M. Mapping the world’s degraded lands. Appl. Geogr. 2015, 57, 12–21. [Google Scholar] [CrossRef]
- Liu, M.; Dries, L.; Wim Heijman, W.; Zhu, X.; Deng, X.; Huang, J. Land tenure reform and grassland degradation in Inner Mongolia, China. China Econ. Rev. 2019, 55, 181–198. [Google Scholar] [CrossRef]
- Li, X.L.; Gao, J.; Brierley, G.; Qiao, Y.M.; Zhang, J.; Yang, Y.W. Rangeland degradation on the Qinghai-Tibet plateau: Implications for rehabilitation. Land Degrad. Dev. 2013, 24, 72–80. [Google Scholar] [CrossRef]
- Lin, L.; Li, Y.K.; Xu, X.L.; Zhang, F.W.; Du, Y.G.; Liu, S.L.; Guo, X.W.; Cao, G.M. Predicting parameters of degradation succession processes of Tibetan Kobresia grasslands. Solid Earth 2015, 6, 1237–1246. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M.; Güneralp, B.; Reilly, M.K. A meta-analysis of global urban land expansion. PLoS ONE 2011, 6, e23777. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yang, X.; Zhou, C.; Shao, X.; Shi, Z.; Li, H.; Su, H.; Qin, R.; Chang, T.; Hu, X.; et al. Alpine Grassland Degradation and Its Restoration in the Qinghai–Tibet Plateau. Grasses 2023, 2, 31–46. [Google Scholar] [CrossRef]
- Zhang, W.; Xue, X.; Peng, F.; You, Q.; Hao, A. Meta-analysis of the effects of grassland degradation on plant and soil properties in the alpine meadows of the Qinghai-Tibetan Plateau. Glob. Ecol. Conserv. 2019, 20, e00774. [Google Scholar] [CrossRef]
- Xie, H.; Zhang, Y.; Wu, Z.; Lv, T. A Bibliometric Analysis on Land Degradation: Current Status, Development, and Future Directions. Land 2020, 9, 28. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Veldman, J.W.; Buisson, E.; Durigan, G.; Fernandes, G.W.; Le Stradic, S.; Mahy, G.; Negreiros, D.; Overbeck, G.E.; Veldman, R.G.; Zaloumis, N.P.; et al. Toward an old-growth concept for grasslands, savannas, and woodlands. Front. Ecol. Environ. 2015, 13, 154–162. [Google Scholar] [CrossRef]
- Wick, A.F.; Geaumont, B.A.; Sedivec, K.; Hendrickson, J. Grassland degradation. In Biological and Environmental Hazards, Risks and Disasters; Shroder, J.F., Sivanpillai, R., Eds.; Elsevier: New York, NY, USA, 2016; Volume 8, pp. 257–276. ISBN 9780123964717. [Google Scholar]
- Andrade, B.O.; Marchesi, E.; Burkart, S.; Setubal, R.B.; Lezama, F.; Perelman, S.; Schneider, A.A.; Trevisan, R.; Overbeck, G.E.; Boldrini, I.I. Vascular plant species richness and distribution in the Río de la Plata grasslands. Bot. J. Linn. Soc. 2018, 188, 250–256. [Google Scholar] [CrossRef]
- Gang, C.C.; Zhou, W.; Chen, Y.Z.; Wang, Z.Q.; Sun, Z.G.; Li, J.L.; Odeh, I. Quantitative assessment of the contributions of climate change and human activities on global grassland degradation. Environ. Earth Sci. 2014, 72, 4273–4282. [Google Scholar] [CrossRef]
- Zhou, W.; Gang, C.; Zhou, L.; Chen, Y.; Li, J.; Ju, W.; Odeh, I. Dynamic of grassland vegetation degradation and its quantitative assessment in the northwest China. Acta Oecol. 2014, 55, 86–96. [Google Scholar] [CrossRef]
- Lark, T.J.; Spawn, S.A.; Bougie, M.; Gibbs, H.K. Cropland expansion in the United States produces marginal yields at high costs to wildlife. Nat. Commun. 2020, 11, 4295. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, L.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef] [PubMed]
- Siyabulela, S.; Tefera, S.; Wakindiki, I.; Keletso, M. Comparison of grass and soil conditions around water points in different land use systems in semi-arid South African rangelands and implications for management and current rangeland paradigms. Arid. Land Res. Manag. 2020, 34, 207–230. [Google Scholar] [CrossRef]
- Kamp, J.; Koshkin, M.A.; Bragina, T.M.; Katzner, T.E.; Milner-Gulland, E.J.; Schreiber, E.; Sheldon, R.; Shmalenko, A.; Smelansky, I.; Terraube, J.; et al. Persistent and novel threats to the biodiversity of Kazakhstan’s steppes and semi-deserts. Biodivers. Conserv. 2016, 25, 2521–2541. [Google Scholar] [CrossRef]
- Shahriary, E.; Langford, R.P.; Gill, T.E.; Hussein, M.; Hargrove, W.L.; Golding, P. Partitioning variation in vegetation communities around Lajaneh Piosphere, Iran. Arid. Land Res. Manag. 2021, 35, 32–54. [Google Scholar] [CrossRef]
- Gaitán, J.J.; Bran, D.E.; Oliva, G.E.; Aguiar, M.R.; Buono, G.G.; Ferrante, D.; Nakamatsu, V.; Ciari, G.; Salomone, J.M.; Massara, V.; et al. Aridity and overgrazing have convergent effects on ecosystem structure and functioning in Patagonian rangelands. Land Degrad. Dev. 2018, 29, 210–218. [Google Scholar] [CrossRef]
- Abu Hammad, A.; Tumeizi, A. Land degradation: Socioeconomic and environmental causes and consequences in the eastern Mediterranean. Land Degrad. Dev. 2012, 23, 216–226. [Google Scholar] [CrossRef]
- Vetter, S.; Bond, W.J. Changing predictors of spatial and temporal variability in stocking rates in a severely degraded communal rangeland. Land Degrad. Dev. 2012, 23, 190–199. [Google Scholar] [CrossRef]
- Kovácsné Koncz, N.; Béri, B.; Deák, B.; Kelemen, A.; Tóth, K.; Kiss, R.; Radócz, S.; Miglécz, T.; Tóthmérész, B.; Valkó, O. Meat production and maintaining biodiversity: Grazing by traditional and crossbred beef cattle breeds in marshes and grasslands. Appl. Veg. Sci. 2020, 23, 139–148. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z. Quantitative Analysis of Driving Factors of Grassland Degradation: A Case Study in Xilin River Basin, InnerMongolia. Sci. World J. 2012, 2012, 169724. [Google Scholar] [CrossRef] [PubMed]
- Mor-Mussery, A.; Abu-Glaion, H.; Shuker, S.; Zaady, E. The influence of trampling by small ruminants on soil fertility in semi-arid rangelands. Arid. Land Res. Manag. 2020, 35, 189–197. [Google Scholar] [CrossRef]
- Borer, E.T.; Seabloom, E.W.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Lind, E.M.; Adler, P.B.; Alberti, J.; Anderson, T.M.; Bakker, J.D.; et al. Herbivores and nutrients control grassland plant diversity via light limitation. Nature 2014, 508, 517. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.; Deng, X.; Song, W.; Li, Z.; Chen, J. What is the main cause of grassland degradation? A case study of grassland ecosystem service in the middle-south Inner Mongolia. CATENA 2017, 150, 100–107. [Google Scholar] [CrossRef]
- Evans, R. Overgrazing and soil erosion on hill pastures with particular reference to the Peak District. Grass Forage Sci. Soc. 2006, 32, 65–76. [Google Scholar] [CrossRef]
- Zhao, Y.; Peth, S.; Krummelbein, J.; Horn, R.; Wang, Z.Y.; Steffens, M.; Hoffmann, C.; Peng, X.H. Spatial variability of soil properties affected by grazing intensity in Inner Mongolia Grassland. Ecol. Model. 2007, 205, 241–254. [Google Scholar] [CrossRef]
- Jiang, A.; Jing, L.H.; Mipam, T.-D.; Tian, L.M. Progress in research on the effects of grazing on grassland litter decomposition. Acta Pratacult. Sin. 2022, 32, 208–220. [Google Scholar]
- Isselstein, J.; Jeangros, B.; Pavlů, V. Agronomic aspects of biodiversity targeted management of temperate grasslands in Europe—A review. Agric. Res. 2005, 3, 139–151. [Google Scholar]
- Perevolotsky, A.; Seligman, N.G. Role of grazing in Mediterranean rangeland ecosystems. Bioscience 1998, 48, 1007–1017. [Google Scholar] [CrossRef]
- Bakker, J.P.; Berendse, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol. Evol. 1999, 14, 63–68. [Google Scholar] [CrossRef]
- Bajor, Z.; Zimmermann, Z.; Szabó, G.; Fehér, Z.; Járdi, I.; Lampert, R.; Kerény-Nagy, V.; Penksza, P.; Szabó, Z.L.; Székely, Z.; et al. Effect of conservation management practices on sand grassland vegetation in Budapest, Hungary. Appl. Ecol. Environ. Res. 2016, 14, 233–247. [Google Scholar] [CrossRef]
- Erdős, L.; Bátori, Z.; Tölgyesi, C.; Körmöczi, L. The moving split window (MSW) analysis in vegetation science—An overview. Appl. Ecol. Environ. Res. 2014, 12, 787–805. [Google Scholar] [CrossRef]
- Erdős, L.; Cserhalmi, D.; Bátori, Z.; Kiss, T.; Morschhauser, T.; Benyhe, B.; Dénes, A. Shrub encroachment in a wooded-steppe mosaic: Combining GIS methods with landscape historical analysis. Appl. Ecol. Environ. Res. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Erdős, L.; Tölgyesi, C.; Dénes, A.; Darányi, N.; Fodor, A.; Bátori, Z.; Tolnay, D. Comparative analysis of the natural and semi–natural plant communities of Mt Nagy and other parts of the Villány (Mts south Hungary). Thaiszia J. Bot. 2014, 24, 1–21. [Google Scholar]
- Szentes, S.; Sutyinszki, Z.; Szabó, G.; Zimmermann, Z.; Házi, J.; Wichmann, B.; Hufnágel, L.; Penksza, K.; Bartha, S. Grazed Pannonian grassland beta-diversity changes due to C4 yellow bluestem. Cent. Eur. J. Biol. 2012, 7, 1055–1065. [Google Scholar] [CrossRef]
- Ma, L.; Yao, Z.; Zheng, X.; Zhang, H.; Wang, K.; Zhu, B.; Wang, R.; Zhang, W.; Liu, C. Increasing grassland degradation stimulates the non-growing season CO2 emissions from an alpine meadow on the Qinghai–Tibetan Plateau. Environ. Sci. Pollut. Res. 2018, 25, 26576–26591. [Google Scholar] [CrossRef]
- Bartha, S.; Szentes, S.; Horváth, A.; Házi, J.; Zimmermann, Z.; Molnár, C.; Dancza, I.; Margóczi, K.; Pál, R.; Purger, D.; et al. Impact of mid-successional dominant species on the diversity and progress of succession in regenerating temperate grasslands. Appl. Veg. Sci. 2014, 17, 201–213. [Google Scholar] [CrossRef]
- Wright, C.K.; Wimberly, M.C. Recent land use change in the western corn belt threatens grasslands and wetlands. Proc. Natl. Acad. Sci. USA 2013, 110, 4134–4139. [Google Scholar] [CrossRef]
- Sauer, J.R.; Link, W.A.; Fallon, J.E.; Pardieck, K.L.; Ziolkowski, D.J., Jr. The North American breeding bird survey 1966e2011: Summary analysis and species accounts. N. Am. Fauna 2012, 79, 1–32. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Archer, D.; Hendrickson, J.; Kronberg, S.; Liebig, M.; Nichols, K.; Schmer, M.; Tanaka, D.; Aguilar, J. Diversification and ecosystem services for conservation agriculture: Outcomes from pastures and integrated crop-livestock systems. Renew. Agric. Food Syst. 2013, 28, 129–144. [Google Scholar] [CrossRef]
- Varga, K.; Csízi, I. Túllegeltetett természetközeli gyeptársulás rekultivációja legeltetés kizárással. Gyepgazdálkodási Közlemények 2020, 18, 45–53. [Google Scholar] [CrossRef]
- Fernandez-Gimenez, M.E.; Le Febre, S. Mobility in pastoral systems: Dynamic flux or downward trend? The International Journal of Sustainable. Dev. World Ecol. 2006, 13, 341–362. [Google Scholar] [CrossRef]
- Tasi, J.; Bajnok, M.; Halász, A.; Szabó, F.; Harkányiné Székely, Z.; Láng, V. Magyarországi komplex gyepgazdálkodási adatbázis létrehozásának első lépései és eredményei. Gyepgazdálkodási Közlemények 2014, 12, 57–58. [Google Scholar] [CrossRef]
- Hortobágyi, T.; Simon, T. Növényföldrajz, Társulástan és Ökológia; Nemzeti Tankönyvkiadó: Budapest, Hungary, 2002; pp. 1–538. [Google Scholar]
- Šefferová Stanová, V.; Janák, M.; Ripka, J. Management of Natura 2000 Habitats. In 1530*Pannonic Salt Steppes and Salt Marshes; European Commission: Brussels, Belgium, 2008. [Google Scholar]
- Balázs, F. A gyepek termésbecslése növénycönológia alapján. Agrártudományok 1949, 1, 25–35. [Google Scholar]
- Király, G.; Virók, V.; Szmorad, F.; Molnár, V.A. Új magyar füvészkönyv: Magyarország hajtásos növényei: Határozókulcsok; Aggteleki Nemzeti Park Igazgatóság: Jósvafő, Hungray, 2009; pp. 1–616. [Google Scholar]
- Borhidi, A. A Magyar Flóra Szociális Magatartástípusa, Természetességi és Relatív Ökológiai Értékszámai; KTM-OTVH-JPTE: Pécs, Hungary, 1993; pp. 1–93. [Google Scholar]
- Penksza, K.; Házi, J.; Tóth, A.; Wichmann, B.; Pajor, F.; Gyuricza, C.S.; Póti, P.; Szentes, S. Eltérő hasznosítású szürkemarha legelő szezonális táplálóanyag-tartalom alakulás, fajdiverzitás változása és ennek hatása a biomassza mennyiségére és összetételére nedves pannon gyepekben. Növénytermelés 2013, 62, 73–94. [Google Scholar]
- Török, P.; Janišová, M.; Kuzemko, A.; Rūsiņa, S.; Stevanović, Z.D. Grasslands, their threats and management in Eastern Europe. In Grasslands of the World: Diversity, Management and Conservation; CRC Press: Boca Raton, FL, USA, 2018; pp. 67–88. [Google Scholar]
- Házi, J.; Bartha, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural grassland management by mowing of Calamagrostis epigejos in Hungary. Plant Biosyst. 2011, 145, 699–707. [Google Scholar] [CrossRef]
- Házi, J.; Penksza, K.; Bartha, S.; Hufnagel, L.; Tóth, A.; Gyuricza, C.; Szentes, S. Cut mowing and grazing Effects with grey cattle on plant species composition in case of Pannon wet grasslands. Appl. Ecol. Environ. Res. 2012, 10, 223–231. [Google Scholar] [CrossRef]
- Valkó, O.; Venn, S.; Zmihoski, M.; Biurrun, I.; Labadessa, R.; Loos, J. The challenge of abandonment for the sustainable management of Palaearctic natural and semi-natural grasslands. Hacquetia 2018, 17, 5–16. [Google Scholar] [CrossRef]
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóth, E.; Tóthmérész, B. Managing for composition or species diversity? Pastoral and year-round grazing systems in alkali grasslands. Agric. Ecosyst. Environ. 2016, 234, 23–30. [Google Scholar] [CrossRef]
- Török, P.; Valkó, O.; Deák, B.; Kelemen, A.; Tóthmérész, B. Traditional cattle grazing in a mosaic alkali landscape: Effects on grassland biodiversity along a moisture gradient. PLoS ONE 2014, 9, e97095. [Google Scholar] [CrossRef]
- Pápay, G. Cserjeirtás után magára hagyott, legeltetett és kaszált gyepterületek vegetációjának összehasonlító elemzése parádóhutai (Mátra) mintaterületen. Gyepgazdálkodási Közlemények 2016, 14, 37–48. [Google Scholar] [CrossRef]
- Pápay, G.; Penksza, K.; Szabó, G.; Ibadzane, M.; Járdi, I.; Wichmann, B. Természetvédelmi kezelések hatása hegyi rétek vegetációjára a Gyöngyösi Sár-hegy TT területén. Gyepgazdálkodási Közlemények 2017, 15, 37–46. [Google Scholar] [CrossRef]
- Penksza, K.; Pápay, G.; Házi, J.; Tóth, A.; S-Falusi, E.; Saláta, D.; Kerényi-Nagy, V.; Wichmann, B. Gyepregeneráció erdőirtással kialakított gyepekben mátrai (Fallóskút) mintaterületeken. Gyepgazdálkodási Közlemények 2015, 13, 31–44. [Google Scholar] [CrossRef]
- Tóth, E.; Deák, B.; Valkó, O.; Kelemen, A.; Miglécz, T.; Tóthmérész, B.; Török, P. Livestock type is more crucial than grazing intensity: Traditional cattle and sheep grazing in shortgrass steppes. Land Degrad. Dev. 2016, 29, 231–239. [Google Scholar] [CrossRef]
- Nagy, G. A gyephasználat és a vidékfejlesztés összefüggései. In Gyepgazdálkodásunk helyzete és kilátásai; Debreceni Gyepgazdálkodási Napok 17: Debrecen, Hungary, 2001; pp. 24–25. [Google Scholar]
- Dufour, A.; Gadallah, F.; Wagner, H.H.; Guisan, A.; Buttler, A. Plant species richness and environmental heterogeneity in a mountain landscape: Effects of variability and spatial configuration. Ecography 2006, 29, 573–584. [Google Scholar] [CrossRef]
- Vickery, J.A.; Tallowin, J.R.; Feber, R.E.; Asteraki, E.J.; Atkinson, P.W.; Fuller, R.J.; Brown, V.K. The management of lowland neutral grasslands in Britain: Effects of agricultural practices on birds and their food resources. J. Appl. Ecol. 2001, 38, 647–664. [Google Scholar] [CrossRef]
- Czóbel, S.; Szirmai, O.; Németh, Z.; Gyuricza, C.S.; Házi, J.; Tóth, A.; Schellenberger, J.; Vasa, L.; Penksza, K.P. Shortterm effects of grazing exclusion on net ecosystem CO2 exchange and net primary production in a Pannonian sandy grassland. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 67–72. [Google Scholar] [CrossRef][Green Version]
- Stefán, E. Az alsószuhai szőlőhegy tájtörténeti és botanikai vizsgálata. Bot. Közlemények 2018, 105, 129–142. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 2017 | 2018 | 2019 | 2020 | |
|---|---|---|---|---|
| Yearly precipitation (mm) | 527.50 | 557.80 | 505.10 | 648.50 |
| Yearly average temperature (°C) | 11.20 | 12.50 | 13.30 | 11.70 |
| Treatment/Plant Group | Year | Overgrazing (OG) | Mowing (M) | Zero Utilisation (Z) | Mowing and Grazing (M + G) |
|---|---|---|---|---|---|
| Rare specialists (Sr) | 2017 | 0.00 | 0.00 | 0.00 | 0.52 |
| 2018 | 0.00 | 0.00 | 0.00 | 0.52 | |
| 2019 | 0.00 | 0.00 | 0.00 | 0.52 | |
| 2020 | 0.00 | 0.00 | 0.00 | 0.52 | |
| Specialists (S) | 2017 | 2.08 | 0.00 | 0.00 | 2.08 |
| 2018 | 1.04 | 2.08 | 0.00 | 1.56 | |
| 2019 | 1.04 | 0.00 | 0.00 | 1.04 | |
| 2020 | 1.04 | 0.00 | 0.00 | 2.08 | |
| Competitors (C) | 2017 | 28.13 | 54.02 | 62.50 | 68.23 |
| 2018 | 19.27 | 73.44 | 42.71 | 49.13 | |
| 2019 | 14.06 | 36.46 | 48.44 | 57.81 | |
| 2020 | 14.06 | 40.63 | 49.48 | 56.25 | |
| Generalists (G) | 2017 | 6.77 | 37.50 | 14.58 | 10.94 |
| 2018 | 6.25 | 18.75 | 27.08 | 9.90 | |
| 2019 | 4.69 | 34.38 | 19.79 | 13.02 | |
| 2020 | 4.69 | 33.33 | 47.58 | 11.98 | |
| Natural pioneers (NP) | 2017 | 0.52 | 0.00 | 0.00 | 1.56 |
| 2018 | 0.00 | 0.00 | 0.00 | 0.52 | |
| 2019 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 2020 | 0.00 | 0.00 | 0.00 | 0.52 | |
| Disturbance tolerant plants (DT) | 2017 | 27.60 | 1.56 | 17.71 | 9.90 |
| 2018 | 29.17 | 3.65 | 15.63 | 10.42 | |
| 2019 | 33.85 | 6.25 | 21.35 | 12.50 | |
| 2020 | 30.21 | 2.60 | 17.19 | 13.02 | |
| Weeds (W) | 2017 | 25.52 | 0.00 | 1.04 | 2.08 |
| 2018 | 30.21 | 0.00 | 1.04 | 4.69 | |
| 2019 | 32.81 | 4.69 | 0.52 | 3.13 | |
| 2020 | 41.67 | 4.17 | 8.85 | 3.13 | |
| Ruderal competitors (RC) | 2017 | 5.21 | 5.21 | 2.08 | 3.65 |
| 2018 | 3.65 | 2.08 | 4.17 | 5.21 | |
| 2019 | 4.17 | 13.54 | 6.25 | 5.21 | |
| 2020 | 4.17 | 14.58 | 11.46 | 6.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, K.; Csízi, I.; Halász, A.; Mezőszentgyörgyi, D.; Nagy, D. Monitoring the Degradation of Semi-Natural Grassland Associations under Different Land-Use Patterns. Agronomy 2024, 14, 35. https://doi.org/10.3390/agronomy14010035
Varga K, Csízi I, Halász A, Mezőszentgyörgyi D, Nagy D. Monitoring the Degradation of Semi-Natural Grassland Associations under Different Land-Use Patterns. Agronomy. 2024; 14(1):35. https://doi.org/10.3390/agronomy14010035
Chicago/Turabian StyleVarga, Krisztina, István Csízi, András Halász, Dávid Mezőszentgyörgyi, and Dominik Nagy. 2024. "Monitoring the Degradation of Semi-Natural Grassland Associations under Different Land-Use Patterns" Agronomy 14, no. 1: 35. https://doi.org/10.3390/agronomy14010035
APA StyleVarga, K., Csízi, I., Halász, A., Mezőszentgyörgyi, D., & Nagy, D. (2024). Monitoring the Degradation of Semi-Natural Grassland Associations under Different Land-Use Patterns. Agronomy, 14(1), 35. https://doi.org/10.3390/agronomy14010035