
The Effect of Soil Microbial Residues-Mediated Nitrogen Conservation and Supply during the Growing Season on Nitrogen Uptake by Wheat

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
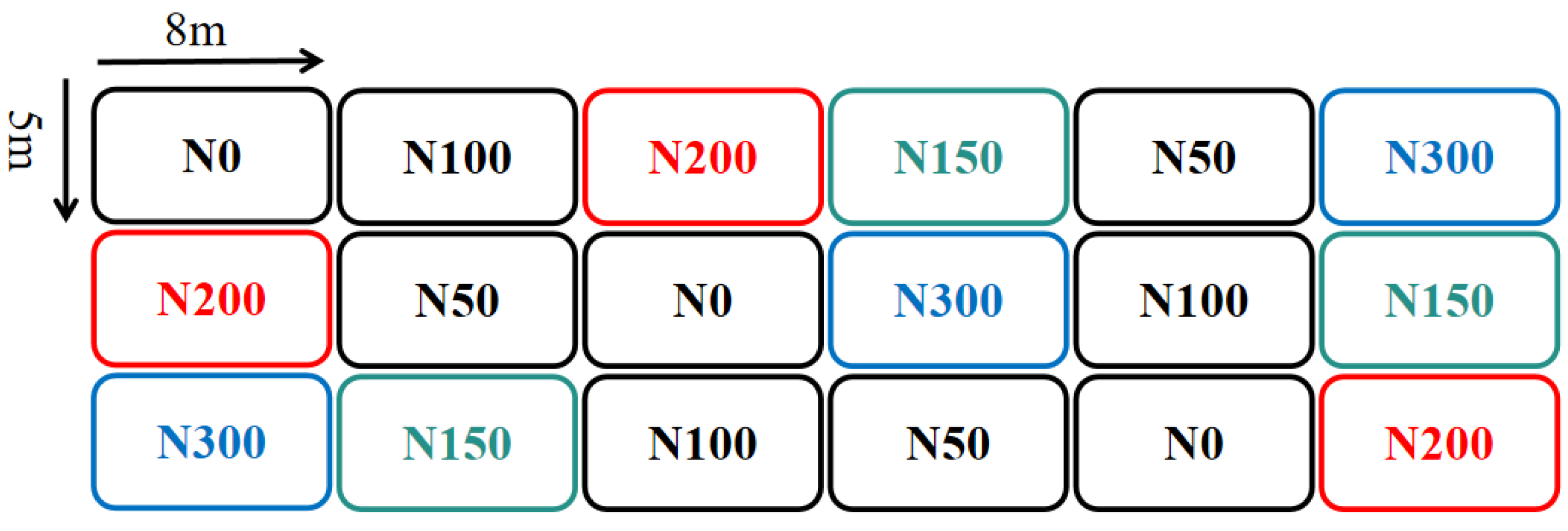
2.1. Site Description and Experimental Design
2.2. Sample Collection and Measurement
2.2.1. Soil and Plant Sample Collection
2.2.2. Determination of Total N and 15N Abundance in Soil and Plants
2.2.3. Analysis of Amino Sugars and 15N Enrichment
2.3. Calculation
2.3.1. Remaining Fertilizer N in Soil
2.3.2. Fertilizer-Derived Fungal, Bacterial, and Total Microbial Necromass N in Soil
2.4. Statistical Analysis
3. Results
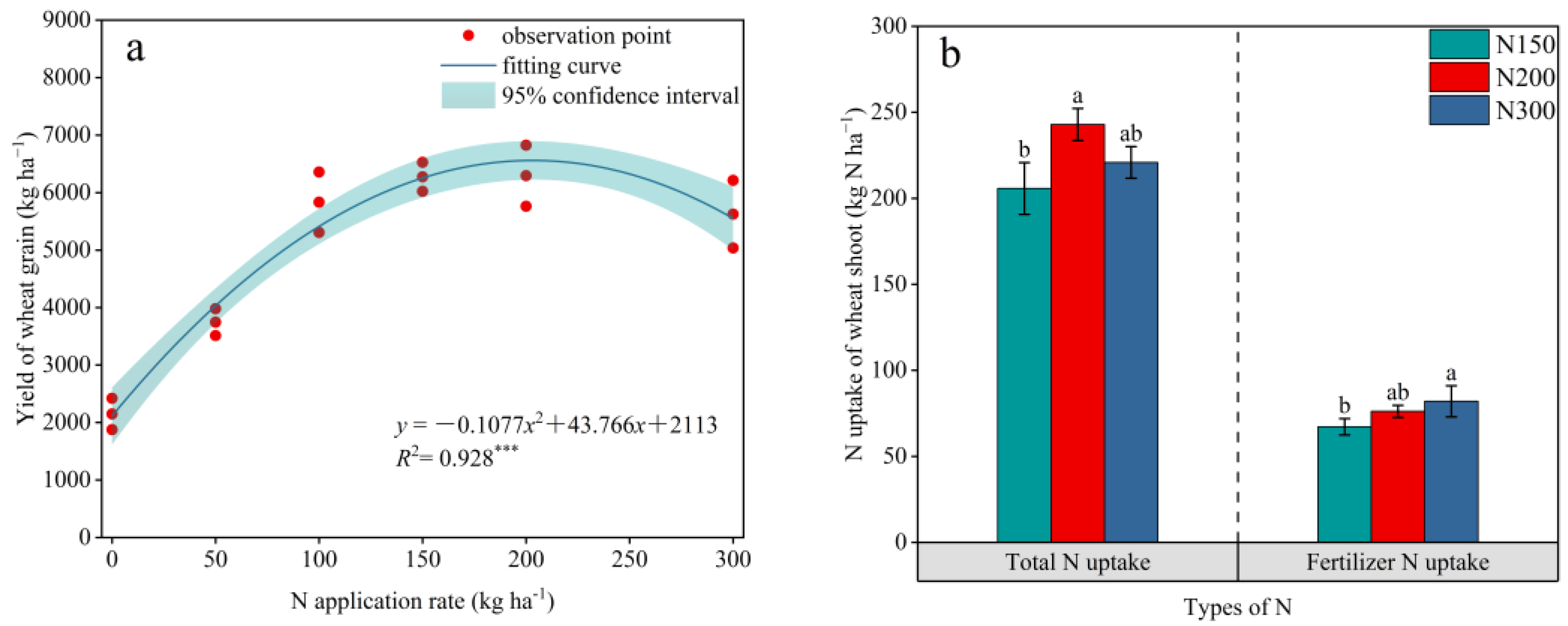
3.1. Effects of Long-Term Different N Application on Crop Yield and N Uptake
3.2. Effects of Long-Term Fertilization on Soil Physical and Chemical Properties
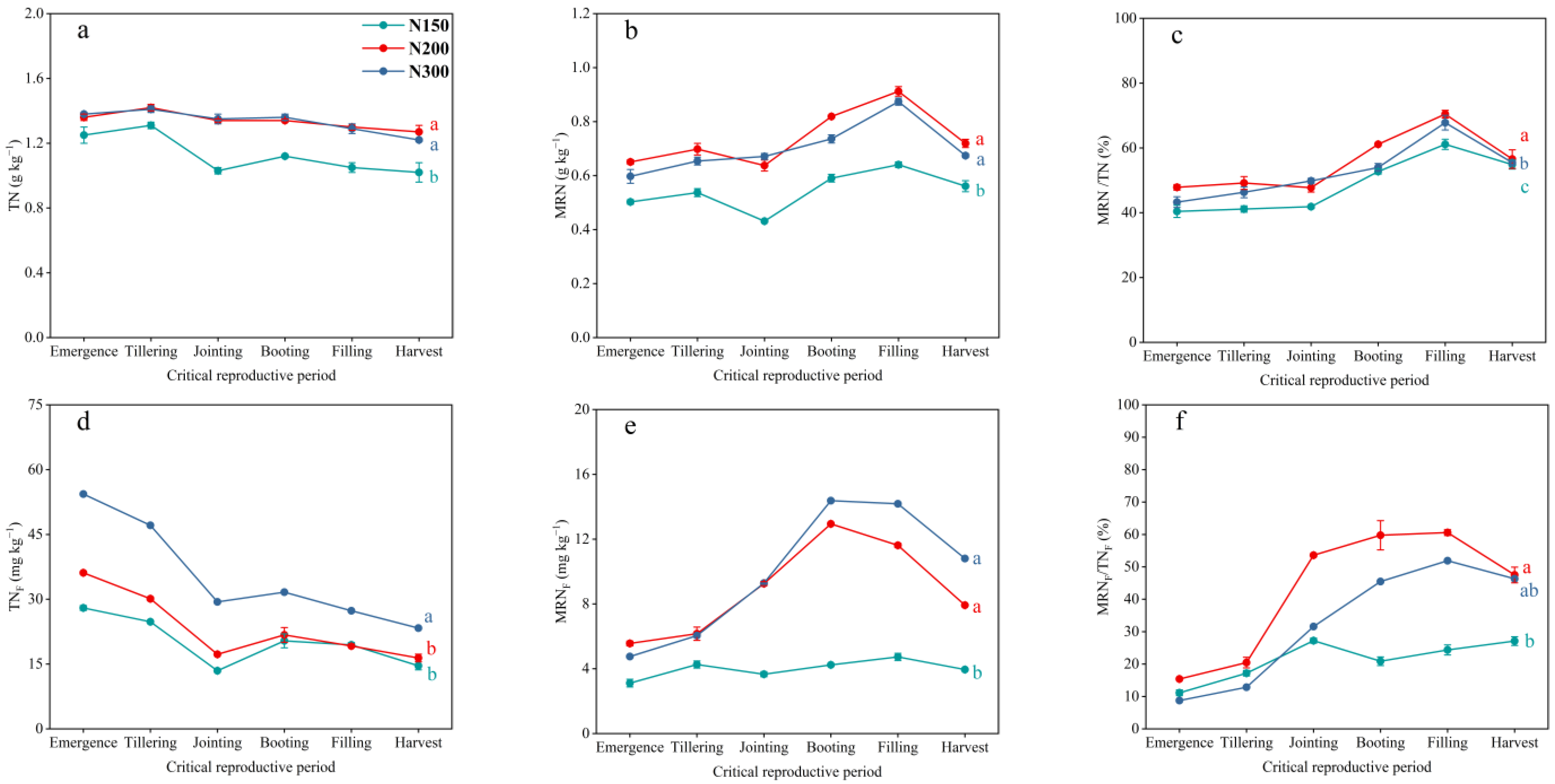
3.3. Dynamics of Soil TN and MRN Pools and Fertilizer N in Those Pools
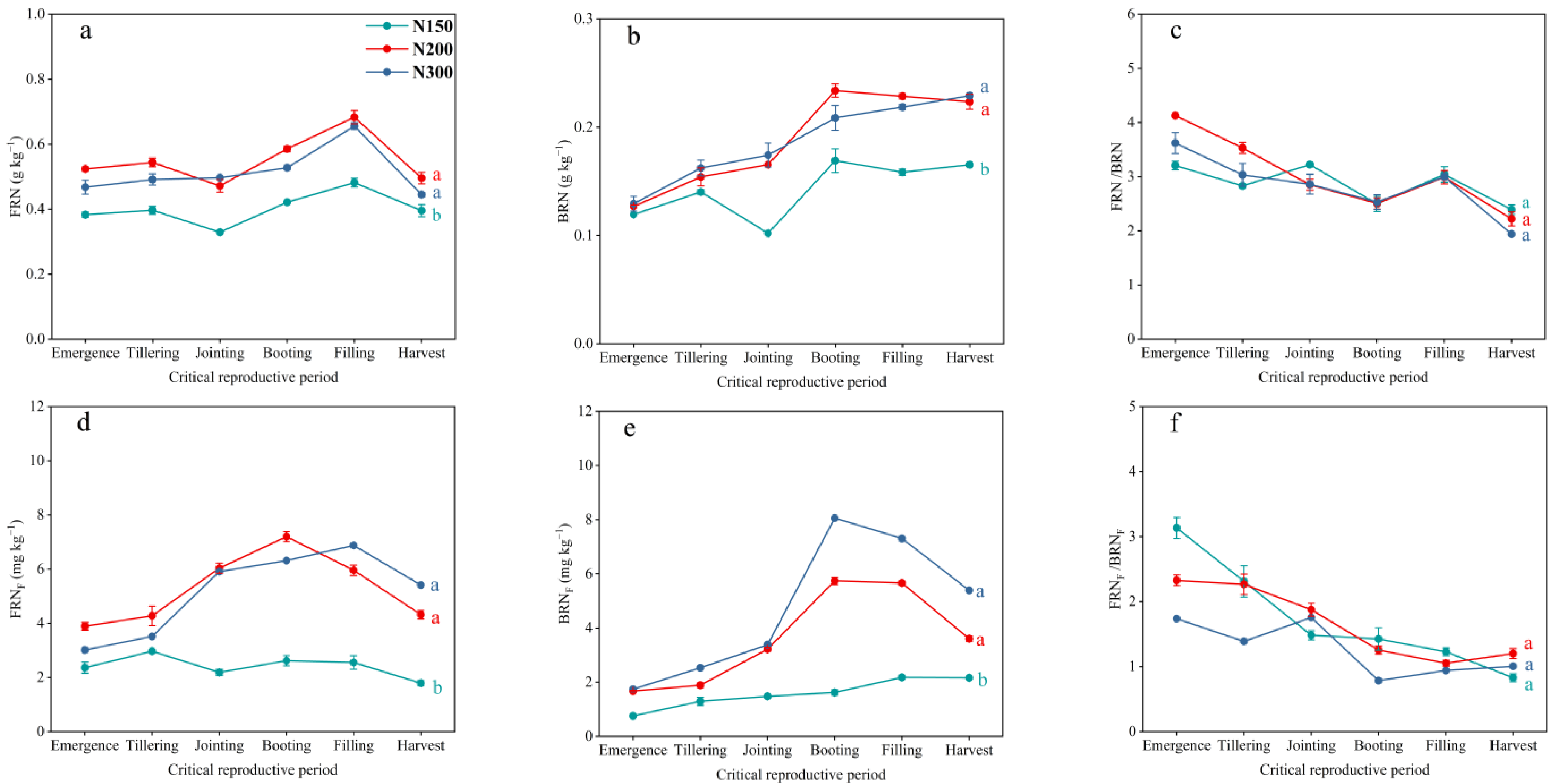
3.4. Dynamics of Soil FRN and BRN Pools and Fertilizer N in Those Pools
3.5. Dynamic Changes in the Efficiency of Fertilizer Nitrogen-Synthesizing Microbial Residue Nitrogen Driven by Different Long-Term Nitrogen Application Levels
3.6. Dynamics of Rhizosphere Soil MRN, FRN, and BRN Pools and Fertilizer N in Those Pools
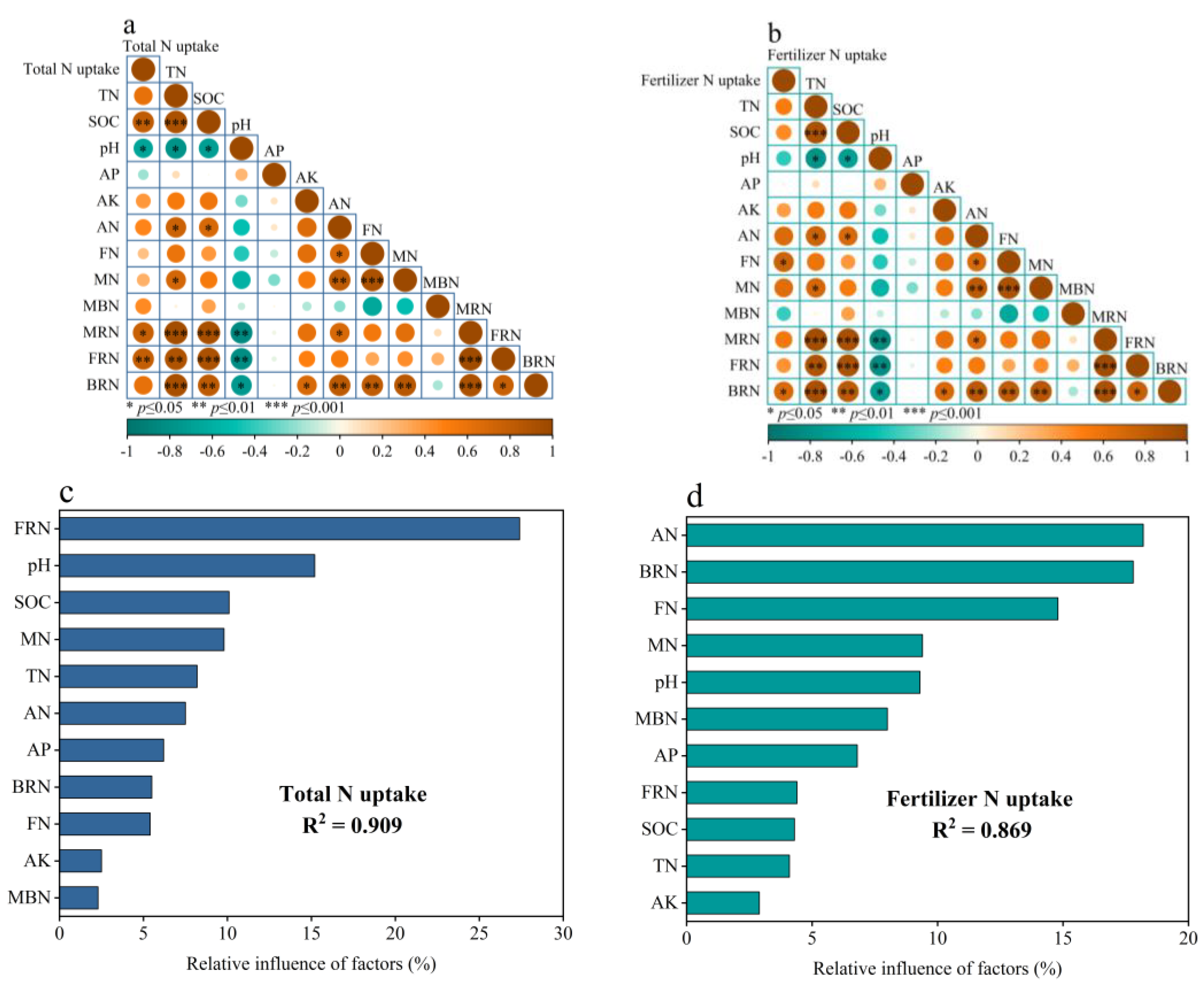
3.7. Correlation between Crop Nitrogen Uptake and Soil Biotic and Abiotic Factors
4. Discussion
4.1. The Effect of Different Nitrogen Levels on Microbial Residue
4.2. The Effect of Microbial Residue Nitrogen on Crop Nitrogen Uptake
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. The Fate of Nitrogen from Soil to Plants: Influence of Agricultural Practices in Modern Agriculture. Agriculture 2021, 11, 944. [Google Scholar] [CrossRef]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a Century of Ammonia Synthesis Changed the World. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing Nitrogen for Sustainable Development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Zhang, X.; Davidson, E.A.; Zhu, F.; Li, S.; Zhao, X.; Chen, X.; Zhang, L.; He, J.; Wei, W.; et al. Fates and Use Efficiency of Nitrogen Fertilizer in Maize Cropping Systems and Their Responses to Technologies and Management Practices: A Global Analysis on Field 15N Tracer Studies. Earth’s Future 2021, 9, e2020EF001514. [Google Scholar] [CrossRef]
- Martínez-Dalmau, J.; Berbel, J.; Ordóñez-Fernández, R. Nitrogen Fertilization. A Review of the Risks Associated with the Inefficiency of Its Use and Policy Responses. Sustainability 2021, 13, 5625. [Google Scholar] [CrossRef]
- Stevens, W.B.; Hoeft, R.G.; Mulvaney, R.L. Fate of Nitrogen-15 in a Long-Term Nitrogen Rate Study: II. Nitrogen Uptake Efficiency. Agron. J. 2005, 97, 1046–1053. [Google Scholar] [CrossRef]
- Pandey, A.; Eldridge, S.M.; Weatherley, A.; Willett, I.R.; Myint, A.K.; Oo, A.N.; Ngwe, K.; Mang, Z.T.; Chen, D. High Fertilizer Nitrogen Input Increases Nitrogen Mining in Sandy Paddy Soils. Nutr. Cycl. Agroecosyst. 2023, 125, 77–88. [Google Scholar] [CrossRef]
- Dan, X.; He, M.; Meng, L.; He, X.; Wang, X.; Chen, S.; Cai, Z.; Zhang, J.; Zhu, B.; Müller, C. Strong Rhizosphere Priming Effects on N Dynamics in Soils with Higher Soil N Supply Capacity: The ‘Matthew Effect’ in Plant-Soil Systems. Soil Biol. Biochem. 2023, 178, 108949. [Google Scholar] [CrossRef]
- Singh, B. Are Nitrogen Fertilizers Deleterious to Soil Health? Agronomy 2018, 8, 48. [Google Scholar] [CrossRef]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R.; Boast, C.W. The Myth of Nitrogen Fertilization for Soil Carbon Sequestration. J. Environ. Qual. 2007, 36, 1821–1832. [Google Scholar] [CrossRef]
- Chen, Z.; Zhou, X.; Geng, S.; Miao, Y.; Cao, Y.; Chen, Z.; Zhang, J.; Han, S. Interactive Effect of Nitrogen Addition and Throughfall Reduction Decreases Soil Aggregate Stability through Reducing Biological Binding Agents. For. Ecol. Manag. 2019, 445, 13–19. [Google Scholar] [CrossRef]
- Wang, F.; Liu, Y.; Liang, B.; Liu, J.; Zong, H.; Guo, X.; Wang, X.; Song, N. Variations in Soil Aggregate Distribution and Associated Organic Carbon and Nitrogen Fractions in Long-Term Continuous Vegetable Rotation Soil by Nitrogen Fertilization and Plastic Film Mulching. Sci. Total Environ. 2022, 835, 155420. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, X.; Reis, S.; Gu, B. Socioeconomic Barriers of Nitrogen Management for Agricultural and Environmental Sustainability. Agric. Ecosyst. Environ. 2022, 333, 107950. [Google Scholar] [CrossRef]
- Hayatsu, M.; Tago, K.; Saito, M. Various Players in the Nitrogen Cycle: Diversity and Functions of the Microorganisms Involved in Nitrification and Denitrification. Soil Sci. Plant Nutr. 2008, 54, 33–45. [Google Scholar] [CrossRef]
- Ma, Q.; Wu, Z.; Pan, F.; Wang, J.; Zhou, H.; Jiang, C.; Xu, Y.; Yu, W. Effect of Glucose Addition on the Fate of Urea-15N in Fixed Ammonium and Soil Microbial Biomass N Pools. Eur. J. Soil Biol. 2016, 75, 168–173. [Google Scholar] [CrossRef]
- Pajares, S.; Bohannan, B.J.M. Ecology of Nitrogen Fixing, Nitrifying, and Denitrifying Microorganisms in Tropical Forest Soils. Front. Microbiol. 2016, 7, 1045. [Google Scholar] [CrossRef] [PubMed]
- Liang, C. Soil Microbial Carbon Pump: Mechanism and Appraisal. Soil Ecol. Lett. 2020, 2, 241–254. [Google Scholar] [CrossRef]
- Liang, C.; Zhu, X. The Soil Microbial Carbon Pump as a New Concept for Terrestrial Carbon Sequestration. Sci. China Earth Sci. 2021, 64, 545–558. [Google Scholar] [CrossRef]
- Glaser, B.; Millar, N.; Blum, H. Sequestration and Turnover of Bacterial- and Fungal-Derived Carbon in a Temperate Grassland Soil under Long-Term Elevated Atmospheric p CO2. Glob. Chang. Biol. 2006, 12, 1521–1531. [Google Scholar] [CrossRef]
- Joergensen, R.G. Amino Sugars as Specific Indices for Fungal and Bacterial Residues in Soil. Biol. Fertil. Soils 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Mason, K.E.; McNamara, N.P.; Ostle, N.; Puissant, J.; Goodall, T.; Griffiths, R.I.; Stott, A.W.; Whitaker, J. Environmental and Microbial Controls on Microbial Necromass Recycling, an Important Precursor for Soil Carbon Stabilization. Commun. Earth Environ. 2020, 1, 36. [Google Scholar] [CrossRef]
- Glaser, B.; Turrión, M.-B.; Alef, K. Amino Sugars and Muramic Acid—Biomarkers for Soil Microbial Community Structure Analysis. Soil Biol. Biochem. 2004, 36, 399–407. [Google Scholar] [CrossRef]
- He, H.; Xie, H.; Zhang, X. A Novel GC/MS Technique to Assess 15N and 13C Incorporation into Soil Amino Sugars. Soil Biol. 2006, 38, 1083–1091. [Google Scholar] [CrossRef]
- Zhou, F.; Zhang, X.; Ma, S.; Li, Y.; Zhu, M.; Zhang, W.; Li, J.; Liu, X.; Hu, G.; Wang, X.; et al. Soil Microbial Necromass Regulation of Long-Term Fertilizer N Retention Influenced by Maize Stover Mulching. Geoderma 2023, 433, 116453. [Google Scholar] [CrossRef]
- Anning, D.K.; Li, Z.; Qiu, H.; Deng, D.; Zhang, C.; Ghanney, P.; Shen, Q. Divergent Accumulation of Microbial Residues and Amino Sugars in Loess Soil after Six Years of Different Inorganic Nitrogen Enrichment Scenarios. Appl. Sci. 2021, 11, 5788. [Google Scholar] [CrossRef]
- Hu, Q.; Liu, T.; Ding, H.; Li, C.; Tan, W.; Yu, M.; Liu, J.; Cao, C. Effects of Nitrogen Fertilizer on Soil Microbial Residues and Their Contribution to Soil Organic Carbon and Total Nitrogen in a Rice-Wheat System. Appl. Soil Ecol. 2023, 181, 104648. [Google Scholar] [CrossRef]
- Zhou, R.; Liu, Y.; Dungait, J.A.J.; Kumar, A.; Wang, J.; Tiemann, L.K.; Zhang, F.; Kuzyakov, Y.; Tian, J. Microbial Necromass in Cropland Soils: A Global Meta-analysis of Management Effects. Glob. Chang. Biol. 2023, 29, 1998–2014. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, D.; Luan, H.; Liu, D.; Wang, X.; Sun, J.; Zhou, W.; Liang, G. Living and Dead Microorganisms in Mediating Soil Carbon Stocks Under Long-Term Fertilization in a Rice-Wheat Rotation. Front. Microbiol. 2022, 13, 854216. [Google Scholar] [CrossRef]
- Groffman, P.M.; Fahey, T.J.; Fisk, M.C.; Yavitt, J.B.; Sherman, R.E.; Bohlen, P.J.; Maerz, J.C. Earthworms Increase Soil Microbial Biomass Carrying Capacity and Nitrogen Retention in Northern Hardwood Forests. Soil Biol. Biochem. 2015, 87, 51–58. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L.; et al. Reducing Environmental Risk by Improving N Management in Intensive Chinese Agricultural Systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The Importance of Anabolism in Microbial Control over Soil Carbon Storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Creamer, C.; Whitaker, J. Deconstructing the Microbial Necromass Continuum to Inform Soil Carbon Sequestration. Funct. Ecol. 2022, 36, 1396–1410. [Google Scholar] [CrossRef]
- Liao, S.; Tan, S.; Peng, Y.; Wang, D.; Ni, X.; Yue, K.; Wu, F.; Yang, Y. Increased Microbial Sequestration of Soil Organic Carbon under Nitrogen Deposition over China’s Terrestrial Ecosystems. Ecol. Process. 2020, 9, 52. [Google Scholar] [CrossRef]
- Hu, J.; Huang, C.; Zhou, S.; Liu, X.; Dijkstra, F.A. Nitrogen Addition Increases Microbial Necromass in Croplands and Bacterial Necromass in Forests: A Global Meta-Analysis. Soil Biol. Biochem. 2022, 165, 108500. [Google Scholar] [CrossRef]
- Zhang, W.; Cui, Y.; Lu, X.; Bai, E.; He, H.; Xie, H.; Liang, C.; Zhang, X. High Nitrogen Deposition Decreases the Contribution of Fungal Residues to Soil Carbon Pools in a Tropical Forest Ecosystem. Soil Biol. Biochem. 2016, 97, 211–214. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and Mechanisms of Responses by Soil Microbial Communities to Nitrogen Addition. Soil Biol. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-Term Effects of Mineral Fertilizers on Soil Microorganisms—A Review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Engelking, B.; Flessa, H.; Joergensen, R.G. Shifts in Amino Sugar and Ergosterol Contents after Addition of Sucrose and Cellulose to Soil. Soil Biol. Biochem. 2007, 39, 2111–2118. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, W.; Zhang, X.; Bao, X.; Xie, H.; Li, J.; He, H.; Liang, C.; Zhang, X. Dynamics of Microbial Necromass in Response to Reduced Fertilizer Application Mediated by Crop Residue Return. Soil Biol. Biochem. 2022, 165, 108512. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, H.; Mao, Z.; Bao, X.; He, H.; Zhang, X.; Liang, C. Fungi Determine Increased Soil Organic Carbon More than Bacteria through Their Necromass Inputs in Conservation Tillage Croplands. Soil Biol. Biochem. 2022, 167, 108587. [Google Scholar] [CrossRef]
- Xu, Y.; Gao, X.; Liu, Y.; Li, S.; Liang, C.; Lal, R.; Wang, J. Differential Accumulation Patterns of Microbial Necromass Induced by Maize Root vs. Shoot Residue Addition in Agricultural Alfisols. Soil Biol. Biochem. 2022, 164, 108474. [Google Scholar] [CrossRef]
- Su, F.; Chen, X.; Zhang, L.; Hao, M.; Wei, X. Dynamics of Microbial Residues in Highland Agroecosystems as Affected by Cropping Systems and Fertilisation in a 31-year-long Experiment. Eur. J. Soil Sci. 2022, 73, e13205. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Y.; Zhang, Y.; Huang, D.; Li, X.; Gregorich, E.; McLaughlin, N.; Zhang, X.; Chen, X.; Zhang, S.; et al. Effect of Long-Term Tillage and Cropping System on Portion of Fungal and Bacterial Necromass Carbon in Soil Organic Carbon. Soil Tillage Res. 2022, 218, 105307. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Brabcová, V.; Štursová, M.; Davidová, A.; Jansa, J.; Cajthaml, T.; Baldrian, P. Decomposer Food Web in a Deciduous Forest Shows High Share of Generalist Microorganisms and Importance of Microbial Biomass Recycling. ISME J. 2018, 12, 1768–1778. [Google Scholar] [CrossRef]
- Akroume, E.; Maillard, F.; Bach, C.; Hossann, C.; Brechet, C.; Angeli, N.; Zeller, B.; Saint-André, L.; Buée, M. First Evidences That the Ectomycorrhizal Fungus Paxillus Involutus Mobilizes Nitrogen and Carbon from Saprotrophic Fungus Necromass. Environ. Microbiol. 2019, 21, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Bindschedler, S.; Job, D.; Wick, L.Y.; Filippidou, S.; Kooli, W.M.; Verrecchia, E.P.; Junier, P. Exploiting the Fungal Highway: Development of a Novel Tool for the in Situ Isolation of Bacteria Migrating along Fungal Mycelium. FEMS Microbiol. Ecol. 2015, 91, fiv116. [Google Scholar] [CrossRef] [PubMed]
- Villarino, S.H.; Talab, E.; Contisciani, L.; Videla, C.; Di Geronimo, P.; Mastrángelo, M.E.; Georgiou, K.; Jackson, R.B.; Piñeiro, G. A Large Nitrogen Supply from the Stable Mineral-Associated Soil Organic Matter Fraction. Biol. Fertil. Soils 2023, 59, 833–841. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, L.; Gao, X.; Wang, J. Contrasting Response of Fungal versus Bacterial Residue Accumulation within Soil Aggregates to Long-Term Fertilization. Sci. Rep. 2022, 12, 17834. [Google Scholar] [CrossRef]
- Xue, P.; Pei, J.; Ma, N.; Wang, J. Microbial Residual Nitrogen Distribution in Brown Earth’s Aggregates as Affected by Different Maize Residues and Soil Fertility Levels. Front. Environ. Sci. 2022, 10, 892039. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Treatment | ||
|---|---|---|---|
| N150 | N200 | N300 | |
| SOC (g kg−1) | 9.25 ± 0.32 c | 11.39 ± 0.16 a | 10.39 ± 0.05 b |
| pH | 8.53 ± 0.08 a | 8.39 ± 0.01 b | 8.44 ± 0.03 ab |
| AP (mg kg−1) | 35.41 ± 2.83 a | 35.02 ± 3.10 a | 34.85 ± 3.18 a |
| AK (mg kg−1) | 125.80 ± 2.90 a | 130.85 ± 5.75 a | 132.81 ± 2.18 a |
| AN (mg kg−1) | 123.23 ± 2.24 b | 134.76 ± 6.20 a | 136.58 ± 7.19 a |
| FN (mg kg−1) | 148.76 ± 0.73 c | 156.68 ± 0.53 b | 170.41 ± 0.49 a |
| MN (mg kg−1) | 33.39 ± 1.50 c | 40.59 ± 3.14 b | 46.31 ± 3.48 a |
| MBN (mg kg−1) | 2.96 ± 0.18 a | 3.25 ± 0.12 a | 2.51 ± 0.18 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.; Xing, L.; Zhang, C.; Li, J.; Li, Y.; Zhang, Y.; He, H.; Hu, C.; Li, X.; Zhang, L.; et al. The Effect of Soil Microbial Residues-Mediated Nitrogen Conservation and Supply during the Growing Season on Nitrogen Uptake by Wheat. Agronomy 2024, 14, 193. https://doi.org/10.3390/agronomy14010193
Han J, Xing L, Zhang C, Li J, Li Y, Zhang Y, He H, Hu C, Li X, Zhang L, et al. The Effect of Soil Microbial Residues-Mediated Nitrogen Conservation and Supply during the Growing Season on Nitrogen Uptake by Wheat. Agronomy. 2024; 14(1):193. https://doi.org/10.3390/agronomy14010193
Chicago/Turabian StyleHan, Jian, Li Xing, Chuang Zhang, Jundi Li, Yannan Li, Yuming Zhang, Hongbo He, Chunsheng Hu, Xiaoxin Li, Lijuan Zhang, and et al. 2024. "The Effect of Soil Microbial Residues-Mediated Nitrogen Conservation and Supply during the Growing Season on Nitrogen Uptake by Wheat" Agronomy 14, no. 1: 193. https://doi.org/10.3390/agronomy14010193
APA StyleHan, J., Xing, L., Zhang, C., Li, J., Li, Y., Zhang, Y., He, H., Hu, C., Li, X., Zhang, L., Dong, W., Qin, S., & Liu, X. (2024). The Effect of Soil Microbial Residues-Mediated Nitrogen Conservation and Supply during the Growing Season on Nitrogen Uptake by Wheat. Agronomy, 14(1), 193. https://doi.org/10.3390/agronomy14010193