

Identification of Donors for Fresh Seed Dormancy and Marker Validation in a Diverse Groundnut Mini-Core Collection

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
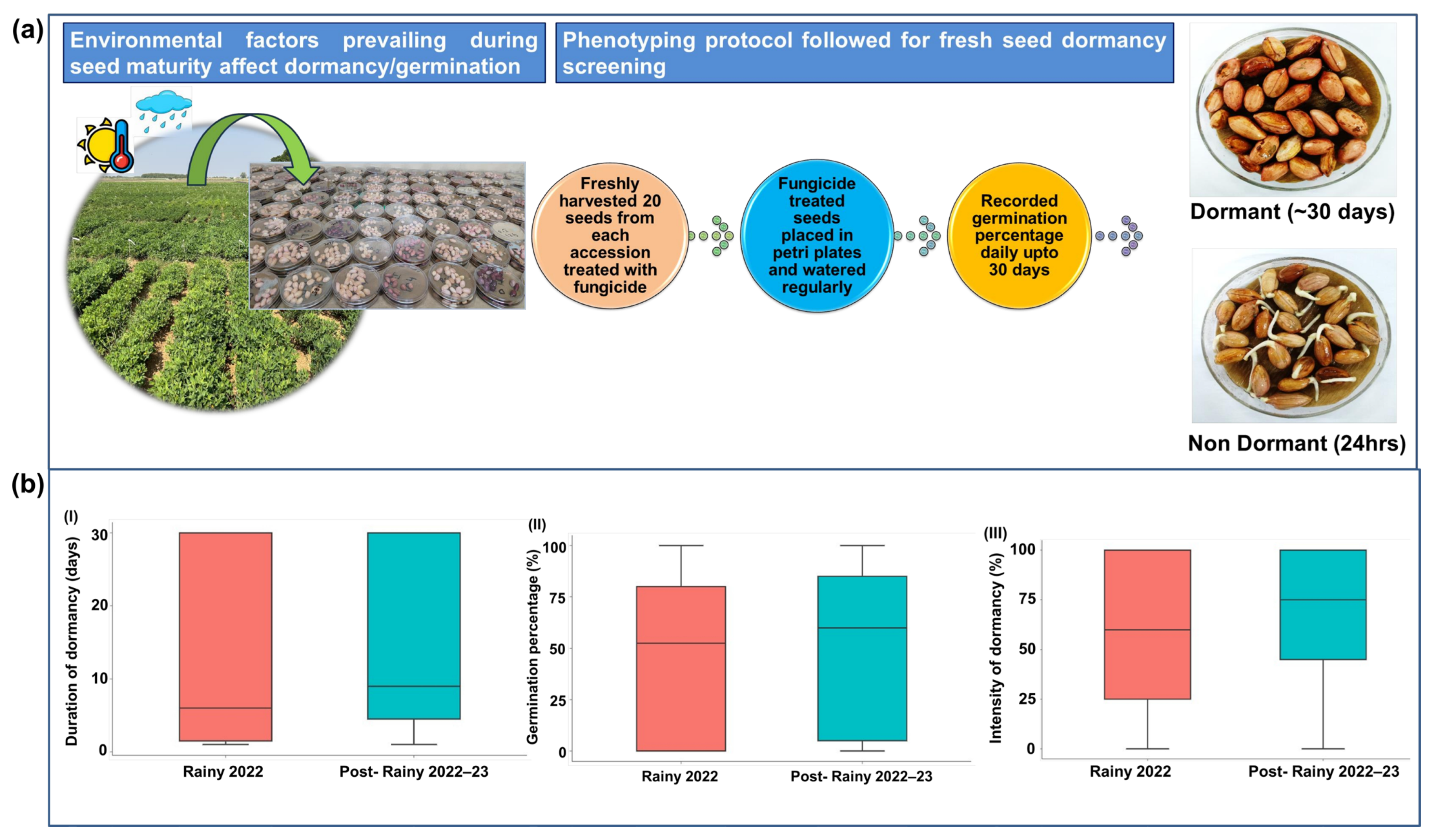
2.2. Phenotyping Protocol and Parameters Estimated for Fresh Seed Dormancy
2.3. Statistical Analysis
2.4. Validation of Allele-Specific Markers Associated with Fresh Seed Dormancy
3. Results and Discussion
3.1. Analysis of Variance
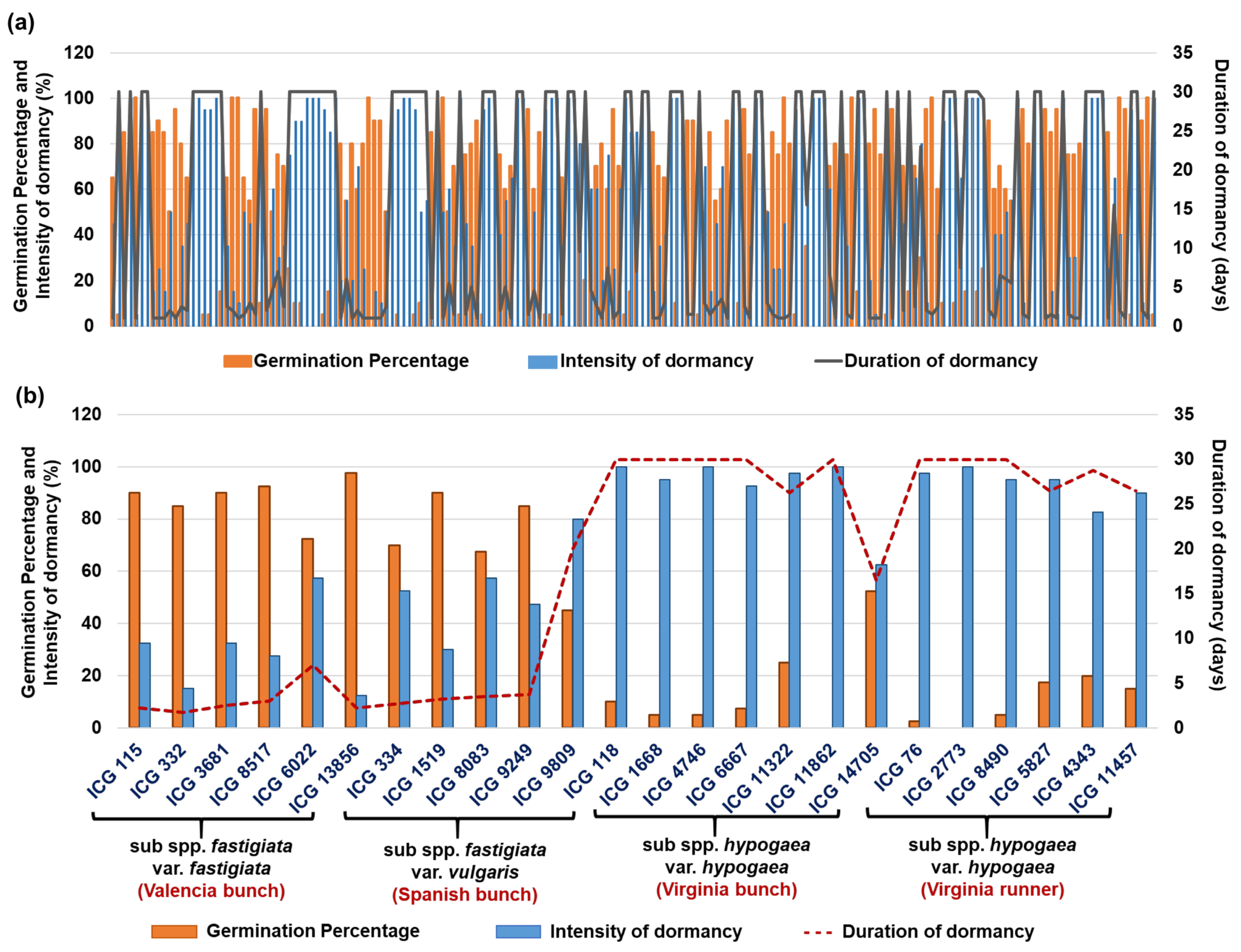
3.2. Germination Percentage, Duration, and Intensity of Dormancy
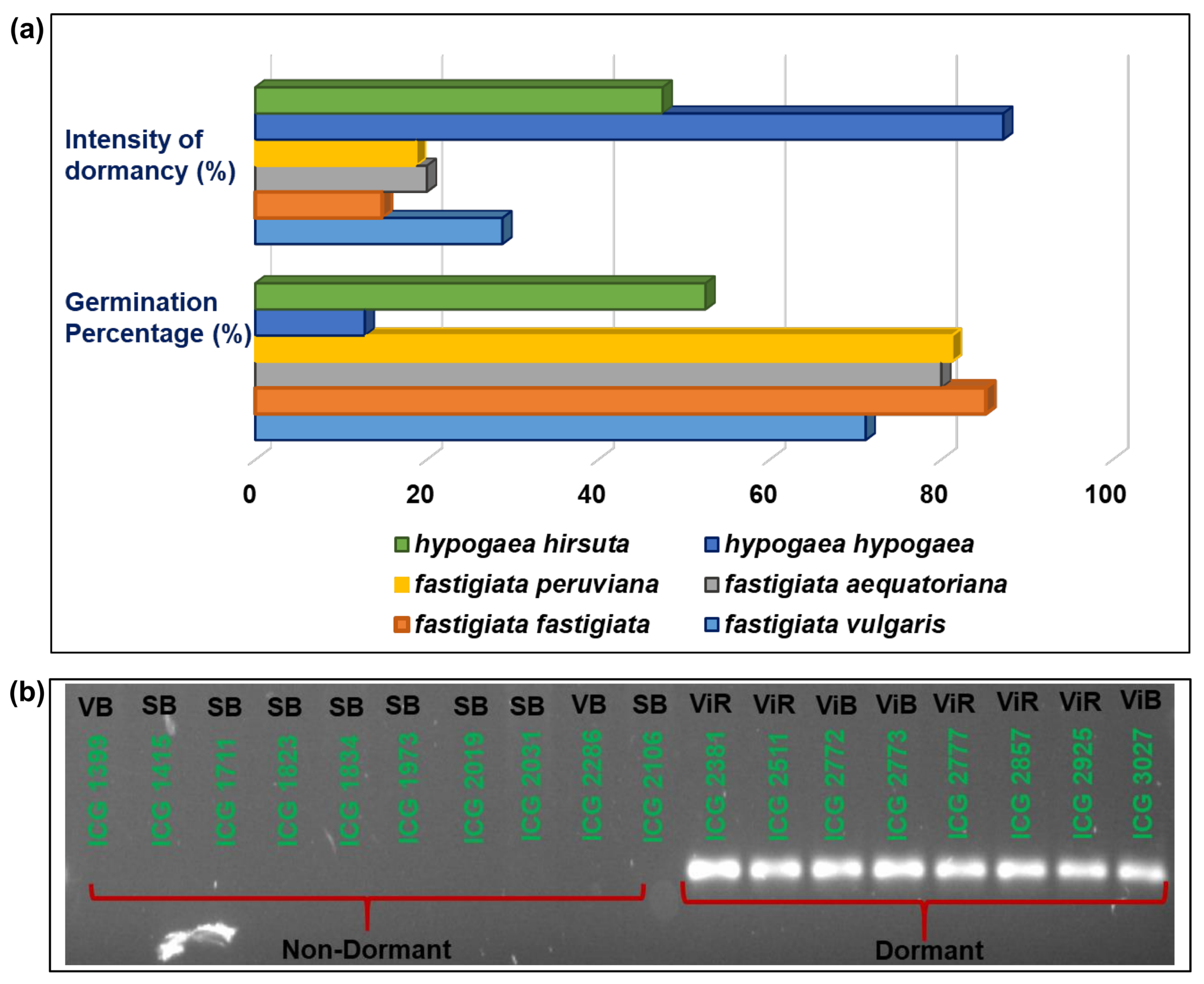
3.3. The Validation of Mini-Core Accessions with Allele-Specific Markers Associated with Fresh Seed Dormancy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krapovickas, A. The origin, variability and spread of the groundnut (Arachis hypogaea). In The Domestication and Exploitation of Plant and Animals; Ucko, R.J., Dimbledy, W.C., Eds.; Greald Duckworth Co., Ltd.: London, UK, 1969; pp. 427–441. [Google Scholar]
- Krapovickas, A. Evolution of the genus Arachis. In Agricultural Genetics-Selected Topics; Moav, R., Ed.; Wiley: New York, NY, USA, 1973; pp. 135–151. [Google Scholar]
- Upadhyaya, H.D.; Nigam, S.N. Inheritance of fresh seed dormancy in peanut. Crop Sci. 1999, 39, 98–101. [Google Scholar] [CrossRef]
- Naganagoudar, Y.B.; Kenchanagoudar, P.V.; Rathod, S.; Keerthi, C.M.; Nadaf, H.L.; Channappagoudar, B.B. Inheritance of fresh seed dormancy in recombinant inbred lines (RIL) developed for mapping population TAG 24 x GPBD 4 in groundnut (Arachis hypogaea L.). Legum. Res. 2016, 39, 844–846. [Google Scholar] [CrossRef]
- Vishwakarma, M.K.; Pandey, M.K.; Shasidhar, Y.; Manohar, S.S.; Nagesh, P.; Janila, P.; Varshney, R.K. Identification of two major quantitative trait locus for fresh seed dormancy using the diversity arrays technology and diversity arrays technology-seq based genetic map in Spanish-type peanuts. Plant Breed. 2016, 135, 367–375. [Google Scholar] [CrossRef]
- Zhang, C.; Selvaraj, J.N.; Yang, Q.; Liu, Y. A survey of aflatoxin-producing Aspergillus sp. from peanut field soils in four agroecological zones of China. Toxins 2017, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Patro, H.K.; Ray, M. Seed dormancy in groundnut—A review. Int. J. Trop. Agric. 2016, 34, 31–37. [Google Scholar]
- Wang, M.L.; Wang, H.; Zhao, C.; Tonnis, B.; Tallury, S.; Wang, X.; Clevenger, J.; Guo, B. Identification of QTLs for seed dormancy in cultivated peanut using a recombinant inbred line mapping population. Plant Mol. Biol. Rep. 2021, 40, 208–217. [Google Scholar] [CrossRef]
- Gupta, R.K.; Singh, S.S.; Verma, M.M. Induction of dormancy in groundnut (Arachis hypogea L.) variety T−64 by maleic hydra zide. J. Agric. Res. 1985, 19, 82–86. [Google Scholar]
- Footitt, S.; Douterelo-Soler, I.; Clay, H.; Finch-Savage, W.E. Dormancy cycling in Arabidopsis seeds is controlled by seasonally distinct hormone-signaling pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 20236–20241. [Google Scholar] [CrossRef]
- Yaw, A.J.; Richard, A.; Safo-Kantanka, O.; Adu-Dapaah, H.K.; Ohemeng-Dapaah, S.; Agyeman, A. Inheritance of fresh seed dormancy in groundnut. Afr. J. Biotechnol. 2008, 7, 421–424. [Google Scholar]
- Zhang, M.; Zeng, Q.; Liu, H.; Qi, F.; Sun, Z.; Miao, L.; Li, X.; Li, C.; Liu, D.; Guo, J.; et al. Identification of a stable major QTL for fresh-seed germination on chromosome Arahy. 04 in cultivated peanut (Arachis hypogaea L.). Crop J. 2022, 10, 1767–1773. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Bramel, P.J.; Ortiz, R.; Singh, S. Developing a mini core of peanut for utilization of genetic resources. Crop Sci. 2002, 42, 2150–2156. [Google Scholar] [CrossRef]
- Bomireddy, D.; Gangurde, S.S.; Variath, M.T.; Janila, P.; Manohar, S.S.; Sharma, V.; Parmar, S.; Deshmukh, D.; Reddisekhar, M.; Reddy, D.M.; et al. Discovery of major quantitative trait loci and candidate genes for fresh seed dormancy in groundnut. Agronomy 2022, 12, 404. [Google Scholar] [CrossRef]
- Puppala, N.; Nayak, S.N.; Sanz-Saez, A.; Chen, C.; Devi, M.J.; Nivedita, N.; Bao, Y.; He, G.; Traore, S.M.; Wright, D.A.; et al. Sustaining yield and nutritional quality of peanuts in harsh environments: Physiological and molecular basis of drought and heat stress tolerance. Front. Genet. 2023, 14, 1121462. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Luo, H.; Zhang, H.; Yung, W.S.; Li, M.W.; Lam, H.M.; Huang, C. Feeding the world using speed breeding technology. Trends Plant Sci. 2023, 28, 372–373. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, B.; Zhao, J.; Zhang, F.; Xie, T.; Xu, K.; Gao, G.; Yan, G.; Li, H.; Li, L.; et al. Genomic selection and genetic architecture of agronomic traits during modern rapeseed breeding. Nat. Genet. 2022, 54, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of cultivated peanut, Arachis hypogaea, yields insights into genome evolution and oil improvement. Mol. Plant 2019, 12, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.-C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef]
- Wang, J.; Yan, C.; Shi, D.; Zhao, X.; Yuan, C.; Sun, Q.; Mou, Y.; Chen, H.; Li, Y.; Li, C.; et al. The genetic base for peanut height-related traits revealed by a meta-analysis. Plants 2021, 10, 1058. [Google Scholar] [CrossRef]
- Pandey, M.K.; Agarwal, G.; Kale, S.M.; Clevenger, J.; Nayak, S.N.; Sriswathi, M.; Chitikineni, A.; Chavarro, C.; Chen, X.; Upadhyaya, H.D.; et al. Development and evaluation of a high density genotyping ‘Axiom_Arachis’ array with 58 K SNPs for accelerating genetics and breeding in groundnut. Sci. Rep. 2017, 7, 40577. [Google Scholar] [CrossRef]
- Kumar, R.; Janila, P.; Vishwakarma, M.K.; Khan, A.W.; Manohar, S.S.; Gangurde, S.S.; Variath, M.T.; Shasidhar, Y.; Pandey, M.K.; Varshney, R.K.; et al. Whole-genome resequencing-based QTL-seq identified candidate genes and molecular markers for fresh seed dormancy in groundnut. Plant Biotechnol. J. 2020, 18, 992–1003. [Google Scholar] [CrossRef] [PubMed]
- Miller, O.H.; Burns, E.E. Internal color of Spanish peanut hulls as an index of kernel maturity. J. Food Sci. 1971, 36, 666–670. [Google Scholar] [CrossRef]
- Kumar, A.S.T.; Gowda, M.V.C.; Nadaf, H.L. Seed dormancy in erect bunch genotypes of groundnut (Arachis hypogaea L.) I. Variability for intensity and duration. J. Oilseeds Res. 1991, 8, 166–172. [Google Scholar]
- Panse, V.G.; Sukhatme, P.V. Statistical Methods for Agricultural Workers; Indian Council of Agricultural Research Publication: New Delhi, India, 1985. [Google Scholar]
- Wang, M.L.; Chen, C.Y.; Pinnow, D.L.; Barkley, N.A.; Pittman, R.N.; Lamb, M.; Pederson, G.A. Seed dormancy variability in the US peanut mini-core collection. Res. J. Seed Sci. 2012, 5, 84–95. [Google Scholar] [CrossRef]
- Kumar, N.; Ajay, B.C.; Dagla, M.C.; Rathnakumar, A.L.; Radhakrishnan, T.; Lal, C.; Samdur, M.Y.; Mathur, R.K.; Manivel, P. Multi-environment evaluation of Spanish bunch groundnut genotypes for fresh seed dormancy. Indian J. Genet. 2019, 79, 571–582. [Google Scholar] [CrossRef]
- Faye, I.; Ndoye, O.; Diop, T.A. Evaluation of fresh seed dormancy on seven peanut (Arachis hypogaea L.) lines derived from crosses between Spanish varieties: Variability on intensity and duration. J. Appl. Sci. Res. 2009, 5, 853–857. [Google Scholar]
- Kumar, N.; Ajay, B.C.; Rathanakumar, A.L.; Radhakrishnan, T.; Lal, C.; Samdur, M.Y.; Mathur, R.K.; Manivel, P.; Chikani, B.M. Genetic variability for fresh seed dormancy in Spanish bunch advanced breeding lines of groundnut (Arachis hypogaea L.). J. Oilseeds Res. 2017, 34, 119–124. [Google Scholar]
- Kumar, N.; Ajay, B.C.; Dagla, M.C.; Rathanakumar, A.L.; Lal, C.; Radhakrishnan, T.; Samdur, M.Y.; Mathur, R.K.; Manivel, P.; Makwana, A.D.; et al. Evaluation of Spanish bunch advanced breeding lines for fresh seed dormancy in groundnut (Arachis hypogaea L.). Front. Crop Improv. 2018, 6, 9–12. [Google Scholar]
- Kumar, N.; Ajay, B.C.; Dagla, M.C.; Rathanakumar, A.L.; Radhakrishnan, T.; Samdur, M.Y.; Mathur, R.K.; Manivel, P.; Lal, C.; Makwana, A.D.; et al. Identification of Spanish bunch advanced breeding lines having fresh seed dormancy in groundnut (Arachis hypogaea L.). Progressive Res. Int. J. 2018, 13, 19–23. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Traits | Source | Df | Sum Sq | Mean Sq | F Value | Pr(>F) | Significance |
|---|---|---|---|---|---|---|---|
| Days to 50% germination | Genotype | 183 | 107,874 | 589.5 | 18.299 | <2 × 10−16 | *** |
| Replication | 1 | 12 | 12.1 | 0.376 | 0.5398 | ||
| Season | 1 | 118 | 117.7 | 3.652 | 0.0565 | ||
| Residuals | 535 | 17,234 | 32.2 | ||||
| Germination Percentage | Genotype | 183 | 946,243 | 5171 | 13.382 | <2 × 10−16 | *** |
| Replication | 1 | 41 | 41 | 0.105 | 0.746 | ||
| Season | 1 | 4735 | 4735 | 12.255 | 0.000503 | *** | |
| Residuals | 535 | 206,716 | 206,716 | ||||
| Intensity of dormancy | Genotype | 183 | 946,243 | 5171 | 13.382 | <2 × 10−16 | *** |
| Replication | 1 | 41 | 41 | 0.105 | 0.746 | ||
| Season | 1 | 4735 | 4735 | 12.255 | 0.000503 | *** | |
| Residuals | 535 | 206,716 | 206,716 |
| Gene ID | SNP_Position | Function | F/R | Allele-Specific Primer Sequence | Length (bp) |
|---|---|---|---|---|---|
| Aradu.D94AQ.1 | A09_115175289 | Zeaxanthin epoxidase | F | CACATCTTCTAGTGAAAGCGGA | 22 |
| R | CCATCTTTCTGATGGAACAACC | 22 | |||
| Intergenic | B05_111598196 | F | TTTTCCTTAATATTTGAAAAATTATCCTAA | 30 | |
| R | CGTCTTTGCCAATTGTGTATAAC | 23 |
| Rainy 2022 | Post Rainy 2022–2023 | |||||||
|---|---|---|---|---|---|---|---|---|
| S. No. | Accessions | Agronomic Type | DG | GP | DI | DG | GP | DI |
| 1 | ICG 5827 | Virginia Runner | 30 | 0 | 100 | 23 | 35 | 90 |
| 2 | ICG 7000 | Virginia Bunch | 30 | 10 | 100 | 22 | 35 | 100 |
| 3 | ICG 9809 | Spanish Bunch | 16 | 35 | 65 | 25 | 55 | 95 |
| 4 | ICG 11322 | Virginia Bunch | 30 | 15 | 95 | 23 | 35 | 100 |
| 5 | ICG 11457 | Virginia Runner | 23 | 30 | 80 | 30 | 0 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bomireddy, D.; Sharma, V.; Senthil, R.; Reddisekhar, M.; Shah, P.; Singh, K.; Reddy, D.M.; Sudhakar, P.; Reddy, B.V.B.; Pandey, M.K. Identification of Donors for Fresh Seed Dormancy and Marker Validation in a Diverse Groundnut Mini-Core Collection. Agronomy 2024, 14, 112. https://doi.org/10.3390/agronomy14010112
Bomireddy D, Sharma V, Senthil R, Reddisekhar M, Shah P, Singh K, Reddy DM, Sudhakar P, Reddy BVB, Pandey MK. Identification of Donors for Fresh Seed Dormancy and Marker Validation in a Diverse Groundnut Mini-Core Collection. Agronomy. 2024; 14(1):112. https://doi.org/10.3390/agronomy14010112
Chicago/Turabian StyleBomireddy, Deekshitha, Vinay Sharma, Ramachandran Senthil, Mangala Reddisekhar, Priya Shah, Kuldeep Singh, Devarapalli Mohan Reddy, Palagiri Sudhakar, Bommu Veera Bhaskara Reddy, and Manish K. Pandey. 2024. "Identification of Donors for Fresh Seed Dormancy and Marker Validation in a Diverse Groundnut Mini-Core Collection" Agronomy 14, no. 1: 112. https://doi.org/10.3390/agronomy14010112
APA StyleBomireddy, D., Sharma, V., Senthil, R., Reddisekhar, M., Shah, P., Singh, K., Reddy, D. M., Sudhakar, P., Reddy, B. V. B., & Pandey, M. K. (2024). Identification of Donors for Fresh Seed Dormancy and Marker Validation in a Diverse Groundnut Mini-Core Collection. Agronomy, 14(1), 112. https://doi.org/10.3390/agronomy14010112