
Determination of Genetic Diversity of Some Upland and Sea Island Cotton Genotypes Using High-Resolution Capillary Electrophoresis Gel

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Isolation
2.3. PCR Amplification and SSR Markers
2.4. Detection of PCR Amplicons
2.5. Data Analysis
3. Results
3.1. Polymorphism Analysis of EST-SSR Markers
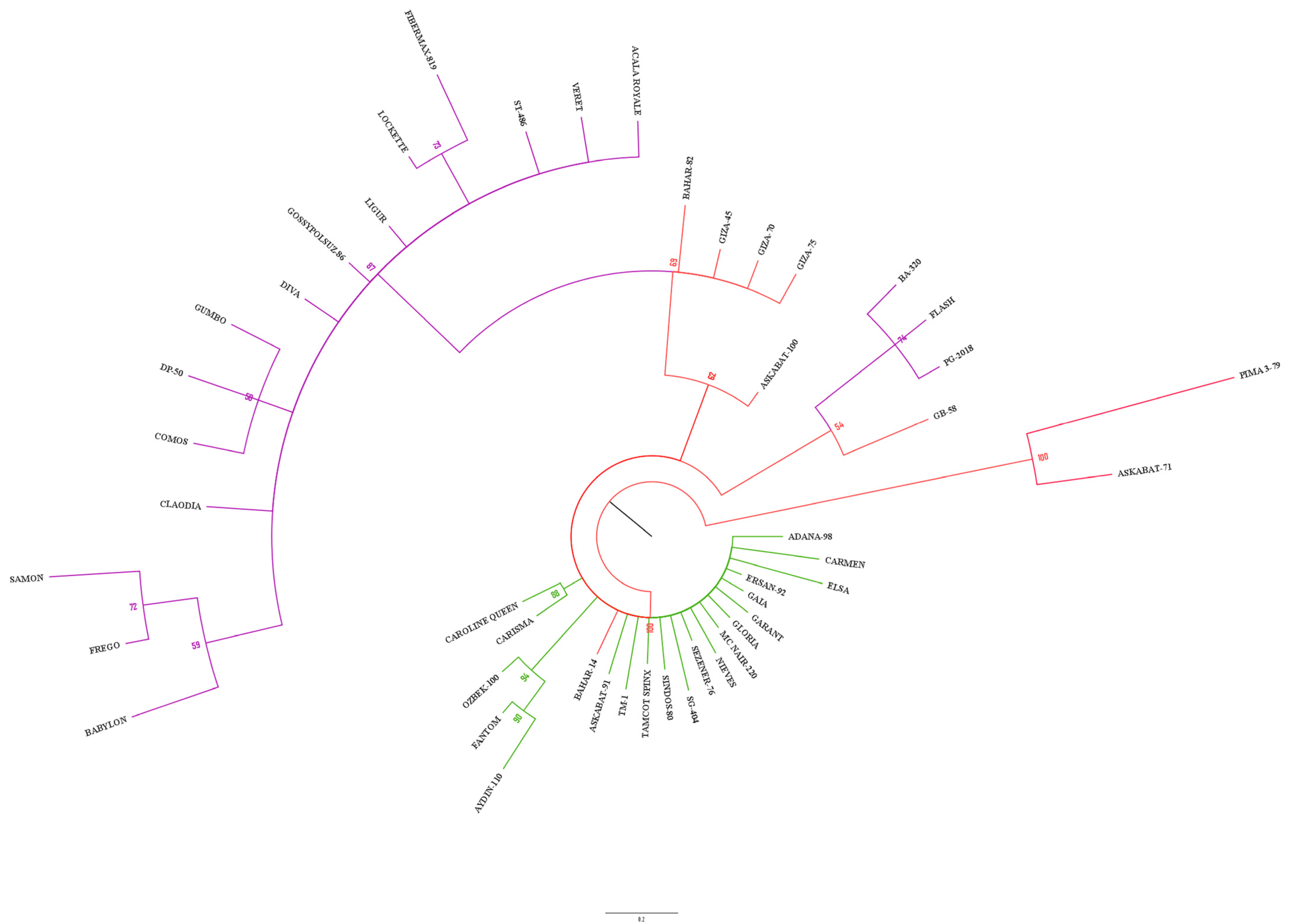
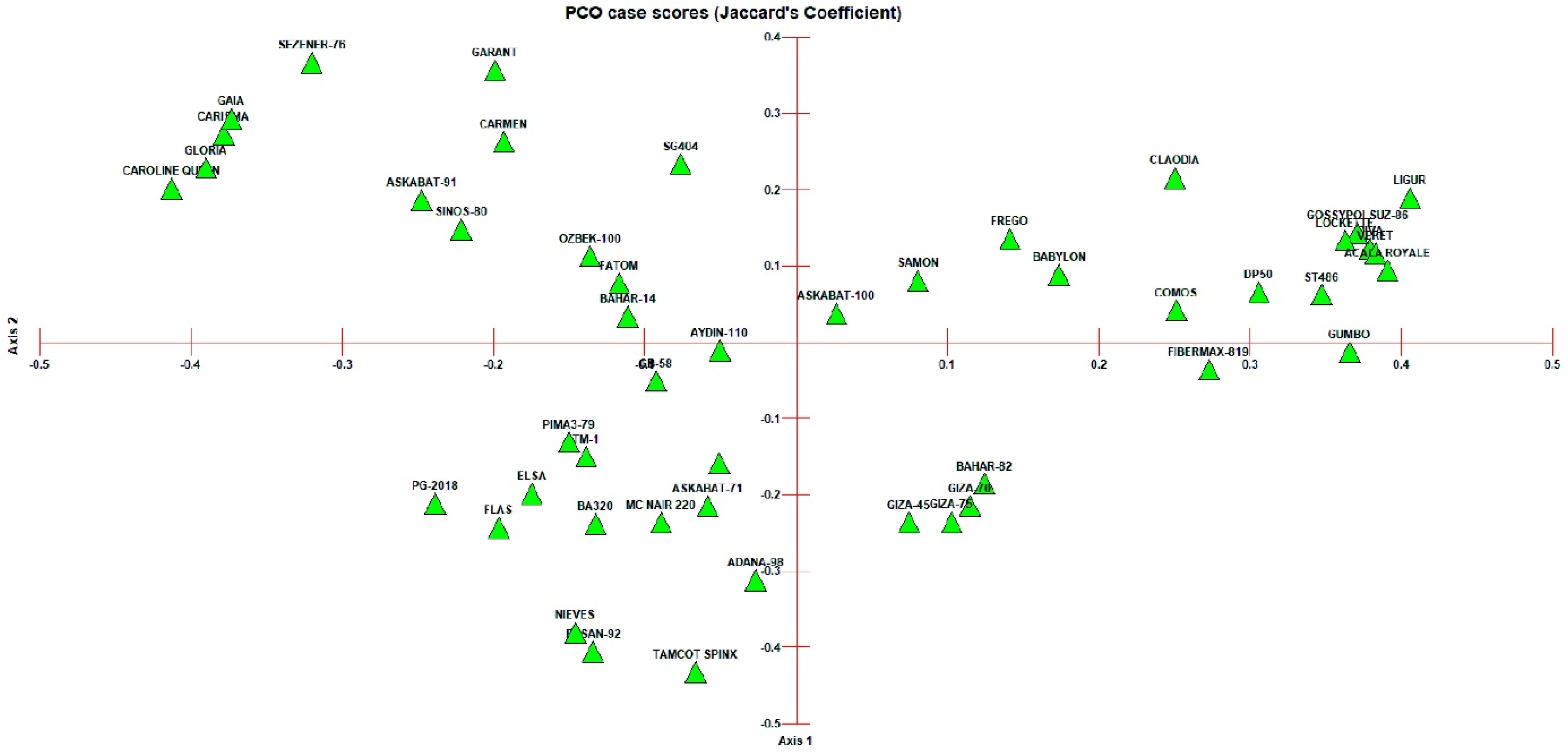
3.2. Clustering and PCoA Analysis
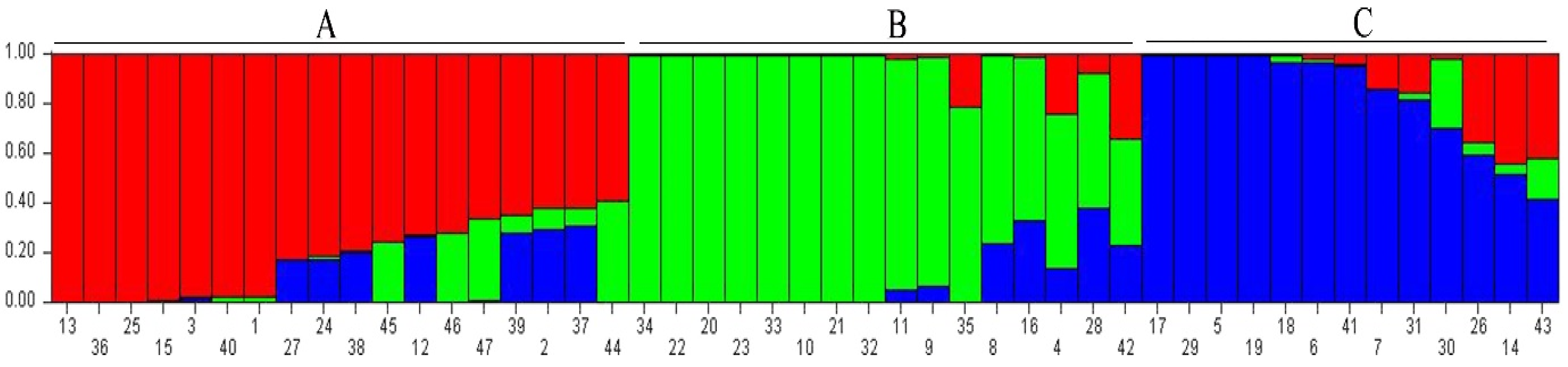
3.3. Population Structure Analysis
4. Discussion
4.1. Effects of Molecular Markers on Population
4.2. Cluster Analysis and Population Structure
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, K.; Wendel, J.F.; Hua, J. Designations for individual genomes and chromosomes in Gossypium. J. Cotton Res. 2018, 1, 3. [Google Scholar] [CrossRef]
- Grover, C.E.; Zhu, X.; Grupp, K.K.; Jareczek, J.J.; Gallagher, J.P.; Szadkowski, E.; Seijo, J.G.; Wendel, J.F. Molecular confirmation of species status for the allopolyploid cotton species, Gossypium ekmanianum Wittmack. Genet. Resour. Crop Evol. 2015, 62, 103–114. [Google Scholar] [CrossRef]
- Anwar, K.; Joshi, R.; Dhankher, O.P.; Singla-Pareek, S.L.; Pareek, A. Elucidating the response of crop plants towards individual, combined and sequentially occurring abiotic stresses. Int. J. Mol. Sci. 2021, 22, 6119. [Google Scholar] [CrossRef] [PubMed]
- Witt, T.W.; Ulloa, M.; Schwartz, R.C.; Ritchie, G.L. Response to deficit irrigation of morphological, yield and fiber quality traits of upland (Gossypium hirsutum L.) and Pima (G. barbadense L.) cotton in the Texas High Plains. Field Crops Res. 2020, 249, 107759. [Google Scholar] [CrossRef]
- Aydın, A. A Study on the Molecular Characterization of Some Commercial Cotton Varieties Registered in Turkey. Ph.D. Dissertation, Akdeniz University, Antalya, Turkey, 2018. [Google Scholar]
- Karaca, M.; Ince, A.G. New non-redundant microsatellite and CAPS-microsatellite markers for cotton (Gossypium L.). Turk. J. Field Crops 2011, 16, 172–178. [Google Scholar]
- Kumlay, A.M.; Eren, B.; Demirel, S.; Demirel, F.; Yıldırım, B. Reveal of Genetic Variation by iPBS Analysis in Some Cotton Varieties. Eur. J. Eng. Sci. Tech. 2021, 21, 67–73. [Google Scholar] [CrossRef]
- Rashid, U.; Anwar, F.; Knothe, G. Evaluation of biodiesel obtained from cottonseed oil. Fuel Process. Technol. 2009, 90, 1157–1163. [Google Scholar] [CrossRef]
- Sekhar, S.C.; Rao, B.V.K. Cottonseed oil as health oil. Pertanika J. Trop. Agric. Sci. 2011, 34, 17–24. [Google Scholar]
- Guo, Q.; Liu, J.; Li, J.; Cao, S.; Zhang, Z.; Zhang, J.; Zhang, Y.; Deng, Y.; Niu, D.; Su, L.; et al. Genetic diversity and core collection extraction of Robinia pseudoacacia L. germplasm resources based on phenotype, physiology, and genotyping markers. Ind. Crops Prod. 2022, 178, 114627. [Google Scholar] [CrossRef]
- Piasecka, A.; Kachlicki, P.; Stobiecki, M. Analytical methods for detection of plant metabolomes changes in response to biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 379. [Google Scholar] [CrossRef]
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential mechanisms of abiotic stress tolerance in crop plants induced by thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Swarup, S.; Cargill, E.J.; Crosby, K.; Flagel, L.; Kniskern, J.; Glenn, K.C. Genetic diversity is indispensable for plant breeding to improve crops. Crop Sci. 2021, 61, 839–852. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, Y.; Wang, N.; Xu, Q.; Huo, F.; Liu, X.; Yan, G. Genetic diversity and population structure analysis of upland cotton (Gossypium hirsutum L.) germplasm in China based on SSR markers. Genet. Resour. Crop Evol. 2023. [Google Scholar] [CrossRef]
- Ghuge, S.B.; Mehetre, S.S.; Chimote, V.P.; Pawar, B.D.; Naik, R.M. Molecular characterization of cotton genotypes using SSR, ISSR and RAPD markers in relation to fiber quality traits. J. Cotton Res. 2018, 32, 1–12. [Google Scholar]
- Kamaluddin; Khan, M.A.; Kiran, U.; Ali, A.; Abdin, M.Z.; Zargar, M.Y.; Ahmad, S.; Sofi, P.A.; Gulzar, S. Molecular Markers and Marker-Assisted Selection in Crop Plants. In Plant Biotechnology: Principles and Applications; Abdin, M., Kiran, U., Kamaluddin, Ali, A., Eds.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Tyagi, P.; Gore, M.A.; Bowman, D.T.; Campbell, B.T.; Udall, J.A.; Kuraparthy, V. Genetic diversity and population structure in the US Upland cotton (Gossypium hirsutum L.). Theor. Appl. Genet. 2014, 127, 283–295. [Google Scholar] [CrossRef]
- Moiana, L.D.; Filho, P.S.V.; Gonçalves-Vidigal, M.C.; de Carvalho, L.P. Genetic diversity and population structure of upland cotton Brazilian cultivars (Gossypium hirsutum L. race latifolium H.) using SSR markers. Aust. J. Crop Sci. 2015, 9, 143–152. [Google Scholar]
- Morales, N.; Ogbonna, A.C.; Ellerbrock, B.J.; Bauchet, G.J.; Tantikanjana, T.; Tecle, I.Y.; Powell, A.F.; Lyon, D.; Menda, N.; Simoes, C.C.; et al. Breedbase: A digital ecosystem for modern plant breeding. G3 2022, 12, jkac078. [Google Scholar] [CrossRef]
- Younis, A.; Ramzan, F.; Ramzan, Y.; Zulfiqar, F.; Ahsan, M.; Lim, K.B. Molecular markers improve abiotic stress tolerance in crops: A review. Plants 2020, 9, 1374. [Google Scholar] [CrossRef]
- Fu, Y.B. Understanding crop genetic diversity under modern plant breeding. Theor. Appl. Genet. 2015, 128, 2131–2142. [Google Scholar] [CrossRef]
- Kumar, S.J.; Susmita, C.; Sripathy, K.V.; Agarwal, D.K.; Pal, G.; Singh, A.N.; Kumar, S.; Rai, A.K.; Simal-Gandara, J. Molecular characterization and genetic diversity studies of Indian soybean (Glycine max (L.) Merr.) cultivars using SSR markers. Mol. Biol. Rep. 2022, 49, 2129–2140. [Google Scholar] [CrossRef]
- Çelik, S. Genetic Diversity Analysis of Some Upland Cotton (Gossypium hirsutum L.) Genotypes Using SSR Markers. TJNS 2022, 11, 80–89. [Google Scholar]
- Zhao, X.; Zhang, Q.; Tang, X.; Xu, Y.; Du, Y.; Zhao, B.; Fan, X.; Zhang, X.; Zhang, W.; Wu, W.; et al. Multiplex detection of transgenic maize by microdroplet PCR combined with capillary gel electrophoresis. Acta Biochim. Biophys. Sin. 2019, 51, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Xia, J.; Gong, J.; Yang, S. Application of capillary electrophoresis detection with fluorescent SSR markers in rice DNA fingerprint identification. Chin. J. Rice Sci. 2011, 25, 672–676. [Google Scholar]
- Wang, Q.; Fang, L.; Chen, J.; Hu, Y.; Si, Z.; Wang, S.; Chang, L.; Guo, W.; Zhang, T. Genome-wide mining, characterization and development of microsatellite markers in Gossypium species. Sci. Rep. 2015, 5, 10638. [Google Scholar] [CrossRef]
- Ince, A.G.; Karaca, M. Species-specific touch-down DAMD-PCR markers for Salvia species. J. Med. Plants Res. 2012, 6, 1590–1595. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Eren, B.; Keskin, B.; Demirel, F.; Demirel, S.; Türkoğlu, A.; Yilmaz, A.; Haliloğlu, K. Assessment of genetic diversity and population structure in local alfalfa genotypes using iPBS molecular markers. Genet. Resour. Crop Evol. 2023, 70, 617–628. [Google Scholar] [CrossRef]
- Demirel, S.; Demirel, F. Molecular identification and population structure of emmer and einkorn wheat lines with different ploidy levels using SSR markers. Genet. Resour. Crop Evol. 2023. [Google Scholar] [CrossRef]
- Karaca, M.; Ince, A.G.; Aydin, A.; Elmasulu, S.Y.; Turgut, K. Microsatellites for genetic and taxonomic research on thyme (Thymus L.). Turk. J. Biol. 2015, 39, 147–159. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 261–285. [Google Scholar] [CrossRef]
- Sabev, P.; Valkova, N.; Todorovska, E.G. Molecular markers and their application in cotton breeding: Progress and future perspectives. Bulg. J. Agric. Sci. 2020, 26, 816–828. [Google Scholar]
- Mishra, N.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Gupta, N.; Sharma, A.; Solanki, R.S.; Tiwari, S. Characterization of soybean genotypes on the basis of yield attributing traits and SSR molecular markers. Innov. Sci. Technol. 2022, 3, 87–106. [Google Scholar]
- Cardle, L.; Ramsay, L.; Milbourne, D.; Macaulay, M.; Marshall, D.; Waugh, R. Computational and experimental characterization of physically clustered simple sequence repeats in plants. Genetics 2000, 156, 847–854. [Google Scholar] [CrossRef]
- Saha, S.; Karaca, M.; Jenkins, J.N.; Zipf, A.E.; Reddy, O.U.K.; Kantety, R.V. Simple sequence repeats as useful resources to study transcribed genes of cotton. Euphytica 2003, 130, 355–364. [Google Scholar] [CrossRef]
- Qureshi, S.N.; Saha, S.; Kantety, R.V.; Jenkins, J.N. EST-SSR: A new class of genetic markers in cotton. J. Cotton Sci. 2004, 8, 112–123. [Google Scholar]
- Karaca, M.; Ince, A.G.; Aydin, A.; Ay, S.T. Cross-genera transferable e-microsatellite markers for 12 genera of the Lamiaceae family. J. Sci. Food Agric. 2013, 93, 1869–1879. [Google Scholar] [CrossRef]
- Noormohammadi, Z.; Rahnama, A.; Sheidai, M. EST-SSR and SSR analyses of genetic diversity in diploid cotton genotypes from Iran. Nucleus 2013, 56, 171–178. [Google Scholar] [CrossRef]
- Saleem, M.A.; Amjid, M.W.; Ahmad, M.Q.; Riaz, H.; Arshad, S.F.; Zia, Z.U. EST-SSR based analysis revealed narrow genetic base of in-use cotton varieties of Pakistan. Pak. J. Bot. 2020, 52, 1667–1672. [Google Scholar] [CrossRef]
- Hinchliffe, D.J.; Turley, R.B.; Naoumkina, M.; Kim, H.J.; Tang, Y.; Yeater, K.M.; Li, P.; Fang, D.D. A combined functional and structural genomics approach identified an EST-SSR marker with complete linkage to the Ligon lintless-2 genetic locus in cotton (Gossypium hirsutum L.). BMC Genom. 2011, 12, 445. [Google Scholar] [CrossRef]
- Wang, B.H.; Zhu, P.; Yuan, Y.L.; Wang, C.B.; Yu, C.M.; Zhang, H.H.; Zhu, X.Y.; Wang, W.; Yao, C.B.; Zhuang, Z.M.; et al. Development of EST-SSR markers related to salt tolerance and their application in genetic diversity and evolution analysis in Gossypium. Genet. Mol. Res. 2014, 13, 3732–3746. [Google Scholar] [CrossRef]
- Han, Z.G.; Guo, W.Z.; Song, X.L.; Zhang, T.Z. Genetic mapping of EST-derived microsatellites from the diploid Gossypium arboreum in allotetraploid cotton. Mol. Genet. Genom. 2004, 272, 308–327. [Google Scholar] [CrossRef]
- Han, Z.; Wang, C.; Song, X.; Guo, W.; Gou, J.; Li, C.; Chen, X.; Zhang, T. Characteristics, development and mapping of Gossypium hirsutum derived EST-SSRs in allotetraploid cotton. Theor. Appl. Genet. 2006, 112, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pérez, R.; Ballester, J.; Dicenta, F.; Arús, P.; Martínez-Gómez, P. Comparison of SSR polymorphisms using automated capillary sequencers, and polyacrylamide and agarose gel electrophoresis: Implications for the assessment of genetic diversity and relatedness in almond. Sci. Hortic. 2006, 108, 310–316. [Google Scholar] [CrossRef]
- McCudden, C.R.; Mathews, S.P.; Hainsworth, S.A.; Chapman, J.F.; Hammett-Stabler, C.A.; Willis, M.S.; Grenache, D.G. Performance comparison of capillary and agarose gel electrophoresis for the identification and characterization of monoclonal immunoglobulins. Am. J. Clin. Pathol. 2008, 129, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.F.; Brubaker, C.L.; Seelanan, T. The origin and evolution of Gossypium. In Physiology of Cotton; Springer: Dordrecht, The Netherlands, 2010; pp. 1–18. [Google Scholar]
- Wendel, J.F. New World tetraploid cottons contain Old World cytoplasm. Proc. Natl. Acad. Sci. USA 1989, 86, 4132–4136. [Google Scholar] [CrossRef]
- Rana, M.K.; Bhat, K.V. RAPD markers for genetic diversity study among Indian cotton cultivars. Curr. Sci. 2005, 88, 1956–1961. [Google Scholar]
- Abdellatif, K.F.; Soliman, Y.A. Genetic relationships of cotton (Gossypium barbadense L.) genotypes as studied by morphological and molecular markers. Afr. J. Biotechnol. 2013, 12, 4736–4746. [Google Scholar]
- Islam, A.A.; Era, F.M.; Khalequzzaman, M.F.; Chakrabarty, S. Estimation of heterosis in hybrids of upland cotton (Gossypium hirsutum L.) for seed cotton yield and related traits. JAPS 2021, 1, 1–12. [Google Scholar]
- Zhang, J.; Percy, R.G.; McCarty, J.C. Introgression genetics and breeding between Upland and Pima cotton: A review. Euphytica 2014, 198, 1–12. [Google Scholar] [CrossRef]
- Li, Y.; Si, Z.; Wang, G.; Shi, Z.; Chen, J.; Qi, G.; Jin, S.; Han, Z.; Gao, W.; Tian, Y.; et al. Genomic insights into the genetic basis of cotton breeding in China. Mol. Plant 2023, 16, 662–677. [Google Scholar] [CrossRef] [PubMed]
- Mehboob-ur-Rahman; Shaheen, T.; Tabbasam, N.; Iqbal, M.A.; Ashraf, M.; Zafar, Y.; Paterson, A.H. Cotton genetic resources. A review. Agron. Sustain. Dev. 2012, 32, 419–432. [Google Scholar]
- Stelly, D.M.; Saha, S.; Raska, D.A.; Jenkins, J.N. Registration of 17 upland (Gossypium hirsutum) cotton germplasm lines disomic for different G. barbadense chromosome or arm substitutions. Crop Sci. 2005, 45, 2663. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Genotype | Species |
|---|---|---|
| 1 | Adana-98 | G. hirsutum |
| 2 | Aydın-110 | G. hirsutum |
| 3 | BA-320 | G. hirsutum |
| 4 | Babylon | G. hirsutum |
| 5 | Carisma | G. hirsutum |
| 6 | Carmen | G. hirsutum |
| 7 | Caroline Queen | G. hirsutum |
| 8 | Claodia | G. hirsutum |
| 9 | Comos | G. hirsutum |
| 10 | Diva | G. hirsutum |
| 11 | DP-50 | G. hirsutum |
| 12 | Elsa | G. hirsutum |
| 13 | Erşan-92 | G. hirsutum |
| 14 | Fantom | G. hirsutum |
| 15 | Flash | G. hirsutum |
| 16 | Frego | G. hirsutum |
| 17 | Gaia | G. hirsutum |
| 18 | Garant | G. hirsutum |
| 19 | Gloria | G. hirsutum |
| 20 | Gossypolsüz 86 | G. hirsutum |
| 21 | Gumbo | G. hirsutum |
| 22 | Ligur | G. hirsutum |
| 23 | Lockette | G. hirsutum |
| 24 | Mc Nair 220 | G. hirsutum |
| 25 | Nieves | G. hirsutum |
| 26 | Özbek-100 | G. hirsutum |
| 27 | PG -2018 | G. hirsutum |
| 28 | Samon | G. hirsutum |
| 29 | Sezener-76 | G. hirsutum |
| 30 | SG-404 | G. hirsutum |
| 31 | Sindos-80 | G. hirsutum |
| 32 | ST-468 | G. hirsutum |
| 33 | Veret | G. hirsutum |
| 34 | Acala Royale | G. hirsutum |
| 35 | Fibermax-819 | G. hirsutum |
| 36 | Tamcot Spinx | G. hirsutum |
| 37 | TM-1 | G. hirsutum |
| 38 | Pima 3–79 | G. barbadense |
| 39 | G.B-58 | G. barbadense |
| 40 | Askabat-71 | G. barbadense |
| 41 | Askabat-91 | G. barbadense |
| 42 | Askabat-100 | G. barbadense |
| 43 | Bahar-14 | G. barbadense |
| 44 | Bahar-82 | G. barbadense |
| 45 | Giza-45 | G. barbadense |
| 46 | Giza-70 | G. barbadense |
| 47 | Giza-75 | G. barbadense |
| Primer ID | Primer Sequence | Motif | Allele Number | Tm (°C) | He | H0 | PIC |
|---|---|---|---|---|---|---|---|
| GA01-2651 | F:AAATCCTACCTCTCCGGCCA R: CCCAGGGCAAAACAATGTCG | (GCCGGC)3 | 6 | 60 | 0.485 | 0.479 | 0.475 |
| GA02-54 | F:GGGAAAGCGCGTCATTGATC R: GCCGAGCCCAGACCTAATAG | (GCCGG)4 | 4 | 60 | 0.450 | 0.096 | 0.437 |
| GA04-1418 | F:GCAGGCAGAATACAAAAGATCGA R: AAGAAAAGGGGGAGGGGAGA | (GCCGGC)3 | 33 | 60 | 0.737 | 0.596 | 0.730 |
| GA07-410 | F:GAAATTACCTTTCCGGCCACC R: GACGTCGTTTTGGAGGGCTA | (GCCGGC)3 | 24 | 60 | 0.720 | 0.528 | 0.708 |
| GA08-323 | F:GAACCGACCTAAGGTGACTGT R: AGAGAGAAGGGAGGGGGAAG | (CCGGCG)3 | 6 | 60 | 0.476 | 0.266 | 0.473 |
| GD01-295 | F:TTTACCTTTCCGACCACCGC R: GGTGCGTTTTGGTCCCCTAT | (GCCGGC)3 | 7 | 60 | 0.704 | 0.415 | 0.693 |
| GD02-301 | F:AAACCCGTTGTGCAACCATG R: GGATGAGGCTGAGAAGGAGC | (CCGGTG)3 | 15 | 60 | 0.898 | 0.883 | 0.889 |
| GD03-2002 | F:GGCCCGGCCCGAATATAATA R: GACTAGACCTGTCCATGGGC | (CCGGGC)3 | 19 | 60 | 0.798 | 0.539 | 0.788 |
| GD06-2808 | F:GGCCCGGCCCGAATATAATAA R: CGGCCCGAAATATGGGCTTA | (CCGGGC)3 | 14 | 60 | 0.625 | 0.560 | 0.616 |
| GD08-420 | F:AGAGAAGGGAGGGGGAAAGG R: GGGCTCTAACACCAAATCGGA | (CGCCGG)3 | 42 | 60 | 0.741 | 0.571 | 0.738 |
| GD09-1296 | F:GGCGCACAAAACACCAAGAT R: AGGGAGGAAGGAAAGGGGG | (GCCGGC)3 | 14 | 60 | 0.745 | 0.546 | 0.737 |
| GD10-1664 | F:GGGCTCTAACACCAAATCGGA R: AGAGAAGGGAGGGGGAAAGG | (GCCGGC)3 | 29 | 60 | 0.679 | 0.493 | 0.675 |
| MK086 | F:CCACCAGTTTGGTAGGTATGAAC R: TCAACAGTGCAAGGACTTCATC | (CAT)8 | 2 | 66 | 0.293 | 0.121 | 0.268 |
| MK105 | F:CAAAGATGCCGAAAGAGAGG R: GTAAGATCGGCGGGTCATC | (CCG)12 | 30 | 66 | 0.666 | 0.319 | 0.659 |
| MK126 | F:ACCGTACCCGTGGCTCTTAT R: TGTTGTTGTGGGAGGCTTCT | (CAT)8 | 4 | 66 | 0.414 | 0.043 | 0.378 |
| MK129 | F:GCTGATGCTGATTCCTCCAT R: TGCCCTTCATCTCGTTTCTT | (CAA)8 | 7 | 66 | 0.373 | 0.479 | 0.345 |
| MK132 | F:AGCAAGGCATGAGCGATACT R: GGTGGTACCTTCCCATGTTG | (TCAGCC)6 | 8 | 66 | 0.541 | 0.391 | 0.511 |
| MK146 | F:ATGGAGGCTGCAAAGACTGT R: CCACTCCGACTAAAAGATCAGC | (GTAGTGAGA)3 | 3 | 66 | 0.577 | 0.787 | 0.487 |
| MK173 | F:GGGGTCCACAGATACAGG R: GTCCAAAACTTGTCCCATTAG | (TATG)9 | 13 | 66 | 0.867 | 0.106 | 0.858 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aydin, A. Determination of Genetic Diversity of Some Upland and Sea Island Cotton Genotypes Using High-Resolution Capillary Electrophoresis Gel. Agronomy 2023, 13, 2407. https://doi.org/10.3390/agronomy13092407
Aydin A. Determination of Genetic Diversity of Some Upland and Sea Island Cotton Genotypes Using High-Resolution Capillary Electrophoresis Gel. Agronomy. 2023; 13(9):2407. https://doi.org/10.3390/agronomy13092407
Chicago/Turabian StyleAydin, Adnan. 2023. "Determination of Genetic Diversity of Some Upland and Sea Island Cotton Genotypes Using High-Resolution Capillary Electrophoresis Gel" Agronomy 13, no. 9: 2407. https://doi.org/10.3390/agronomy13092407
APA StyleAydin, A. (2023). Determination of Genetic Diversity of Some Upland and Sea Island Cotton Genotypes Using High-Resolution Capillary Electrophoresis Gel. Agronomy, 13(9), 2407. https://doi.org/10.3390/agronomy13092407