Abstract
Weeds pose major constraints in crop production. The use of allelochemicals and allelopathic species can provide an effective alternative for sustainable weed management. In a previous study that evaluated the allelopathic activity of wild and cultivated plants in Turkey, Hibiscus sabdariffa demonstrated the strongest inhibitory potential. This study aimed to estimate the phytotoxic influence of the H. sabdariffa water crude extracts on Lactuca sativa L. in a bioassay experiment. High-performance liquid chromatography (HPLC) analysis was used to identify two major compounds, hydroxycitric acid lactone and hydroxy citric acid, and their plant growth inhibitory activities were evaluated by bioassays. Hydroxycitric acid lactone had a stronger growth inhibitory activity on L. sativa L. and was estimated as a major allelochemical in H. sabdariffa calyx. The high concentration (16.7% of the dry weight of the calyx) and strong inhibitory effect (EC50, 73.7 ppm) of the hydroxycitric acid lactone could demonstrate the growth inhibitory activity of the H. sabdariffa calyx extract. This study showed that hydroxycitric acid lactone, a major compound in the calyx of Hibiscus sabdariffa, is a plant growth inhibitor.
1. Introduction
Weeds are a major impediment to the growth and development of field crops, often leading to yield losses [1,2]. The annual global economic loss caused by weeds has been estimated at more than 100 billion US dollars [3]; moreover, the worldwide expenditure on weed management is expected to reach billions of US dollars [4]. Farmers have struggled with weeds intensively since the dawn of agriculture [5], and the control of weeds, therefore, poses a challenging issue for sustainable agriculture [6,7]. Since the discovery of synthetic herbicides, they have become among the most widely used weed-control strategies in most countries [8,9]. Over two hundred functional substances had been recorded for herbicidal applications [9,10]. However, weeds could develop resistance to synthetic herbicides [9].
Allelopathy is a biological occurrence for several plant species, in which plants release bioactive compounds into the surrounding habitat either from their aerial or underground segments through root exudates, leaching by dews and rains, and vaporizing or decomposing plant tissue. The released compounds could influence the growth and development of other organisms, in particular weeds, other plants, animals, and microorganisms, in an inhibitory or stimulatory way in the environment [11,12]. These plant growth inhibitory compounds (allelochemicals) from various donor plant species could act as natural weed inhibitors. Different parts of plants, such as leaves, barks, roots, flowers, seeds, pollens, stems, and fruits, could contain growth-inhibitory compounds [13,14]. The concentration of the inhibitory compounds in the different plant parts may vary, leading to variation in the level of plant growth inhibitory effects [13,15]. Previous studies have shown the effective use of the species containing inhibitory substances and their growth inhibitory compounds against weeds in good plant protection practices [16,17,18,19,20].
Natural products are made by syntheses in alive organisms as their answer to biological or physical incentives. They are usually built via complicated pathways leading to molecules not easily obtainable with ordinary synthetic methods in the laboratory [21,22]. These various motives make natural products a favorable material for the development of replacements for long-establish synthetic agrochemicals [23].
The use of artificially produced herbicides all the time does not point out a fitting implement to fight against resistance-developing weeds but is also asserted as harmful to the environment [24]. Moreover, the application of the species containing inhibitory substances and their growth inhibitory compounds against weeds offers alternatives for the development of ecologically friendly agricultural practices to increase crop productivity and maintain ecosystem stability [25]. Plants with growth-inhibitory properties are thought of as potential sources of biological materials against weeds [26], and growth-inhibitory compounds could be utilized for weed control to lessen the massive reliance on chemically synthesized herbicides and the corresponding toxicity danger for nature [27]. Consequently, studies for other new reliable weed-control practices have become essential, with growing attention paid to the mechanism of plant growth inhibition [28]. The effective use of plant growth inhibitory mechanisms has played a central role in weed control, and plant extracts have the potential to be exploited as natural herbicides [28]. Additionally, the action of taking out inhibitory compounds from growth-inhibitory plant materials by extraction would be applicable to produce bioherbicides [20].
The visible changes that inhibitory compounds cause to the growth and development of target plants, which are also called receptor or susceptible plants, are diverse. These effects on receptor plants could consist of decreasing radicle and shoot length, becoming darker and/or blowing up seeds, inhibited or retarded germination rate, the spiral shaping of the root axis, color defects on seeds, the absence of root hairs, the localized death of living tissues (necrosis), the growing total quantity of seminal roots, and the lessening in the amount of dry weight accumulation, for example [29,30]. These morphological effects are in many cases secondary actions of primary differentiation (the inhibition of cell division and elongation, interference with cell membrane permeability, enzymatic activities, respiration, photosynthesis, and so on) at the cellular or molecular level in susceptible plants [29].
Hibiscus sabdariffa L. is a member of the Malvaceae family of plants, and it is often recognized as Roselle. The plant is an all-around living annual plant in the tropics and subtropics of both hemispheres and many areas of Central and West Africa, Southeast Asia, America, and elsewhere as it is relatively easy to grow. The red calyx of the plant is used in several ways, including in herbal teas, herbal medicines, syrups, and food coloring [31]. H. sabdariffa calyx had shown strong growth inhibitory activity on lettuce seedling elongation; it reduced the lettuce radicle and hypocotyl elongation to 3.2% and 6.6% of the control, respectively, in previous research [32]. The chemical composition of H. sabdariffa has been described as consisting of hydroxycitric acid and its lactone in the aqueous extracts [31,33,34,35,36,37]. Although the whole plant (leaves, stem, and roots) and isolated chemicals from the whole plant indicated strong inhibitory activity on the growth of test plant species [38,39,40,41], the inhibition effect of the calyx alone and its substances has not been researched. (2S,3R)-hydroxycitric acid has been shown to prevent the action of pancreatic-amylase and small intestine glucosidase [34]. H. sabdariffa is implemented in traditional medicine in the form of herbal teas or cold beverages for its hypotensive and diuretic effects and to drop body temperature and blood viscosity [36]. H. sabdariffa dried calyx ethanolic extract in addition to the diet of rats showed a reduction in serum lipids on their serum lipid profile [42].
Hydroxycitric acid (Figure 1a) is a citric acid molecule with a hydroxyl group at the second carbon [35]. This acid has four stereoisomers: (2S,3S), (2R,3R), (2S,3R), and (2R,3S), and their lactone forms [37]. In general solution, hydroxycitric acid is a mixture of non-lactone and lactone forms. (2S,3S)-hydroxycitric acid (a mixture of (2S,3S)-hydroxycitric acid stereoisomer and its lactone form) is found in Garcinia cambogia. (2S,3R)-hydroxycitric acid (a mixture of (2S,3R)-hydroxycitric acid stereoisomer and its lactone form) is found enriched in the calyxes of H. sabdariffa [35]. Hydroxycitric acid lactone (Figure 1b), which is the lactone form of (+)-allo-hydroxycitric acid (Figure 1a), and its derivatives are the major organic compounds in the leaves and calyces extracts of H. sabdariffa [31].
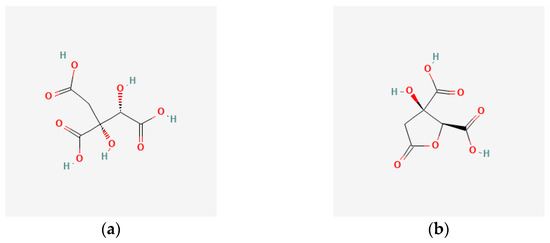
Figure 1.
The target compounds in this study. (a) (2S,3R)-hydroxycitric acid [43]; (b) (2S,3R)-hydroxycitric acid lactone [34,43,44].
Total activity and specific activity are two indicators for the assessment of the growth inhibitory capacity of a plant and an individual compound [15,26,45,46,47,48,49,50,51]. The total activity of a compound in a plant is the proportion of the concentration of an inhibitory compound in the producing plant to its specific activity (EC50), which is the effective concentration of the inhibitory compound required to exert a half-maximum effect on a receiver plant [44]. The activity of inhibitory compounds and the species containing inhibitory substances can be evaluated by their specific activity. The compounds with high biological activity per unit weight of the compound should be contemplated to develop herbicides. These compounds should have small EC50 values, which indicate high specific activity [48,52]. On the other hand, the total activity of considered inhibitory compounds could be assessed to determine the contribution of such compounds to the growth inhibitory effects of the species containing inhibitory substances [50]. The total activity assessment could be applied to approximate the impact of a compound on the inhibitory effect [50]. The compounds with high total activity, which is a functional indicator to declare the inhibitory potential of a compound, should be directed for further plant growth inhibitory research on the path of herbicide development [52,53]. In previous studies, several compounds have been isolated, identified, and evaluated by the total activity approach as inhibitory compounds. Based on the total activity approach, many plant growth inhibitory compounds such as L-3,4-dihydroxyphenylalanine (L-DOPA) in Mucuna pruriens [46], cyanamide in Vicia villosa [47], cis-cinnamoyl glucosides in Spiraea thunbergii [48], rutin in Fagopyrum esculentum [13], angelicin in Heracleum sosnowskyi fruit [49], carnosic acid in Rosmarinus officinalis leaves [15], and indigo in fruit pulp of Couroupita guianensis [26] were identified as the principal inhibitory compounds from reported inhibitory plants.
The introductory screening study for the inhibitory effects of plant species [32] indicated H. sabdariffa calyx as a potential plant growth inhibitory candidate that demonstrated the strongest inhibitory effect between others on the test plant, L. sativa. However, the plant growth inhibitory effect of calyx and the contribution of its two characteristic compounds to the inhibitory effect of calyx remain unknown. In this study, the inhibitory activity of H. sabdariffa calyx was demonstrated by specific activity, where the contribution of two compounds (hydroxycitric acid lactone and hydroxycitric acid) of calyx to its inhibition effect was evaluated by total activity. Consequently, current research focused on (i) the evaluation of the plant growth inhibitory effects of H. sabdariffa calyx water crude extract on L. sativa and (ii) the estimation of the contribution of hydroxycitric acid and hydroxycitric acid lactone to the phytotoxicity of H. sabdariffa calyx.
2. Materials and Methods
2.1. Plant Material and Extraction Procedure
The crude extract was obtained from the air-dried Hibiscus sabdariffa calyx plant sample. The air-dried samples were finely ground, and 160 mg of the sample was extracted with 20 mL distilled water (MilliporeSigmaTM, Burlington, MA, USA) at room temperature. The solution was sonicated (10 min), filtered (No. 1 filter paper, Advantec Toyo, Tokyo, Japan), and operated as a working solution.
2.2. Chemicals and Test Plant for Bioassay
Potassium hydroxycitrate tribasic monohydrate (hydroxycitric acid tripotassium salt) and (+)-garcinia acid ((−)-hydroxycitric acid lactone, (2S,3S)-3-hydroxy-5-oxotetrahydrofuran-2,3-dicarboxylic acid, Garcinia lactone) were bought from Sigma-Aldrich Chemie GmbH (Steinheim, Germany). Both compounds were analytical standards and were used without further purification. Lettuce seeds (Lactuca sativa, cultivar Legacy) were bought from a domestic seed company in Japan, Takii Seed Co., Ltd. (Kyoto, Japan) and used as test plant in the bioassay to assess the phytotoxic effect of pure compounds and Hibiscus sabdariffa calyx extract.
2.3. Inhibitory Effects of H. sabdariffa Crude Extract and Test Compounds
The specific activity of the crude extract and the total activity of the examined standard compounds were estimated using lettuce as a test plant as described by Golisz et al. [13] and Appiah et al. [15]. Concentrations of the crude extracts (10, 50, 100, 250, 500, 1000, and 4000 ppm), hydroxycitric acid (5, 10, 25, 50, 100, 200, 1000, 2000, and 4000 ppm), and hydroxycitric acid lactone (5, 10, 25, 50, 100, and 200 ppm) were prepared with distilled water for the inhibitory activity bioassays. Filter paper (27 mm ø, Toyo Roshi, Ltd., Tokyo, Japan) was laid down in a glass Petri dish (30 mm ø). A total of 0.7 mL of test solution was put into the filter. Five pre-germinated seedlings were left on the filter paper and incubated (CN-25C, Mitsubishi Elec., Tokyo, Japan) for 48 h at 22 °C in dark conditions.
Distilled water was used as the control treatment without crude extract or pure compound. Each treatment was replicated three times. After the incubation period, the lengths of hypocotyls and radicles were determined by millimeter paper with 1 mm accuracy, and the means were put into numbers as a percentage of the control. The elongation percentage was calculated using the Equation (1) below:
where X = treated (crude extract or pure compound) means radicle or hypocotyl length and Y = control means radicle or hypocotyl length.
The contribution of hydroxycitric acid lactone and hydroxycitric acid to the allelopathy of H. sabdariffa calyx crude extract was estimated through the total activity concept, which is derived from the inhibition and concentration of the compounds in the crude extract. Total activity was estimated by the division of hydroxycitric acid lactone or hydroxycitric acid concentration in H. sabdariffa calyx by the specific activity (EC50) of the compound. The concentrations of these compounds in H. sabdariffa calyx and their specific activities have the same unit. Therefore, the total activity is without any unit. Total activity was calculated using Equation (2) below [51]:
2.4. HPLC Analysis of H. sabdariffa Calyx
The ground 160 mg of the H. sabdariffa calyx samples were accurately weighed into the Erlenmeyer and extracted with 20 mL distilled water as shown in the extraction procedure to analyze hydroxycitric acid lactone and hydroxycitric acid concentration by HPLC. An aliquot of the extract was filtered through a 0.2 µm syringe filter before injection (5 µL). HPLC analysis was performed using an LC-20AD liquid chromatograph (Shimadzu, Kyoto, Japan). An Inertsil ODS 3 column (250 × 4.6 mm, 5 µm particles, GL Sciences Inc, Tokyo, Japan) was used in isocratic conditions. The mobile phase was 0.1% H3PO4 (phosphoric acid) in water. The column temperature was maintained at 30 °C, and the flow rate of the mobile phase was set at 0.5 mL/min. The analysis was monitored by using an SPD-M20A detector at 210 nm. The compounds were quantified by comparing the peak areas of the target commercial compounds with the abundance of these compounds in the corresponding standards used in the calibration curve. The quantification was obtained by comparing the peak areas of the target compounds with the abundance of these compounds in the corresponding standards used in the calibration curve. All chemical analyses were done in three replicates.
3. Results and Discussion
3.1. Effects of the Crude Extracts of Hibiscus sabdariffa Calyx on Lettuce Growth
The plant growth inhibitory effects of H. sabdariffa calyx crude extracts were investigated on the radicle and hypocotyl elongation of lettuce (Figure 2). The specific activity (EC50) of the crude extract from the calyx of H. sabdariffa was estimated for radicle and hypocotyl to be 877 ppm and 1880 ppm, respectively. The application of 4000 ppm of the crude calyx extract resulted in the highest inhibition (96.5%) of lettuce radicle growth (i.e., lettuce radicle elongation of 3.5%). Previous studies showed that the leaf, stem, and root of H. sabdariffa had inhibitory activity on the growth of test plant species [38,39]. However, there is no study on the plant growth inhibitory effect of the calyx of H. sabdariffa. This study assessed the influence of H. sabdariffa calyx on plant growth for the first time.

Figure 2.
Effects of water crude extract from H. sabdariffa calyx (HsCrude), hydroxycitric acid (HCA), and hydroxycitric acid lactone (HCAL) on the radicle (a) and hypocotyl (b) growth of lettuce seedlings. The data are the mean ± standard deviation, n = 3.
The specific activity (EC50) of H. sabdariffa calyx obtained in this study (877 ppm) was of a lower value than many species in the existing studies. In other studies, the ethanolic crude extracts of Rosmarinus officinalis [15] and of Phragmites communis Trin., and the methanolic crude extracts of Gliricidia sepium (Jacq.) Kunth, Pachysandra terminalis Siebold and Zucc., Samanea saman (Jacq.) Merr, Brachiaria brizantha (A. Rich.) Stapf, and Tamarindus indica L., demonstrated lower inhibitory effects (EC50 values of 1280, 2130, 1780, 1920, 2200, 2500, and 2510 ppm, respectively) [15,54,55,56]. The species containing inhibitory substances could be evaluated by their specific activity, and such species ought to have small EC50 values for their utilization in weed control [13,15,48]. H. sabdariffa calyx could be assessed as a prospective candidate for this purpose.
3.2. The Content of Pure Compounds in Hibiscus sabdariffa Calyx
Hydroxycitric acid and hydroxycitric acid lactone were found in the crude extracts of the examined calyx of H. sabdariffa. HPLC chromatograms of standard pure chemicals and H. sabdariffa calyx water crude extract with a retention time of 10.5 min for hydroxycitric acid (a) and 14.0 min for hydroxycitric acid lactone (b) are shown in Figure 3. The results of the HPLC analysis of H. sabdariffa calyx water extract confirmed the presence of hydroxycitric acid and hydroxycitric acid lactone in the calyx, Figure 3c. The concentration of hydroxycitric acid lactone was estimated at 167 mg g−1 dry weight of the calyx (16.7%), while that of hydroxycitric acid was 91.5 mg g−1 DW (9.15%). The hydroxycitric acid lactone was more abundant in the calyx of H. sabdariffa than hydroxycitric acid. Similarly, previous studies have also reported hydroxycitric acid lactone [35,57,58] and hydroxycitric acid [35,57,59] as the major organic acids in the calyx extract of H. sabdariffa.

Figure 3.
HPLC chromatograms (a) hydroxycitric acid; (b) hydroxycitric acid lactone; and (c) H. sabdariffa calyx extract.
3.3. Plant Growth Inhibitory Effects of Pure Compounds Present in H. sabdariffa Calyx
Hydroxycitric acid and hydroxycitric acid lactone were investigated for their plant growth inhibitory effect on lettuce. The examined compounds revealed different degrees of inhibitions on the lettuce radicle and hypocotyl elongation (Figure 2). The lettuce radicle elongation was inhibited more than hypocotyl elongation for both compounds examined in this study. Similar to the results of this study, some other reported allelochemicals had very minimal or no influence on the hypocotyl growth of lettuce seedlings [15,26,46]. Previous studies showed that the isolated chemicals (trimethyl allo-hydroxycitrate and β-sitosterol) from the leaf, stem, and root of H. sabdariffa had inhibitory activity on the growth of test plant species [41,42]. However, the allelopathy of the calyx’s substances had not been studied before. Current research assessed the worth of the influence of H. sabdariffa calyx’s characteristic compounds (hydroxycitric acid lactone and hydroxycitric acid) on the plant growth for the first time.
The specific activity (EC50) on lettuce radicle elongation was estimated at 1730 ppm for hydroxycitric acid and 73.7 ppm for hydroxycitric acid lactone, whereas EC50 on lettuce hypocotyl elongation of hydroxy citric acid lactone was about ≈36-fold lower than that of hydroxycitric acid. Moreover, the hormesis effect can explain a low-dose stimulatory effect, and a high-dose inhibitory effect of hydroxycitric acid on the hypocotyl growth (Figure 2b). Calabrese et al. [60] described this kind of dose-response relationship formerly. Hydroxycitric acid lactone demonstrated a stronger inhibitory effect on the L. sativa seedling growth than hydroxycitric acid had. Current research is reporting for the first time that hydroxycitric acid lactone could be an important plant growth inhibitor from the calyx of H. sabdariffa.
This study showed that hydroxycitric acid lactone, a saturated lactone form, is a stronger plant growth inhibitor than its non-lactone form, hydroxy citric acid. Previous studies have also shown that lactone-form compounds of some species have plant-growth inhibitory properties. Some bioactive natural chemicals, such as coumarin [61,62], umbelliferone [63,64], esculetin [14], scopoletin [65,66], patulin [67,68], psoralen [69,70], and angelicin [49], are α-β unsaturated lactones and have allelopathic potentials. Many unsaturated lactones are potent antimicrobial compounds. It was presumed that unsaturated lactone antibiotics may prevent enzyme activity by uniting with SH and possibly amino groups of enzyme proteins. This could be a basic mechanism of all unsaturated lactone inhibitors as well as coumarins and protoanemonin [71].
Moreover, sesquiterpene lactones and hydroxycitric acid lactone have the same lactone ring structure, which is in saturated form. Sesquiterpene lactones are broadly distributed in plants and the potential phytotoxicity [72] of dehydrocostus lactone [73,74], inuloxins A–D [75], artemisinin [76], and cnicin [77] has been reported. Momilactone [78] and acremolactone A, B, C [79,80] are other natural-origin compounds that demonstrated plant growth inhibitory effects on target plants in previous studies. Considering all the above presented, it could be thought that the lactone units of the compounds could be the core functional group required for biological activity such as the plant growth inhibitory effects of hydroxycitric acid lactone and the above-mentioned natural compounds.
Other organic acids such as citric acid, malic acid, and ascorbic acid were reported in the calyx of H. sabdariffa, aside from hydroxycitric acid lactone and hydroxycitric acid [31]. However, these organic acids have not shown the plant growth inhibitory effect; rather, they have revealed the plant growth improving effect on the target plants in several studies [52,81,82,83,84]. The application of citric acid has enhanced physiological parameters in numerous plant species such as Polianthes tuberosa, Lilium spp., and Phaseolus vulgaris (ordinary bean) [81] and had a growth stimulatory effect on L. sativa, for example [52]. Treatment with malic acid has significantly enhanced the growth of Miscanthus sacchariflorus under cadmium stress [82]. The foliar application of ascorbic acid has improved the shoot fresh and dry weights of cucumber [83] and early spring maize seedling establishment [84] in previous studies. Hence, the low EC50 of hydroxycitric acid lactone not only indicates the plant growth inhibitory effect of this compound but also further shows the influence and power of the compound for the plant growth inhibitory effect of calyx.
3.4. Estimation of Contributions of the Compounds to the Growth Inhibitory Activity of H. sabdariffa Calyx
The contribution of the inhibitory effects of hydroxycitric acid and hydroxycitric acid lactone in the crude extracts of H. sabdariffa calyx on the radicle elongation of lettuce seedlings was weighed up by the total activity approach. This approach was used to estimate which of the two compounds of H. sabdariffa calyx had a stronger contribution to the growth inhibitory activity of H. sabdariffa calyx on the radicle elongation of L. sativa. The total activity approach of evaluating the contribution of a plant growth inhibitor compound is based on the concentration and inhibitory effect (specific activity, EC50) of the compound [45]. The inhibitory effect of H. sabdariffa calyx extract on the radicle growth of lettuce could be described by the existence of hydroxycitric acid lactone in the extract (Figure 2). The total activity of hydroxycitric acid lactone and hydroxycitric acid was approximated at 2270 and 53.0, respectively. Thus, the plant growth inhibitory effect of hydroxycitric acid lactone was about ≈43-fold stronger than the inhibitory effect of hydroxycitric acid. One of the fundamental findings of this study is that the high concentration of hydroxycitric acid lactone in H. sabdariffa calyx coupled with low EC50 further shows the influence and dominance of the compound in the plant growth inhibition recognized. The inhibitory effect of H. sabdariffa calyx on both the radicle and hypocotyl growth of lettuce could be described by hydroxycitric acid lactone in the extract (Figure 2).
The total activity (TA) of hydroxycitric acid lactone in this study is 2270, and this value is relatively high when compared to many other plant growth inhibitory compounds in the existing literature. Based on the total activity evaluation, hydroxycitric acid lactone showed higher inhibitory effects than juglone (TA: 2000) [85], coumarin (TA: 2000) [86], L-DOPA (TA: 250) [46], indigo (TA: 58) [26], and cyanamide (TA: 40) [47]. However, goniothalamin (TA: 3600) [87] had a higher total activity value than hydroxycitric acid lactone.
4. Conclusions
H. sabdariffa calyx demonstrated a strong plant growth inhibitory effect on lettuce growth elongation in the initial screening. The inhibitory effect of the calyx and its substances had not been studied before. The result showed that the calyx of H. sabdariffa had plant growth inhibitory potential, and this is the first report. The crude extract from the calyx of H. sabdariffa and the pure hydroxycitric acid lactone inhibited lettuce radicle elongation by 50% at 877 ppm and 73.7 ppm, respectively. The inhibitory contribution of hydroxycitric acid lactone to the effect of H. sabdariffa calyx extract was estimated (using the total activity approach) to be higher than hydroxycitric acid showed. This is the first report of hydroxycitric acid lactone as a plant growth inhibitor from H. sabdariffa calyxes. Future studies should focus on evaluating the herbicidal potential of H. sabdariffa calyx and hydroxycitric acid lactone under both greenhouse and field conditions.
Author Contributions
Conceptualization, T.G.I.O. and Y.F.; methodology, T.G.I.O., K.S.A. and Y.F.; software, Microsoft Office 2016; validation, T.G.I.O., E.A. and Y.F.; formal analysis, T.G.I.O.; investigation, T.G.I.O.; resources, T.G.I.O. and Y.F.; data curation, T.G.I.O. and Y.F.; writing—original draft preparation, T.G.I.O.; writing—review and editing, T.G.I.O., K.S.A. and Y.F.; visualization, T.G.I.O. and K.S.A.; supervision, Y.F.; project administration, T.G.I.O. and Y.F.; and funding acquisition, Y.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly supported by JST CREST Grant Number JPMJCR1702 and JSPS KAKENHI Grant Number 26304024.
Data Availability Statement
Not applicable. Data related to this publication is shared in the manuscript.
Acknowledgments
We would like to thank Kohinoor Begum for her technical support during the experiments. We also thank our other colleagues and laboratory members of the International Environmental and Agricultural Sciences, Tokyo University of Agriculture and Technology.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oerke, E.-C.; Dehne, H.-W. Safeguarding production—Losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Appleby, A.P.; Muller, F.; Carpy, S. Weed control. In Agrochemicals; Muller, F., Ed.; Wiley: New York, NY, USA, 2000; pp. 687–707. [Google Scholar]
- Kraehmer, H.; Baur, P. Weed Anatomy; Wiley-Blackwell: Chichester, UK, 2013. [Google Scholar]
- Scavo, A.; Mauromicale, G. Integrated Weed Management in Herbaceous Field Crops. Agronomy 2020, 10, 466. [Google Scholar] [CrossRef]
- Gawronska, H.; Ciarka, D.; Bernat, W.; Gawronski, S.W. Sunflower-desired Allelopathic Crop for Sustainable and Organic Agriculture. In Allelopathy New Concept and Methodology; Fujii, Y., Hiradate, S., Eds.; Science Publishers: Enfield, CT, USA, 2007; pp. 185–210. [Google Scholar]
- Scavo, A.; Mauromicale, G. Crop Allelopathy for Sustainable Weed Management in Agroecosystems: Knowing the Present with a View to the Future. Agronomy 2021, 11, 2104. [Google Scholar] [CrossRef]
- Duke, S.O. Why have no new herbicide modes of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef]
- Cárdenas, D.M.; Bajsa-Hirschel, J.; Cantrell, C.L.; Rial, C.; Varela, R.M.; Molinillo, J.M.G.; Macías, F.A. Evaluation of the phytotoxic and antifungal activity of C17-sesquiterpenoids as potential biopesticides. Pest Manag. Sci. 2022, 10, 4240–4251. [Google Scholar] [CrossRef]
- Vencill, W.; Nichols, R.; Webster, T.; Soteres, J.; Mallory-Smith, C.; Burgos, N.; Johnson, W.G.; McClelland, M. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef]
- Fujii, Y.; Parvez, S.S.; Parvez, M.M.; Ohmae, Y.; Iida, O. Screening of 239 medicinal plant species for allelopathic activity using the sandwich method. Weed Biol. Manag. 2003, 3, 233–241. [Google Scholar] [CrossRef]
- Iqbal, Z.; Nasir, H.; Fujii, Y. Allelopathic Activity of Buckwheat: A ground Cover Crop for Weed Control. In Allelopathy New Concept and Methodology; Fujii, Y., Hiradate, S., Eds.; Science Publishers: Enfield, CT, USA, 2007; pp. 173–183. [Google Scholar]
- Golisz, A.; Lata, B.; Gawronski, S.W.; Fujii, Y. Specific and total activities of the allelochemicals identified in buckwheat. Weed Biol. Manag. 2007, 7, 164–171. [Google Scholar] [CrossRef]
- Haig, T. Allelochemicals in Plants. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, R.S., Mallik, A.U., Luo, S.M., Eds.; Springer: New York, NY, USA, 2008; pp. 63–104. [Google Scholar]
- Appiah, K.S.; Mardani, H.K.; Omari, R.A.; Eziah, V.Y.; Ofosu-Anim, J.; Onwona-Agyeman, S.; Amoatey, C.A.; Kawada, K.; Katsura, K.; Oikawa, Y.; et al. Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves. Toxins 2018, 10, 498. [Google Scholar] [CrossRef]
- Vyvyan, J.R. Allelochemicals as Leads for New Herbicides and Agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Barney, J.N.; Hay, A.G.; Weston, L.A. Isolation and Characterization of Allelopathic Volatiles from Mugwort (Artemisia vulgaris). J. Chem. Ecol. 2005, 31, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. CRC Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowsk, A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides: Current Research and Case Studies in Use; Price, A.J., Kelton, J.A., Eds.; IntechOpen: London, UK, 2013; pp. 444–466. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Mauromicale, G. Leaf extracts of cultivated cardoon as potential bioherbicide. Sci. Hortic. 2020, 261, 109024. [Google Scholar] [CrossRef]
- Duke, S.O.; Dayan, F.E. Modes of action of microbially-produced phytotoxins. Toxins 2011, 3, 1038–1064. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Dayan, F.E.; Romagni, J.G.; Rimando, A.M. Natural products as sources for new mechanisms of herbicidal action. Crop Prot. 2000, 40, 99–111. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 60, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Molinillo, J.M.G.; Galindo, J.C.G.; Varela, R.M.; Simonet, A.M.; Castellano, D. The Use of Allelopathic Studies in the Search for Natural Herbicides. J. Crop Prod. 2001, 4, 237–255. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Begum, K.; Motobayashi, T.; Hasan, N.; Appiah, K.S.; Shammi, M.; Fujii, Y. Indigo as a Plant Growth Inhibitory Chemical from the Fruit Pulp of Couroupita guianensis Aubl. Agronomy 2020, 10, 1388. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T.; An, M. Screening method for the evaluation of crop allelopathic potential. Bot. Rev. 2001, 67, 403–415. [Google Scholar] [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Lombardo, S.; Pesce, G.R.; Mauromicale, G. Allelopathic potential of leaf aqueous extracts from Cynara cardunculus L. on the seedling growth of two cosmopolitan weed species. Ital. J. Agron. 2019, 14, 78–83. [Google Scholar] [CrossRef]
- Bhadoria, P. Allelopathy: A Natural Way towards Weed Management. Am. J. Exp. Agric. 2011, 1, 7–20. [Google Scholar] [CrossRef]
- Appiah, K.S.; Li, Z.; Zeng, R.S.; Luo, S.; Oikawa, Y.; Fujii, Y. Determination of allelopathic potentials in plant species in Sino-Japanese floristic region by sandwich method and dish pack method. Int. J. Basic Appl. Sci. 2015, 4, 381–394. [Google Scholar] [CrossRef]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.—A phytochemical and pharmacological review. Food Chem. 2014, 165, 423–443. [Google Scholar] [CrossRef]
- Isin Ozkan, T.G.; Akalin Urusak, E.; Appiah, K.S.; Fujii, Y. First Broad Screening of Allelopathic Potential of Wild and Cultivated Plants in Turkey. Plants 2019, 8, 532. [Google Scholar] [CrossRef]
- Zarrabal, O.; Barradas-Dermitz, D.M.; Orta-Flores, Z.; Hayward-Jones, P.M.; Nolasco-Hipólito, C.N.; Aguilar-Uscanga, M.G.; Miranda-Medina, A.; Bujang, K.B. Hibiscus sabdariffa L., roselle calyx, from ethnobotany to pharmacology. J. Exp. Pharmacol. 2012, 4, 25–39. [Google Scholar] [CrossRef]
- Hida, H.; Yamada, T.; Yamada, Y. Absolute Configuration of Hydroxycitric Acid Produced by Microorganisms. Biosci. Biotechnol. Biochem. 2006, 70, 1972–1974. [Google Scholar] [CrossRef]
- Yamada, T.; Hida, H.; Yamada, Y. Chemistry, physiological properties, and microbial production of hydroxycitric acid. Appl. Microbiol. Biotechnol. 2007, 75, 977–982. [Google Scholar] [CrossRef]
- Zheoat, A.M.; Gray, A.I.; Igoli, J.O.; Kennedy, A.R.; Ferro, V.A. Crystal structures of hibiscus acid and hibiscus acid dimethyl ester isolated from Hibiscus sabdariffa (Malvaceae). Acta Cryst. 2017, 73, 1368–1371. [Google Scholar] [CrossRef]
- Hida, H.; Yamada, T.; Yamada, Y. Genome shuffling of Streptomyces sp. U121 for improved production of hydroxycitric acid. Appl. Microbiol. Biotechnol. 2007, 73, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Piyatida, P.; Kato-Noguchi, H. Screening of Allelopathic Activity of Eleven Thai Medicinal Plants on Seedling Growth of Five Test Plant Species. Asian J. Plant Sci. 2010, 9, 486–491. [Google Scholar] [CrossRef]
- Pukclai, P.; Kato-Noguchi, H. Evaluation of allelopathic activity of Hibiscus sabdariffa L. Adv. Biol. Res. 2011, 5, 366–372. [Google Scholar]
- Suwitchayanon, P.; Pukclai, P.; Ohno, O.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of an allelopathic substance from Hibiscus sabdariffa. Nat. Prod. Commun. 2015, 10, 765–766. [Google Scholar] [CrossRef] [PubMed]
- Piyatida, P.; Kimura, P.; Sato, M.; Kato-Noguchi, H. Isolation of β-sitosterol from Hibiscus sabdariffa L. Allelopath. J. 2013, 32, 289–300. [Google Scholar]
- Carvajal-Zarrabal, O.; Waliszewski, S.M.; Barradas-Dermitz, D.M.; Orta-Flores, Z.; Hayward-Jones, P.M.; Nolasco-Hipólito, C.; Angulo-Guerrero, O.; Sánchez-Ricaño, R.; Infanzón, R.M.; Trujillo, P.R. The consumption of Hibiscus sabdariffa dried calyx ethanolic extract reduced lipid profile in rats. Plant Foods Hum. Nutr. 2005, 60, 153–159. [Google Scholar] [CrossRef]
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/ALLO-2-hydroxycitric-acid (accessed on 23 June 2023).
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/6481826 (accessed on 23 June 2023).
- Hiradate, S.; Ohse, K.; Furunayashi, A.; Fujii, Y. Quantitative evaluation of allelopathic potentials in soils: Total activity approach. Weed Sci. 2010, 58, 258–264. [Google Scholar] [CrossRef]
- Fujii, Y.; Shibuya, T.; Yasuda, T. Allelopathy of mucuna: Its discrimination and identification of L-DOPA as a candidate of allelopathic substance. Jpn. Agric. Res. Q. 1992, 25, 238–247. [Google Scholar]
- Kamo, T.; Hiradate, S.; Fujii, Y. First Isolation of Natural Cyanamide as a Possible Allelochemical from Hairy Vetch Vicia villosa. J. Chem. Ecol. 2003, 29, 275–283. [Google Scholar] [CrossRef]
- Hiradate, S.; Morita, S.; Sugie, H.; Fujii, Y.; Harada, J. Phytotoxic Cis-Cinnamoyl Glucosides from Spiraea thunbergii. Phytochemistry 2004, 65, 731–739. [Google Scholar] [CrossRef]
- Mishyna, M.; Laman, N.; Prokhorov, V.; Fujii, Y. Angelicin as the Principal Allelochemical in Heracleum sosnowskyi Fruit. Nat. Prod. Commun. 2015, 10, 767–770. [Google Scholar] [CrossRef] [PubMed]
- Hiradate, S.; Morita, S.; Furubayashi, A.; Fujii, Y.; Harada, J. Plant Growth Inhibition by Cis-Cinnamoyl Glucosides and Cis-Cinnamic Acid. J. Chem. Ecol. 2005, 31, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Hiradate, S. A Critical Survey of Allelochemicals in Action—The Importance of Total Activity and the Weed Suppression Equation. In Establishing the Scientific Base; Harper, J.D.I., An, M., Wu, H., Kent, J.H., Eds.; Centre for Rural Social Research, Charles Sturt University: Wagga Wagga, Australia, 2005; pp. 73–76. [Google Scholar]
- Syed, S.; Ahmed, Z.I.; Al-haq, M.I.; Mohammad, A.; Fujii, Y. The possible role of organic acids as allelochemicals in Tamarindus indica L. leaves. Acta Agric. Scand. 2014, 64, 511–517. [Google Scholar] [CrossRef]
- Hiradate, S. Isolation strategies for finding bioactive compounds: Specific activity vs. total activity. In Natural Products for Pest Management; Rimando, A.M., Duke, S.O., Eds.; American Chemical Society: Washington, DC, USA, 2006; pp. 113–126. [Google Scholar]
- Takemura, T.; Sakuno, E.; Kamo, T.; Hiradate, S.; Fujii, Y. Screening of the Growth-Inhibitory Effects of 168 Plant Species against Lettuce Seedlings. Am. J. Plant Sci. 2013, 4, 1095–1104. [Google Scholar] [CrossRef]
- Li, F.; Hu, H. Isolation and Characterization of a Novel Antialgal Allelochemical from Phragmites communis. J. Appl. Environ. Microbiol. 2005, 71, 6545–6553. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kato-Noguchi, H. The Seasonal Variations of Allelopathic Activity and Allelopathic Substances in Brachiaria brizantha. Bot. Stud. 2015, 56, 25. [Google Scholar] [CrossRef]
- Rodríguez-Medina, I.C.; Beltrán-Debón, R.; Molina, V.M.; Alonso-Villaverde, C.; Joven, J.; Menéndez, J.A.; Fernández-Gutiérrez, A. Direct characterization of aqueous extract of Hibiscus sabdariffa using HPLC with diode array detection coupled to ESI and ion trap MS. J. Sep. Sci. 2009, 32, 3441–3448. [Google Scholar] [CrossRef]
- Ramirez-Rodrigues, M.M.; Plaza, M.L.; Azeredo, A.; Balaban, M.O.; Marshall, M.R. Physicochemical and phytochemical properties of cold and hot water extraction from Hibiscus sabdariffa. J. Food Sci. 2011, 76, C428–C435. [Google Scholar] [CrossRef]
- Beltrán-Debón, R.; Alonso-Villaverde, C.; Aragones, G.; Rodriguez-Medina, I.; Rull, A.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Camps, J.; Joven, J. The aqueous extract of Hibiscus sabdariffa calices modulates the production of monocyte chemoattractant protein-1 in humans. Phytomedicine 2010, 17, 186–191. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Bachmann, K.A.; Bailer, A.J.; Bolger, P.M.; Borak, J.; Cai, L.; Cedergreen, N.; Cherian, M.G.; Chiueh, C.C.; Clarkson, T.W.; et al. Biological stress response terminology: Integrating the concepts of adaptive response and preconditioning stress within a hormetic dose–response framework. Toxicol. Appl. Pharmacol. 2007, 222, 122–128. [Google Scholar] [CrossRef]
- Haig, T.J.; Seal, A.N.; Pratley, J.E.; An, M.; Wu, H. Lavender as a source of novel plant compounds for the development of a natural herbicide. J. Chem. Ecol. 2009, 35, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhao, G.; Liu, D.; Liu, S.; Gun, X.; Tang, Q. Discovery and Weed Inhibition Effects of Coumarin as the Predominant Allelochemical of Yellow Sweet clover (Melilotus officinalis). Int. J. Agric. Biol. 2016, 18, 168–175. [Google Scholar] [CrossRef]
- Real, M.; Gamiz, B.; Lopez-Cabeza, R.; Celis, R. Sorption, persistence, and leaching of the allelochemical umbelliferone in soils treated with nanoengineered sorbents. Sci. Rep. 2019, 9, 9764. [Google Scholar] [CrossRef]
- Morikawa, C.I.O.; Miyaura, R.; Kamo, T.; Hiradate, S.; Chávez Pérez, J.A.; Fujii, Y. Isolation of Umbelliferone as a Principal Allelochemical from the Peruvian Medicinal Plant Diplostephium foliosissimum (Asteraceae). Rev. Soc. Quim. Peru 2011, 77, 285–291. [Google Scholar]
- Narwal, S.S. Allelochemicals. In Allelopathy in Crop Production; Kumar, P., Ed.; Scientific Publishers: Jodhpur, India, 2004; pp. 6–17. ISBN 978-8172-337-77-3. [Google Scholar]
- Fay, P.; Duke, W. An Assessment of Allelopathic Potential in Avena Germ Plasm. Weed Sci. 1977, 25, 224–228. [Google Scholar] [CrossRef]
- Einhellig, F. Allelopathy-A Natural Protection, Allelochemicals. In CRC Handbook of Natural Pesticides: Methods Volume I: Theory, Practice, and Detection; Mandava, N.B., Ed.; CRC Press: Boca Raton, FL, USA, 2017; pp. 161–200. ISBN 9781351072717. [Google Scholar]
- Chen, J.; Wei, J.; Gao, J.M.; Ye, X.; Mcerlan, C.S.P.; Ma, Y. Allelopathic inhibitory effects of Penicillium griseofulvum produced patulin on the seed germination of Orobanche cumana Wallr. and Phelipanche aegyptiaca Pers. Allelopath. J. 2017, 41, 65–80. [Google Scholar] [CrossRef]
- Putnam, A.R. Weed Allelopathy. In Weed Physiology: Volume I: Reproduction and Ecophysiology; Duke, S.O., Ed.; Reissued 2018; CRC Press: Boca Raton, FL, USA, 1985; Volume 1, pp. 131–156. ISBN 0-8493-6313-6. [Google Scholar]
- Shilling, D.G.; Dusky, J.A.; Mossier, M.A.; Bewick, T.A. Allelopathic Potential of Celery Residues on Lettuce. J. Am. Soc. Hortic. Sci. 1992, 117, 308–312. Available online: https://journals.ashs.org/jashs/downloadpdf/journals/jashs/117/2/article-p308.pdf (accessed on 25 December 2022). [CrossRef]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press, Inc.: Orlando, FL, USA, 1984; pp. 233, 268, 344. ISBN 978-0-12-587055-9. [Google Scholar]
- Macias, F.A.; Fernandez, A.; Varela, R.M.; Molinillo, J.M.G.; Torres, A.; Alves, P.L.C.A. Sesquiterpene Lactones as Allelochemicals. J. Nat. Prod. 2006, 69, 795–800. [Google Scholar] [CrossRef]
- Toda, Y.; Shigemori, H.; Ueda, J.; Miyamoto, K. Isolation and identification of polar auxin transport inhibitors from Saussurea costus and Atractylodes japonica. Acta Agrobot. 2017, 70, 1700. [Google Scholar] [CrossRef]
- Toda, Y.; Okada, K.; Ueda, J.; Miyamoto, K. Dehydrocostus lactone, a naturally occurring polar auxin transport inhibitor, inhibits epicotyl growth by interacting with auxin in etiolated Pisum sativum seedlings. Acta Agrobot. 2019, 72, 1779. [Google Scholar] [CrossRef]
- Andolfi, A.; Zermane, N.; Cimmino, A.; Avolio, F.; Boari, A.; Vurro, M.; Evidente, A. Inuloxins A-D, phytotoxic bi-and tri-cyclic sesquiterpene lactones produced by Inula viscosa: Potential for broomrapes and field dodder management. Phytochemistry 2013, 86, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Wang, C.; Chen, J.; Peng, J.; Cao, Q. A novel sensitive electrochemical sensor based on in-situ polymerized molecularly imprinted membranes at graphene modified electrode for artemisinin determination. Biosens. Bioelectron. 2015, 64, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Schabes, F.I.; Sigstad, E.E. A calorimetric study of the allelopathic effect of cnicin isolated from Centaurea diffusa Lam. on the germination of soybean (Glicine max) and radish (Raphanus sativus). Thermochim. Acta 2007, 458, 84–87. [Google Scholar] [CrossRef]
- Xu, M.; Galhano, R.; Wiemann, P.; Bueno, E.; Tiernan, M.; Wu, W.; Chung, I.M.; Gershenzon, J.; Tudzynski, B.; Sesma, A.; et al. Genetic evidence for natural product-mediated plant–plant allelopathy in rice (Oryza sativa). New Phytol. 2012, 193, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Kinoshita, H.; Nukina, M.; Sugiyama, T. Acremolactone A, a novel herbicidal epoxydihydropyranyl γ-lactone from Acremonium roseum I4267. J. Antibiot. 1998, 5, 967–969. [Google Scholar] [CrossRef] [PubMed]
- Sassa, T.; Ooi, T.; Kinoshita, H. Isolation and Structures of Acremolactones B and C, Novel Plant-Growth Inhibitory γ-Lactones from Acremonium roseum I4267. Biosci. Biotechnol. Biochem. 2004, 68, 2633–2636. [Google Scholar] [CrossRef] [PubMed]
- Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric Acid-Mediated Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Guo, H.; Chen, H.; Hong, C.; Jiang, D.; Zheng, B. Exogenous malic acid alleviates cadmium toxicity in Miscanthus sacchariflorus through enhancing photosynthetic capacity and restraining ROS accumulation. Ecotoxicol. Environ. Saf. 2017, 141, 119–128. [Google Scholar] [CrossRef]
- Naz, H.; Akram, N.A.; Ashraf, M. Impact of Ascorbic Acid on Growth and Some Physiological Attributes of Cucumber (Cucumber sativus) Plants under Water-Deficit Conditions. Pak. J. Bot. 2016, 48, 877–883. [Google Scholar]
- Ahmad, I.; Basra, S.M.A.; Afzal, I.; Farooq, M.; Wahid, A. Growth improvement in spring maize through exogenous application of ascorbic acid, salicylic acid and hydrogen peroxide. Int. J. Agric. Biol. 2013, 15, 95–100. [Google Scholar]
- Jung, K.; Fujii, Y.; Yoshizaki, S.; Kobori, H. Evaluation of total allelopathic of heartseed walnut (Juglans ailanthifolia Carr.) and its potential to control black locust (Robinia pseudoacacia L.). Allelopath. J. 2010, 26, 243–253. [Google Scholar]
- Yamamoto, Y.; Fujii, Y. Exudation of allelopathic compound from plant roots of sweet vernal grass (Anthoxanthum odoratum). J. Weed Sci. Technol. 1997, 42, 31–35. [Google Scholar] [CrossRef]
- Raihan, I.; Miyaura, R.; Baki, B.B.; Fujii, Y. Assessment of allelopathic potential of goniothalamin allelochemical from Malaysian plant Goniothalamus andersonii J. Sinclair by sandwich method. Allelopath. J. 2019, 46, 25–40. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).