
Knowledge of Cover Crop Seed Traits and Treatments to Enhance Weed Suppression: A Narrative Review



Abstract
1. Introduction
2. Factors Affecting Cover Crops’ Seed Germination
2.1. Abiotic Factors
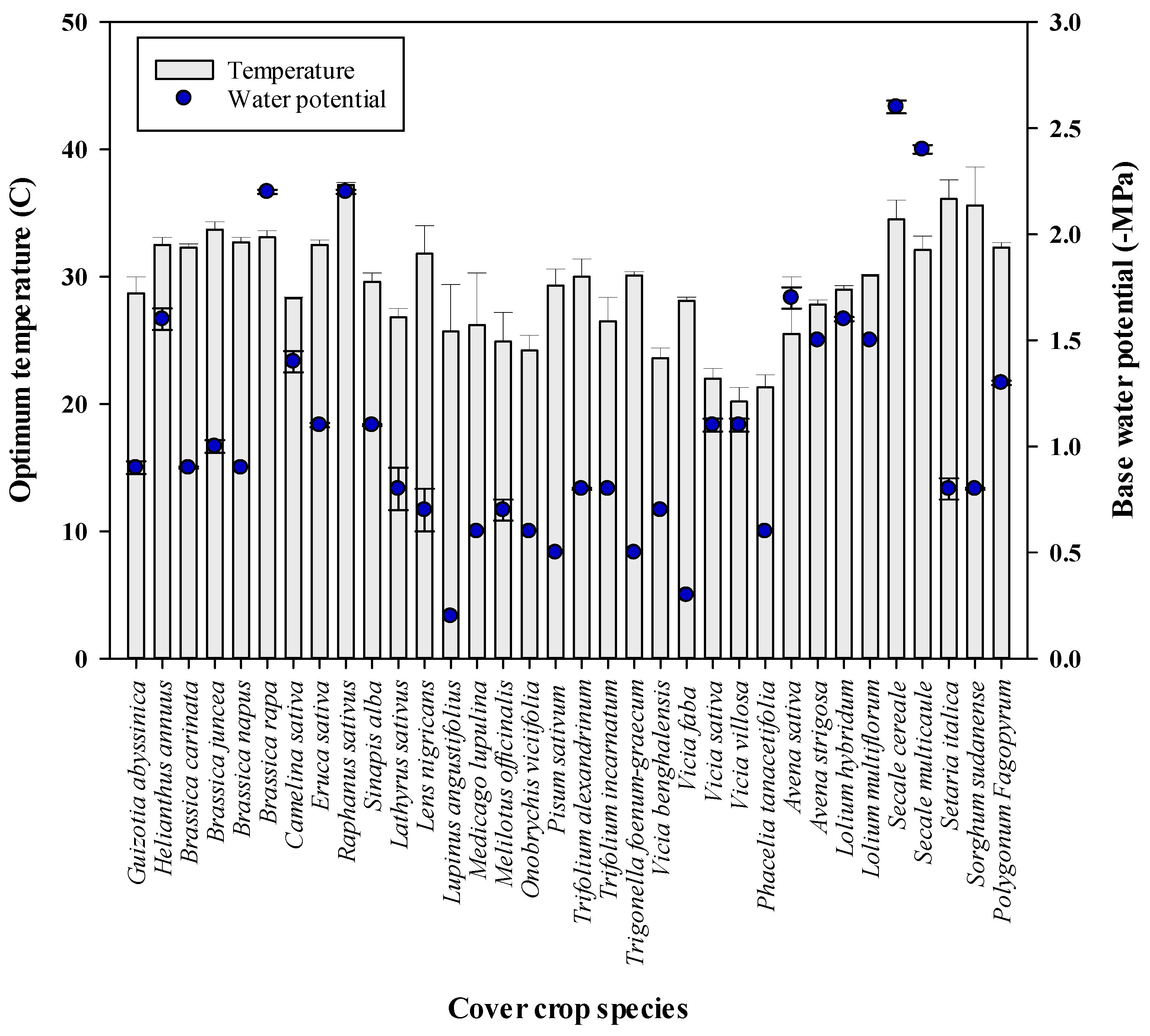
Temperature and Soil Moisture
2.2. Seed Dormancy
3. Seed Traits of Main Cover Crop Species
3.1. Fabaceae
3.1.1. Vicia sp.
3.1.2. Trifolium sp.
3.2. Poaceae
3.3. Brassicaceae
4. Solutions for Enhancing Cover Crop Seed Germinability
4.1. Agronomic Practices
Sowing Time and Planting Geometry
4.2. Seed Pre-Treatment
4.2.1. Seed Priming
4.2.2. Seed Coating
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bond, W.; Turner, R.; Grundy, A. A review of non-chemical weed management. In HDRA, the Organic Organisation; Ryton Organic Gardens: Coventry, UK, 2003; p. 81. [Google Scholar]
- Colbach, N.; Petit, S.; Chauvel, B.; Deytieux, V.; Lechenet, M.; Munier-Jolain, N.; Cordeau, S. The Pitfalls of Relating Weeds, Herbicide Use, and Crop Yield: Don’t Fall Into the Trap! A Critical Review. Front. Agron. 2020, 2, 33. [Google Scholar] [CrossRef]
- Travlos, I.; De Prado, R.; Chachalis, D.; Bilalis, D.J. Herbicide Resistance in Weeds: Early Detection, Mechanisms, Dispersal, New Insights and Management Issues; Frontiers Media SA: Lausanne, Switzerland, 2020. [Google Scholar]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot. Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef]
- Adeux, G.; Vieren, E.; Carlesi, S.; Bàrberi, P.; Munier-Jolain, N.; Cordeau, S. Mitigating crop yield losses through weed diversity. Nat. Sustain. 2019, 2, 1018–1026. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 19 March 2020).
- Stoate, C.; Baldi, A.; Beja, P.; Boatman, N.; Herzon, I.; Van Doorn, A.; De Snoo, G.; Rakosy, L.; Ramwell, C. Ecological impacts of early 21st century agricultural change in Europe—A review. J. Environ. Manag. 2009, 91, 22–46. [Google Scholar] [CrossRef]
- Weber, J.F.; Kunz, C.; Peteinatos, G.G.; Zikeli, S.; Gerhards, R. Weed control using conventional tillage, reduced tillage, no-tillage, and cover crops in organic soybean. Agriculture 2017, 7, 43. [Google Scholar] [CrossRef]
- Petit, S.; Cordeau, S.; Chauvel, B.; Bohan, D.; Guillemin, J.-P.; Steinberg, C. Biodiversity-based options for arable weed management. A review. Agron. Sustain. Dev. 2018, 38, 48. [Google Scholar] [CrossRef]
- Bhaskar, V.; Westbrook, A.S.; Bellinder, R.R.; DiTommaso, A. Integrated management of living mulches for weed control: A review. Weed Technol. 2021, 35, 856–868. [Google Scholar] [CrossRef]
- Nosratti, I.; Sabeti, P.; Chaghamirzaee, G.; Heidari, H. Weed problems, challenges, and opportunities in Iran. Crop Prot. 2020, 134, 104371. [Google Scholar] [CrossRef]
- Kumar, V.; Obour, A.; Jha, P.; Liu, R.; Manuchehri, M.R.; Dille, J.A.; Holman, J.; Stahlman, P.W. Integrating cover crops for weed management in the semiarid US Great Plains: Opportunities and challenges. Weed Sci. 2020, 68, 311–323. [Google Scholar] [CrossRef]
- Teasdale, J.; Brandsaeter, L.; Calegari, A.; Neto, F.S.; Upadhyaya, M.; Blackshaw, R. Cover crops and weed management. In Non Chemical Weed Management Principles; Concepts and Technology; CABI: Wallingford, UK, 2007; pp. 49–64. [Google Scholar]
- Scavo, A.; Fontanazza, S.; Restuccia, A.; Pesce, G.R.; Abbate, C.; Mauromicale, G. The role of cover crops in improving soil fertility and plant nutritional status in temperate climates. A review. Agron. Sustain. Dev. 2022, 42, 93. [Google Scholar] [CrossRef]
- Rouge, A.; Adeux, G.; Busset, H.; Hugard, R.; Martin, J.; Matejicek, A.; Moreau, D.; Guillemin, J.-P.; Cordeau, S. Weed suppression in cover crop mixtures under contrasted levels of resource availability. Eur. J. Agron. 2022, 136, 126499. [Google Scholar] [CrossRef]
- Rouge, A.; Adeux, G.; Busset, H.; Hugard, R.; Martin, J.; Matejicek, A.; Moreau, D.; Guillemin, J.-P.; Cordeau, S. Carry-over effects of cover crops on weeds and crop productivity in no-till systems. Field Crop. Res. 2023, 295, 108899. [Google Scholar] [CrossRef]
- Osipitan, O.A.; Dille, J.A.; Assefa, Y.; Knezevic, S.Z. Cover crop for early season weed suppression in crops: Systematic review and meta-analysis. Agron. J. 2018, 110, 2211–2221. [Google Scholar] [CrossRef]
- Mahé, I.; Chauvel, B.; Colbach, N.; Cordeau, S.; Gfeller, A.; Reiss, A.; Moreau, D. Deciphering field-based evidences for crop allelopathy in weed regulation. A review. Agron. Sustain. Dev. 2022, 42, 50. [Google Scholar] [CrossRef]
- Teasdale, J.; Hatfield, J.; Buhler, D.; Stewart, B. Cover crops, smother plants, and weed management. Integr. Weed Soil Manag. 1998, 247, 270. [Google Scholar]
- Tursun, N.; Işık, D.; Demir, Z.; Jabran, K. Use of Living, Mowed, and Soil-Incorporated Cover Crops for Weed Control in Apricot Orchards. Agronomy 2018, 8, 150. [Google Scholar] [CrossRef]
- Ward, M.J.; Ryan, M.R.; Curran, W.S.; Barbercheck, M.E.; Mortensen, D.A. Cover crops and disturbance influence activity-density of weed seed predators Amara aenea and Harpalus pensylvanicus (Coleoptera: Carabidae). Weed Sci. 2011, 59, 76–81. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Moonen, A.; Barberi, P. Size and composition of the weed seedbank after 7 years of different cover-crop-maize management systems. Weed Res. 2004, 44, 163–177. [Google Scholar] [CrossRef]
- Lawley, Y.E.; Teasdale, J.R.; Weil, R.R. The mechanism for weed suppression by a forage radish cover crop. Agron. J. 2012, 104, 205–214. [Google Scholar] [CrossRef]
- Johnson, G.A.; Defelice, M.S.; Helsel, Z.R. Cover crop management and weed control in corn (Zea mays). Weed Technol. 1993, 7, 425–430. [Google Scholar] [CrossRef]
- Alonso-Ayuso, M.; Gabriel, J.L.; García-González, I.; Del Monte, J.P.; Quemada, M. Weed density and diversity in a long-term cover crop experiment background. Crop Prot. 2018, 112, 103–111. [Google Scholar] [CrossRef]
- Büchi, L.; Wendling, M.; Amossé, C.; Jeangros, B.; Charles, R. Cover crops to secure weed control strategies in a maize crop with reduced tillage. Field Crop. Res. 2020, 247, 107583. [Google Scholar] [CrossRef]
- Smith, R.G.; Warren, N.D.; Cordeau, S. Are cover crop mixtures better at suppressing weeds than cover crop monocultures? Weed Sci. 2020, 68, 186–194. [Google Scholar] [CrossRef]
- Derrouch, D.; Chauvel, B.; Felten, E.; Dessaint, F. Weed Management in the Transition to Conservation Agriculture: Farmers’ Response. Agronomy 2020, 10, 843. [Google Scholar] [CrossRef]
- Cordeau, S.; Smith, R.G.; Gallandt, E.R.; Brown, B.; Salon, P.; DiTommaso, A.; Ryan, M.R. Timing of tillage as a driver of weed communities. Weed Sci. 2017, 65, 504–514. [Google Scholar] [CrossRef]
- Adeux, G.; Cordeau, S.; Antichi, D.; Carlesi, S.; Mazzoncini, M.; Munier-Jolain, N.; Bàrberi, P. Cover crops promote crop productivity but do not enhance weed management in tillage-based cropping systems. Eur. J. Agron. 2021, 123, 126221. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Osipitan, O.A.; Dille, J.A.; Assefa, Y.; Radicetti, E.; Ayeni, A.; Knezevic, S.Z. Impact of cover crop management on level of weed suppression: A meta-analysis. Crop Sci. 2019, 59, 833–842. [Google Scholar] [CrossRef]
- Mennan, H.; Jabran, K.; Zandstra, B.H.; Pala, F. Non-chemical weed management in vegetables by using cover crops: A review. Agronomy 2020, 10, 257. [Google Scholar] [CrossRef]
- Grünwald, N.J.; Hu, S.; van Bruggen, A.H.C. Short-term Cover Crop Decomposition in Organic and Conventional Soils: Characterization of Soil C, N, Microbial and Plant Pathogen Dynamics. Eur. J. Plant Pathol. 2000, 106, 37–50. [Google Scholar] [CrossRef]
- Constantin, J.; Le Bas, C.; Justes, E. Large-scale assessment of optimal emergence and destruction dates for cover crops to reduce nitrate leaching in temperate conditions using the STICS soil–crop model. Eur. J. Agron. 2015, 69, 75–87. [Google Scholar] [CrossRef]
- Yang, C.; Sun, R.; Lu, X.; Jin, T.; Peng, X.; Zhang, N.; Wang, J.; Wang, H.; Liu, W. Seed-germination ecology of Vicia villosa Roth, a cover crop in orchards. Agronomy 2022, 12, 2488. [Google Scholar] [CrossRef]
- Bradford, K.J. A water relations analysis of seed germination rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef]
- Yin, X.; Kropff, M.J.; McLaren, G.; Visperas, R.M. A nonlinear model for crop development as a function of temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef]
- Constantin, J.; Dürr, C.; Tribouillois, H.; Justes, E. Catch crop emergence success depends on weather and soil seedbed conditions in interaction with sowing date: A simulation study using the SIMPLE emergence model. Field Crop. Res. 2015, 176, 22–33. [Google Scholar] [CrossRef]
- Brisson, N.; Gary, C.; Justes, E.; Roche, R.; Mary, B.; Ripoche, D.; Zimmer, D.; Sierra, J.; Bertuzzi, P.; Burger, P.; et al. An overview of the crop model stics. Eur. J. Agron. 2003, 18, 309–332. [Google Scholar] [CrossRef]
- Tribouillois, H.; Dürr, C.; Demilly, D.; Wagner, M.H.; Justes, E. Determination of Germination Response to Temperature and Water Potential for a Wide Range of Cover Crop Species and Related Functional Groups. PLoS ONE 2016, 11, e0161185. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055. [Google Scholar] [CrossRef]
- Nosratti, I.; Almaleki, S.; Chauhan, B.S. Seed Germination Ecology of Soldier Thistle (Picnomon acarna): An Invasive Weed of Rainfed Crops in Iran. Weed Sci. 2019, 67, 261–266. [Google Scholar] [CrossRef]
- Payamani, R.; Nosratti, I.; Amerian, M. Variations in the germination characteristics in response to environmental factors between the hairy and spiny seeds of hedge parsley (Torilis arvensis Huds.). Weed Biol. Manag. 2018, 18, 176–183. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Allaby, R.G.; Stevens, C. Domestication as innovation: The entanglement of techniques, technology and chance in the domestication of cereal crops. World Archaeol. 2010, 42, 13–28. [Google Scholar] [CrossRef]
- Kissing Kucek, L.; Riday, H.; Rufener, B.P.; Burke, A.N.; Eagen, S.S.; Ehlke, N.; Krogman, S.; Mirsky, S.B.; Reberg-Horton, C.; Ryan, M.R. Pod Dehiscence in Hairy Vetch (Vicia villosa Roth). Front. Plant Sci. 2020, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Parker, T.A.; Mier y Teran, J.C.B.; Palkovic, A.; Jernstedt, J.; Gepts, P. Genetic control of pod dehiscence in domesticated common bean: Associations with range expansion and local aridity conditions. bioRxiv 2019. [Google Scholar] [CrossRef]
- Nosratti, I.; Soltanabadi, S.; Honarmand, S.J.; Chauhan, B.S. Environmental factors affect seed germination and seedling emergence of invasive Centaurea balsamita. Crop Pasture Sci. 2017, 68, 583–589. [Google Scholar] [CrossRef]
- Dürr, C.; Dickie, J.B.; Yang, X.Y.; Pritchard, H.W. Ranges of critical temperature and water potential values for the germination of species worldwide: Contribution to a seed trait database. Agric. For. Meteorol. 2015, 200, 222–232. [Google Scholar] [CrossRef]
- Mischler, R.; Duiker, S.W.; Curran, W.S.; Wilson, D. Hairy vetch management for no-till organic corn production. Agron. J. 2010, 102, 355–362. [Google Scholar] [CrossRef]
- Keene, C.; Curran, W.; Wallace, J.; Ryan, M.; Mirsky, S.; VanGessel, M.; Barbercheck, M. Cover crop termination timing is critical in organic rotational no-till systems. Agron. J. 2017, 109, 272–282. [Google Scholar] [CrossRef]
- Bekker, R.; Bakker, J.; Grandin, U.; Kalamees, R.; Milberg, P.; Poschlod, P.; Thompson, K.; Willems, J. Seed size, shape and vertical distribution in the soil: Indicators of seed longevity. Funct. Ecol. 1998, 12, 834–842. [Google Scholar] [CrossRef]
- Renzi, J.P.; Chantre, G.R.; Cantamutto, M.A. Development of a thermal-time model for combinational dormancy release of hairy vetch (Vicia villosa ssp. villosa). Crop Pasture Sci. 2014, 65, 470–478. [Google Scholar] [CrossRef]
- De Morais, L.F.; Deminicis, B.B.; de Pádua, F.T.; Morenz, M.J.; Araujo, R.P.; de Nepomuceno, D.D. Methods for breaking dormancy of seeds of tropical forage legumes. Am. J. Plant Sci. 2014, 5, 46580. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Smýkal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar]
- Soltani, E.; Gruber, S.; Oveisi, M.; Salehi, N.; Alahdadi, I.; Javid, M.G. Water stress, temperature regimes and light control induction, and loss of secondary dormancy in Brassica napus L. seeds. Seed Sci. Res. 2017, 27, 217–230. [Google Scholar] [CrossRef]
- Huang, S.; Gruber, S.; Stockmann, F.; Claupein, W. Dynamics of dormancy during seed development of oilseed rape (Brassica napus L.). Seed Sci. Res. 2016, 26, 245–253. [Google Scholar] [CrossRef]
- Gorecki, M.; Long, R.; Flematti, G.; Stevens, J. Parental environment changes the dormancy state and karrikinolide response of Brassica tournefortii seeds. Ann. Bot. 2012, 109, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Vercellino, R.B.; Pandolfo, C.E.; Cerrota, A.; Cantamutto, M.; Presotto, A. The roles of light and pericarp on seed dormancy and germination in feral Raphanus sativus (Brassicaceae). Weed Res. 2019, 59, 396–406. [Google Scholar] [CrossRef]
- Malik, M.S.; Norsworthy, J.K.; Riley, M.B.; Bridges, W. Temperature and light requirements for wild radish (Raphanus raphanistrum) germination over a 12-month period following maturation. Weed Sci. 2010, 58, 136–140. [Google Scholar] [CrossRef]
- Adkins, S.W.; Bellairs, S.M.; Loch, D.S. Seed dormancy mechanisms in warm season grass species. Euphytica 2002, 126, 13–20. [Google Scholar] [CrossRef]
- Stump, W.L.; Westra, P. The seedbank dynamics of feral rye (Secale cereale). Weed Technol. 2000, 14, 7–14. [Google Scholar] [CrossRef]
- Goggin, D.E.; Steadman, K.J.; Emery, R.J.N.; Farrow, S.C.; Benech-Arnold, R.L.; Powles, S.B. ABA inhibits germination but not dormancy release in mature imbibed seeds of Lolium rigidum Gaud. J. Exp. Bot. 2009, 60, 3387–3396. [Google Scholar] [CrossRef]
- Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar]
- Poljakoff-Mayber, A.; Popilevski, I.; Belausov, E.; Ben-Tal, Y. Involvement of phytohormones in germination of dormant and non-dormant oat (Avena sativa L.) seeds. Plant Growth Regul. 2002, 37, 7–16. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Hernández-González, M.; Fernández-Pascual, E.; Toorop, P.; Frischie, S.; Gálvez-Ramírez, C. Germination ecology of winter annual grasses in Mediterranean climates: Applications for soil cover in olive groves. Agric. Ecosyst. Environ. 2018, 262, 29–35. [Google Scholar] [CrossRef]
- Cordeau, S.; Wayman, S.; Reibel, C.; Strbik, F.; Chauvel, B.; Guillemin, J.P. Effects of drought on weed emergence and growth vary with the seed burial depth and presence of a cover crop. Weed Biol. Manag. 2018, 18, 12–25. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Devine, T.E.; Mosjidis, J.A.; Bellinder, R.R.; Beste, C.E. Growth and development of hairy vetch cultivars in the northeastern United States as influenced by planting and harvesting date. Agron. J. 2004, 96, 1266–1271. [Google Scholar] [CrossRef]
- Sainju, U.M.; Singh, B.P. Nitrogen storage with cover crops and nitrogen fertilization in tilled and nontilled soils. Agron. J. 2008, 100, 619–627. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Abbate, C.; Lombardo, S.; Fontanazza, S.; Pandino, G.; Anastasi, U.; Mauromicale, G. Trifolium subterraneum cover cropping enhances soil fertility and weed seedbank dynamics in a Mediterranean apricot orchard. Agron. Sustain. Dev. 2021, 41, 70. [Google Scholar] [CrossRef]
- Hyvönen, T.; Ketoja, E.; Salonen, J.; Jalli, H.; Tiainen, J. Weed species diversity and community composition in organic and conventional cropping of spring cereals. Agric. Ecosyst. Environ. 2003, 97, 131–149. [Google Scholar] [CrossRef]
- Dabney, S.; Delgado, J.; Reeves, D. Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant 2001, 32, 1221–1250. [Google Scholar] [CrossRef]
- Van Assche, J.A.; Debucquoy, K.L.; Rommens, W.A. Seasonal cycles in the germination capacity of buried seeds of some Leguminosae (Fabaceae). New Phytol. 2003, 158, 315–323. [Google Scholar] [CrossRef]
- Zhang, Q.; Tu, B.; Liu, C.; Liu, X. Pod anatomy, morphology and dehiscing forces in pod dehiscence of soybean (Glycine max (L.) Merrill). Flora 2018, 248, 48–53. [Google Scholar] [CrossRef]
- Volesky, J.; Mowrey, D.; Smith, G. Performance of rose clover and hairy vetch interseeded into Old World bluestem. Rangel. Ecol. Manag./J. Range Manag. Arch. 1996, 49, 448–451. [Google Scholar] [CrossRef]
- Andersen, B.J.; Samarappuli, D.P.; Wick, A.; Berti, M.T. Faba bean and pea can provide late-fall forage grazing without affecting maize yield the following season. Agronomy 2020, 10, 80. [Google Scholar] [CrossRef]
- Vann, R.; Reberg-Horton, S.; Castillo, M.; McGee, R.; Mirsky, S. Winter Pea, Crimson Clover, and Hairy Vetch Planted in Mixture with Small Grains in the Southeast United States. Agron. J. 2019, 111, 805–815. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Lombardo, S.; Fontanazza, S.; Abbate, C.; Pandino, G.; Anastasi, U.; Onofri, A.; Mauromicale, G. Improving soil health, weed management and nitrogen dynamics by Trifolium subterraneum cover cropping. Agron. Sustain. Dev. 2020, 40, 18. [Google Scholar] [CrossRef]
- Liu, Q.; Xu, R.; Yan, Z.; Jin, H.; Cui, H.; Lu, L.; Zhang, D.; Qin, B. Phytotoxic allelochemicals from roots and root exudates of Trifolium pratense. J. Agric. Food Chem. 2013, 61, 6321–6327. [Google Scholar] [CrossRef] [PubMed]
- Wyngaarden, S.L.; Gaudin, A.; Deen, W.; Martin, R.C. Expanding red clover (Trifolium pratense) usage in the corn–soy–wheat rotation. Sustainability 2015, 7, 15487–15509. [Google Scholar] [CrossRef]
- Ross, S.M.; King, J.R.; Izaurralde, R.C.; O’Donovan, J.T. Weed suppression by seven clover species. Agron. J. 2001, 93, 820–827. [Google Scholar] [CrossRef]
- Nichols, P.; Foster, K.; Piano, E.; Pecetti, L.; Kaur, P.; Ghamkhar, K.; Collins, W. Genetic improvement of subterranean clover (Trifolium subterraneum L.). 1. Germplasm, traits and future prospects. Crop Pasture Sci. 2013, 64, 312–346. [Google Scholar] [CrossRef]
- Baresel, J.P.; Nichols, P.; Charrois, A.; Schmidhalter, U. Adaptation of ecotypes and cultivars of subterranean clover (Trifolium subterraneum L.) to German environmental conditions and its suitability as living mulch. Genet. Resour. Crop Evol. 2018, 65, 2057–2068. [Google Scholar] [CrossRef]
- Balfourier, F.; Imbert, C.; Charmet, G. Evidence for phylogeographic structure in Lolium species related to the spread of agriculture in Europe. A cpDNA study. Theor. Appl. Genet. 2000, 101, 131–138. [Google Scholar] [CrossRef]
- Sebastian, J.; Dinneny, J.R. Setaria viridis: A model for understanding panicoid grass root systems. In Genetics and Genomics of Setaria; Springer: Cham, Switzerland, 2017; pp. 177–193. [Google Scholar]
- Bakker, J. Seeds, ecology, biogeography and evolution of dormancy, and germination. CC Baskin & JM Baskin. Plant Ecol. 2001, 152, 204. [Google Scholar]
- Wilson, M.L.; Baker, J.M.; Allan, D.L. Factors Affecting Successful Establishment of Aerially Seeded Winter Rye. Agron. J. 2013, 105, 1868–1877. [Google Scholar] [CrossRef]
- Miedaner, T. Breeding wheat and rye for resistance to Fusarium diseases. Plant Breed. 1997, 116, 201–220. [Google Scholar] [CrossRef]
- Sarwar, M.; Kirkegaard, J. Biofumigation potential of brassicas: II. Effect of environment and ontogeny on glucosinolate production and implications for screening. Plant Soil 1998, 201, 91–101. [Google Scholar] [CrossRef]
- Haramoto, E.R.; Gallandt, E.R. Brassica cover cropping: I. Effects on weed and crop establishment. Weed Sci. 2005, 53, 695–701. [Google Scholar] [CrossRef]
- Krato, C.; Petersen, J. Competitiveness and yield impact of volunteer oilseed rape (Brassica napus) in winter and spring wheat (Triticum aestivum). J. Plant Dis. Prot. 2012, 119, 74–82. [Google Scholar] [CrossRef]
- Gruber, S.; Pekrun, C.; Claupein, W. Population dynamics of volunteer oilseed rape (Brassica napus L.) affected by tillage. Eur. J. Agron. 2004, 20, 351–361. [Google Scholar] [CrossRef]
- Baraibar, B.; Mortensen, D.A.; Hunter, M.C.; Barbercheck, M.E.; Kaye, J.P.; Finney, D.M.; Curran, W.S.; Bunchek, J.; White, C.M. Growing degree days and cover crop type explain weed biomass in winter cover crops. Agron. Sustain. Dev. 2018, 38, 65. [Google Scholar] [CrossRef]
- Tribouillois, H.; Constantin, J.; Justes, E. Analysis and modeling of cover crop emergence: Accuracy of a static model and the dynamic STICS soil-crop model. Eur. J. Agron. 2018, 93, 73–81. [Google Scholar] [CrossRef]
- Cordeau, S.; Guillemin, J.P.; Reibel, C.; Chauvel, B. Weed species differ in their ability to emerge in no-till systems that include cover crops. Ann. Appl. Biol. 2015, 166, 444–455. [Google Scholar] [CrossRef]
- Baumann, D.T.; Bastiaans, L.; Kropff, M.J. Competition and crop performance in a leek–celery intercropping system. Crop Sci. 2001, 41, 764–774. [Google Scholar] [CrossRef]
- Fisher, K.; Momen, B.; Kratochvil, R. Is broadcasting seed an effective winter cover crop planting method? Agron. J. 2011, 103, 472–478. [Google Scholar] [CrossRef]
- Haramoto, E.R. Species, seeding rate, and planting method influence cover crop services prior to soybean. Agron. J. 2019, 111, 1068–1078. [Google Scholar] [CrossRef]
- Brar, G.; Gomez, J.; McMichael, B.; Matches, A.; Taylor, H. Germination of twenty forage legumes as influenced by temperature. Agron. J. 1991, 83, 173–175. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Ryan, M.R.; Teasdale, J.R.; Curran, W.S.; Reberg-Horton, C.S.; Spargo, J.T.; Wells, M.S.; Keene, C.L.; Moyer, J.W. Overcoming weed management challenges in cover crop–based organic rotational no-till soybean production in the eastern United States. Weed Technol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Noland, R.L.; Wells, M.S.; Sheaffer, C.C.; Baker, J.M.; Martinson, K.L.; Coulter, J.A. Establishment and function of cover crops interseeded into corn. Crop Sci. 2018, 58, 863–873. [Google Scholar] [CrossRef]
- Koehler-Cole, K.; Elmore, R.W. Seeding Rates and Productivity of Broadcast Interseeded Cover Crops. Agronomy 2020, 10, 1723. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Wallace, J.M.; Curran, W.S.; Crockett, B.C. Hairy vetch seedbank persistence and implications for cover crop management. Agron. J. 2015, 107, 2391–2400. [Google Scholar] [CrossRef]
- Sauer, T.J.; Hatfield, J.L.; Prueger, J.H. Corn Residue Age and Placement Effects on Evaporation and Soil Thermal Regime. Soil Sci. Soc. Am. J. 1996, 60, 1558–1564. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Wortmann, C. Crop residue removal and soil erosion by wind. J. Soil Water Conserv. 2017, 72, 97A–104A. [Google Scholar] [CrossRef]
- Boyd, N.S.; Brennan, E.B.; Smith, R.F.; Yokota, R. Effect of Seeding Rate and Planting Arrangement on Rye Cover Crop and Weed Growth. Agron. J. 2009, 101, 47–51. [Google Scholar] [CrossRef]
- Gaba, S.; Perronne, R.; Fried, G.; Gardarin, A.; Bretagnolle, F.; Biju-Duval, L.; Colbach, N.; Cordeau, S.; Fernández-Aparicio, M.; Gauvrit, C. Response and effect traits of arable weeds in agro-ecosystems: A review of current knowledge. Weed Res. 2017, 57, 123–147. [Google Scholar] [CrossRef]
- Mahé, I.; Cordeau, S.; Bohan, D.A.; Derrouch, D.; Dessaint, F.; Millot, D.; Chauvel, B. Soil seedbank: Old methods for new challenges in agroecology? Ann. Appl. Biol. 2021, 178, 23–38. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Badalzadeh, A.; Shahraki, A.D. Effect of Hydro-priming and Salinity Stress on Germination Indices of Niger (Guizotia abyssinica Cass.). Acta Univ. Agric. Silvic. Mendel. Brun. 2021, 69, 46. [Google Scholar] [CrossRef]
- Kaya, M.D.; Okçu, G.; Atak, M.; Cıkılı, Y.; Kolsarıcı, Ö. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron. 2006, 24, 291–295. [Google Scholar] [CrossRef]
- Allen, R.; Hollingsworth, L.; Thomas, J. Sunflower planting and emergence with coated seed. Trans. ASAE 1983, 26, 665–0668. [Google Scholar] [CrossRef]
- Snapp, S.; Price, R.; Morton, M. Seed priming of winter annual cover crops improves germination and emergence. Agron. J. 2008, 100, 1506–1510. [Google Scholar] [CrossRef]
- Stassinos, P.M.; Rossi, M.; Borromeo, I.; Capo, C.; Beninati, S.; Forni, C. Enhancement of Brassica napus Tolerance to High Saline Conditions by Seed Priming. Plants 2021, 10, 403. [Google Scholar] [CrossRef]
- Bijanzadeh, E.; Nosrati, K.; Egan, T. Influence of seed priming techniques on germination and emergence of rapeseed (Brassica napus L.). Seed Sci. Technol. 2010, 38, 242–247. [Google Scholar] [CrossRef]
- Willenborg, C.J.; Gulden, R.H.; Johnson, E.N.; Shirtliffe, S.J. Germination characteristics of polymer-coated canola (Brassica napus L.) seeds subjected to moisture stress at different temperatures. Agron. J. 2004, 96, 786–791. [Google Scholar] [CrossRef]
- Begum, N.; Gul, H.; Hamayun, M.; Rahman, I.U.; Ijaz, F.; Sohail, Z.I.; Afzal, A.; Afzal, M.; Ullah, A.; Karim, S. Influence of seed priming with ZnSO and CuSO on germination. Middle-East J. Sci. Res. 2014, 22, 879–885. [Google Scholar]
- Rao, S.; Akers, S.; Ahring, R. Priming Brassica Seed to Improve Emergence under Different Temperatures and Soil Moisture Conditions 1. Crop Sci. 1987, 27, 1050–1053. [Google Scholar] [CrossRef]
- Chin, J.M.; Lim, Y.Y.; Ting, A.S.Y. Biopolymers for biopriming of Brassica rapa seeds: A study on coating efficacy, bioagent viability and seed germination. J. Saudi Soc. Agric. Sci. 2021, 20, 198–207. [Google Scholar] [CrossRef]
- Ahmad, M.; Waraich, E.A.; Hussain, S.; Ayyub, C.M.; Ahmad, Z.; Zulfiqar, U. Improving Heat Stress Tolerance in Camelina sativa and Brassica napus Through Thiourea Seed Priming. J. Plant Growth Regul. 2021, 41, 2886–2902. [Google Scholar] [CrossRef]
- Huang, P.; He, L.; Abbas, A.; Hussain, S.; Hussain, S.; Du, D.; Hafeez, M.B.; Balooch, S.; Zahra, N.; Ren, X. Seed priming with sorghum water extract improves the performance of camelina (Camelina sativa (L.) crantz.) under salt stress. Plants 2021, 10, 749. [Google Scholar] [CrossRef]
- Pimpini, F.; Sambo, P. Seed germination of rocket (Eruca sativa Mill.) as affected by osmo-priming. In Proceedings of the Atti V Giornate Scientifiche SOI, Sirmione, Italy, 28–30 March 2000. [Google Scholar]
- Kaymak, H.Ç.; Güvenç, İ.; Yarali, F.; Dönmez, M.F. The effects of bio-priming with PGPR on germination of radish (Raphanus sativus L.) seeds under saline conditions. Turk. J. Agric. For. 2009, 33, 173–179. [Google Scholar] [CrossRef]
- Gheidary, S.; Akhzari, D.; Pessarakli, M. Effects of salinity, drought, and priming treatments on seed germination and growth parameters of Lathyrus sativus L. J. Plant Nutr. 2017, 40, 1507–1514. [Google Scholar] [CrossRef]
- Kintl, A.; Huňady, I.; Vymyslický, T.; Ondrisková, V.; Hammerschmiedt, T.; Brtnický, M.; Elbl, J. Effect of Seed Coating and PEG-Induced Drought on the Germination Capacity of Five Clover Crops. Plants 2021, 10, 724. [Google Scholar] [CrossRef] [PubMed]
- Kavandi, A.; Jafari, A.A.; Jafarzadeh, M. Effect of seed priming on enhancement of seed germination and seedling growth of annual sainfoin (Onobrychis crista-galli (L.) Lam.) in medium and long-term collections of gene bank. J. Rangel. Sci. 2018, 8, 117–128. [Google Scholar]
- Ahmad, F.; Kamal, A.; Singh, A.; Ashfaque, F.; Alamri, S.; Siddiqui, M.; Khan, M. Seed priming with gibberellic acid induces high salinity tolerance in Pisum sativum through antioxidants, secondary metabolites and up-regulation of antiporter genes. Plant Biol. 2021, 23, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Tsegay, B.A.; Andargie, M. Seed Priming with Gibberellic Acid (GA 3) Alleviates Salinity Induced Inhibition of Germination and Seedling Growth of Zea mays L., Pisum sativum Var. abyssinicum A. Braun and Lathyrus sativus L. J. Crop Sci. Biotechnol. 2018, 21, 261–267. [Google Scholar] [CrossRef]
- Skwarek, M.; Wala, M.; Kołodziejek, J.; Sieczyńska, K.; Lasoń-Rydel, M.; Ławińska, K.; Obraniak, A. Seed Coating with Biowaste Materials and Biocides—Environment-Friendly Biostimulation or Threat? Agronomy 2021, 11, 1034. [Google Scholar] [CrossRef]
- Jisha, K.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Bortolin, G.S.; Teixeira, S.B.; de Mesquita Pinheiro, R.; Ávila, G.E.; Carlos, F.S.; da Silva Pedroso, C.E.; Deuner, S. Seed priming with salicylic acid minimizes oxidative effects of aluminum on Trifolium seedlings. J. Soil Sci. Plant Nutr. 2020, 20, 2502–2511. [Google Scholar] [CrossRef]
- Qiu, Y.; Amirkhani, M.; Mayton, H.; Chen, Z.; Taylor, A.G. Biostimulant Seed Coating Treatments to Improve Cover Crop Germination and Seedling Growth. Agronomy 2020, 10, 154. [Google Scholar] [CrossRef]
- Mondal, S.; Bose, B. Impact of micronutrient seed priming on germination, growth, development, nutritional status and yield aspects of plants. J. Plant Nutr. 2019, 42, 2577–2599. [Google Scholar] [CrossRef]
- Galhaut, L.; de Lespinay, A.; Walker, D.J.; Bernal, M.P.; Correal, E.; Lutts, S. Seed priming of Trifolium repens L. improved germination and early seedling growth on heavy metal-contaminated soil. Water Air Soil Pollut. 2014, 225, 1905. [Google Scholar] [CrossRef]
- Souguir, M.; Hassiba, F.; Hannachi, C. Effect of NaCl priming on seed germination of Tunisian fenugreek (Trigonella foenum-graecum L.) under salinity conditions. J. Stress Physiol. Biochem. 2013, 9, 86–96. [Google Scholar]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biológica Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- M’Sehli, W.; Kallala, N.; Jaleli, K.; Bouallegue, A.; Mhadhbi, H. Monopotassium phosphate (KH2PO4) and salicylic acid (SA) as seed priming in Vicia faba L. and Vicia sativa L. Biosci. J. 2020, 36, 2078–2091. [Google Scholar] [CrossRef]
- Kalsa, K.K.; Abebie, B. Influence of seed priming on seed germination and vigor traits of Vicia villosa ssp. dasycarpa (Ten.). Afr. J. Agric. Res. 2012, 7, 3202–3208. [Google Scholar]
- Yusefi-Tanha, E.; Fallah, S.; Pessarakli, M. Effects of seed priming on growth and antioxidant components of hairy vetch (Vicia villosa) seedlings under chilling stress. J. Plant Nutr. 2019, 42, 428–443. [Google Scholar] [CrossRef]
- Tiryaki, I.; Keles, H. Reversal of the inhibitory effect of light and high temperature on germination of Phacelia tanacetifolia seeds by melatonin. J. Pineal Res. 2012, 52, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Mao, P. Comparative Time-Course Physiological Responses and Proteomic Analysis of Melatonin Priming on Promoting Germination in Aged Oat (Avena sativa L.) Seeds. Int. J. Mol. Sci. 2021, 22, 811. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Kontturi, M.; Peltonen, J. Phosphorus seed coating enhancement on early growth and yield components in oat. Agron. J. 2006, 98, 206–211. [Google Scholar] [CrossRef]
- Lee, K.-A.; Kim, Y.; Alizadeh, H.; Leung, D.W. Protection of Italian ryegrass (Lolium multiflorum L.) seedlings from salinity stress following seed priming with L-methionine and casein hydrolysate. Seed Sci. Res. 2021, 31, 51–59. [Google Scholar] [CrossRef]
- Scott, J.; Mitchell, C.; Blair, G. Effect of nutrient seed coating on the emergence and early growth of perennial ryegrass. Aust. J. Agric. Res. 1985, 36, 221–231. [Google Scholar] [CrossRef]
- Khazaie, H.; Earl, H.; Sabzevari, S.; Yanegh, J.; Bannayan, M. Effects of osmo-hydropriming and drought stress on seed germination and seedling growth of rye (Secale montanum). ProEnvironment Promediu 2013, 6, 496–507. [Google Scholar]
- Riazi, A.; Sharifzadeh, F.; Ahmadi, A. Effect of osmopriming on seeds germination of forage millet. Res. Constr. Agric. Hortic. 2007, 77, 72–82. (In Persian) [Google Scholar]
- Aune, J.B.; Ousman, A. Effect of seed priming and micro-dosing of fertilizer on sorghum and pearl millet in Western Sudan. Exp. Agric. 2011, 47, 419–430. [Google Scholar] [CrossRef]
- Den Hollander, N.; Bastiaans, L.; Kropff, M. Clover as a cover crop for weed suppression in an intercropping design: I. Characteristics of several clover species. Eur. J. Agron. 2007, 26, 92–103. [Google Scholar] [CrossRef]
- Karssen, C.M.; Haigh, A.; Van der Toorn, P.; Weges, R. Physiological mechanisms involved in seed priming. In Recent Advances in the Development and Germination of Seeds; Springer: New York, NY, USA, 1989; pp. 269–280. [Google Scholar]
- Willenborg, C.J.; Wildeman, J.C.; Miller, A.K.; Rossnagel, B.G.; Shirtliffe, S.J. Oat germination characteristics differ among genotypes, seed sizes, and osmotic potentials. Crop Sci. 2005, 45, 2023–2029. [Google Scholar] [CrossRef]
- Eriksson, O. Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis. Acta Oecol. 1999, 20, 61–66. [Google Scholar] [CrossRef]
- Giri, G.S.; Schillinger, W.F. Seed priming winter wheat for germination, emergence, and yield. Crop Sci. 2003, 43, 2135–2141. [Google Scholar] [CrossRef]
- Parera, C.A.; Cantliffe, D.J. Presowing seed priming. Hortic. Rev. 1994, 16, 109–141. [Google Scholar]
- Dutta, P. Seed priming: New vistas and contemporary perspectives. In Advances in Seed Priming; Springer: Singapore, 2018; pp. 3–22. [Google Scholar]
- Nawaz, J.; Hussain, M.; Jabbar, A.; Nadeem, G.A.; Sajid, M.; Subtain, M.U.; Shabbir, I. Seed priming a technique. Int. J. Agric. Crop Sci. 2013, 6, 1373. [Google Scholar]
- Pedrini, S.; Merritt, D.J.; Stevens, J.; Dixon, K. Seed coating: Science or marketing spin? Trends Plant Sci. 2017, 22, 106–116. [Google Scholar] [CrossRef]
- Amirkhani, M.; Netravali, A.N.; Huang, W.; Taylor, A.G. Investigation of soy protein–based biostimulant seed coating for broccoli seedling and plant growth enhancement. HortScience 2016, 51, 1121–1126. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]


{kind=link}
| Cover Crop Species | Dominant Dormancy Pattern | Main Method of Breaking Dormancy | References |
|---|---|---|---|
| Fabaceae | |||
| Vicia spp., Trifolium spp., Lathyrus sativus, Pisum sativum, Melilotus officinalis, Lupinus spp., Faba spp., and Eruca sativa | Physical (hard seed), physiological | Mechanical abrasion, after-ripening | [54,55,56,57] |
| Brassicaceae | |||
| Brassica spp. | Induced secondary dormancy | Alternating temperatures and the presence of light | [58,59,60] |
| Raphanus sativus | Mechanical resistance and non-leachable chemical inhibitors associated with the pericarp | Dry storage | [61,62] |
| Poaceae | |||
| Sorghum spp. | Seed covering structures (mechanical, permeable, and chemical barriers) | Removal of seed coat structures | [63] |
| Secale cereale | limited innate and induced dormancy | [64] | |
| Lolium spp. | Non-deep physiological dormancy | Chilling at low temperatures and dry after-ripening | [65,66] |
| Avena spp. | High temperature dormancy | After-ripening in dry storage at temperatures higher than 20 °C | [66,67] |
| Setaria spp. | Presence of germination inhibitors in the seed coat | Seed coats removed | [68] |
| Aegilops spp., Anisantha spp., Anisantha spp., Bromus spp., Hordeum spp., and Trachynia spp. | Non-deep physiological dormancy | High temperatures through dry after-ripening | [68] |
| Cover Crop Species | 1000-Seed Weight (mg) § | Seed Treatment | |
|---|---|---|---|
| Priming | Coating | ||
| Guizotia abyssinica | 3.3 | + [112] | Unknown |
| Helianthus annuus | 48.0 | + [113] | + [114] |
| Brassica carinata | 5.0 | Unknown | Unknown |
| Brassica juncea | 3.0 | + [115] | Unknown |
| Brassica napus | 2.7 | + [116,117] | + [118] |
| Brassica rapa | 3.7 | + [119,120] | + [121] |
| Camelina sativa | 1.3 | + [122,123] | Unknown |
| Eruca sativa | 1.3 | + [124] | Unknown |
| Raphanus sativus | 13.0 | + [125] | Unknown |
| Sinapis alba | 8.0 | Unknown | Unknown |
| Lathyrus sativus | 176.0 | + [126] | Unknown |
| Lens nigricans | 21.5 | Unknown | Unknown |
| Lupinus angustifolius | 179.4 | Unknown | Unknown |
| Medicago lupulina | 1.5 | Unknown | + [127] |
| Melilotus officinalis | 2.5 | Unknown | Unknown |
| Onobrychis viciifolia | 23.0 | + [128] | + [127] |
| Pisum sativum | 168.8 | + [129,130] | + [131] |
| Trifoliumalexandrinum | 3.0 | + [132] | Unknown |
| Trifolium incarnatum | 4.7 | Unknown | Unknown |
| Trifolium hybridum | 0.83 | Unknown | |
| Trifolium resupinatum | 1.48 | Unknown | Unknown |
| Trifolium pratense | 2.04 | + [133] | + [134] |
| Trifolium subterraneum | 6.28 | + [135] | Unknown |
| Trifolium repense | 075 | + [136] | + [127] |
| Trigonella foenum graecum | 16.0 | + [137] | Unknown |
| Vicia faba | 359.6 | + [138] | Unknown |
| Vicia sativa | 53.8 | + [139] | Unknown |
| Vicia villosa | 26.7 | + [140,141] | Unknown |
| Phacelia tanacetifolia | 1.8 | + [142] | Unknown |
| Avena sativa | 39.4 | + [143] | + [144] |
| Lolium hybridum | 3.4 | Unknown | Unknown |
| Lolium multiflorum | 2.7 | + [145] | + [146] |
| Secale cereale | 32.3 | + [147] | Unknown |
| Secale multicaule | 18.8 | Unknown | Unknown |
| Setaria italica | 2.2 | + [148] | Unknown |
| Sorghum sudanense | 13.8 | + [149] | Unknown |
| Fagopyrum esculentum | 25.0 | Unknown | Unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nosratti, I.; Korres, N.E.; Cordeau, S. Knowledge of Cover Crop Seed Traits and Treatments to Enhance Weed Suppression: A Narrative Review. Agronomy 2023, 13, 1683. https://doi.org/10.3390/agronomy13071683
Nosratti I, Korres NE, Cordeau S. Knowledge of Cover Crop Seed Traits and Treatments to Enhance Weed Suppression: A Narrative Review. Agronomy. 2023; 13(7):1683. https://doi.org/10.3390/agronomy13071683
Chicago/Turabian StyleNosratti, Iraj, Nicholas E. Korres, and Stéphane Cordeau. 2023. "Knowledge of Cover Crop Seed Traits and Treatments to Enhance Weed Suppression: A Narrative Review" Agronomy 13, no. 7: 1683. https://doi.org/10.3390/agronomy13071683
APA StyleNosratti, I., Korres, N. E., & Cordeau, S. (2023). Knowledge of Cover Crop Seed Traits and Treatments to Enhance Weed Suppression: A Narrative Review. Agronomy, 13(7), 1683. https://doi.org/10.3390/agronomy13071683