
The Ammonium/Nitrate Ratio Affects the Growth and Shikonin Accumulation in Arnebia euchroma
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Regents
2.2. Medium Formulation
2.3. Experimental Design
2.4. Mass Weight Measurement and Root Morphology Analysis
2.5. Quantification of Shikonin Compounds
2.6. Determination of NO3-N and NH2-N Contents
2.7. RNA Extraction and qRT-PCR Analysis
2.8. Sample Preparation for Hormone Determination
2.9. Quantification of Hormones in AEHR
2.10. Statistical Analysis
3. Results
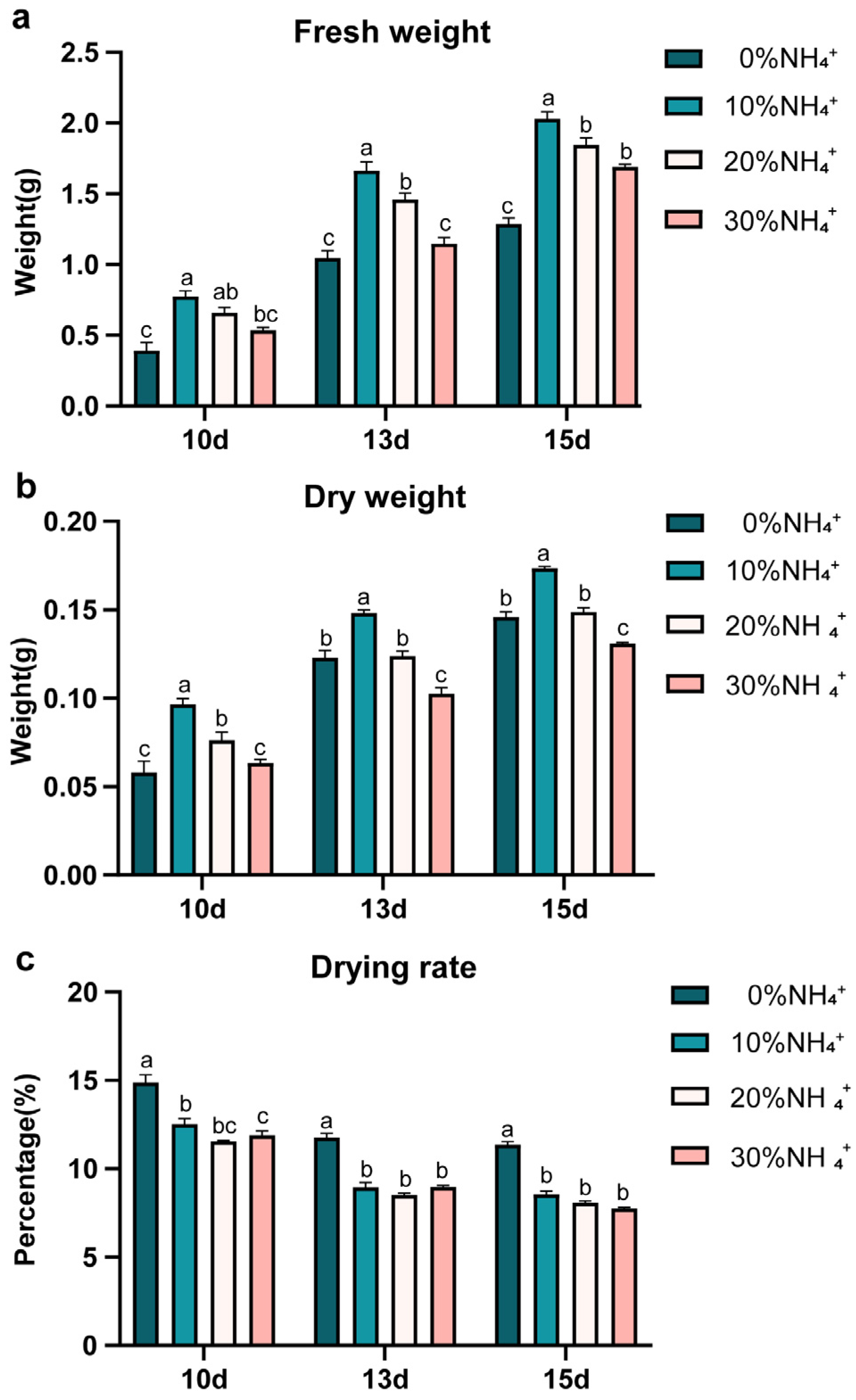
3.1. Growth of AEHR in Response to Ammonium
3.2. Accumulation of Shikonin in AEHR in Response to Ammonium
3.3. Development and Morphology of AEHR in Reponse to Ammonium
3.4. Nitrogen Assimilation in AEHR under Short-Term Ammonium Treatments
3.5. Shikonin Biosynthesis in AEHR under Short-Term Ammonium Treatments
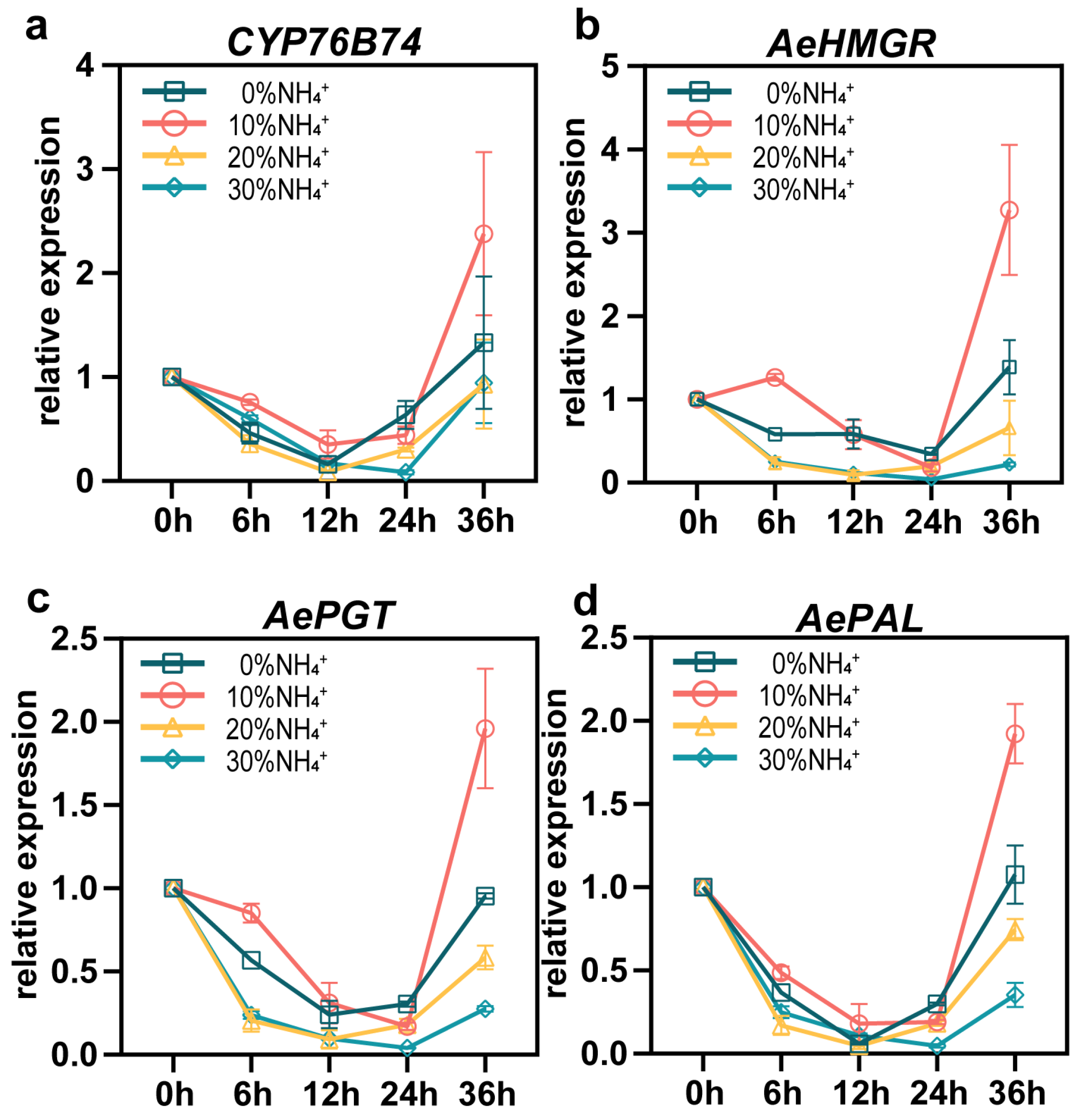
3.6. Gene Expression of Key Enzymes in the Shikonin Biosynthesis Pathway
3.7. Hormone Metabolism of AEHR under NH4+ Toxicity
4. Conclusions
5. Discussion
5.1. Arnebiae Euchroma Is an Ammonium-Sensitive Species
5.2. Compare to a Sole Nitrate Nitrogen Source, A. euchroma Prefers an Appropriate Ammonium/Nitrate Ratio
5.3. Auxin and Cytokinin Might Regulate the Growth and Architecture of AEHR under NH4+ Toxicity
5.4. Shikonin Synthesis in AEHR under NH4+ Toxicity and Its Possible Hormonal Regulation Mechanism
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shindo, S.; Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Matsuo, T. Shikonin Inhibits Inflammatory Cytokine Production in Human Periodontal Ligament Cells. Inflammation 2016, 39, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, F.; DeVico, A.L.; Yarchoan, R.; Redfield, R.; Cleghorn, F.; Blattner, W.A.; Garzino-Demo, A.; Colombini-Hatch, S.; Margolis, D.; Gallo, R.C. Higher macrophage inflammatory protein (MIP)-1alpha and MIP-1beta levels from CD8+ T cells are associated with asymptomatic HIV-1 infection. Proc. Natl. Acad. Sci. USA 2000, 97, 13812–13817. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Bai, L.; Gong, R.; Chuan, J.; Duan, X.; Zhu, Y. Shikonin Protects PC12 Cells Against β-amyloid Peptide-Induced Cell Injury Through Antioxidant and Antiapoptotic Activities. Sci. Rep. 2018, 8, 26. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, Z.; Wang, J.; Li, W.; Liu, G.; Zhang, B.; Tang, Y. Insights into the mechanism of Arnebia euchroma on leukemia via network pharmacology approach. BMC Complement. Med. Ther. 2020, 20, 322. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.Z.; Baik, K.U.; Kweon, G.R.; Lim, K.; Hwang, B.D. Acylshikonin analogues: Synthesis and inhibition of DNA topoisomerase-I. J. Med. Chem. 1995, 38, 1044–1047. [Google Scholar] [CrossRef]
- Yazaki, K. Lithospermum erythrorhizon cell cultures: Present and future aspects. Plant Biotechnol. 2017, 34, 131–142. [Google Scholar] [CrossRef]
- Papageorgiou, V.P.; Assimopoulou, A.N.; Couladouros, E.A.; Hepworth, D.; Nicolaou, K.C. The Chemistry and Biology of Alkannin, Shikonin, and Related Naphthazarin Natural Products. Angew. Chem. Int. Ed. Engl. 1999, 38, 270–301. [Google Scholar] [CrossRef]
- Wang, S.; Wang, R.; Liu, T.; Lv, C.; Liang, J.; Kang, C.; Zhou, L.; Guo, J.; Cui, G.; Zhang, Y.; et al. CYP76B74 Catalyzes the 3″-Hydroxylation of Geranylhydroquinone in Shikonin Biosynthesis. Plant Physiol. 2019, 179, 402–414. [Google Scholar] [CrossRef]
- Tabata, M.; Mizukami, H.; Hiraoka, N.; Konoshima, M. Pigment formation in callus cultures of Lithospermum erythrorhizon. Phytochemistry 1974, 13, 927–932. [Google Scholar] [CrossRef]
- Fu, S.L.; Shang, T.M.; Xiao, P.G. Analysis of naphthoquinone pigments in several Arnebiae Radix. Acta Pharm. Sin. 1984, 19, 921–925. [Google Scholar] [CrossRef]
- Fujita, Y.; Hara, Y.; Ogino, T.; Suga, C. Production of shikonin derivatives by cell suspension cultures of Lithospermum erythrorhizon: I. Effects of nitrogen sources on the production of shikonin derivatives. Plant Cell Rep. 1981, 1, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Arghavani, P.; Haghbeen, K.; Mousavi, A. Enhancement of Shikalkin Production in Arnebia euchroma Callus by a Fungal Elicitor, Rhizoctonia solani. Iran. J. Biotechnol. 2015, 13, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Zou, A.; Zhao, H.; Wu, F.; Zhu, Y.; Zhao, H.; Liao, Y.; Tang, R.J.; Pang, Y.; Yang, R.; et al. Transgenic studies reveal the positive role of LeEIL-1 in regulating shikonin biosynthesis in Lithospermum erythrorhizon hairy roots. BMC Plant. Biol. 2016, 16, 121. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, V.; Ryu, D.D. Improvement of shikonin productivity in Lithospermum erythrorhizon cell culture by alternating carbon and nitrogen feeding strategy. Biotechnol. Bioeng. 1993, 42, 793–799. [Google Scholar] [CrossRef]
- Tani, M.; Takeda, K.; Yazaki, K.; Tabata, M. Effects of oligogalacturonides on biosynthesis of shikonin in Lithospermum cell cultures. Phytochemistry 1993, 34, 1285–1290. [Google Scholar] [CrossRef]
- Zhao, H.; Baloch, S.K.; Kong, L.R.; Zhang, W.J.; Zou, A.L.; Wang, X.M.; Qi, J.L.; Yang, Y.H. Molecular cloning, characterization, and expression analysis of LeMYB1 from Lithospermum erythrorhizon. Biol. Plant. 2014, 58, 436–444. [Google Scholar] [CrossRef]
- Sun, M.H.; Lu, X.P.; Cao, X.J.; Li, J.; Xiong, J.; Xie, X.X. Effect of different forms of nitrogen on the activity of nitrate reductase and expression of the relative genes in Citrus sinensis ×Poncirus trifoliate. J. Fruit Sci. 2017, 34, 410–417. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, K.C.; Zheng, C.X.; Zhu, B.C.; Wang, Y.L.; Yang, J.W.; Wang, W. Effect of different nitrogenous forms on growth and chemical component in tuber of Pinellia pedatisecta Schott. J. Plant Nutr. Fertil. 2014, 20, 1038–1043. [Google Scholar] [CrossRef]
- Crawford, N.M.; Glass, A.D.M. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998, 3, 389–395. [Google Scholar] [CrossRef]
- Migocka, M.; Warzybok, A.; Kłobus, G. The genomic organization and transcriptional pattern of genes encoding nitrate transporters 1 (NRT1) in cucumber. Plant Soil 2012, 364, 245–260. [Google Scholar] [CrossRef]
- Wang, S.; Guo, L.P.; Xie, T.; Yang, J.; Tang, J.F.; Li, X.; Wang, X.; Huang, L.Q. Different secondary metabolic responses to MeJA treatment in shikonin-proficient and shikonin-deficient cell lines from Arnebia euchroma (Royle) Johnst. Plant Cell Tissue Organ Cult. PCTOC 2014, 119, 587–598. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Tellabati, M.; Nelli, R.K.; White, G.A.; Perez, B.B.; Sebastian, S.; Slomka, M.J.; Brookes, S.M.; Brown, I.H.; Dunham, S.P.; et al. 18S rRNA is a reliable normalisation gene for real time PCR based on influenza virus infected cells. Virol. J. 2012, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.F.; Tsai, C.L.; Tsai, C.Y. Interaction of potassium with the form and amount of nitrogen nutrition on growth and nitrogen uptake of maize. J. Plant Nutr. 1992, 15, 23–33. [Google Scholar] [CrossRef]
- Song, W.; Zhuang, Y.; Liu, T. CYP82AR Subfamily Proteins Catalyze C-1′ Hydroxylations of Deoxyshikonin in the Biosynthesis of Shikonin and Alkannin. Org. Lett. 2021, 23, 2455–2459. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, K.; Kunihisa, M.; Fujisaki, T.; Sato, F. Geranyl diphosphate:4-hydroxybenzoate geranyltransferase from Lithospermum erythrorhizon. Cloning and characterization of a ket enzyme in shikonin biosynthesis. J. Biol. Chem. 2002, 277, 6240–6246. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. Int. J. Exp. Plant Biol. 2016, 248, 92–101. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Fukui, H.; Tabata, M. Effect of gibberellin A3 on shikonin production in Lithospermum callus cultures. Phytochemistry 1986, 25, 621–622. [Google Scholar] [CrossRef]
- Kumar, P.; Saini, M.; Bhushan, S.; Warghat, A.R.; Pal, T.; Malhotra, N.; Sood, A. Effect of salicylic acid on the activity of PAL and PHB geranyltransferase and shikonin derivatives production in cell suspension cultures of Arnebia euchroma (Royle) Johnst—A medicinally important plant species. Appl. Biochem. Biotechnol. 2014, 173, 248–258. [Google Scholar] [CrossRef]
- Zhao, H.; Chang, Q.S.; Zhang, D.X.; Fang, R.J.; Zhao, H.; Wu, F.Y.; Wang, X.M.; Lu, G.H.; Qi, J.L.; Yang, Y.H. Overexpression of LeMYB1 enhances shikonin formation by up-regulating key shikonin biosynthesis-related genes in Lithospermum erythrorhizon. Biol. Plant. 2015, 59, 429–435. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, N.; Gao, K.; Chen, F.; Yuan, L.; Mi, G. Ammonium inhibits primary root growth by reducing the length of meristem and elongation zone and decreasing elemental expansion rate in the root apex in Arabidopsis thaliana. PLoS ONE 2013, 8, e61031. [Google Scholar] [CrossRef]
- Chen, H.; Jia, Y.; Xu, H.; Wang, Y.; Zhou, Y.; Huang, Z.; Yang, L.; Li, Y.; Chen, L.-S.; Guo, J. Ammonium nutrition inhibits plant growth and nitrogen uptake in citrus seedlings. Sci. Hortic. 2020, 272, 109526. [Google Scholar] [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate Addition Alleviates Ammonium Toxicity Without Lessening Ammonium Accumulation, Organic Acid Depletion and Inorganic Cation Depletion in Arabidopsis thaliana Shoots. Plant Cell Physiol. 2012, 53, 577–591. [Google Scholar] [CrossRef]
- Hessini, K.; Kronzucker, H.J.; Abdelly, C.; Cruz, C. Drought stress obliterates the preference for ammonium as an N source in the C4 plant Spartina alterniflora. J. Plant Physiol. 2017, 213, 98–107. [Google Scholar] [CrossRef]
- Hill, T.D.; Sommer, N.R.; Kanaskie, C.R.; Santos, E.A.; Oczkowski, A.J. Nitrogen uptake and allocation estimates for Spartina alterniflora and Distichlis spicata. J. Exp. Mar. Biol. Ecol. 2018, 21, 466–472. [Google Scholar] [CrossRef]
- Garnica, M.; Houdusse, F.; Zamarreno, A.M.; Garcia-Mina, J.M. The signal effect of nitrate supply enhances active forms of cytokinins and indole acetic content and reduces abscisic acid in wheat plants grown with ammonium. J. Plant Physiol. 2010, 167, 1264–1272. [Google Scholar] [CrossRef]
- Petersen, F.; Demann, J.; Restemeyer, D.; Ulbrich, A.; Olfs, H.W.; Westendarp, H.; Appenroth, K.J. Influence of the Nitrate-N to Ammonium-N Ratio on Relative Growth Rate and Crude Protein Content in the Duckweeds Lemna minor and Wolffiella hyalina. Plants 2021, 10, 1741. [Google Scholar] [CrossRef]
- Mantovani, C.; Prado, R.M.; Pivetta, K.F.L. Impact of Nitrate and Ammonium ratio on Nutrition and Growth of two Epiphytic Orchids. An. Acad. Bras. Cienc. 2018, 90, 3423–3431. [Google Scholar] [CrossRef]
- Carr, N.F.; Boaretto, R.M.; Mattos, D., Jr. Coffee seedlings growth under varied NO3−:NH4+ ratio: Consequences for nitrogen metabolism, amino acids profile, and regulation of plasma membrane H+-ATPase. Plant Physiol. Biochem. 2020, 154, 11–20. [Google Scholar] [CrossRef]
- Hao, H.; Lei, C.; Dong, Q.; Shen, Y.; Chi, J.; Ye, H.; Wang, H. Effects of exogenous methyl jasmonate on the biosynthesis of shikonin derivatives in callus tissues of Arnebia euchroma. Appl. Biochem. Biotechnol. 2014, 173, 2198–2210. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.S.; Li, H.L.; Wu, Y.L.; Lu, D.Y. Studies on cell growth and Product Synthesis in Production Stage Culture of Arnebia euchroma Cell II: Fermentation culture. Nat. Prod. Res. Dev. 2003, 15, 429–432. [Google Scholar] [CrossRef]
- Ge, F.; Yuan, X.; Wang, X.; Zhao, B.; Wang, Y. Cell growth and shikonin production of Arnebia euchroma in a periodically submerged airlift bioreactor. Biotechnol. Lett. 2006, 28, 525–529. [Google Scholar] [CrossRef]
- Malik, S.; Bhushan, S.; Sharma, M.; Ahuja, P.S. Physico-chemical factors influencing the shikonin derivatives production in cell suspension cultures of Arnebia euchroma (Royle) Johnston, a medicinally important plant species. Cell Biol. Int. 2011, 35, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Bhushan, S.; Verma, S.C.; Sharma, N.; Sinha, A.K.; Sharma, M.; Ahuja, P.S. Production of naphthoquinone pigments in cell suspension cultures of Arnebia euchroma (Royle) Johnston: Influence of pH on growth kinetics and acetylshikonin. Med. Aromat. Plant Sci. Biotechnol. 2008, 2, 43–49. [Google Scholar]
- Liang, J.W.; Li, T.; Wang, R.S.; Zhou, L.; Yang, Q.; Wang, S.; Guo, L.P. Establishment of RNA interfered hairy root system of two CYP450 genes in Arnebia euchroma and its influence. China J. Chin. Mater. Med. 2020, 45, 3422–3431. [Google Scholar] [CrossRef]
- Lu, W.H.; Chen, Y.F.; Wang, F.; Dai, N.B.; Hao, A.H.; Li, C.F.; Jia, S.E. Effects of physical and chemical conditions on hairy root culture and content of Arnebia euchromu. J. Huazhong Agric. Univ. 2012, 31, 50–54. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, F.; Zhu, C. Influence of fungal elicitor and macroporous resin on shikonin accumulation in hairy roots of Arnebia euchroma (Royle) Johnst. Chin. J. Biotechnol. 2013, 29, 214–223. [Google Scholar] [CrossRef]
- Yang, Y.H.; Lu, J.; Zhao, Q.H.; Cao, R.Q. Effect of water extract of algae on growth and pigment formation in Lithospermuml erythrorhizon and Arnebia euchromu cell cultures. J. Plant Resour. Environ. 1992, 1, 39–44. [Google Scholar]
- Ge, S.N.; Zhang, S.X.; Wang, F.; Wei, H.; Xie, W.L.; Xie, J.; Li, C.; Zhao, H. Effects of Exogenous Substances on the Content of Secondary Metabolites in Arnebia euchroma Johnst Hairy Roots. Food Sci. 2016, 37, 160–164. [Google Scholar] [CrossRef]
- Li, M.; Pan, Q.; Wang, F.; Zhang, P. Research of Effects on Arnebia euchroma (Royle) Johnst Hairy Roots Growth Factors. Xinjiang Agric. Sci. 2012, 49, 2062–2068. [Google Scholar] [CrossRef]
- Lu, W.H.; Pan, Q.; Wang, F.; Chen, Y.F.; Dai, N.B. Influence of Culture Conditions on Arnebia euchromaHairy Roots Growth and Shikonin Content. Acta Bot. Boreali Occident. Sin. 2012, 32, 1686–1691. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, H.; Cao, R. Effect of Brassinolide on Growth and Shikonin Formation in Cultured Onosma paniculatum Cells. J. Plant Growth Regul. 1999, 18, 89–92. [Google Scholar] [CrossRef]
- Yang, H.; von der Fecht-Bartenbach, J.; Friml, J.; Lohmann, J.U.; Neuh User, B.; Ludewig, U. Auxin-modulated root growth inhibition in Arabidopsis thaliana seedlings with ammonium as the sole nitrogen source. Funct. Plant Biol. 2015, 42, 239–251. [Google Scholar] [CrossRef]
- Dziewit, K.; Pěnčík, A.; Dobrzyńska, K.; Novák, O.; Szal, B.; Podgórska, A. Spatiotemporal auxin distribution in Arabidopsis tissues is regulated by anabolic and catabolic reactions under long-term ammonium stress. BMC Plant Biol. 2021, 21, 602. [Google Scholar] [CrossRef]
- Péret, B.; De Rybel, B.; Casimiro, I.; Benková, E.; Swarup, R.; Laplaze, L.; Beeckman, T.; Bennett, M.J. Arabidopsis lateral root development: An emerging story. Trends Plant Sci. 2009, 14, 399–408. [Google Scholar] [CrossRef]
- Barreto, R.F.; de Mello Prado, R.; Lúcio, J.C.B.; López-Díaz, I.; Carrera, E.; Carvalho, R.F. Ammonium Toxicity Alleviation by Silicon is Dependent on Cytokinins in Tomato cv. Micro-Tom. J. Plant Growth Regul. 2021, 41, 417–428. [Google Scholar] [CrossRef]
- Walch-Liu, P.; Neumann, G.; Bangerth, F.; Engels, C. Rapid effects of nitrogen form on leaf morphogenesis in tobacco. J. Exp. Bot. 2000, 51, 227–237. [Google Scholar] [CrossRef]
- Fang, D.Q.; Hou, C.S.; Li, X.M.; Ye, H.C.; Li, G.F. Effects of pH and hormone on cell growth and synthesis of Shikonin derivatives in suspension culture of Arnebia euchroma. Plant Sci. J. 1994, 2, 159–164. [Google Scholar]
- Zhu, W.H.; Fan, H.X.; Hu, Q.; Zhu, H.Q. Induction culture of callus and selection of superior clones of Onosma paniculatum. J. Chin. Med. Mater. 1990, 1, 6–9. [Google Scholar] [CrossRef]
- Yan, H.Y.; Cao, R.Q. Influencing factors of Shikonin formation in callus culture of Lithospermum erythorhizon. Acta Agric. Boreali Sin. 2002, 17, 116–120. [Google Scholar] [CrossRef]
- Ahmad, M.; Alonso, A.V.; Koletti, A.E.; Rodić, N.; Reichelt, M.; Rödel, P.; Assimopoulou, A.N.; Paun, O.; Declerck, S.; Schneider, C.; et al. Dynamics of alkannin/shikonin biosynthesis in response to jasmonate and salicylic acid in Lithospermum officinale. Sci. Rep. 2022, 12, 17093. [Google Scholar] [CrossRef]
- Ding, J.; Shi, S.; Jiang, B.-H.; Yang, Y.-H.; Huang, J.; Shen, H.-G.; Xia, K.; Zhang, J.; Jiang, X. Effects of Methyl jasmonate with indole-3-acetic acid and 6-benzylaminopurine on the secondary metabolism of cultured Onosma paniculatum cells. Vitr. Cell. Dev. Biol. Plant 2004, 40, 581–585. [Google Scholar] [CrossRef]
- Zhu, Y. Cloning, Expression and Functional Analysis of LeTCP4 Gene in Lithospermum Erythrorhizon. Master’s Thesis, Nanjing University, Nanjing, China, 2012. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Step | Medium | NH4+ (mM) | NO3− (mM) | Total Content (mM) | NH4+/NO3− Ratio | References |
|---|---|---|---|---|---|---|---|
| L. erythorhizon suspension cell cultures | Two-step for growth | LS | 20.61 | 39.4 | 60.01 | 34.34% | [11] |
| MG-5 | 6.25 | 54.22 | 60.47 | 10.34% | [6] | ||
| Two-step for shikonin production | White | 0 | 3.33 | 3.33 | 0.00% | [11] | |
| M9 | 0 | 6.67 | 6.67 | 0.00% | [11] | ||
| L. erythorhizon hairy roots | Two-step for growth | B5 | 2.02 | 24.73 | 26.75 | 7.55% | [16,31] |
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [16,31] | |
| A. erchroma suspension cell cultures | One-step | AG-7 | 7 | 18.79 | 25.79 | 27.14% | [41,42] |
| Two-step for growth | AG-7 | 7 | 18.79 | 25.79 | 27.14% | [42] | |
| LS | 20.61 | 39.4 | 60.01 | 34.34% | [43] | ||
| MS | 20.61 | 39.4 | 60.01 | 34.34% | [44,45] | ||
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [42,46,47,48,49,50,51] | |
| M10 | 0 | 10.97 | 10.97 | 0.00% | [43] | ||
| APM | 0 | 17.74 | 17.74 | 0.00% | [44,45] | ||
| A. erchroma hairy roots | Two-step for growth | SH | 2.61 | 24.73 | 27.34 | 9.55% | [51] |
| SH without NH4+ | 0 | 24.73 | 24.73 | 0.00% | [47,50,52] | ||
| MS without NH4+ | 0 | 18.79 | 18.79 | 0.00% | [47,52] | ||
| B5 without NH4+ | 0 | 24.73 | 24.73 | 0.00% | [47] | ||
| 1/2MS without NH4+ | 0 | 9.395 | 9.395 | 0.00% | [46,52] | ||
| LS without NH4+ | 0 | 18.79 | 18.79 | 0.00% | [47] | ||
| MG-5 without NH4+ | 0 | 47.97 | 47.97 | 0.00% | [52] | ||
| MS | 20.61 | 39.4 | 60.01 | 34.34% | [47] | ||
| B5 | 2.02 | 24.73 | 26.75 | 7.55% | [49] | ||
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [47,50] | |
| Onosma paniculatum suspension cell cultures | Two-step for growth | B5 | 2.02 | 24.73 | 26.75 | 7.55% | [53] |
| Two-step for shikonin production | M9 | 0 | 6.67 | 6.67 | 0.00% | [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Liang, J.; Wang, R.; Wan, X.; Yan, B.; Zhang, Y.; Chen, M.; Liu, C.; Li, Q.; Wang, S.; et al. The Ammonium/Nitrate Ratio Affects the Growth and Shikonin Accumulation in Arnebia euchroma. Agronomy 2023, 13, 1318. https://doi.org/10.3390/agronomy13051318
Shi L, Liang J, Wang R, Wan X, Yan B, Zhang Y, Chen M, Liu C, Li Q, Wang S, et al. The Ammonium/Nitrate Ratio Affects the Growth and Shikonin Accumulation in Arnebia euchroma. Agronomy. 2023; 13(5):1318. https://doi.org/10.3390/agronomy13051318
Chicago/Turabian StyleShi, Linyuan, Jiuwen Liang, Ruishan Wang, Xiufu Wan, Binbin Yan, Yan Zhang, Meilan Chen, Changzheng Liu, Qi Li, Sheng Wang, and et al. 2023. "The Ammonium/Nitrate Ratio Affects the Growth and Shikonin Accumulation in Arnebia euchroma" Agronomy 13, no. 5: 1318. https://doi.org/10.3390/agronomy13051318
APA StyleShi, L., Liang, J., Wang, R., Wan, X., Yan, B., Zhang, Y., Chen, M., Liu, C., Li, Q., Wang, S., & Guo, L. (2023). The Ammonium/Nitrate Ratio Affects the Growth and Shikonin Accumulation in Arnebia euchroma. Agronomy, 13(5), 1318. https://doi.org/10.3390/agronomy13051318