Abstract
High-yielding cultivars with high water use efficiency are a main target in maize breeding; yet, no comprehensive study about the genetic regulation of photosynthesis, or about the ranges of variability for gas exchange-related parameters in maize has been published. Here, a large maize panel of 731 inbred lines previously genotyped was evaluated to elucidate the genetic basis of photosynthesis-related parameters, measured 15–30 days after silking, across three years. Large phenotypic and genotypic variations were observed in this panel with dramatic fluctuations in heritability for various traits. We detected 27 minor Quantitative Trait Loci (QTL), comprising 39 significant trait-Single Nucleotide Polymorphisms (SNPs), located mostly on chromosomes 5 and 8. Most candidates genes were novel, though a few of them were functionally associated with the theory of source-to-sink translocation presented in previous studies. Genomic selection of favorable alleles to enhance photosynthesis, along with other tools, could be a practical, and a promising, approach in the future. Leaf net photosynthesis and stomatal conductance are the most promising targets for breeding programs. We did not find any marker associated with intrinsic water use efficiency, which highlights that developing maize cultivars with more efficient use of water by genomic selection is not straightforward; however, given the high heritability value for this trait, phenotypic selection could be implemented.
1. Introduction
Maize, with a global production of 1.1 billion tons in 2020, is the first cereal crop that accounts for 12% of the global production of primary crops. However, the increasing world population, the environmental problems associated with climate change, and the losses of agricultural land due to urbanization and land degradation, require global crop productions to increase. Therefore, increasing global crop production and yields are major challenges of the modern era [1].
Photosynthesis is the principal sunlight storage process on Earth. It has been traditionally accepted that leaf photosynthesis and crop growth and yield are strongly correlated, since carbon, incorporated via photosynthesis, is crucial for biomass production [2,3]. Nowadays, this relationship is considered controversial [4,5,6], but it is important to consider the way photosynthesis is measured. Photosynthesis at canopy (per ground area) level has been proved critical for increasing crop yields, and over the past decades breeding to magnify plant architecture and optimize resource acquisition have led to higher yielding maize varieties [7,8,9,10]. Nonetheless, many researchers consider the “core” chemical process of photosynthesis (i.e., the photosynthetic rate as CO2 fixation per unit leaf area) the only major agronomic trait unimproved by the green revolution. Although genetic variation exists for photosynthesis-related processes, it has been largely ignored in breeding programs until recent years [11,12].
Besides the need to enhance crop yields, the increasing frequency and intensity of drought episodes, associated with climate change, is also a major threat to global food security. In this context, breeding to enhance yields, while using less water, is mandatory. In a general way, water use efficiency (WUE) is considered an indicator of the biomass produced and the associated water cost. It can be used to explore plants’ performance under drought stress, as well as to elucidate if improving yield, while optimizing water consumption, is possible. Improving the structural and functional properties of the photosynthetic machinery, via biotechnological manipulations, can lead to the development of more resource use-efficient varieties that are more tolerant to environmental stresses, including drought [13,14,15]. Greater WUE can enhance crop productivity in water-limited environments [16,17,18]. Since genetic variation exists in traits that can contribute to an increased WUE, searching for sources of genetic variation for such traits is desirable to indirectly improve WUE [19,20,21].
Recently, researchers have begun to use genome-scale analysis to allele mining focusing on genes simultaneously involved in photosynthesis- and yield-related traits. Several genome-wide association analysis studies (GWASs) on C3 plants, like soybean [22,23] and chickpea [3], have found natural variation and novel loci [24] detected 159 quantitative trait loci within 31 genomic regions for four photosynthesis-related traits under different phosphorus stress conditions [25] reported 239 unique candidate genes for photosynthetic traits related to WUE in sorghum. However, these studies were not focused on primary cereal crops [26] presented six clusters of quantitative trait loci for several photosynthesis-related traits recorded at the young leaf stage in 120 maize recombinant inbred lines evaluated in a greenhouse under water deprivation [9] used integrated genome-scale analysis to identify novel genes for the maximum quantum efficiency of PSII (Fv/Fm) and networks underlying senescence in maize. Therefore, maize variability for photosynthesis-related traits and genomic regions implicated in those traits are not well known because GWAS has been rarely used for photosynthesis-related parameters in adult maize plants at post-flowering stage.
Traditional plant breeding led to a great loss of genetic diversity, but there is still a large intra- and inter-specific genetic variation for plant photosynthesis. However, to the best of our knowledge, no comprehensive study about the genetic regulation of gas exchange-related parameters, or about the ranges of variability for gas exchange parameters in maize, has been published to date. Therefore, research efforts must focus on a better understanding of the photosynthetic process and on harnessing this natural variation for breeding programs [6,10,11,27,28]. The use of large diversity panels is ideal to identify the genetic basis underlying variations in photosynthesis-related parameters, and to design specific breeding programs.
Given these perspectives, a large maize panel with 731 accessions [29,30] was used to characterize the loci and candidate genes for five photosynthetic traits measured 15–30 days after silking, across three consecutive years. The analysis results provide new insights into the understanding of natural variation and the genetics of complex photosynthesis-related traits, as well as shed light for further selection strategies to enhance photosynthetic capacity.
2. Results
The photosynthetic traits under study were the following: leaf net photosynthesis (AN), stomatal conductance (gS), transpiration rate (E), intrinsic water use efficiency (WUEi) and instantaneous water use efficiency (WUEinst).
3. Variance Analysis and Heritability
In the current panel, we observed large phenotypic variations for all traits, as is shown in Figure 1. We found significant differences among the six checks for the photosynthetic parameters evaluated (Table 1).
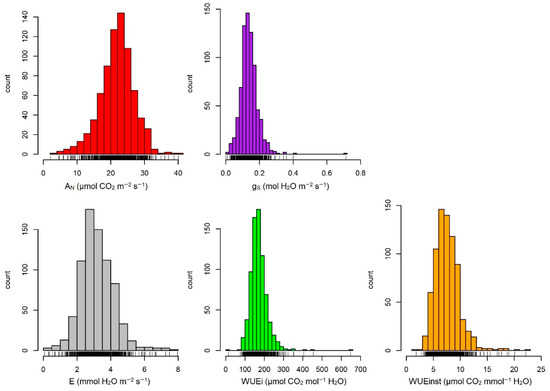
Figure 1.
Histogram of best linear unbiased estimators for photosynthesis-related traits in a large maize association panel across three years. AN: Leaf net photosynthesis; gS: Stomatal conductance; E: Transpiration rate; WUEi: Intrinsic Water use efficiency; WUEinst: Instantaneous Water use efficiency.

Table 1.
Comparison of means (± standard deviations) of the six maize inbred lines used as checks for photosynthetic traits.
Based on combined analyses of variance, significant differences among genotypes and among years were found for AN, gS, and E (Table 2). For WUEi, differences were significant among genotypes, but not among years, while the reverse was true for WUEinst.
Table 2.
Mean squares of the combined analysis of variance of the inbred panel across three years and heritability (± standard deviation) estimates for photosynthetic traits.
In line with the results stated above, heritability was high for AN, gS, E and WUEi (Table 2). Among these traits, we observed low-to-moderately significant Pearson correlations (Table 3). Surprisingly enough, even though the calculation for WUEi is AN/gS; WUEi is negatively and weakly correlated with AN, and AN positively correlated with gS.
Table 3.
Pearson correlation coefficients among best linear unbiased estimators for photosynthetic traits in a large maize association panel evaluated across three consecutive years.
4. Association Mapping: QTLs, SNPs and Candidate Genes
SNPs with p values ≤ 1 × 10−4 were regarded as significant. Given that the causative regulatory variants may be far away from functional genes [31], all significant SNPs within 5 kb for the same trait were declared as the same QTL.
For the five photosynthetic parameters evaluated across three years, we found 27 QTL, comprising 39 significant trait-SNP associations, on ten chromosomes, with half of them located on chromosomes 5 and 8. The QTL number ranged from zero for WUEi to 13 for gS. (Table 4, Supplementary Figure S1). These QTL had minor effects with a contribution to phenotypic variation (R2) ≤ 4%. Most of the observed SNPs were trait-specific. Only two significant SNPs exhibited pleiotropic effects; S8_152322485 for E and WUEinst, and S8_165286201 for gS and E (Table 4). Fifteen of the significant SNPs were also detected for being significant via single environment analysis in two years (2018, 2019) and two of them across the three years.
Table 4.
The characteristics of SNPs most significantly associated to photosynthetic traits within each QTL: bin, p-value, proportion of total variance explained by the SNP (R2), additive effect (across years, 2017, 2018, 2019), and number of lines of the maize association panel homozygous for each allele.
We found 13 QTL for gS on chromosomes 2, 3, 5, 8, and 10. Two QTL were mapped at bin 2.09; one of them comprised eight significant SNPs (Table 4).
We found eight significant SNPs within six QTL at chromosomes 1, 8, and 9 for E. The most significant SNP, S1_29241480, was significant via combined analyses across three years and in all single environment analyses.
Only four significant SNPs for WUEinst were detected across the years analysis and were clustered into three QTL at bins 1.03, 5.05, and 8.06 (Table 4).
Regarding AN, five significant SNPs were simultaneously detected via combined analyses on chromosomes 2, 3, 6, 8, and 9. Four favorable alleles for increasing AN were found in the majority of those inbred. We looked at the distribution of favorable alleles (increased AN) for five of the most significantly associated loci in the combined analysis (Supplementary Figure S2). Eighteen lines showed high AN, and the inbred lines carrying all five favorable alleles had higher AN value than those carrying only one (p < 0.05), indicating that pyramiding favorable alleles with additive effects could enhance AN and could be implemented in marker-assisted selection.
Additionally, we searched for candidate genes within a ±500 bp region around each significant SNP and found 36 candidate genes within 27 QTL for five photosynthetic traits, according to B73 RefGen_v4 (MaizeGDB genome browser [32]). Of these, 16 were annotated as having uncharacterized function (Supplementary Table S1), but most of them displayed functional annotations as genes codifying for tRNA, lincRNA, hydrolase, kinase, and ligase, among others.
5. Discussion
In comparison with previous GWASs, the current study used a larger number of inbred lines at the reproductive stage [9,33] and allowing dissection of genomics [26]. The current study used a larger maize panel, with 731 involved in photosynthesis at the adult plant stage, and the identification of novel genetic and phenotypic variations.
The h2 was high for AN and gS; while E and WUEinst presented high stand errors and quite low heritability. These results are in accordance with the results from the combined variance analysis that showed high influence of the environment (light, temperatures, etc.) and no influence of maize genotype, and are in line with the evidence reported by [26], who found strong environmental effects on physiological performance. Zhao and Zhong [33] estimated lower h2 for AN and gS, but higher for E and WUEi. In our experiment, the different subsets of samples collected from the panel each year (depending on the weather conditions and the availability of resources) could have affected the estimates.
Most loci had minor effects on photosynthetic traits, were unevenly distributed across the ten chromosomes, and were environment-specific; besides which, they only explained a low percentage of the phenotypic variation across years. This is in accordance with the trends for the evaluation of the same panel for cold tolerance [29], which highlights the heavy influence of variable environments and diverse genetic backgrounds on photosynthetic capacity [34]. Similarly, [26] also reported that QTL were environment-specific for drought and control conditions in a limited collection of maize inbred lines. Practical ways to increase photosynthesis should concurrently combine optimization of light capture by the leaf canopy and bridge elite lines with high photosynthesis [35,36].
Little attention has been paid to the genomics involved in photosynthesis-related traits at the maize adult stage, with the scarce QTL previously found co-localized with many QTL uncovered in the current study. Given that two loci around 500 bp across different mapping populations were regarded as one QTL, five QTL mapped to bins 1.03, 5.02, 8.08, and 10.06 in previous studies [9,29]. Sekhon et al. [9] found a highly significant SNP on chromosome 8, lying in gene ZmBLGU42, and putatively encoding a b-glucosidase orthologous to Arabidopsis BGLU42 [37] and rice Os1bglu4 [38]. They emphasized that the differential post-transcriptional activity of ZmBLGU42 alleles underlines differences in senescence phenotype. Zhao and Zhong [33] identified 54 QTL by using SSR in two maize mapping populations; among which thirteen collocated in the same bin of the QTL reported here. Furthermore, the significant SNPs, S2_9085639 (bin 2.02) and S9_123361111 (bin 9.04), associated with AN were co-localized with the QTL presented by Pelleschi et al. [26]), related to net CO2-uptake. However, a large number of novel loci have not been reported in previous studies, which demonstrates that using a large panel could uncover new genetic variation for photosynthesis-related traits in maize. Genomic recurrent selection for environment-specific alleles for a given ecological region could be an effective approach.
The functional annotations of candidate genes indicate a comprehensive genetic basis underlying photosynthesis in maize. Identification of four candidate genes, Zm00001d004448, Zm00001d004125 (ACC1; [39]), Zm00001d007576 (cle14), and Zm00001d004779 (rpl7), in the present study (Supplementary Table S1) support source-sink translocation [40]. As evidence of candidate genes associated with AN, Zm00001d004448, encoding a pectate lyase 8, and ACC1, functionally annotated as acetyl-coenzyme A carboxylase, are involved in cell wall modification [41] and carbohydrate metabolism and remobilization [42], respectively. Notably, a recent study [43] found that ACC1, identified as a central player for defense module 30, interacts with pathways associated with secondary metabolism and microbial carbon and nitrogen metabolism. The third gene, rpl7, found in the confidence interval of the SNP S5_135301104 associated with gS, was also proposed as a candidate gene for hemicellulose QTL [44]. The gene Zm00001d004085, in the confidence interval of S5_79113622 associated with gS, was found to be related with leaf length and width in a maize nested association mapping (NAM) population [45], highlighting the potential association between leaf photosynthetic capacity and leaf size. Accordingly, as observed in a previous study [46]), leaf width shapes intrinsic WUE via coordination of vein and stomatal traits and by affecting stomatal aperture. These associations heavily address the influence of the environment on photosynthetic traits. However, most of the new candidate genes in the current research have not been reported and validated, though the putative functions for the genes were annotated on the public Maizegdb database. None of the candidate genes detected in our study were found by Zhao and Zhong [33], but these authors reported different candidate genes with similar functions to those detected here (photomorphogenesis, photosynthesis, and stress response). These results indicated newly natural variation discovered in a large maize panel and indicated that there are a variable number of genes involved in genetic regulation, depending on genetic background and the environment.
6. Conclusions
Our study expands the knowledge of photosynthesis in adult maize plants by identifying, in a large panel, novel natural genetic variation. Genomic selection for favorable and minor alleles and bridging elite lines with high photosynthesis into commercial lines are recommendations to improve the photosynthetic capacity in a given environment. However, producing high-yielding and photosynthetic-efficient cultivars is a systematic project which also calls for improved heterosis and biotic and abiotic tolerances. We expect that dissecting the genetic basis underlying maize photosynthesis could lay a solid foundation for uncovering its molecular basis in the future. These results suggest that photosynthetic rate and stomatal conductance are the most promising selection targets for phenotypic selection aiming at developing more efficient maize cultivars. Improving water use efficiency is possible, though challenging.
7. Materials and Methods
7.1. Plant Materials
A subset of 836 maize inbred lines from the Ames association panel [30] (provided by the North Central Regional Plant Introduction Station (USA)) were chosen for the current study. The same subset was used in a previous GWAS for cold tolerance [29]. The panel, along with five dent inbred checks, A619, A632, A662, A665, and PH207, and one flint inbred check EP42 [29,47] were cultivated at Misión Biológica de Galicia (Pontevedra, Spain, 42°24′ N, 8°38′ W, 20 m above the sea level).
7.2. Field Experiment and Phenotypic Measurements
All seeds were sown in the experimental field of Misión Biológica de Galicia. All of the sowing details are reported by Yi et al. [29]. The sowing dates were 19 April 2017, 19 April 2018, and 30 April 2019. The average daily accumulated rain during the summer months (June-August) was, 0.68 mm, 1.21 mm and 2.52 mm in 2017, 2018 and 2019, respectively (Information provided by Agencia Estatal de Meterología, Spain [48]). The experiments followed an augmented design with 17 blocks with 50 non-replicated inbred lines and the six checks stated above per block. Each plot contained one row corresponding to one inbred line, 2.4 m long and with 13 plants. Row and planting spacings were 0.8 m and 0.21 m, respectively.
For each plot, days to silking (day) were recorded during maize flowering time, as described by Buckler et al. [49], and photosynthesis-related traits were measured in three plants per plot once in a period of 15–30 days after silking to ensure that all inbred lines were measured at the same developmental stage. Given the wide variability enclosed in this panel, measurement periods expanded from 12 July to 21 August in 2017, 6 July to 31 August in 2018 and 27 June to 5 September in 2019. Survey measurements of leaf net photosynthesis (AN), stomatal conductance (gS) and transpiration rate (E) were recorded on primary ear leaves after equilibration to a steady-state using a portable photosynthesis system equipped with a 6 cm2 LED light source chamber (LI-6400XT and 6400-02B, Li-Cor Inc., Lincoln, NE, USA). Environmental conditions in the leaf chamber were adjusted to match average external conditions during the measurement time as best as possible, minimizing the time taken for gas exchange parameters to stabilize. All measurements were done at a light intensity of 1000 µmol m−2 s−1 and a CO2 concentration in the leaf chamber of 400 µmol m−2 s−1, achieved with a CO2 mixer. Relative humidity and air temperature inside the chamber were not controlled. Measures were recorded on sunny days, from 10:00 a.m. to 2:00 p.m. (average temperature and standard deviation: 25.3 ± 3.4 °C; https://www.worldweatheronline.com/pontevedra-weather-history/galicia/es.aspx (accessed on 3 July 2022)). No midday depression of photosynthesis was observed. Intrinsic Water use efficiency (WUEi) was later calculated as the ratio between AN and gS, and instantaneous water use efficiency (WUEinst) as the ratio between AN and E [50].
The numbers of inbred lines measured each year varied depending on the weather; 248 inbred lines were measured in 2017 and 581 and 565 in 2018 and 2019, respectively. The average values for each plot were used for further analyses. In the end, 731 inbred lines (from the original 836) were investigated in the current study.
7.3. Statistical Analysis Filtering of Genotypic Data and Association Mapping
Considering that the checks followed a randomized complete block design in the presented experiments, combined analysis of variances for each photosynthetic trait were performed using the general linear procedure of SAS software version 9.4 [51]: . The value is the observed value, μ is the mean, is the block effect l within year n, is the year effect n, is the genotype effect m, is the interaction effect between genotype m and year n, and is the residual effect. Aside from taking as the fixed effect, the other factors were considered as random effects. Mean comparisons among checks for each trait were carried out using the Fisher's protected least significant difference (LSD). Heritability (h2) for each trait across three years was calculated using the six checks on a family-mean basis, as described by Holland et al. [52].
For each trait, we calculated the best linear unbiased estimator (BLUE) of each inbred line across environments and for each environment, using the mixed model procedure (PROC MIXED) of SAS software version 9.4 [52]. Inbred lines were considered as a fixed effect, and year, block within the year, and the inbred × year interaction as random factors. We used the BLUEs across all environments to calculate simple Pearson correlation coefficients among traits with PROC CORR in SAS software version 9.4 [52].
The Ames maize inbred line panel [30] was previously genotyped using genotyping-by-sequencing (GBS). Data is publicly available at Panzea (http://www.panzea.org (accessed on 11 September 2019)). For the 731 inbred lines investigated, 152.670 filtered SNPs were obtained after excluding SNPs with >20% missing data and a minor allele frequency < 0.05 in TASSEL 5.2.54 [52]. Using the BLUEs across the years, a GWAS was completed with Tassel 5 based on a mixed linear model using a genotype–phenotype matrix. A kinship matrix, obtained by the centered IBS method, was used to avoid false-positive associations caused by the underlying population structure. The associations between SNPs and photosynthetic traits were obtained based on a mixed linear model (MLM) in Tassel 5.2.54 [52,53]. The analyzed MLM was y = Xβ + Zu + e; where y is the vector of BLUEs for each trait, β is termed as a vector of the SNP marker fixed-effects parameters, u denotes a vector of random additive effects (inbreds), X and Z represent the design matrices for the fixed effects and the random effects, respectively; e refers to a vector of random residuals. The other details followed our previous study [29]. An association between a trait and an SNP was considered significant when the observed −log10 (p) values showed a clear derivation from the distribution of the expected −log10 (p) that holds under the null hypothesis (i.e., no significant association) in quantile–quantile (QQ) plots [54,55,56].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13030801/s1, Figure S1: Manhattan plot and Quantile–quantile plot from a mixed linear model for photosynthesis-related traits in a large maize association population via single environment analysis. SNPs with p values ≤the threshold of 1 × 10−4 were considered significantly associated with the trait. The different colors in the Manhattan plot represent the 10 different chromosomes of maize. Figure S2: Comparison of Leaf net photosynthesis (AN) among groups with the different number of favorable alleles carried in a large maize panel. The x-axis indicates groups 1, 2, 3, 4, and 5 with inbred lines carrying 1, 2, 3, 4, and 5 favorable alleles in a large maize panel, respectively. The different upper letters above the mean lines denote significant differences at p < 0.05. The number of inbred lines contained in each group is presented below the lines. A total of 18 inbred lines, including EP32, GE129, NC250, Pa392, Oh43E, J47, Mo46, MS24, MS78, MS1, CH9, P39M96, A681, H29w, AusTRCF306303, AusTRCF305819, A665, and ND301, carried five favorable alleles to increase Pn. Table S1: Candidate genes associated with photosynthesis-related traits in a large maize association panel across three environments.
Author Contributions
P.R. conceived and designed the study. Q.Y., P.R. and L.Á.-I. carried out the field trial and participated in the data collection; Q.Y. and A.L.-M. performed data analysis; Q.Y. wrote the first draft; L.Á.-I. contributed on the writing of the introduction; A.L.-M. contributed on the writing of material and methods and results; M.C.R. contributed on results and discussion; A.L.-M. was in charge of the preparation and submission of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministerio de Innovación y Universidades (MCIU), the Agencia Estatal de Investigación (AEI) and the European Fund for Regional Development (FEDER), UE (project code PID2019-108127RB-I00) and by the European Union under H2020 framework programme (Proj. 1586, Ref PCI2021-121912). There was also the support of the Guizhou Provincial Science and Technology Plan Project (Qian Kehe Support [2022]key026).
Data Availability Statement
Genotypic data is publicly available at Panzea (http://www.panzea.org(accessed on 11 September 2019)). The data sets used and/or analyzed during the current study will be available upon request to Qiang Yi (yiqiang990@hotmail.com).
Acknowledgments
Qiang Yi would like to acknowledge the support of the Guizhou Provincial Science and Technology Plan Project (Qian Kehe Support [2022]key026). We thank Ana Butrón for the English language review.
Conflicts of Interest
The authors declare no competing interest.
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Yin, X.; Stomph, T.J.; Struik, P.C. Can Exploiting Natural Genetic Variation in Leaf Photosynthesis Contribute to Increasing Rice Productivity? A Simulation Analysis. Plant Cell Environ. 2014, 37, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Basu, U.; Bajaj, D.; Sharma, A.; Malik, N.; Daware, A.; Narnoliya, L.; Thakro, V.; Upadhyaya, H.D.; Kumar, R.; Tripathi, S.; et al. Genetic Dissection of Photosynthetic Efficiency Traits for Enhancing Seed Yield in Chickpea. Plant Cell Environ. 2019, 42, 158–173. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ainsworth, E.A.; Ort, D.R. A Meta-Analysis of Responses of Canopy Photosynthetic Conversion Efficiency to Environmental Factors Reveals Major Causes of Yield Gap. J. Exp. Bot. 2013, 64, 3723–3733. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Rufty, T.W.; Lewis, R.S. Increasing Photosynthesis: Unlikely Solution For World Food Problem. Trends Plant Sci. 2019, 24, 1032–1039. [Google Scholar] [CrossRef]
- Hussain, S.; Ulhassan, Z.; Brestic, M.; Zivcak, M.; Zhou, W.; Allakhverdiev, S.I.; Yang, X.; Safdar, M.E.; Yang, W.; Liu, W. Photosynthesis Research under Climate Change. Photosynth. Res. 2021, 150, 5–19. [Google Scholar] [CrossRef]
- Stewart, D.W.; Costa, C.; Dwyer, L.M.; Smith, D.L.; Hamilton, R.I.; Ma, B.L. Canopy Structure, Light Interception, and Photosynthesis in Maize. Agron. J. 2003, 95, 1465–1474. [Google Scholar] [CrossRef]
- Lee, E.A.; Tollenaar, M. Physiological Basis of Successful Breeding Strategies for Maize Grain Yield. Crop Sci. 2007, 47, S-202. [Google Scholar] [CrossRef]
- Sekhon, R.S.; Saski, C.; Kumar, R.; Flinn, B.S.; Luo, F.; Beissinger, T.M.; Ackerman, A.J.; Breitzman, M.W.; Bridges, W.C.; De Leon, N.; et al. Integrated Genome-Scale Analysis Identifies Novel Genes and Networks Underlying Senescence in Maize. Plant Cell 2019, 31, 1968–1989. [Google Scholar] [CrossRef]
- Yan, Y.; Hou, P.; Duan, F.; Niu, L.; Dai, T.; Wang, K.; Zhao, M.; Li, S.; Zhou, W. Improving Photosynthesis to Increase Grain Yield Potential: An Analysis of Maize Hybrids Released in Different Years in China. Photosynth. Res. 2021, 150, 295–311. [Google Scholar] [CrossRef]
- Flood, P.J.; Harbinson, J.; Aarts, M.G.M. Natural Genetic Variation in Plant Photosynthesis. Trends Plant Sci. 2011, 16, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Faralli, M.; Matthews, J.; Lawson, T. Exploiting Natural Variation and Genetic Manipulation of Stomatal Conductance for Crop Improvement. Curr. Opin. Plant Biol. 2019, 49, 1–7. [Google Scholar] [CrossRef]
- Huo, Y.; Wang, M.; Wei, Y.; Xia, Z. Overexpression of the Maize PsbA Gene Enhances Drought Tolerance through Regulating Antioxidant System, Photosynthetic Capability, and Stress Defense Gene Expression in Tobacco. Front. Plant Sci. 2016, 6, 1223. [Google Scholar] [CrossRef] [PubMed]
- Salesse-Smith, C.E.; Sharwood, R.E.; Busch, F.A.; Kromdijk, J.; Bardal, V.; Stern, D.B. Overexpression of Rubisco Subunits with RAF1 Increases Rubisco Content in Maize. Nat. Plants 2018, 4, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Yoon, D.K.; Ishiyama, K.; Suganami, M.; Tazoe, Y.; Watanabe, M.; Imaruoka, S.; Ogura, M.; Ishida, H.; Suzuki, Y.; Obara, M.; et al. Transgenic Rice Overproducing Rubisco Exhibits Increased Yields with Improved Nitrogen-Use Efficiency in an Experimental Paddy Field. Nat. Food 2020, 1, 134–139. [Google Scholar] [CrossRef]
- Leakey, A.D.; Ferguson, J.N.; Pignon, C.P.; Wu, A.; Jin, Z.; Hammer, G.L.; Lobell, D.B. Water Use Efficiency as a Constraint and Target for Improving the Resilience and Productivity of C3 and C4 Crops. Annu. Rev. Plant Biol. 2019, 70, 781–808. [Google Scholar] [CrossRef]
- Prado, S.A.; Cabrera-Bosquet, L.; Grau, A.; Coupel-Ledru, A.; Millet, E.J.; Welcker, C.; Tardieu, F. Phenomics Allows Identification of Genomic Regions Affecting Maize Stomatal Conductance with Conditional Effects of Water Deficit and Evaporative Demand. Plant. Cell Environ. 2018, 41, 314–326. [Google Scholar] [CrossRef]
- Ghannoum, O. How Can We Breed for More Water Use-Efficient Sugarcane? J. Exp. Bot. 2016, 67, 557–559. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Liu, S.; Ferjani, A.; Li, J.; Yan, J. Genetic Variation in ZmVPP1 Contributes to Drought Tolerance in Maize Seedlings. Nat. Genet. 2016, 48, 1233–1241. [Google Scholar] [CrossRef]
- Djemel, A.; Álvarez-Iglesias, L.; Pedrol, N.; López-Malvar, A.; Ordás, A.; Revilla, P. Identification of Drought Tolerant Populations at Multi-Stage Growth Phases in Temperate Maize Germplasm. Euphytica 2018, 214, 138. [Google Scholar] [CrossRef]
- Djemel, A.; Álvarez-Iglesias, L.; Santiago, R.; Malvar, R.A.; Pedrol, N.; Revilla, P. Algerian Maize Populations from the Sahara Desert as Potential Sources of Drought Tolerance. Acta Physiol. Plant. 2019, 41, 12. [Google Scholar] [CrossRef]
- Lopez, M.A.; Xavier, A.; Rainey, K.M. Phenotypic Variation and Genetic Architecture for Photosynthesis and Water Use Efficiency in Soybean (Glycine max L. Merr). Front. Plant Sci. 2019, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, Y.; Zhang, S.; Che, Z.; Yuan, W.; Yu, D. GWAS Reveals Two Novel Loci for Photosynthesis-Related Traits in Soybean. Mol. Genet. Genom. 2020, 295, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Lü, H.; Yang, Y.; Li, H.; Liu, Q.; Zhang, J.; Yin, J.; Chu, S.; Zhang, X.; Yu, K.; Lv, L.; et al. Genome-Wide Association Studies of Photosynthetic Traits Related to Phosphorus Efficiency in Soybean. Front. Plant Sci. 2018, 9, 1226. [Google Scholar] [CrossRef]
- Pignon, C.P.; Fernandes, S.B.; Valluru, R.; Bandillo, N.; Lozano, R.; Buckler, E.; Gore, M.A.; Long, S.P.; Brown, P.J.; Leakey, A.D.B. Phenotyping Stomatal Closure by Thermal Imaging for GWAS and TWAS of Water Use Efficiency-Related Genes. Plant Physiol. 2021, 187, 2544–2562. [Google Scholar] [CrossRef]
- Pelleschi, S.; Leonardi, A.; Rocher, J.P.; Cornic, G.; De Vienne, D.; Thévenot, C.; Prioul, J.L. Analysis of the Relationships between Growth, Photosynthesis and Carbohydrate Metabolism Using Quantitative Trait Loci (QTLs) in Young Maize Plants Subjected to Water Deprivation. Mol. Breed. 2006, 17, 21–39. [Google Scholar] [CrossRef]
- Flood, P.J. Using Natural Variation to Understand the Evolutionary Pressures on Plant Photosynthesis. Curr. Opin. Plant Biol. 2019, 49, 68–73. [Google Scholar] [CrossRef]
- Morales, F.; Ancín, M.; Fakhet, D.; González-Torralba, J.; Gámez, A.L.; Seminario, A.; Soba, D.; Ben Mariem, S.; Garriga, M.; Aranjuelo, I. Photosynthetic Metabolism under Stressful Growth Conditions as a Bases for Crop Breeding and Yield Improvement. Plants 2020, 9, 88. [Google Scholar] [CrossRef]
- Yi, Q.; Álvarez-Iglesias, L.; Malvar, R.A.; Romay, M.C.; Revilla, P. A Worldwide Maize Panel Revealed New Genetic Variation for Cold Tolerance. Theor. Appl. Genet. 2021, 134, 1083–1094. [Google Scholar] [CrossRef]
- Romay, M.C.; Millard, M.J.; Glaubitz, J.C.; Peiffer, J.A.; Swarts, K.L.; Casstevens, T.M.; Elshire, R.J.; Acharya, C.B.; Mitchell, S.E.; Flint-garcia, S.A.; et al. Comprehensive Genotyping of the USA National Maize Inbred Seed Bank. Genome Biol. 2013, 14, R55. [Google Scholar] [CrossRef]
- Wallace, J.G.; Bradbury, P.J.; Zhang, N.; Gibon, Y.; Stitt, M.; Buckler, E.S. Association Mapping across Numerous Traits Reveals Patterns of Functional Variation in Maize. PLoS Genet. 2014, 10, e1004845. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, M.R.; Cannon, E.K.; Portwood, J.L.; Harper, L.C.; Gardiner, J.M.; Schaeffer, M.L.; Andorf, C.M. A Pan-Genomic Approach to Genome Databases Using Maize as a Model System. BMC Plant Biol. 2021, 21, 385. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Zhong, Y. Genetic Dissection of the Photosynthetic Parameters of Maize (Zea mays L.) in Drought-Stressed and Well-Watered Environments. Russ. J. Plant Physiol. 2021, 68, 1125–1134. [Google Scholar] [CrossRef]
- Duncan, W.G.; Hesketh, J.D. Net Photosynthetic Rates, Relative Leaf Growth Rates, and Leaf Numbers of 22 Races of Maize Grown at Eight Temperatures1. Crop Sci. 1968, 8, 670–674. [Google Scholar] [CrossRef]
- Long, S.P.; Marshall-Colon, A.; Zhu, X.G. Meeting the Global Food Demand of the Future by Engineering Crop Photosynthesis and Yield Potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic Strategies for Improving Crop Yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Zamioudis, C.; Hanson, J.; Pieterse, C.M.J. β-Glucosidase BGLU42 Is a MYB72-Dependent Key Regulator of Rhizobacteria-Induced Systemic Resistance and Modulates Iron Deficiency Responses in Arabidopsis Roots. New Phytol. 2014, 204, 368–379. [Google Scholar] [CrossRef]
- Rouyi, C.; Baiya, S.; Lee, S.K.; Mahong, B.; Jeon, J.S.; Ketudat- Cairns, J.R.; Ketudat-Cairns, M. Recombinant Expression and Characterization of the Cytoplasmic Rice β-Glucosidase Os1BGlu4. PLoS ONE 2014, 9, e96712. [Google Scholar] [CrossRef]
- Castorina, G.; Domergue, F.; Chiara, M.; Zilio, M.; Persico, M.; Ricciardi, V.; Horner, S.; Consonni, G. Drought-Responsive ZmFDL1 / MYB94 Regulates Cuticle Biosynthesis and Cuticle-Dependent Leaf Permeability. Plant Physiol. 2020, 184, 266–282. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-Sink Transport of Sugar and Regulation by Environmental Factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef]
- Rajhi, I.; Yamauchi, T.; Takahashi, H.; Nishiuchi, S.; Shiono, K.; Watanabe, R.; Mliki, A.; Nagamura, Y.; Tsutsumi, N.; Nishizawa, N.K.; et al. Identification of Genes Expressed in Maize Root Cortical Cells during Lysigenous Aerenchyma Formation Using Laser Microdissection and Microarray Analyses. New Phytol. 2011, 190, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, S.; Bertin, P.; Presterl, T.; Jamin, P.; Coubriche, D.; Gouesnard, B.; Laborde, J.; Charcosset, A. Association Mapping for Phenology and Plant Architecture in Maize Shows Higher Power for Developmental Traits Compared with Growth Influenced Traits. Hered 2016, 118, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; He, X.; Baer, M.; Beirinckx, S.; Tian, T.; Moya, Y.A.T.; Zhang, X.; Deichmann, M.; Frey, F.P.; Bresgen, V.; et al. Plant Flavones Enrich Rhizosphere Oxalobacteraceae to Improve Maize Performance under Nitrogen Deprivation. Nat. Plants 2021, 7, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, H.; Hu, X.; Liu, Z.; Wu, Y.; Huang, C. Genome-Wide Association Study Reveals the Genetic Basis of Stalk Cell Wall Components in Maize. PLoS ONE 2016, 11, e0158906. [Google Scholar] [CrossRef]
- Monir, M.M.; Zhu, J. Dominance and Epistasis Interactions Revealed as Important Variants for Leaf Traits of Maize NAM Population. Front. Plant Sci. 2018, 9, 627. [Google Scholar] [CrossRef]
- Pan, L.; George-Jaeggli, B.; Borrell, A.; Jordan, D.; Koller, F.; Al-Salman, Y.; Cano, F.J. Coordination of Stomata and Vein Patterns with Leaf Width Underpins Water-use Efficiency in a C4 Crop. Plant Cell Environ. 2022, 45, 1612–1630. [Google Scholar] [CrossRef]
- Revilla, P.; Rodríguez, V.M.; Ordás, A.; Rincent, R.; Charcosset, A.; Giauffret, C.; Melchinger, A.E.; Schön, C.C.; Bauer, E.; Altmann, T.; et al. Cold Tolerance in Two Large Maize Inbred Panels Adapted to European Climates. Crop Sci. 2014, 54, 1981–1991. [Google Scholar] [CrossRef]
- Gobierno de España Agencia Estatal de Meteorología—AEMET. Available online: https://www.aemet.es/ (accessed on 18 November 2022).
- Buckler, E.S.; Holland, J.B.; Bradbury, P.J.; Acharya, C.B.; Brown, P.J.; Browne, C.; Ersoz, E.; Flint-Garcia, S.; Garcia, A.; Glaubitz, J.C.; et al. The Genetic Architecture of Maize Flowering Time. Science 2009, 325, 714–718. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Bort, J.; Nogués, S.; Araus, J.L. The Combined Effect of Constant Water Deficit and Nitrogen Supply on WUE, NUE and Δ13C in Durum Wheat Potted Plants. Ann. Appl. Biol. 2007, 151, 277–289. [Google Scholar] [CrossRef]
- SAS/STAT, Statistical Analysis System Proprietary Software Release 9.4; SAS Institute Inc.: Cary, NC, USA, 2007.
- Holland, J.B.; Nyquist, W.E.; Cervantes-Martínez, C.T. Estimated and Interpreting Heritability for Plant Breeding: An Update. Plant Breed. Rev. 2003, 22, 9–122. [Google Scholar]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Bi, I.V.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A Unified Mixed-Model Method for Association Mapping That Accounts for Multiple Levels of Relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Becker, L.C.; Becker, D.M.; Starmer, J.D.; Province, M.A. Avoiding the High Bonferroni Penalty in Genome-Wide Association Studies. Genet. Epidemiol. 2010, 34, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Gowda, M.; Das, B.; Makumbi, D.; Babu, R.; Semagn, K.; Mahuku, G.; Olsen, M.S.; Bright, J.M.; Beyene, Y.; Prasanna, B.M. Genome-Wide Association and Genomic Prediction of Resistance to Maize Lethal Necrosis Disease in Tropical Maize Germplasm. Theor. Appl. Genet. 2015, 128, 1957–1968. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).