Abstract
Modern vegetable production systems are often characterized by monoculture fields and the intensive use of tillage and/or synthetic agrochemicals for managing weeds. A growing public interest in more sustainable and eco-friendly production practices has resulted in increased demand for crops to be produced with lower inputs. Field studies were conducted over three field seasons to investigate the use of conservation tillage in concert with an interplanted living mulch and/or cover crop residue for managing weeds in sweet corn as compared with the standard practice of using conventional tillage and pre-emergence residual herbicides. Whole plot treatments included: (1) conventional till, (2) no-till with cover crop residue, (3) living mulch + cover crop residue, and (4) living mulch + winter killed residue. The split-plot factor consisted of herbicide treatments: (1) at-planting application of residual herbicides or (2) no herbicide. The cover crop systems suppressed weeds as well as the standard practice throughout the cropping cycle in all three years. In addition, there was no significant improvement in weed suppression with the application of herbicides within the cover crop treatments. Crop development and yield were similar among treatments in year 2. However, reduced yields were encountered in all cover crop treatments during year 3 relative to the conventional tillage treatment.
1. Introduction
Modern crop production systems are typically characterized by large monoculture fields and the intensive use of synthetic agrochemicals for managing weeds [1]. This lack of diversity has resulted in undesirable side effects including the development of herbicide-resistant weeds, health-related issues resulting from occupational or consumer exposure to chemical residues, and increased farm expense. Integrated weed management programs designed to reduce chemical inputs often rely on mechanical weed control methods such as deep tillage and cultivation, as well as hand weeding. These tactics demand large amounts of fossil fuel or human labor and can therefore significantly increase production costs in conventional and organic farming systems [2,3]. Furthermore, labor costs and shortages contribute to a need to reduce the amount of hand weeding, especially in organic operations [4,5]. An interest in more sustainable and eco-friendly production practices has resulted in a growing demand for crops to be produced with lower inputs [6].
In addition to the cost associated with intensive herbicide use, this tactic has several additional drawbacks. For example, there are a limited number of herbicides registered for use in vegetable crops, and many provide only partial weed control [7]. Furthermore, commonly used herbicides in vegetables may reduce yields by causing injury and stunting [8]. Subsequently, many are not labeled for postemergence broadleaf weed control unless applied with shielded sprayers to prevent injury. Moreover, herbicide-resistant weed populations in many areas have resulted in the reduced efficacy of many commonly used herbicides [9] and, due to the relatively small market size, chemical companies lack financial incentives to develop replacement products for many vegetable crops [10]. These limitations suggest that additional weed management options are needed.
Increasing the number of available weed management options via the addition of cover crop residues and living mulches can help create cropping environments that are more diverse and less reliant on chemical inputs. Using conservation tillage practices together with cover cropping can reduce weed pressure and management cost through a reduction in hand weeding, herbicide sprays, and fuel usage [11]. Cover crops suppress weeds, in part by filling ecological niches otherwise occupied by weeds. Moreover, different cover crop species, species combinations, and cover cropping tactics may result in varying levels of weed suppression. For example, cover crop residue that remains on the soil surface may suppress weed establishment by reducing the light levels needed for seed germination and by acting as a physical barrier to seedling germination and growth [12]. Furthermore, its efficacy may differ whether it is a grass or broadleaf cover crop [13]. Additionally, allelopathic compounds released by certain decomposing cover crops can hinder weed seed germination or act as plant growth inhibitors [14,15,16]. Past research has highlighted the ability of surface residue created from fall-planted cover crops to reduce weed populations during the subsequent growing season [17,18]. Furthermore, many studies have demonstrated greater weed suppression with increasing amounts of cover crop biomass [19,20]. Cover crop surface residues are typically created by terminating living cover crops using a roller-crimper, whereby rolling drums are fitted with dull blades designed to crimp stems rather than chop them.
Most research aimed at determining the impacts of conservation tillage and cover cropping on weed establishment was directed at agronomic crops. This has resulted in enhanced adoption of these practices in crops such as field corn and soybean [21]. Previous attempts to incorporate cover crop residue and conservation tillage into vegetable production have often resulted in lower yields and/or delayed harvests caused by seedling interference and cooler soil temperatures [22]. Furthermore, cover crop residues decompose over time and consequently may provide only early-season weed suppression. This can lead to weed establishment later in the season, particularly in the inter-row area where the soil surface is not covered by the main crop’s canopy [23]. Alternatively, cover crops used as living mulches and interplanted within the cash crop maintain the ability to suppress weeds throughout the cash crop cycle [24]. Nevertheless, living mulches are limited to suppressing weeds in the inter-row area, and if not properly managed, may compete with the main crop causing yield reductions [20]. Furthermore, the incorporation of living mulches into crop production may require banded herbicide sprays or strip tillage to clear out zones for establishing the crop rows. Strip tillage is a minimum tillage tactic that confines soil disturbance to a narrow zone within the crop row [25]. However, soil disruption in the intra-row area may result in weeds emerging within the crop row. As such, weed management remains an important challenge in strip-tilled operations [7,26]. A potential method for eliminating the need for banded herbicides and strip tillage from living mulch crop production is to plant the living mulch in alternating strips with annual cover crops, such that the living mulch is restricted to the inter-row areas.
The purpose of this research was to investigate the use of conservation tillage in concert with an interplanted living mulch and/or cover crop residue for managing weeds in sweet corn and compare this with the traditional practice of using conventional tillage and a pre-emergence herbicide application. Furthermore, we investigated the use of cover crop diculture systems, whereby alternating strips of living mulch and either a spring-terminated cover crop residue or frost-killed dying mulch were planted as a means of restricting the living mulch to the inter-row areas. We hypothesized that combining a living mulch with a terminated cover crop residue or frost-killed dying mulch would result in similar weed suppression compared to joint herbicides and tillage programs, and that both living mulch treatments would result in greater weed suppression than the no-till treatment by the end of the sweet corn growing season. Furthermore, we hypothesized that sweet corn productivity and yield would be similar among treatments.
2. Materials and Methods
2.1. Experimental Design and Field Operations
Field operations were conducted in 2019, 2020, and 2021 at the Central Maryland Research and Education Center in Upper Marlboro, MD (38.859079°, −76.778731°, year 1), and roughly 16 km away in Beltsville, MD (39.011440°, −76.833356°, years 2 and 3). Soils at the study sites are Annapolis series (fine-loamy, glauconitic, mesic Typic Hapludults) (Upper Marlboro) and a Russett-Christiana complex where the Russett surface soil is classified as loam or sandy loam and the Christiana surface soil is classified as silt loam (Beltsville). Treatments were arranged in a Latin square: split-plot design with four replicates. Whole plot treatments included: (1) conventional till (CT), (2) no-till with cover crop residue (NT), (3) living mulch + cover crop residue (LMRye), and (4) living mulch + winter killed residue (LMFR). The split-plot factor consisted of herbicide treatments: (1) an at-planting application of residual herbicides (herbicide) or (2) no herbicide application (no herbicide). Main plots measured 82.8 m2 (9.1 m × 9.1 m) and each subplot measured 41.9 m2 (4.6 m × 9.1 m).
During early fall, a mixture of crimson clover (Trifolium incarnatum; 3.36 kg ha−1), forage radish (Raphanus sativus; 3.9 kg ha−1), and cereal rye (Secale cereale L. ‘Aroostook’; 62.8 kg ha−1) was planted in CT and NT plots. In living mulch treatments, rows alternated between two rows of red clover (Trifolium pratense) and three rows of cereal rye (75.1 kg ha−1) in LMRye or forage radish (11.2 kg ha−1) in LMFR. Red clover was seeded at a rate of 9 kg ha−1 in LMRye plots and 16.8 kg ha−1 in LMFR plots. All cover crops were drilled at an interrow spacing of 15.2 cm. In the spring, when the rye reached anthesis, cover crops in CT plots were mowed, plowed, and incorporated into the soil. Crimson clover senesced naturally, and the forage radish was winter killed in 2019 and 2020. A roller crimper was used to terminate the rye in the NT and LMRye treatments, and temporarily slow red clover growth in LMRye and LMFR plots. In late May, sweet corn (Zea mays convar. saccharata var. rugosa) [variety: Providence (Syngenta, Wilmington, DE)] was seeded into each plot at an inter-row spacing of 76.2 cm, resulting in 12 crop rows per plot. In LMRye and LMFR plots, sweet corn seeds were planted within the center of the strips of forage radish or rye residue. A pre-emergence herbicide combination of 1.68 kg ai ha−1 atrazine and 1.42 kg ai ha−1 S-metolachlor was immediately applied to herbicide subplots following sweet corn planting. Herbicides were broadcasted in NT and CT herbicide subplots and banded within the strips (intra-row area) of the forage radish (LMFR) or rye (LMRye) herbicide subplots. Plots were overhead irrigated as needed to mitigate periods of low rainfall and a split-application of nitrogen fertilizer was applied according to recommended production practices. The timing of field tasks is provided in Table 1.

Table 1.
Timing of field operations in 2019 (Upper Marlboro, MD), 2020, and 2021 (Beltsville, MD).
2.2. Cover Crop and Weed Biomass
The cover crop and winter annual weed biomass were collected from each plot just prior to cover crop termination by clipping shoot tissue at the ground level from two 0.3 m × 0.3 m quadrats. Each quadrat was placed randomly in CT and NT plots and within the intra-row areas of the LMRye and LMFR treatments. Plant material within each quadrat was separated by a cover crop or weed, dried, and weighed. Biomass measurements of the red clover were not taken because the red clover remained a living mulch and continued to develop throughout the cropping cycle. However, it was noted that the red clover completely covered the soil surface in the inter-row areas.
To assess treatment impacts on weed biomass accumulation in the absence of any weed management intervention, two 0.78 m2 unmanaged areas were established in each subplot immediately after sweet corn planting. Plot areas outside the unmanaged zones were manually weeded to assess the direct impact of treatment on the yield. Biomass samples were collected from one unmanaged area six weeks after sweet corn planting (WAP) and the second immediately following the final sweet corn harvest. Measurements were taken by clipping and collecting all the weeds at the soil level. Weeds were separated by species, dried at 60 °C (>1 week), and weighed. Dry weight measurements were combined for each species to determine total biomass per treatment, and species measurements were used to calculate the relative abundances of the most common species.
2.3. Crop Development and Yield
Stand counts were initiated in all treatments at less than 10 d after planting. Counts were repeated every 3–4 d until all the viable seedlings had emerged. To assess the treatment impact on sweet corn growth, the developmental stage, extended leaf height, and chlorophyll content of five randomly selected plants per subplot were recorded. Measurements were taken weekly beginning at the V1 stage (the first leaf fully emerged) and continuing through VT (tassels fully visible). The corn development stage was determined according to [27]. The extended leaf height was measured from the soil surface to the tallest extended leaf. Chlorophyll content was measured as an indicator of nitrogen level using a Soil Plant Analysis Development (SPAD)-502 chlorophyll meter (Spectrum Technologies Inc., Plainfield, IL, USA). Three measurements were taken per leaf (base, center, and leaf tip) from the last fully emerged leaf and averaged to provide a single value per plant. Chlorophyll meters were reported to detect nitrogen deficiencies by the V8 stage [28]. As such, the analysis of SPAD readings and crop growth parameters focused on those measurements taken between the V8 and VT stages. To estimate yield, all sweet corn ears located in the center 6.15 m of the four center rows within each subplot were harvested, and all the marketable ears were counted and weighed. The yield was collected over multiple dates until all the marketable primary ears were harvested.
2.4. Statistical Analysis
Cover crop biomass was averaged within each treatment over all the experiment years and the standard error of the total biomass per treatment calculated. All plant growth and development metrics were analyzed using linear mixed models (LMM) to test for differences in emergence, crop chlorophyll content, extended leaf height, development stage with treatment (CT, LMFR, LMRye, and NT), subplot treatment (herbicide vs. no herbicide), and their interaction as fixed effects. When the LMM indicated a significant difference between the treatment means, post-hoc pairwise means comparisons were performed using Tukey-adjusted p-values [29]. Similar analyses were performed on sweet corn yield and weed biomass data. Data were log transformed when necessary to stabilize the variances. For weed abundance, species that made up at least 5% of the total number of weeds (2019) or total biomass (2020, 2021) were listed. Data for all the experiment years were analyzed together unless a significant interaction between the year and cover crop treatment was detected. Seedling emergence and delays in harvest maturity were determined by calculating the proportion of the total emerged/harvested on each sample date. All statistical analyses were performed using R (v. 4.1.2; R Core Team 2021). Linear mixed effect models were built using the package ‘lme4′ [30]. Post-hoc means comparisons were performed using the package ‘emmeans’ [29]. All figures were made using the ‘ggplot2′ package [31].
3. Results
3.1. Cover Crop and Weed Biomass
Similar amounts of cover crop biomass were collected from each treatment each year. As such, the biomass of each cover crop species per treatment was averaged over all the study years. A high production of the cover crop biomass and comparatively low winter annual weed biomass occurred across all the treatments (Figure 1). CT biomass averaged 3891 kg ha−1 and 2887 kg ha−1 and NT biomass averaged 3926 kg ha−1 and 2727 kg ha−1 for rye and crimson clover, respectively. Rye biomass in LMRye averaged 7976 kg ha−1 but was only planted in 60% of the plot area.
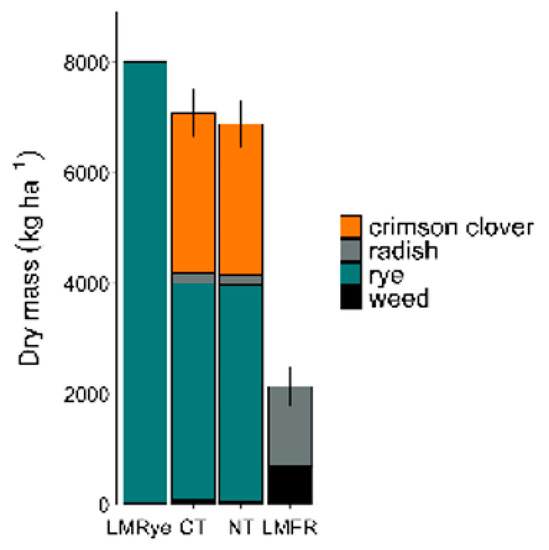
Figure 1.
Mean biomass of cover crop and winter annual weeds averaged over all experiment years in plots designated as living mulch + rye (LMRye), conventional tillage (CT), no-till (NT), and living much + forage radish (LMFR).
The dominant weed species encountered six weeks after sweet corn planting across all the experiment years and locations were cutleaf evening-primrose (Oenothera laciniata Hill), goosegrass (Eleusine indica L.), and common lambsquarters (Chenopodium album L.). However, the substantial variation in species abundance occurred between treatments. All weed species comprising >5% of the total biomass within each experiment year at 6 WAP are shown in Table 2.
Table 2.
Percent of abundance of all weed species making up >5% total abundance.
No significant year effect was detected for the weed biomass measurements taken 6 WAP. As such, data from all the study years were combined and analyzed together. Significant treatment (χ2 = 11.32, df = 3, p < 0.01), herbicide (χ2 = 33.93, df = 1, p < 0.001), and treatment by herbicide interaction (χ2 = 11.86, df = 3, p < 0.01) effects were found. Significantly greater weed biomass was collected in the no herbicide CT subplots compared to all the other whole-subplot treatment combinations except for NT no herbicide (Figure 2A). No differences were detected in the amount of weed biomass collected from herbicide and no herbicide subplots within each living mulch treatment (LMFR and LMRye) and amounts of weed biomass collected in all the cover crop subplots were similar to the herbicide-treated CT subplot. A significantly greater biomass was collected in the NT no herbicide subplot compared to the LMRye herbicide subplot. A significant effect of the study year was detected for biomass samples taken at crop harvest. However, p-value corrections resulting from Tukey pairwise comparisons did not detect any significant differences. Thus, study years were again combined for the analysis. Treatment effects on weed biomass at harvest were similar to those detected at 6 WAP. Significant treatment (χ2 = 16.60, df = 3, p < 0.001), herbicide (χ2 = 15.95, df = 1, p < 0.001), and interaction (χ2 = 13.11, df = 3, p < 0.01) effects were found and the no herbicide CT treatment contained more weeds than all other subplot treatment combinations except for NT no herbicide (Figure 2B). No differences were detected in the amount of biomass collected from the herbicide and no herbicide subplots within each cover crop treatment and the amounts of weed biomass collected in all the cover crop subplots were similar to the herbicide-treated CT subplot.
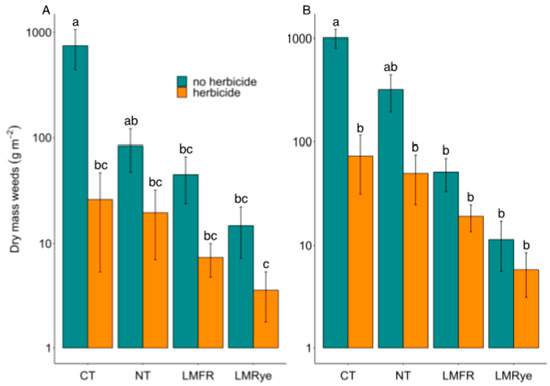
Figure 2.
Weed biomass collected from unweeded areas (A) six weeks after sweet corn planting and (B) following crop harvest over three experimental years in conventional till (CT), no-till (NT), living mulch + forage radish residue (LMFR), and living mulch + rye residue (LMRye). Y axis presented on a log scale. Different letters indicate significant differences.
3.2. Crop Development and Yield
Equipment used in 2019 caused poor seed placement and required replanting after poor initial establishment. Therefore, crop development and yield could not be measured in 2019. In 2020, corn seedling emergence was roughly 1–2 days slower in NT and LMRye treatments compared to CT and LMFR (Figure 3A), while seedling emergence was approximately one day slower in LMRye compared to all the other treatments in 2021 (Figure 3B). In 2020, no significant treatment or herbicide differences were detected in the final stand count, leaf chlorophyll concentration, extended leaf height, crop development stage, or yield (Table 3).
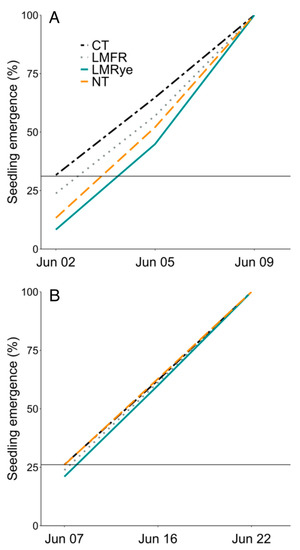
Figure 3.
Percent of total sweet corn seedling emergence by sample date in (A) 2020 and (B) 2021 in conventional till (CT), no-till (NT), living mulch + forage radish residue (LMFR), and living mulch + rye residue (LMRye). Horizontal line marks the initial stand count in CT.
Table 3.
Mean ± SEM plant development and yield metrics for 2020 and 2021.
In 2021, no significant treatment or herbicide differences were detected in the final stand count. However, the average chlorophyll content differed by treatment (χ2 = 15.06, df = 3, p < 0.01) and herbicide (χ2 = 12.00, df = 1, p < 0.001). Lower chlorophyll levels were detected in no herbicide LMRye and LMFR compared to herbicide treated CT (Table 3). The extended leaf height differed by treatment (χ2 = 14.73, df = 3, p < 0.01) and herbicide (χ2 = 26.68, df = 1, p < 0.001) and an interaction effect was detected (χ2 = 22.65, df = 3, p < 0.001). Corn height was shorter in no herbicide subplots of NT, LMFR, and LMRye than their herbicide subplot equivalents and both CT subplot treatments. Similarly, in 2021, sweet corn stage differed according to treatment (χ2 = 16.25, df = 3, p < 0.01), herbicide (χ2 = 27.03, df = 1, p < 0.001), and their interaction (χ2 = 38.72, df = 3, p < 0.001). A developmental delay was detected in all the no herbicide cover crop treatments compared to their herbicide equivalents and both CT subplot treatments. In 2021, the total yield differed by treatment (χ2 = 28.08, df = 3, p < 0.001) and herbicide (χ2 = 51.48, df = 1, p < 0.001) and there was an interaction effect (χ2 = 21.50, df = 3, p < 0.001). The no herbicide cover crop subplots had lower yields than the no herbicide CT treatment (Table 3). Furthermore, all the no herbicide cover crop subplots experienced lower yields than their herbicide subplot equivalents. Although no differences in the total crop yield were detected in 2020, a harvest maturity delay of approximately one day in LMRye and two days in LMFR occurred compared to CT (Figure 4A). In 2021, a harvest maturity delay of approximately one day was detected in all the cover crop treatments compared to CT (Figure 4B).
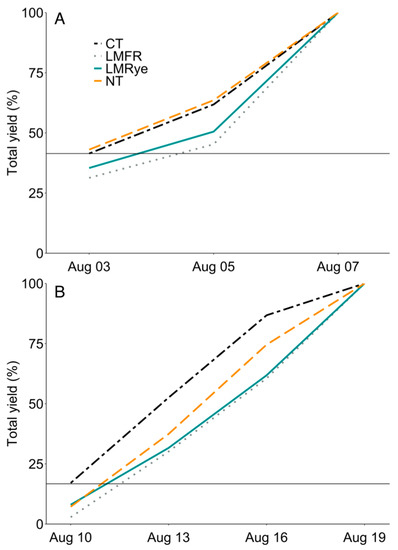
Figure 4.
Percent of total sweet corn yield harvested on each sample date in 2020 (A) and 2021 (B) in conventional till (CT), no-till (NT), living mulch + forage radish residue (LMFR), and living mulch + rye residue (LMRye). Horizontal line marks the initial yield in CT.
4. Discussion
The high production of cover crop biomass and comparatively low winter annual weed biomass occurred across CT, NT, and LMRye treatments. Previous studies have associated high cover crop biomass with reduced weed biomass [12,32] and likely explains the low biomass of winter annual weeds observed in the CT, NT, and LMRye treatments across all the study years. Greater winter annual weed biomass was found in the intra-row areas of LMFR treatments, likely resulting from increased sunlight and warmer soil temperatures that occurred as forage radish residue decomposed.
Managing weeds in strip tillage vegetable systems can be challenging [7,26]. This is in part due to differences in the weed communities typically found between the inter- and intra-row areas. Soil disturbance and reduced residue cover stimulate the germination of annual weeds in tilled zones [33,34,35], while untilled areas are more often plagued by a buildup of perennial weeds [7]. The cover cropping systems investigated during this study which involved combining a living mulch with a dying or killed organic mulch effectively suppressed weeds in the inter- and intra-row areas. As such, we determined that soil disturbances and subsequent weed flushes associated with strip tillage operations aimed to remove the living mulch from the intra-rows could be eliminated if the living mulch is restricted to the inter-rows and the intra-rows consist of organic mulch or decomposed cover crop residue. Our findings contrast previous research investigating the impact of interplanting sweet corn with strips of white clover (Trifolium repens L.) living mulch on weed establishment. Mohler (1991) found greater weed biomass in white clover treatments compared with sweet corn grown in rye residue or conventional tillage [36]. However, this finding was attributed to patchy white clover stands which allowed weed establishment. During the current study, it was also found that combining the living mulch treatments with an at-planting, pre-emergent herbicide application did not provide any additional weed suppressive benefits, except in NT treatments where the breakdown of residue allowed for late-season weed establishment in the absence of residual herbicides. Furthermore, combining rye residue with a living mulch resulted in significantly less weed biomass at crop harvest than the no-herbicide NT treatment and similar biomass to the NT herbicide subplot, suggesting that combining rye residue with a living mulch can provide similar late-season weed suppression to residual herbicides.
Variable treatment effects on sweet corn development and final yield were detected between experiment years. Previous research has found that lower soil temperatures under surface residues in minimum tillage systems can reduce or delay warm-season vegetable emergence and growth [22,37], and cooler soil temperatures can cause a delay in sweet corn seedling emergence [38]. However, treatment-specific emergence delays detected in 2020 and 2021 did not impact plant development metrics, chlorophyll content, or yield. Furthermore, no differences were detected in the final sweet corn stand counts during both study years. As such, growing sweet corn interplanted with a living mulch and/or within cover crop residue did not negatively affect crop establishment. The breakdown of winter-killed forage radish residues in LMFR treatments resulted in early-season bare-ground strips which allowed for warming of the soil prior to planting. As a result, unlike the NT and LMRye treatments, seedling emergence was not delayed in the LMFR treatment during 2020. Thus, the combination of living mulch with winter-killed cover crop residues presents a compatible option for warm-season vegetables susceptible to temperature-induced development delays.
In 2021, sweet corn in no herbicide living mulch subplots contained reduced chlorophyll content compared to the herbicide-treated CT treatment. Measurements of leaf chlorophyll content using SPAD meters are a proven method for detecting nitrogen deficiencies in corn [39]. Previous studies have shown that water shortages can reduce nitrogen uptake, thus resulting in reduced chlorophyll content, and that competition for water is an important mechanism by which living mulches may reduce crop yield [24,40]. Limited rainfall resulted in wilted, water-stressed plants in living mulch treatments during the early summer of 2021. As such, it is possible that reductions in sweet corn yield observed in living mulch subplots in 2021 resulted from either competition for water or nitrogen. In a prior study, competition for water with an interplanted white clover living mulch was also suggested as a factor contributing to reduced sweet corn [36]. Additionally, severe early-season stink bug damage in 2021 occurred primarily in NT, LMRye, and LMFR treatments. Previous research by [41] found that stink bug damage in seedling corn can cause reduced plant height and yield [41]. As such, the incurred stink bug damage likely contributed to a greater proportion of stunted sweet corn plants and lower yields in the cover crop compared to CT treatment in 2021.
Overall, interplanting sweet corn with a red clover living mulch and planting the corn rows into rolled rye or winter-killed forage radish residue resulted in the suppression of annual and perennial broadleaf and grass weeds in the inter- and intra-row areas throughout the cropping cycle. Furthermore, the weed suppressive effect was similar to the standard practice of tilling in the cover crop and applying a pre-emergent herbicide mixture at planting. This system can optimize the weed suppressive benefits of an interplanted living mulch while minimizing competition. As such, the cover crop diculture systems researched here may be a viable option for diversifying integrated weed management programs in sweet corn and other vegetable plantings.
Author Contributions
Conceptualization, V.L.Y. and C.R.R.H.; Data curation, V.L.Y.; Formal analysis, V.L.Y. and A.W.L.; Funding acquisition, V.L.Y., A.W.L. and C.R.R.H.; Investigation, V.L.Y.; Methodology, V.L.Y., A.W.L. and C.R.R.H.; Project administration, V.L.Y.; Visualization, V.L.Y.; Writing—original draft, V.L.Y.; Writing—review and editing, A.W.L. and C.R.R.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Northeast SARE Student Research Grant (GNE19-224) and by Research for Novel Approaches (LNE20-406R).
Data Availability Statement
Data available on request.
Acknowledgments
We are grateful to T. Patton, G. Dively, and M. Dimock for their assistance in the field. We would also like to thank the field crews at the Central Maryland Research and Education Centers in Beltsville and Upper Marlboro, Maryland, for their time, patience, and expert technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Rüegg, W.T.; Quadranti, M.; Zoschke, A. Herbicide research and development: Challenges and opportunities. Weed Res. 2007, 47, 271–275. [Google Scholar] [CrossRef]
- Sosnoskie, L.M.; Culpepper, A.S. Glyphosate-Resistant Palmer Amaranth (Amaranthus palmeri) Increases Herbicide Use, Tillage, and Hand-Weeding in Georgia Cotton. Weed Sci. 2014, 62, 393–402. [Google Scholar] [CrossRef]
- Archer, D.W.; Jaradat, A.A.; Johnson, J.M.F.; Weyers, S.L.; Gesch, R.W.; Forcella, F.; Kludze, H.K. Crop productivity and economics during the transition to alternative cropping systems. Agron. J. 2007, 99, 1538–1547. [Google Scholar] [CrossRef]
- Taylor, J.E.; Charlton, D.; Yuńez-Naude, A. The End of Farm Labor Abundance. Appl. Econ. Perspect. Policy 2012, 34, 587–598. [Google Scholar] [CrossRef]
- Zahniser, S.; Taylor, J.E.; Hertz, T.; Charlton, D. Farm Labor Markets in the United States and Mexico Pose Challenges for U. S. Agriculture. USDA Econ. Res. Serv. 2018, 201, 1–40. [Google Scholar]
- Rana, J.; Paul, J. Consumer behavior and purchase intention for organic food: A review and research agenda. J. Retail. Consum. Serv. 2017, 38, 157–165. [Google Scholar] [CrossRef]
- Brainard, D.C.; Peachey, R.E.; Haramoto, E.R.; Luna, J.M.; Rangarajan, A. Weed Ecology and Nonchemical Management under Strip-Tillage: Implications for Northern U.S. Vegetable Cropping Systems. Weed Technol. 2013, 27, 218–230. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, C.; Doohan, D. Safety of Bicyclopyrone on Several Vegetable Crops and Efficacy of Weed Control. Weed Technol. 2018, 32, 498–505. [Google Scholar] [CrossRef]
- Kniss, A.R. Genetically Engineered Herbicide-Resistant Crops and Herbicide-Resistant Weed Evolution in the United States. Weed Sci. 2018, 66, 260–273. [Google Scholar] [CrossRef]
- Gast, R.E. Industry Views of Minor Crop Weed Control. Weed Technol. 2008, 22, 385–388. [Google Scholar] [CrossRef]
- Mitchell, J.P.; Shrestha, A.; Horwath, W.R.; Southard, R.J.; Madden, N.; Veenstra, J.; Munk, D.S. Tillage and cover cropping affect crop yields and soil carbon in the San Joaquin valley, California. Agron. J. 2015, 107, 588–596. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Curran, W.S.; Mortenseny, D.M.; Ryany, M.R.; Shumway, D.L. Timing of Cover-Crop Management Effects on Weed Suppression in No-Till Planted Soybean using a Roller-Crimper. Weed Sci. 2011, 59, 380–389. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Mohler, C.L. Light Transmittance, Soil Temperature, and Soil Moisture under Residue of Hairy Vetch and Rye. Agron. J. 1993, 85, 673–680. [Google Scholar] [CrossRef]
- Ohno, T.; Doolan, K.; Zibilske, L.M.; Liebman, M.; Gallandt, E.R.; Berube, C. Phytotoxic effects of red clover amended soils on wild mustard seedling growth. Agric. Ecosyst. Environ. 2000, 78, 187–192. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Evidence for Allelopathy by Residues and Aqueous Extracts of Rye (Secale cereale). Weed Sci. 1986, 34, 384–390. [Google Scholar] [CrossRef]
- Creamer, N.G.; Bennett, M.A.; Stinner, B.R.; Cardina, J.; Regnier, E.E. Mechanisms of weed suppression in cover crop-based production systems. HortScience 1996, 31, 410–413. [Google Scholar] [CrossRef]
- Kruidhof, H.M.; Bastiaans, L.; Kropff, M.J. Ecological weed management by cover cropping: Effects on weed growth in autumn and weed establishment in spring. Weed Res. 2008, 48, 492–502. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Mohler, C.L. The quantitative relationship between weed emergence and the physical properties of mulches. Weed Sci. 2000, 48, 385–392. [Google Scholar] [CrossRef]
- Florence, A.M.; Higley, L.G.; Drijber, R.A.; Francis, C.A.; Lindquist, J.L. Cover crop mixture diversity, biomass productivity, weed suppression, and stability. PLoS ONE 2019, 14, e0206195. [Google Scholar] [CrossRef]
- Mennan, H.; Jabran, K.; Zandstra, B.H.; Pala, F. Non-chemical weed management in vegetables by using cover crops: A review. Agronomy 2020, 10, 257. [Google Scholar] [CrossRef]
- Wade, T.; Claassen, R.; Wallander, S. Conservation-Practice Adoption Rates Vary Widely by Crop and Region. United States Dep. Agric. Econ. Res. Serv. 2015, EIB-147, 40. [Google Scholar]
- Hoyt, G.D. Tillage and Cover Residue Affects on Vegetable Yields. HortTechnology 1999, 9, 351–358. [Google Scholar] [CrossRef]
- Teasdale, J.R.; Coffman, C.; Mangum, R. Potenital long-term benefits of no-tillage and organic cropping systems for grain production and soil improvement. Agron. J. 2007, 99, 1297–1305. [Google Scholar]
- Hartwig, N.L.; Ammon, H.U. 50th Anniversary—Invited Article Cover crops and living mulches. Weed Sci. 2002, 1745, 688–699. [Google Scholar] [CrossRef]
- Luna, J.M.; Staben, M.L. Strip tillage for sweet corn production: Yield and economic return. HortScience 2002, 37, 1040–1044. [Google Scholar] [CrossRef]
- Lowry, C.J.; Brainard, D.C. Strip intercropping of rye-vetch mixtures: Effects on weed growth and competition in strip-tilled sweet corn. Weed Sci. 2019, 67, 114–125. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Hanway, J.J.; Benson, G.O.; Herman, J.C. How a Corn Plant Develops, Special Report No. 48; Iowa State University: Ames, IA, USA, 1993. [Google Scholar]
- Varvel, G.E.; Schepers, J.S.; Francis, D.D. Ability for In-Season Correction of Nitrogen Deficiency in Corn Using Chlorophyll Meters. Soil Sci. Soc. Am. J. 1997, 61, 1233–1239. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squared Means. R Package Version 1.8.4-1. 2023. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 26 January 2023).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Buchanan, A.L.; Hooks, C.R.R. Influence of winter cover crop mulch on arthropods in a reduced tillage Cucurbit system. Environ. Entomol. 2018, 47, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Teasdale, J.R.; Beste, C.E.; Potts, W.E. Response of Weeds to Tillage and Cover Crop Residue. Weed Sci. 1991, 39, 195–199. [Google Scholar] [CrossRef]
- Wesson, G.; Wareing, P.F. The role of light in the germination of naturally occurring populations of buried weed seeds. J. Exp. Bot. 1969, 20, 402–413. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Chancellor, R.J.; Drennan, D.S.H. The Effects of Seed Burial and Soil Disturbance on Emergence and Survival of Arable Weeds in Relation to Minimal Cultivation. J. Appl. Ecol. 1984, 21, 629. [Google Scholar] [CrossRef]
- Mohler, C.L. Effects of Tillage and Mulch on Weed Biomass and Sweet Corn Yield. Weed Technol. 1991, 5, 545–552. [Google Scholar] [CrossRef]
- Kaspar, T.C.; Erbach, D.C. Improving stand establishment in no-till with residue-clearing planter attachments. Trans. Am. Soc. Agric. Eng. 1998, 41, 301–306. [Google Scholar]
- Garcia, A.G.; Guerra, L.C.; Hoogenboom, G. Impact of planting date and hybrid on early growth of sweet corn. Agron. J. 2009, 101, 193–200. [Google Scholar] [CrossRef]
- Bullock, D.G.; Anderson, D.S. Evaluation of the Minolta SPAD-502 chlorophyll meter for nitrogen management in corn. J. Plant Nutr. 1998, 21, 741–755. [Google Scholar] [CrossRef]
- Kurtz, T.; Melsted, S.W.; Bray, R.H. The Importance of Nitrogen and Water in Reducing Competition Between Intercrops and Corn 1. Agron. J. 1952, 44, 13–17. [Google Scholar] [CrossRef]
- Hardman, W.C.; Catchot, A.L.; Gore, J.; Henry, W.B.; Cook, D.R. The Impact of Brown Stink Bug (Hemiptera: Pentatomidae) Damage during the Seedling Stage on Field Corn Growth and Yield. J. Econ. Entomol. 2021, 114, 1607–1612. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).