Abstract
Solar radiation entering a high-wire tomato greenhouse is mostly intercepted by the top of the crop canopy, while the role of lower leaves diminishes with age, turning them into sink organs rather than sources. Accordingly, the defoliation of basal leaves is a widely applied agronomic practice in high-wire greenhouse cultivation management. However, the recent increase in the application of supplemental light emitting diode (LED) lighting for high-density tomato production may affect the role of basal leaves, promoting their source role for fruit development and growth. The present research aims to explore the application of supplementary LED lighting on Solanum lycopersicum cv. Siranzo in the Mediterranean area during the cold season in combination with two regimes of basal defoliation. The defoliation factors consisted of the early removal of the leaves (R) right under the developing truss before the fruit turning stage and a non-removal (NR) during the entire cultivation cycle. The lighting factors consisted of an artificial LED lighting treatment with red and blue diodes for 16 h d−1 (h 8-00) with an intensity of 180 µmol s−1 m−2 (RB) and a control cultivated under natural light only (CK). The results demonstrated a great effect of the supplemental LED light, which increased the total yield (+118%), favoring fruit setting (+46%) and faster ripening (+60%) regardless of defoliation regimes, although the increased energy prices hinder the economic viability of the technology. Concerning fruit quality, defoliation significantly reduced the soluble solid content, while it increased the acidity when combined with natural light.
1. Introduction
Defoliation is an agronomic practice widely used for tomato production. It consists of the removal of leaves below a cluster before the plant’s harvest, with timing and intensity defined by the management choices of the producer. The objectives of defoliation include increasing aeration between plants, limiting disease development, promoting vertical light penetration within the canopy, and facilitating crop operations [1]. Basal leaf removal may also affect yield, reducing the sink effect of the less-illuminated leaves in the lower part of the canopy and promoting the accumulation of assimilates in developing fruits [2].
The link between defoliation and yield is determined by the translocation and partitioning of assimilates between plant organs. Translocation refers to the movement of photo-assimilates from the producing organs, or sources, to the recipient organs, or sinks, while partitioning refers to the proportion by which assimilates are allocated to the various sinks [3,4]. The two processes are crucial in determining the accumulation of dry matter in fruits, and therefore affect the final yield [5]. The differentiation of organs into source and sink is not always clear, since one organ can take on both roles [6]. Moreover, each organ possesses a certain sink strength, being more or less capable of attracting the products of photosynthesis due to both physical (e.g., number of cells) and molecular (e.g., enzymatic activity) features [7,8].
Cultivation density, and consequently vertical light distribution, can affect leaves’ sink–source role. Research showed that in high planting density conditions (approximately 30 cm of spacing on the row), basal leaves seem to experience a strong competition for light, contributing little to the plants’ net photosynthesis and becoming sink organs. On the other hand, a larger spacing (approximately 50 cm of spacing on the row) can significantly contribute to the production of assimilates, making defoliation before harvest not favorable for fruit development [9]. Highly dense cultivation is the standard in high-tech greenhouses, in which the use of supplemental LED interlighting was already proven to be capable of increasing yield and other quality aspects of greenhouse tomato production, affecting plant photosynthesis and the plants’ photoreceptor responses [10,11]. The present research aims to evaluate if the application of two defoliation regimes, namely, removal at the fruit turning stage and non-removal, combined with two lighting regimes, namely, supplemental LED light treatment and a control grown with natural light, can significantly affect the role of basal leaves and, consequently, the yield and quality of tomato production.
2. Materials and Methods
2.1. Plants Growing Conditions and Treatments
Truss-tomato plants (Solanum lycopersicum L. cv. Siranzo; Rijk Zwaan, The Netherlands) were cultivated in a glass-glazed greenhouse in Bologna, Italy (44°29′38″ N, 11°20′34″ E). The seedlings were produced in rockwool cubes (Grodan Vital, Roermond, The Netherlands) by a local nursery from mid-August and were transplanted in perlite bags at a distance of 20 cm on the row on 23 September 2021. The environmental conditions (temperature, relative humidity, and solar radiation) were monitored daily during the entire growing period. Both passive (lateral and top openings) and active (fan and heat pump) climate control strategies were used to maintain constant conditions (Tmean 22 °C; RHmean 64%). Fertigation was performed using an open drip irrigation system, providing a solution with an average pH of 6.6 and an electrical conductivity (EC) of 2.8 dS m−1 (Table 1). Supplemental lighting was provided by single-LED interlighting lamps (Flygrow Interlight, Flytech LED Technology, Belluno, Italy) located at 30 cm of distance from the stem and at a height of 1.40 cm from the rockwool cube throughout the entire growing period (Figure S1). Two lighting regimes were applied: a control illuminated with natural light only (namely CK), and an illuminated treatment supplied with red (660 nm) and blue (465 nm) LED lights (RB ratio of 3) with a photosynthetic photon flux density (PPFD) of 180 µmol s−1 m−2 (measured at 30 cm from the plant) and a photoperiod of 16 h d−1 (namely, RB). Lighting treatments were applied from the transplanting day (23 September 2021) until the end of the experiment (16 February 2022). Since the beginning of November, the plants under the two lighting regimes underwent two defoliation regimes, which consisted of an early removal of the leaves right under the developing truss before the fruit turning point (R) and a non-removal during the entire cultivation cycle (NR). A split-plot design with four replicates was used, and each sub-plot contained five plants.

Table 1.
Formulation of standard nutrient solution used for tomato cultivation.
2.2. Plant Vegetative, Physiological, and Biochemical Measurements
Collar diameter was measured once at the end of October at height of 1 cm from the rockwool cube. The internodes length was measured at the end of October as the distance among the fourth and fifth fruit trusses. Plant topping occurred on 23 November, above the sixth truss. The total leaf area was evaluated at the end of the experiment using a leaf area meter (LI-3100C Area Meter, LI-COR Biosciences, Lincoln, NE, USA) on NR plants cultivated with RB or CK treatment. Leaves and stems were weighted with a digital scale, both fresh and after being dried at 60 °C for 4 days. Weight measurements were used to evaluate the leaf dry matter content (LDMC), which is the ratio between the leaf dry mass and the leaf fresh mass (mg g−1), and the specific leaf area (SLA), which is the ratio between leaf area and leaf dry mass (m2 kg−1) [12].
Chlorophyll content of leaves was evaluated in mid-December, considering two points of the first leaf right under the third and fourth fruit trusses. A SPAD-502PLUS (Konica Minolta, Tokyo, Japan) was used to non-disruptively estimate the chlorophyll content. Leaf gas exchange and chlorophyll fluorescence were evaluated using a portable LI-COR 6400 (LI-COR Biosciences, Lincoln, United States), which was set as reported by Calone et al. [13]. In particular, the following parameters were measured: stomatal conductance (GS, in mmol m−2 s−1), under-leaf CO2 concentration (Ci), leaf transpiration (E, in mmol m−2 s−1), net photosynthesis (A, in µmol m−2 s−1), and the effective quantum yield efficiency of PSII (ΦPSII). The ΦPSII, which represents the capacity of photosystem II (PSII) to absorb photon energy, has been calculated as (Fm’-Fs)/Fm’, where Fm’ represents the maximum fluorescence in a leaf adapted to light and Fs represents the steady-state fluorescence. The ΦPSII has been further split in two components: the PSII maximum efficiency (Fv’/Fm’) and the level of photochemical quenching of PSII (qP). The first component (Fv’/Fm’) represents the maximum operational efficiency in a leaf adapted to light. It is calculated as (Fm’—Fo’)/Fm’, where Fo’ is the minimum fluorescence. The second component (qP) represents the real number of active reaction centers of PSII, and is calculated as (Fm’-Fs)/(Fm’-Fo’).
2.3. Fruit Development and Yield
Fruit development was monitored on proximal and distal fruit of both the first cluster (Follow-Up 1) and the third cluster (Follow-Up 2) by measuring the equatorial and polar diameters with a digital Vernier caliper before harvesting. Fruit volume (mm3) was estimated as the volume of an ellipsoid of rotation V = (4/3) πab2, where a is one half of the polar diameter and b is one half of the equatorial diameter [14]. Fruit ripening was evaluated on the same fruit two weeks before harvesting by using a DA-Meter (Sinteleia, Bologna, Italy), which non-destructively evaluated the chlorophyll degradation and correlated it with a ripening index.
From the end of November until the end of the trial, fruits were harvested (in total, 6 clusters per plant). The fresh weight of the total clusters of each plant in each treatment was measured with a digital scale. The number of fruits per cluster was counted for each plant. At harvesting, fruits were divided, counted, and weighted as mature (dark orange or red tomatoes) or immature (green, light green, and light orange) [15]. Fruit productive units (flowers and buds) were counted at the beginning of truss development and were confronted with the number of fruits at harvesting to assess fruit setting.
2.4. Fruit Quality
2.4.1. Non-Destructive Measurements
Tomatoes used for qualitative analysis (n = 12) were selected considering a DA-Meter range index ranging 1.30 to 1.50. The fruit hardness was assessed using a Durofel device (Giraud Technologies, Cavaillon, France) on four opposite sides of the equatorial diameter of each fruit per treatment per block. The instrument non-destructively measured the elasticity of the fruit exocarp, expressing it in a Durofel Index ranging from 0 to 100.
Color was evaluated using a CIE Lab color space analysis, where the L* component represents the lightness from black (0) to white (100), the a* component is a value ranging from green (−) to red (+), and the b* component is a value ranging from blue (−) to yellow (+). A colorimeter (Chroma Meter CR-400, Minolta, Tokyo, Japan) was used to assess the values. The measures were performed on four opposite sides at an equatorial level. Two indexes, the HUE angle (h) and the Chroma (C), were deduced from the a* and b* components by applying the formulas tan−1 (b*/a*)2 and (a*2 + b*2)0.5, respectively [16].
2.4.2. Destructive Measurements
Destructive measurements included the pulp firmness, soluble solids content, and titratable acidity evaluation. Pulp firmness was determined using a fruit-texture analyzer (FTA GÜSS, Strand, South Africa), evaluating the force required to penetrate the fruit. The penetration was performed with a cylindrical and flat-end probe with a 6 mm diameter, at a depth equal to 11 mm and a speed of 30 mm s−1. Measurements were performed on four opposite sides of the equatorial diameter, peeling the fruit before penetration. The soluble solids content was evaluated on each centrifuged fruit using a digital refractometer model PAL-1 (Atago Co., Ltd., Tokyo, Japan). The titratable acidity was measured with an automatic TitroMatic (Compact-S titrator, Crison, Modena, Italy) by diluting 20 mL of tomato juice in 20 mL of distilled water. The titratable acidity was estimated by titrating with 0.1 N NaOH until the titration endpoint at a pH of 8.1.
The fruit dry matter content (FDMC) was evaluated on 3 fruits per treatment per block, other than those fruits used for destructive and non-destructive qualitative analysis, as the ratio between the fruits’ fresh weight and dried weight at 65 °C per 1 week.
2.5. Biochemical Analysis
Lycopene and β-carotene content
Lycopene and β-carotene content were evaluated on 6 fruits per treatment using the methodology described by Anthon and Barrett [17], applying slight modifications. An extraction solution was prepared by mixing hexane, acetone, and ethanol in a v:v:v proportion of 2:1:1, with the addition of 0.5 g L−1 of butylated hydroxytoluene. Then, 0.5 g of homogenized, frozen sample, including the exocarp and mesocarp, were mixed with 10 mL of the extraction solution. The material was left in darkness for 30 min and then centrifuged at 2000× g for 5 min. Finally, 1 mL of supernatant was read at 503 and 444 nm with a spectrophotometer (Biochrom Ltd., Cambridge, England).
The lycopene content was calculated using the following formula (Anthon and Barrett, 2006):
where X is the volume of hexane phase (mL, sew below), Y the weight of the fruit tissue (g), A503 is the absorbance at 503 nm, and 3.12 is the extinction coefficient. β-carotene was calculated with the following equation [17]:
where 444 is the absorbance at 444 nm, 503 is the absorbance at 503 nm, 0.55 is the ratio of the final hexane layer volume to the volume of mixed solvents added for hexane:acetone:ethanol (2:1:1), is the volume of mixed solvents, W is the fresh weight of the sample, and 537 (g mol−1) is the molecular weight of β-carotene.
2.6. Energy Cost Assessment
The energy cost assessment was performed considering the actual consumption of a lamp with red (660 nm) and blue (465 nm) diodes at a ratio of 3 (1.68 kWh) applied for 16 h per day. Costs were estimated per plant per day, considering that a single lamp was able to provide supplementary lighting to approximately 6 plants. The cost per plant was then used to calculate the cost per kg. The price of electricity was acquired from a EUROSTAT [18] dataset, considering electricity prices for non-household consumers (excluding VAT and other recoverable taxes and levies) in Italy, referring to both the second semester of 2021 (0.1853 € kWh−1) and the first semester of 2022 (0.2525 € kWh−1). For the final electricity cost calculation, considering the growing cycle of 147 days, the price of the second semester of 2021 was used for 100 days (from 23 September 2021 to 31 December 2021), and the price of the first semester of 2022 was used for 47 days (from 1 January 2022 to 16 February 2022).
2.7. Statistical Analysis
Data analysis was performed with a two-way ANOVA by comparing the lighting factor with the defoliation factor. Data were analyzed by using IBM SPSS Statistics 28.0.1.0.
3. Results
Results of the two-way ANOVA are reported in Table S1. The evaluation of the vegetative parameters showed a significant increase in the collar diameter of plants grown with a supplementary LED light (+34%) (RB 13.1 ± 2.2 mm, CK 9.8 ± 1.5 mm), regardless of the type of defoliation (data not shown). On the contrary, internode length, total leaf area, leaf fresh and dry weight, LDMC, plant total fresh and dry weight, and stem length did not show any statistical difference among treatments (data not shown). However, the measurement of SLA resulted in a significant increase (+41%) in the case of the CK (181 ± 21 cm2 g−1) treatment compared to the RB treatment (128 ± 19 cm2 g−1).
The physiological response evaluation considered several parameters. Significant differences were, however, only observed in the case of chlorophyll content (SPAD Index), net photosynthesis (A), effective quantum yield efficiency of PSII (ΦPSII and photochemical quenching of PSII (qP). Particularly, SPAD Index results were significantly higher in plants under RB treatment, both in the leaves under the third (+23%) and fourth (+17%) trusses, independent of the defoliation regime (Figure 1). Similarly, the net photosynthesis (A) was higher in plants exposed to the RB treatment compared to the CK but was not altered by the defoliation regime (Table 2). On the other hand, ΦPSII and qP showed significant differences not only among light treatments but also among defoliations regimes, reporting higher levels in the case of RB light treatment and leaf removal (R) (Table 2), although it did not show an interaction among factors (Table S1). The PSII maximum efficiency (Fv/Fm), stomatal conductance (GS), transpiration (E) and under-leaf CO2 concentration (Ci) were not affected by the lighting and defoliation factors (data not shown).
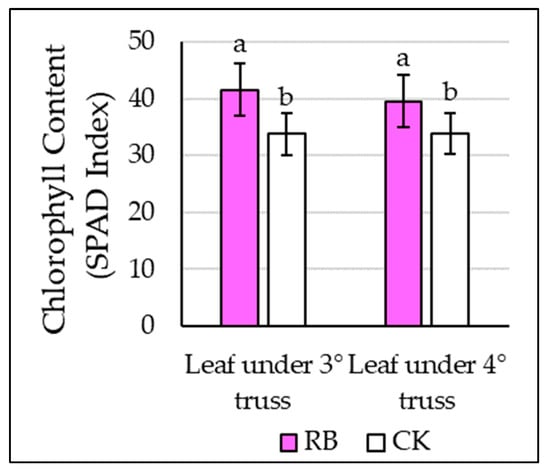
Figure 1.
Effects of supplemental red and blue LED light (RB) and natural light only (CK) on leaves’ chlorophyll content (SPAD Index). Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05). Vertical bars represent standard errors.
Table 2.
Effects of the two separated factors, light (supplemental red and blue LED light, RB, and natural light, CK) and defoliation (early leaf removal, R, and no-removal, NR), on net photosynthesis (A), effective quantum yield efficiency of PSII (ΦPSII) and photochemical quenching of PSII (qP). Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05).
In Follow-Up 1, the volume of the proximal fruit was significantly higher in the case of plants subjected to defoliation (R) but was not affected by the lighting regime (Table 3). On the contrary, the distal fruit of Follow-Up 1 were significantly bigger in the case of RB treatment compared to CK but were not significantly influenced by defoliation (Table 3). During the Follow-Up 2, the presence of supplemental RB light significantly affected fruit dimension in both the proximal and distal fruit, while the defoliation regime did not cause any significant effect (Table 3). A significant interaction between the lighting and defoliation factors was observed for the ripening (measured by DA-Meter values) of proximal fruit during Follow-Up 1 (Table S1). Indeed, the ripening of proximal fruit was increased by early defoliation (R) in RB treated plants compared to the NR plants, while no significant effects were observed in the CK plants (Figure 2a). Conversely, ripening was not affected by either lighting or defoliation in the distal fruit during Follow-Up 1 (data not shown). During Follow-Up 2, RB light significantly increased the DA-Meter values in both proximal and distal fruit (Figure 2b,c). In general, two weeks before harvesting, the RB treatment resulted in a higher rate (+60%) of ripened fruits when compared to CK.
Table 3.
Effects of the two separated factors, light (supplemental red and blue LED light, RB, and natural light, CK) and defoliation (early leaf removal, R, and no-removal, NR), on fruit volume (cm3) in proximal and distal tomatoes of Follow-Up 1 and Follow-Up 2 Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05).
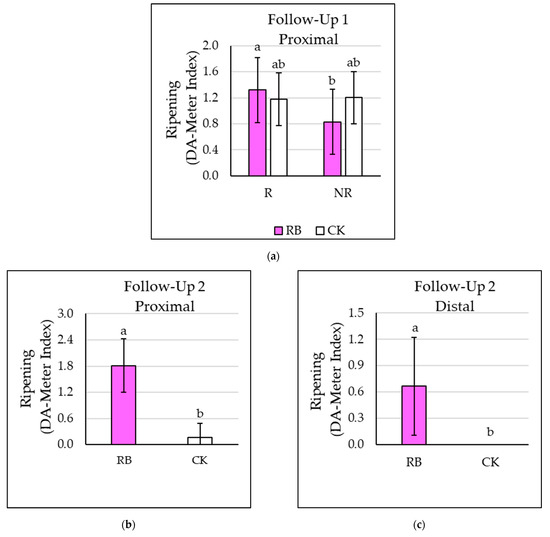
Figure 2.
Effect of light (supplemental red and blue LED light, RB, and natural light only, CK) and defoliation (leaves early removal, R, and no-removal, NR) on (a) proximal fruits of Follow-Up 1 (b) proximal fruits of Follow-Up 2, and (c) distal fruits of Follow-Up 2. Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05). Vertical bars represent standard errors.
The total fruit yield results were significantly increased in the case of supplemental RB light application, with a doubled production (+118%) when compared to the CK (Table 4). LED light application also significantly affected the yield of green and red tomatoes, as is summarized in Table 4. Furthermore, differences were observed for the number of aborted and productive units (flowers and buds), respectively, showing a decrease (−54%) in flower abortions (RB 0.7 ± 1.0, CK 1.6 ± 1.5 of aborted units) and an increase (+46%) in fruit setting (RB 6 ± 1.5, CK 4.1 ± 2.2 of set fruits) in the case of RB light application when compared to the CK (data not shown). However, no significant differences were observed in the cases of different defoliation regimes for yield, flower abortion, and fruit setting (Table S1).
Table 4.
Effect of supplemental red and blue LED light (RB) and natural light only (CK) on tomato yield. Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05).
The fruit dry matter content (FDMC), Chroma (C), fruit hardness, and pulp firmness were not affected by both factors (data not shown). Among color determinations, the HUE angle (h) and lightness (L*) presented significantly higher levels in the case of tomatoes grown under natural light only (CK) (Figure 3a,b), while no significant differences were observed depending on defoliation regime. The soluble solids content showed statistically significant higher levels in the case of tomatoes from plants not subjected to leaf removal (NR) (+3%) (Figure 3c), while no differences were observed depending on the light treatment. A significant interaction between lighting and defoliation factors was observed for fruit acidity (Table S1). Indeed, fruit acidity was increased by early defoliation (R) in control plants when compared to RB-treated ones, while no differences between lighting regimes were observed in NR plants (Figure 3d).
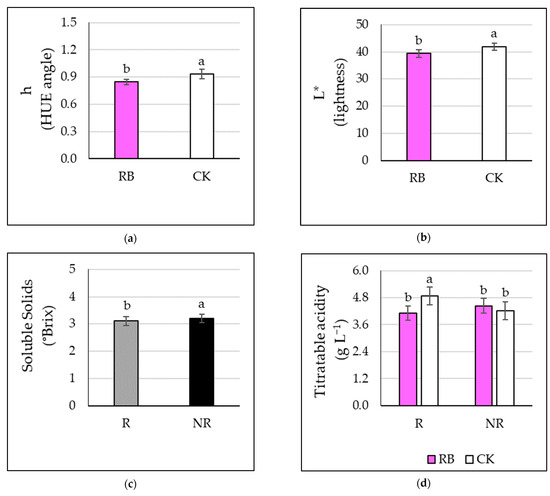
Figure 3.
Effect of light (supplemental red and blue LED light, RB, and natural light only, CK) and defoliation (leaves early removal, R, and no-removal, NR) on (a) HUE angle, (b) lightness, (c) soluble solids and (d) acidity of tomato. Different letters indicate significant differences at ANOVA (significant level p ≤ 0.05). Vertical bars represent standard errors.
Finally, none of the applied factors resulted in changes in either lycopene or β-carotene content (data not shown).
The energy consumption resulted in approximately 1.68 kWh per day per lamp. Considering this consumption and the electricity costs (as reported in Section 2.6), the total cost of supplemental LED lighting during the fall–winter period was approximately 8.51 € plant−1, leading to 2.65 € kg−1.
4. Discussion
Light is a fundamental factor for plant vegetative and architectural development. The effect of specific wavelengths on plant morphology are determined by the stimulation of photoreceptors, which can induce responses that ameliorate leaf exposition to solar radiation and, consequently, photosynthesis [19,20]. This is particularly evident in case of the so called Shade Avoidance Syndrome, in which a low Red/Far-Red ratio (R:FR) can induce phytochromes, the photoreceptors sensitive to the red wavelengths, to promote plant elongation and escape sub-optimal lighting conditions [21]. Blue light can also influence plant morphology, affecting the leaf area expansion [22]. In the present research, red and blue light limitedly influenced the vegetative parameters, reporting a significant increase only in the cases of collar diameter and SLA (Table S1). A similar increase in the tomato collar diameter in response to a supplemental RB light application was formerly observed by Paucek et al. [15]. On the other hand, the significantly lower SLA value in RB seems to confirm the common response of plants in the case of high light intensity, in which leaf thickness, which is determined by increased palisade tissue, can serve as a protective response against excessive irradiation [23].
Regarding physiological parameters, the higher chlorophyll content (Figure 1) and, consequently, the higher net photosynthesis (Table 2) observed in leaves under supplemental RB light can be associated to the actions of phytochromes and cryptochromes, which are stimulated by red and blue light components, respectively [24,25]. Regardless of the lighting factors in this study, defoliation of leaves normally reduces water evapotranspiration [26]. However, the hereby presented results showed absence of significant effects. Previous research observed that a reduction in the source–sink ratio may reduce transpiration and increase the efficiency of PSII in tomato plants, especially under limited nutritive conditions [27].
Defoliation affected fruit dimension only in the case of proximal fruits during Follow Up 1, while no differences were observed for distal fruit or during Follow-Up 2 (Table 3). On the contrary, supplemental LED light significantly influenced the development of both proximal and distal fruits during Follow-Up 2 (Table 3). While in the case of fruits of Follow-Up 1, supplemental LED light application began with proximal fruits already formed on the inflorescence, the fruits of Follow-Up 2 began their development already under the light treatment. As observed in the previous research [28,29], the major dimension of RB fruits may be related to a major sink strength, which is, in turn, determined by a possible stimulation of initial cell division using LED light rather than a stimulation during the enlargement phase.
As was already observed by other authors [15], fruit maturation precocity seemed to be significantly increased by LED light, showing more evident effects, especially later along in the autumn season (December—Follow-Up 2). Regarding fruit dimensions, leaf removal seemed to have a significantly higher effect on ripening only during the earlier stages of production (Follow-Up 1) and in the case of proximal fruits (Figure 3a). Although these effects can be associated with lower assimilates’ competition with other sink organs, including lower leaves, this observation was only related to the first period of production, during which the outside lighting condition was not excessively limited.
The qualitative characteristics of tomatoes, such as their color, flavor, and firmness, are of fundamental importance as they influence consumers’ perception [30]. In the present research, supplemental LED light limitedly affected the qualitative traits. The carotenoid content was often reported to increase in the case of LED treatments, especially in the case of red light application, given a possible involvement of fruit phytochromes [31,32,33]. However, the present study did not present any significant difference. On the other hand, defoliation determined a significant decrease in the soluble solids content (Figure 3c), a possible consequence of the imbalance between the sink/source equilibrium [34].
From an economic point of view, the additional electricity costs incurred when greenhouse artificial lighting is applied should be counterbalanced by the observed yield increase (+118%). However, if an overall increase in costs associated with energy consumption could be estimated at 2.65 € kg−1, when deducting the achievable yield under solar radiation only from the calculation, the actual costs per kg were even higher, namely 4.82 € kg−1. Considering that the producers’ price of vine tomatoes in January 2022 accounted for 1.82 € kg−1 in Italy [35], the application of supplemental LED lighting has not been proven to be economically viable. However, a number of concurrent factors may have affected these performances. From a cost perspective, energy costs increased from January 2021 (when the experiment was designed) by 15 and 37% after 6 and 12 months, respectively. Although an extended, artificial photoperiod was used (16 h d−1) throughout the growing cycle, some authors suggested avoiding supplementary lighting when solar radiation is sufficient [36] or limiting it to specific phenological stages to reduce energy consumption. On the other hand, the estimated income is linked to both agronomic yield and producer price. The short-term nature of the experiment may have hindered the final yield. The research was performed by top-pruning plants at the sixth truss instead of the minimal 15–20 trusses commonly obtained from the same cultivar in local, commercial greenhouses [15]. In addition, the adopted cultivar, though adapted to high-wire intensive cultivation, is associated with low prices in the Italian market (e.g., approximately half the price of cherry tomatoes [35]).
5. Conclusions
Although defoliation is considered the best practice to guarantee easier management, favor ventilation, and control pest development, this research revealed that this practice can limitedly influence the tomato yield and quality in the case of supplemental LED light application. The use of supplemental LED light demonstrated interesting potentialities for tomato production during the cold season in Mediterranean countries, increasing the total yield (+118%), favoring fruit setting (+46%), and hastening ripening (+60%). In relation to the energy costs and measured yield, however, the use of supplemental lighting was not proven to be economically feasible under the tested conditions. Accordingly, future studies should match the energy saving strategies (e.g., reducing light integrals by disruptive lighting protocols) with a further characterization of both genotypic and phenotypic responses to lighting. Finally, the application of new greenhouse technologies that transform the solar spectrum into electricity, namely, photoconversion covers that integrate semi-transparent photovoltaic cells [37], may represent a solution to integrating supplemental lighting for crop cultivation with reducing electricity costs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13030671/s1, Table S1: Results of two-way ANOVA. Figure S1. Descriptive scheme of LED lamp location.
Author Contributions
E.A. designed and performed the experiment and data collection and drafted the manuscript. I.P. contributed to the data collection and critically revised the manuscript. L.M. contributed to the data collection. G.P., F.O., X.G., and G.G. supervised the research and critically revised the manuscript. All authors contributed to the article and approved the submitted version. All authors have read and agreed to the published version of the manuscript.
Funding
The research leading to this publication has received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement no. 862663. Furthermore, this study was carried out within the Agritech National Research Center and received funding from the European Union Next-GenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR)—MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4—D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions: neither the Research Executive Agency (REA), the European Union, nor the European Commission can be considered responsible for them.
Data Availability Statement
Data are available in AMS Acta repository at the following link: http://amsacta.unibo.it/id/eprint/7180.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Heuvelink, E.; Okello, R.C. Developmental processes. In Tomatoes; Heuvelink, E., Ed.; CABI: Wallingford, UK, 2018; Volume 27, pp. 59–88. [Google Scholar] [CrossRef]
- Jo, W.J.; Shin, J.H. Effect of Leaf-Area Management on Tomato Plant Growth in Greenhouses. Hortic. Environ. Biotechnol. 2020, 61, 981–988. [Google Scholar] [CrossRef]
- Osorio, S.; Ruan, Y.L.; Fernie, A.R. An update on source-to-sink carbon partitioning in tomato. Front. Plant Sci. 2014, 5, 516. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, R.K.; Bishnoi, C. Assimilate partitioning and distribution in fruit crops: A review. J. Pharmacogn. Phytochem. 2017, 6, 479–484. [Google Scholar] [CrossRef]
- Ronga, D.; Zaccardelli, M.; Lovelli, S.; Perrone, D.; Francia, E.; Milc, J.; Ulrici, A.; Pecchioni, N. Biomass production and dry matter partitioning of processing tomato under organic vs conventional cropping systems in a Mediterranean environment. Sci. Hortic. 2017, 224, 163–170. [Google Scholar] [CrossRef]
- Aslani, L.; Gholami, M.; Mobli, M.; Sabzalian, M.R. The influence of altered sink-source balance on the plant growth and yield of greenhouse tomato. Physiol. Mol. Biol. Plants 2020, 26, 2109–2123. [Google Scholar] [CrossRef] [PubMed]
- Bihmidine, S.; Hunter III, C.T.; Johns, C.E.; Koch, K.E.; Braun, D.M. Regulation of assimilate import into sink organs: Update on molecular drivers of sink strength. Front. Plant Sci. 2013, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Forney, C.; Bondada, B.; Leng, F.; Xie, Z.S. The molecular regulation of carbon sink strength in grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 11, 606918. [Google Scholar] [CrossRef]
- Hachmann, T.L.; Echer, M.d.M.; Dalastra, G.M.; Vasconcelos, E.S.; Guimarães, V.F. Tomato cultivation under different spacings and different levels of defoliation of basal leaves. Bragantia 2014, 73, 399–406. [Google Scholar] [CrossRef]
- Appolloni, E.; Orsini, F.; Pennisi, G.; Gabarrell Durany, X.; Paucek, I.; Gianquinto, G. Supplemental LED lighting effectively enhances the yield and quality of greenhouse truss tomato production: Results of a meta-analysis. Front. Plant Sci. 2021, 12, 596927. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.O. Spectral effects of artificial light on plant physiology and secondary metabolism: A review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Garnier, E.; Shipley, B.; Roumet, C.; Laurent, G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Funct. Ecol. 2001, 15, 688–695. [Google Scholar] [CrossRef]
- Calone, R.; Cellini, A.; Manfrini, L.; Lambertini, C.; Gioacchini, P.; Simoni, A.; Barbanti, L. The C4 Atriplex halimus vs. The C3 Atriplex hortensis: Similarities and Differences in the Salinity Stress Response. Agronomy 2021, 11, 1967. [Google Scholar] [CrossRef]
- Li, T.; Heuvelink, E.P.; Marcelis, L.F. Quantifying the source–sink balance and carbohydrate content in three tomato cultivars. Front. Plant Sci. 2015, 6, 416. [Google Scholar] [CrossRef] [PubMed]
- Paucek, I.; Pennisi, G.; Pistillo, A.; Appolloni, E.; Crepaldi, A.; Calegari, B.; Spinelli, F.; Cellini, A.; Gabarrell, X.; Orsini, F. Supplementary LED interlighting improves yield and precocity of greenhouse tomatoes in the Mediterranean. Agronomy 2020, 10, 1002. [Google Scholar] [CrossRef]
- Lopez Camelo, A.; Gomez, P. Modelling postharvest colour changes in long shelf life tomatoes. In XXV International Horticultural Congress; Acta Horticulturae: Brussel, Belgium, 1998; p. 9. [Google Scholar]
- Anthon, G.; Barrett, D.M. Standardization of a rapid spectrophotometric method for lycopene analysis. Acta Hortic. 2006, 758, 111–128. [Google Scholar] [CrossRef]
- EUROSTAT. Available online: https://ec.europa.eu/eurostat/en/ (accessed on 14 December 2022).
- Li, L.; Ljung, K.; Breton, G.; Schmitz, R.J.; Pruneda-Paz, J.; Cowing-Zitron, C.; Cole, B.J.; Ivans, L.J.; Pedmale, U.V.; Jung, H.S. Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 2012, 26, 785–790. [Google Scholar] [CrossRef]
- Dierck, R.; Dhooghe, E.; Van Huylenbroeck, J.; Van Der Straeten, D.; De Keyser, E. Light quality regulates plant architecture in different genotypes of Chrysanthemum morifolium Ramat. Sci. Hortic. 2017, 218, 177–186. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Wang, X.Y.; Xu, X.M.; Cui, J. The importance of blue light for leaf area expansion, development of photosynthetic apparatus, and chloroplast ultrastructure of Cucumis sativus grown under weak light. Photosynthetica 2015, 53, 213–222. [Google Scholar] [CrossRef]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blue plus red light control of chlorophyll content and CO2 gas exchange in barley leaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef]
- Balducci, L.; Fierravanti, A.; Rossi, S.; Delzon, S.; De Grandpré, L.; Kneeshaw, D.D.; Deslauriers, A. The paradox of defoliation: Declining tree water status with increasing soil water content. Agric. For. Meteorol. 2020, 290, 108025. [Google Scholar] [CrossRef]
- Glanz-Idan, N.; Wolf, S. Upregulation of photosynthesis in mineral nutrition-deficient tomato plants by reduced source-to-sink ratio. Plant. Signal. Behav. 2020, 15, 1712543. [Google Scholar] [CrossRef] [PubMed]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I.A. Supplemental light-emitting diode inter-lighting increases tomato fruit growth through enhanced photosynthetic light use efficiency and modulated root activity. Front. Plant Sci. 2020, 10, 1656. [Google Scholar] [CrossRef]
- Appolloni, E.; Paucek, I.; Pennisi, G.; Stringari, G.; Gabarrell Durany, X.; Orsini, F.; Gianquinto, G. Supplemental LED Lighting Improves Fruit Growth and Yield of Tomato Grown under the Sub-Optimal Lighting Condition of a Building Integrated Rooftop Greenhouse (i-RTG). Horticulturae 2022, 8, 771. [Google Scholar] [CrossRef]
- Pinheiro, J.; Alegria, C.; Abreu, M.; Gonçalves, E.M.; Silva, C.L. Kinetics of changes in the physical quality parameters of fresh tomato fruits (Solanum lycopersicum, cv.‘Zinac’) during storage. J. Food Eng. 2013, 114, 338–345. [Google Scholar] [CrossRef]
- Alba, R.; Cordonnier-Pratt, M.M.; Pratt, L.H. Fruit-localized phytochromes regulate lycopene accumulation independently of ethylene production in tomato. Plant. Physiol. 2000, 123, 363–370. [Google Scholar] [CrossRef]
- Nájera, C.; Guil-Guerrero, J.L.; Enríquez, L.J.; Álvaro, J.E.; Urrestarazu, M. LED-enhanced dietary and organoleptic qualities in postharvest tomato fruit. Postharvest Biol. Technol. 2018, 145, 151–156. [Google Scholar] [CrossRef]
- Dannehl, D.; Schwend, T.; Veit, D.; Schmidt, U. Increase of yield, lycopene, and lutein content in tomatoes grown under continuous PAR spectrum LED lighting. Front. Plant Sci. 2021, 12, 611236. [Google Scholar] [CrossRef]
- Casierra-Posada, F.; Torres, I.D.; Blanke, M.M. Fruit quality and yield in partially defoliated strawberry plants in the tropical highlands. Gesunde Pflanz. 2013, 65, 107–112. [Google Scholar] [CrossRef]
- EC (2022). Available online: https://agriculture.ec.europa.eu/data-and-analysis/markets/overviews/market-observatories/fruit-and-vegetables/tomatoes-statistics_en (accessed on 14 December 2022).
- Niu, Y.; Lyu, H.; Liu, X.; Zhang, M.; Li, H. Effects of supplemental lighting duration and matrix moisture on net photosynthetic rate of tomato plants under solar greenhouse in winter. Comput. Electron. Agric. 2022, 198, 107102. [Google Scholar] [CrossRef]
- Paskhin, M.O.; Yanykin, D.V.; Gudkov, S.V. Current Approaches to Light Conversion for Controlled Environment Agricultural Applications: A Review. Horticulturae 2022, 8, 885. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).