Abstract
Wheat production worldwide is severely challenged by different pest and pathogen species, and misguided application of chemicals in farming significantly stimulates crop invaders to develop insecticide resistance. The growing desire for sustainable agriculture has promoted the breeding of wheat cultivars with durable pests and diseases resistance tolerance traits, which have become increasingly imperative for wheat production. Various genetic breeding techniques, including transgenic breeding, molecular marker-assisted breeding, molecular breeding with gene pyramiding, gene silencing, and gene editing, have been successfully deployed into wheat cultivar improvement, and have significantly promoted the process of breeding cultivars with multiple substantial resistance traits. Although the success of practical applications of some of these resistant wheat cultivars in farming has suggested that these approaches are effective strategies for combating fungal diseases and herbivorous insects, there remains significant challenges for wheat molecular breeding programs. Thus, this review illustrates and summarizes the recent advances and drawbacks related to the measures associated with molecular breeding in common wheat cultivar improvement.
1. Introduction
Bread wheat (Triticum aestivum L.) is the most cultivated cereal worldwide, occupying approximately 1/6 of total global arable land area (FAOSTAT, 2021). The rapidly growing world population and urbanization have dramatically increased the demand for improving wheat production at an annual rate of 2% in limited land resources [1,2]. However, wheat production is severely challenged by different types of biotic and abiotic stresses. Among these stresses, phytopathogens and herbivorous insects, as well as the viruses transmitted by these pests, can cause substantial damage to wheat grain yield and quality. The combined presence of different species of colonizing pests typically further aggravates the yield and quality losses in wheat grain [3,4]. In addition, returning straw litter back into wheat fields also potentially increases the incidence of plant diseases and insect pests in the subsequent season of crop growth [5]. To guarantee that grain yields meet the demands for food security, large amounts of chemical fertilizers and pesticides are being applied to agroecosystems; however, the economically misguided application of these chemicals in farming has considerably exacerbated environmental issues and pest resurgence [6]. Therefore, the building or biofortification of durable pest and disease resistance in diverse elite wheat cultivars have become increasingly imperative for global wheat production.
To effectively breed resistant wheat cultivars, transferring foreign genes (isolated from either the same or distinct species)-based wheat cultivar improvements contributed greatly to increasing crop yields by contrasting new or improved traits in the early post-genomics era. Recently, extensive studies have typically focused on the identification of more inherent pest- and disease-resistance genes or their closely related molecular markers. In addition, the rapid development of high-throughput sequencing and bioinformatics tools have significantly facilitated fine-mapping or cloning of these resistance genes. The application of marker-assisted selection (MAS) and molecular breeding programs integrating gene pyramiding will therefore necessitate accelerating the process of breeding cultivars with multiple substantial resistance traits (Figure 1). Meanwhile, the approaches associated with gene silencing and genome editing have been established and are well developed. Due to the advantages of these techniques, in combination with the newly released genome sequences of wheat and its different species of pests and pathogens, these approaches have become powerful ways to elucidate the molecular mechanisms of interactions between different attackers and cereal plants and have provided more targets for the genetic improvement of wheat cultivars with desired traits. Based on these advances, an increasing number of heterozygous wheat genotypes have been developed by introducing desirable traits and reducing undesirable traits through conventional hybird breeding and advanced genetically modifying breeding. Although some of these resistant wheat cultivars have been able to adversely affect the performance of fungal diseases and herbivorous insects, there have still been some obstacles that have severely limited the large-scale practical applications of resistant cultivars in agriculture.
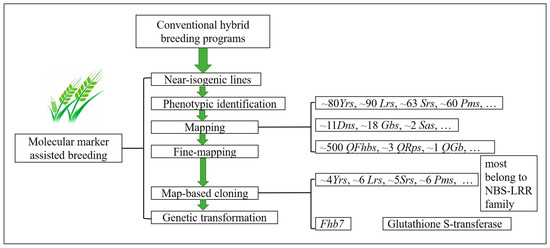
Figure 1.
The regular flow charts of molecular marker-assisted breeding programs involved in wheat cultivar improvements against biotic stresses. The near-isogenic lines developed from conventional hybrid breeding programs were employed for identification and mapping the resistance genes by adopting the diverse molecular markers, and many dominant genes and QTLs had been mapped on wheat chromosomes. Following that, many candidate genes or QTL allenes had been isolated from the wheat germplasms or its donor species with map-based cloning. Most wheat resistance genes belong to the NBS-LRR family, with the Fhb7 gene encoding a protein with Glutathione S-transferase. Yrs, stripe rust resistance genes; Lrs, leaf rust resistance genes; Sr, stem rust resistance genes; Pms, powdery mildew resistance genes; Dns, Russian wheat aphid Diuraphis noxia resistance genes; Gbs, greenbug Schizaphis graminum resistance genes; Sas, English grain aphid Sitobion avenae resistance genes; QFhbs, scab resistance QTLs; QRps, bird cherry-oat aphid Rhopalosiphum padi resistance QTLs; QGb, greenbug resistance QTL.
In this review, we summarize and discuss the recent advances and challenges related to breeding measures for improving common wheat cultivars with pest resistance and disease tolerance.
2. Challenges in Transgenic-Based Wheat Cultivar Improvements Forward to Biotic Stress Resistance
Compared with conventional breeding, transgenic-based wheat cultivar improvements could introduce the desirable traits into wheat plants in a short time. These genetically modified (GM) wheat cultivars developed by adopting recombinant DNA technology exhibit not only improved resistance to different biotic and abiotic stresses but also improved nutrient composition, such as amino acid composition and yield, relative to the requirements for farmed animals and produce. During the early post-genomics era, it had demonstrated that large-scale cultivation of GM wheat plants is a reliable and environmentally friendly approach to combat crop diseases and herbivorous insects in agricultural production.
The systems of genetic transformation and plant regeneration from apoplasts are the rate-limiting steps in the whole process of transgenic wheat breeding, which significantly affects the process of developing the available commercial use of transgenic wheat cultivars. During transformation, Agrobacterium tumefaciens-mediated transformation (ATMT) and particle bombardment mediated transformation (PBMT) are methods used frequently to transfer foreign genes into the cereal genome [7]. In comparison with PBMT, ATMT has significant advantages for successful transformation, such as preciseness of integration, high transformation efficiency, less damage to host cells, and ease of manipulation in vitro [8,9,10,11]. However, the initial successes of ATMT were limited to dicotyledonous plants, probably because dicotyledonous plants are the natural hosts for A. tumefaciens [8,12]. Until the early 1990s, significant progress was made in the ATMT of cereal crops, including rice, maize, and wheat, when phenolics such as acetosyringone (AS) were added to cocultivation media [13], which is necessary for the expression of vir gene clusters and the subsequent activation of the release, transfer, and integration of T-DNA carrying the sequence of genes of interest into the genome of host plants. To date, transgenic wheat disease resistance studies have focused more on powdery mildew and wheat scab. For instance, the gene encoding thaumatin-like protein derived from rice and wheat was integrated into wheat cultivars with the PBMT technique, resulting in resistance to wheat scab and powdery mildew, respectively [13]. Similarly, expression of barley class II chitinase in disease-susceptible wheat cultivar Bobwhite plants transformed with the PBMT technique was able to strengthen the resistance to wheat scab and powdery mildew [14,15], probably because chitinase can break the structural bonds between C1 and C4 of two consecutive chitin N-acetylglucosamines, one of the main components of the fungal cell wall [14]. In addition, the development of transgenic wheat plants expressing the β-1,3-glucanase gene derived from tobacco with ATMT revealed resistance to powdery mildew, likely because β-1,3-glucanase can trigger the hypersensitive response to pathogen infection and break the structural components of fungal cell walls [11,16]. Although there were some efforts focused on the development of transgenic wheat resistant to different cereal viral diseases up until the early 2000s, the accumulating experimental evidence revealed that it was not a trivial task [17], probably because most of reported viral disease resistance genes or alleles did not confer tolerance to all serotypes of viruses, which are transmitted by different species of herbivorous insects. Thus, building host plant resistance associated with transmitted vector pests could be the most efficient and cost-effective measure to suppress the epidemics of crop disease triggered by plant viruses.
For the development of transgenic insect-resistant crops, genes encoding toxins derived from Bacillus thuringiensis (Bt) have been widely transferred into the genome of many monocotyledons to allow them to withstand the damage caused by larval herbivory of major coleopteran or lepidopteran pests. For instance, the integration of the cryIA(c) gene into the winter wheat cultivar Yangmai 158 with ATMT led to successful expression of the Bt endotoxin protein and significantly decreased the survival, development, and fecundity of oriental armyworm [18]. In addition, over the past few decades, considerable progress has been made in the genetic expression of plant lectins in different cereal and vegetable crops. For instance, the pta gene encoding Pinellia ternate agglutinin was successfully engineered into wheat cultivars to strengthen their resistance to wheat aphid, and the performance of aphids was adversely affected when they colonized transgenic plants [18]. Similarly, snowdrop lectin (Galanthus nivalis agglutinin; GNA) has been successfully introduced into the elite wheat cultivar Bobwhite with PBMT, resulting in plants with a higher level of resistance against grain aphids, probably because plant lectins can inhibit the assimilation of sugars in the gut of herbivores [19]. In addition, considerable efforts have focused on the integration of alternative candidate genes into elite crop cultivars to strengthen their resistance to insect pests, including the genes encoding some other plant defense proteins, such as α-amylase inhibitors, secondary metabolites involved in plant defense, and even insecticidal proteins, including avidin, Photorhabdus luminescens toxins, and cholesterol oxidase [20]. Meanwhile, diverse foreign genes associated with the desirable traits of abiotic stress tolerance together with yield and quality were integrated into elite wheat cultivars with ATMT and PBMT approaches, and the cultivars carrying these foreign genes exhibited high yield and quality and exhibited tolerance to different abiotic stresses.
Although the agricultural use of transgenic resistant wheat plants with wide-ranging and durable resistance to biotic stresses could by achieved in increased crop productivity, wheat transgenic plants still face a multitude of challenges, such as low transformation efficiency, pest resistance to or tolerance of foreign genes, trigger the healthy issues and ecological risks, and challenges in the implementation of political policies (Figure 2). This probably restricts the large-scale applications of transgenic resistant wheat cultivars in agricultural production worldwide.
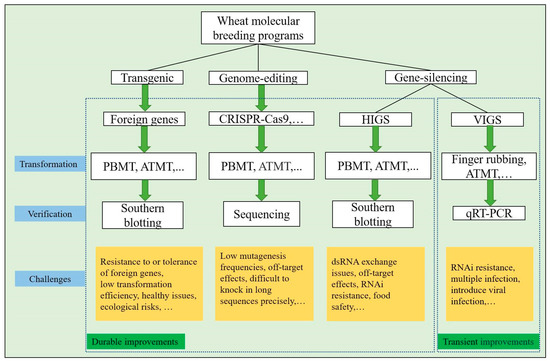
Figure 2.
The principal stages and challenges of prevalent wheat germplasm improvement strategies of molecular breeding programs. The box on the left denotes the durable wheat improvement approaches and principal stages, while the box on the right denotes the transient wheat improvements. The pathways supported by experimental evidence from the literature. ATMT, Agrobacterium tumefaciens-mediated transformation; PBMT, particle bombardment mediated transformation; HIGS, host-induced gene silencing; VIGS, virus-induced gene silencing; qRT–PCR, real-time polymerase chain reaction.
3. Molecular Markers Applied in Selecting Resistance Genes and Their Use in Cultivar Improvements
The growing desire for sustainable strategies in agricultural production has led to a need to quickly transfer the wheat-inherent genes associated with pest and disease resistance to develop additional resistant wheat varieties, especially the considerable progress made in marker-assisted selection (MAS), which has allowed us to identify and subsequently apply the wheat-inherent resistance genotypes (Figure 1). MAS involves the quick identification of traits of interest from germplasms or cross-pollination lines with the assistance of molecular markers [21]. Molecular markers can aid in the detection of variations in genomic or mitochondrial DNA with specifically designed markers on chromosomes, and these fragments with variations can be distinguished by high-resolution gel electrophoresis, such as with nondenaturing polyacrylamide gels [21,22]. Once an agronomic trait of interest is established as being tightly linked with a marker locus, it can be used for the precise selection of agronomic traits of interest in plant breeding, especially for the identification of oligogenic and qualitative traits [21]. During recent decades, various DNA marker-based methods, such as those related to restriction fragment length polymorphism (RFLP), random amplification of polymorphic DNA (RAPD), amplified fragment length polymorphisms (AFLPs), simple sequence repeats (SSRs), and sequence characterized amplified regions (SCAR), have been applied to investigate the genetics and assess the resistance traits in different wheat genotypes [21]. In recent decades, considerable progress has been made in the identification and mapping of dominant resistance genes associated with rust diseases and powdery mildew by adopting different molecular markers, and a variety of reviews have illustrated the identified molecular markers linked to major fungal disease resistance genes and their donors and the adopted approaches. For instance, more than 80 stripe rust resistance genes (Yr1–Yr80), more than 63 wheat stem rust resistance (Sr) genes, more than 90 leaf rust resistance (Lr) genes, and more than 60 genes/alleles associated with powdery mildew (Pm) resistance have been described or cataloged [23,24,25,26]. In addition, more than 30 wheat-dominant genes were reported to confer resistance against cereal aphids, including Diuraphis noxia (Kurdjumov), Schizaphis graminam (Rondani), and Sitobion avenae (Fabricius) [17,27,28,29,30]. To date, the number of resistance genes identified and assigned to different biotic stresses has continually increased (Figure 1). Although these molecular markers, especially PCR-based markers, can be used to identify the dominant genes, the genetic distance between the closely linked molecular markers and the exact locus of resistance genes is too high. This is probably because most molecular markers were developed from chromosomal regions with repetitive DNA, low gene density, or low efficiency of recombination, which significantly affected the effectiveness of fine mapping the genes and the application of molecular breeding to strengthen resistance to different pests [24].
In addition, most traits of interest are controlled by numerous genes or QTLs, and their performance is easily affected by environmental factors, which cannot be effectively identified either by phenotype selection or PCR-based methods [31,32]. Recent advances in high-throughput nucleotide sequencing (HTS) have enabled the efficient discovery of whole-genome sequence variants, which significantly facilitate efficient QTL identification and diversity analysis and the development of ultrahigh-density genetic maps [33]. For instance, the candidate QTLs associated with cereal aphid resistance in wheat recombinant inbred lines from the cross of Seri M82 wheat (susceptible) with the synthetic hexaploid wheat CWI76364 (resistant) were characterized with high-quality sequences obtained by the Illumina HiSeq2000 platform, including QRp.slu.4BL, which conferred antibiosis resistance against the bird cherry-oat aphid (Rhopalosiphum padi L.), and QRp.slu.5AL and QRp.slu.5BL, which conferred tolerance to R. padi. Moreover, a QGb.slu.2DL associated with greenbug (Schizaphis graminum Rondani) resistance was identified in the same wheat recombinant inbred lines [33]. It has been demonstrated that most of the scab resistance in wheat germplasms are controlled by QTL alleles, and over 500 QTLs have been identified and are involved in scab resistance germplasms and their derived lines [34,35]. Some of these QTLs have been applied to develop scab resistance genotypes in elite cereal cultivar backgrounds. However, the current available wheat varieties with resistance to different biotic stresses remain relatively rare, which might be because the wheat germplasm pools lack sufficient QTL for resistance to develop resistant varieties. Therefore, continuing efforts should be focused on constructing high-throughput, high-density, cost-effective, flexible, high-commercialization, and easily manipulatable platforms involved in discovering QTLs associated with biotic stress resistance from available wheat germplasm pools, which will significantly accelerate the process of marker-assisted wheat resistance breeding.
Single-nucleotide polymorphisms (SNPs) are another kind of sequencing-based molecular marker that are the most abundant markers of genetic variation and diversity. Since the late last century, it has been widely used to identify relevant genes associated with qualitative traits and quantitative traits at the single-nucleotide level, which can be present in both coding and noncoding regions [22]. In recent decades, a multitude of SNP tags have been identified by DNA sequencing in combination with fast detection methods such as diversity array technology (DArT) and restriction-site associated DNA sequencing (RAD-seq), and some of these SNPs are closely linked with quantitative traits [36]. For instance, as described previously, SNPs are important and highly accurate biomarkers of variations in desirable traits in cereal crops, such as for high yield and grain quality and for wide-ranging and durable resistance to abiotic and biotic stresses [24,37]. Although the recent reduction in costs of HTS enables the accurate discovery of SNPs, accumulating experimental evidence has demonstrated that constructing high-density SNP arrays could be an alternative that substantially describes the genetic basis of complex traits and identifies potentially associated genes through SNPs. For instance, diverse high-throughput SNP genotyping arrays developed with the aid of HTS, including wheat 9K, 15K, 35K, 55K, 90K, 820K, 921K, and 660K SNP arrays, have been broadly applied to the discovery of wheat SNP markers linked to target genes or QTL [38]. On this basis, genome-wide association studies (GWASs) were developed to comprehensively elucidate the intricate relationships between phenotype and genotype [32,39]. GWASs are based on the determination of the associations of SNPs from a large population of genotypes to identify candidate gene variants; thus, this method has a sufficient frequency of detection of genetic variations associated with both quantitative and qualitative traits [32]. Therefore, developments in GWAS analysis would provide a roadmap for the discovery of potentially new genetic variations of candidate genes associated with pest and disease resistance from diverse germplasm collections. Meanwhile, from the early part of the last century, a multitude of wheat cultivars or germplasms (or genotypes) were characterized as resistant to different species of attackers [27]. These historical collections of resistant genotypes provide sufficient genotypes for identifying genetic variations and phenotypic and genotypic traits through GWAS analysis.
Following tagging and mapping of markers that are closely linked to desirable traits associated with pest and disease resistance, it is possible to introduce them into wheat resistance breeding programs. Multiple gene pyramiding is the most important implementation of the MAS breeding strategy for constructing multiple new traits from diverse germplasms into an elite cultivar background. It can significantly facilitate the process of breeding cultivars with durable resistance to distinct diseases and pests, and sometimes it can be effective against more than one species of biotic stress in agroecosystems. The stacking genotypes were expected to result in stronger resistance to pests and diseases; however, most of them revealed less-than-additive effects [34,40,41]. For instance, the phenotype of the progeny resulting from the stacking of two or three scab-resistance QTL derived from resistant wheat cultivars Sumai 3 and Frontana exhibited less-than-additive effects when compared with the progeny carrying the individual beneficial QTL alleles [34]. Moreover, the properties of the wheat genome and the unstable performance of molecular markers or QTL have greatly impeded the application of molecular marker-assisted breeding for the sustainable control of insect pests and disease. Therefore, prior to conducting a gene pyramiding wheat breeding program, a comprehensive understanding of the relationship between different beneficial genes or QTL alleles is needed.
4. Map-Based Cloning of Resistance Genes and Its Application in Wheat Improvements
To facilitate pyramiding with other resistance genes to develop wheat cultivars with durable resistance, it is better to clone the identified resistance genes or QTL alleles associated with desirable traits [21,42]. Despite the numbers of biotic pest resistance genes identified and deployed in wheat breeding programs, only a small number of these genes have been cloned and characterized (Figure 1). However, the molecular markers linked to resistance loci contribute to map-based cloning of identified genes (Figure 1). Map-based cloning is one of the major strategies for cloning resistance genes or QTL alleles and is mostly dependent on fine genetic and physical maps [43]. For instance, for cloning the leaf rust resistance gene Lr1, the first step was to construct a high-density genetic map of the Lr1 locus by the combination of the different molecular markers and then align these markers to a genomic BAC library of hexaploid wheat and sequence and assemble the full sequence of the resistance gene [44]. This gene was cloned from hexaploid wheat and its ancestor A. tauschii by adopting a similar approach [45]. In recent decades, considerable progress has been made in the fine mapping and cloning of phytopathogen resistance genes.
Based on these advances in the fine mapping of resistance genes in the wheat genome, the stripe rust resistance genes Yr10, Yr18, Yr36, and Yr46 and the leaf rust resistance genes Lr10, Lr21, Lr22a, Lr23, Lr34, and Lr42 were cloned from wheat cultivars by adopting comparative genomic approaches derived from map-based cloning, and the stem rust resistance genes Sr13, Sr33, Sr35, Sr50, and Sr57 were cloned from wheat and its related species with similar methods [46,47]. In addition, more than eight Pm resistance genes have been cloned from common wheat or its donor species, and six of them, Pm3b, Pm21, Pm38, Pm39, Pm46, and Pm60, have been cloned with map-based cloning [42]. For instance, Pm60, encoding a protein with a nucleotide-binding site (NBS) and leucine-rich repeat (LRR) domain, has been cloned from its donor species T. urartu [26]. This gene has been introgressed into common wheat varieties to strengthen its resistance to wheat powdery mildew [48]. Based on the current available references, most plant resistance genes (R genes) that have been cloned over the last two decades belong to the NBS-LRR family, probably because their encoded proteins have a NBS and LRR domain, which are exclusively associated with plant disease resistance [49]. To fully utilize these cloned R genes to breed wheat cultivars that carry disease resistance, extensive genome-wide studies have focused on characterizing the structure and potential functions of NBS-LRR genes isolated from different species of plants. This result suggested that during the process of plant immunity, the LRR domain of the NBS-LRR protein could directly interact with the effectors released by pathogens in their infection or monitor the status of host proteins targeted by pathogen effectors, resulting in the specific recognition of the invading pathogens. The recognition of invading pathogens by the LRR domain was shown to lead to a conformational shift of the NBS domain, which then enabled the N-terminal domains to trigger downstream hypersensitive reactions and caused systemic acquired resistance in infected plant tissues to suppress the spread and proliferation of pathogens [50]. In addition, diverse pests can secrete salivary effectors during infestation, and some effectors are recognized by the LRR domain of the NBS-LRR protein, triggering pest resistance in host plants [51,52,53]. For instance, NBS-LRR genes have been individually isolated from the wild tomato Lycopersicon peruvianum (L.) P. Mill., and muskmelon Cucumis melo (L.), by map-based cloning; their encoding proteins might confer resistance to the potato aphid Macrosiphum euphorbiae (Thomas) and cotton aphid Aphis gossypii (Glover) [54,55,56]. This gene has been widely introduced into different species of crops to strengthen their resistance to different pests and nematodes [56]. As described previously, a multitude of wheat cultivars present NBS-LRR genes [45]; thus, these results suggested that NBS-LRR genes could exhibit resistance to wheat aphids and other pests. Therefore, NBS-LRR genes provide strategic targets for improving the performance of wheat cultivars to strengthen their pest and disease resistance by adopting approaches associated with molecular breeding in the future.
More recently, a Fusarium resistance gene named Fhb7 was cloned from chromosome 7E (7EL, an important donor associated with resistance to fungi or viruses) of Thinopyrum elongatum through a map-based cloning approach, while this gene might encode a glutathione S-transferase [57]. It could be an important donor for breeding elite wheat cultivars with resistance to FHB and crown rot. In contrast to the developments in disease resistance gene cloning, only a small amount of insect resistant genes have been fine mapped and isolated from wheat germplasms in recent decades. Moreover, for rapid development of aphid tolerant wheat cultivars, the differential-display reverse transcription-PCR method was applied to identify candidate expressed sequence tags (ESTs) associated with pest resistance from populations of near-isogenic wheat lines. For instance, six potential ESTs ranging from 128 to 532 bp in length were identified from the populations of backcrossed near-isogenic lines derived from aphid-tolerant wheat genotypes XN98-10-35 and susceptible genotype XN1376 [58,59]. Experimental evidence shows that the biological and morphological traits of XN98-10-35 enable this winter wheat line to convert additional photoassimilates to compensate for the damage caused by aphid feeding and strengthen the cell wall for tolerance to aphid infestation [59]. For cloning the full-length sequences of these ESTs, chromosome walking and rapid amplification of cDNA ends (RACE) techniques could be considered an ideal method for determining the unknown sequences of DNA and mRNA based on known sequences or the markers closest to candidate genes, respectively. Prior to HTS, chromosome walking is an effective tool for the development of EST-STS (sequence-tagged sites) markers to construct a high-resolution genetic map. For instance, the Lr10 and Lr21 resistance locus, which is closely linked to EST-STS markers at a genetic distance of 2.0 centimorgan (cM), has been established in hexaploid wheat by adopting chromosome walking [43,60]. In addition, the constitutive leaf rust resistance gene TcS2A2 has been cloned from the wheat line TcLr35 by adopting the RACE approach, and its derived protein products also have the typical NBS-LRR domains [61]. However, common (hexaploid) wheat is a natural hybrid of Triticum urartu (L.), Aegilops speltoides (Tausch), and A. tauschii (Coss.), and it has an extremely large genome size (16,000 Mbp), highly repetitive sequences, and proximity to the centromere region pose severe challenges for the map-based cloning of resistance genes in hexaploid wheat and its application in wheat improvements [62].
5. Gene Silencing-Based Cultivar Improvements against for Biotic Stresses
Currently, molecular biological studies of common wheat are entering the postgenomics era, and silencing of genes of interest is therefore a crucial method for understanding the potential functions of endogenous genes already present in wheat. During the last few decades, tremendous progress has been made in the genetic manipulation of wheat by the silencing of its endogenous genes, which not only helps to understand the function of many novel genes but also contributes to the generation of improved varieties with economically important traits [63]. RNA silencing, commonly known as RNA interference (RNAi), is a highly conserved phenomenon and is widely found in many eukaryotes, including plants, animals, insects, nematodes, and fungi [64,65]. During coevolution, the invading attackers develop RNA silencing machinery to suppress their hosts’ immune responses. RNAi is always triggered by double-stranded RNA (dsRNA) to silence the expression of target genes. The dsRNA released from the invading attackers can be perceived by the ribonuclease III enzyme Dicer or Dicer-like enzymes and processed for degradation into small interfering RNAs (siRNAs) of 20–30 nucleotides. The binding of siRNA to Argonaute family proteins (AGOs) results in recruitment to the RNA-induced silencing complex (RISC) and suppression of the transcription of target genes associated with defense responses in host plants [66]. Since its discovery, RNAi has been widely used as a novel potential approach for temporarily strengthening the resistance of the crop host to withstand diseases and attacks by herbivorous insects [65,67]. Direct feeding of dsRNA molecules targeting critical genes of attackers via artificial spraying has led to successful knockdown of the expression of target genes and suppressed their performance in many monocot- and dicot-attacker systems [68]. For instance, foliar application with sprayable dsRNA products of the three fungal genes encoding cytochrome P450 lanosterol C-14α-demethylases in F. graminearum onto barley seedlings successfully silenced the transcription of target genes and inhibited fungal growth [69]. However, similar to chemical spraying, to improve the control efficiency, dsRNA molecules should be sprayed multiple times during agricultural production. Meanwhile, the widespread and frequent application of sprayable dsRNA products targeting the crucial genes of pathogens or pests on the foliar of seedlings would significantly stimulate attackers to evolve RNAi resistance and increase the difficulty of pest control. Thus, the specific production of dsRNA derived from critical genes of different biotic stresses in transgenic plants may be a promising measure for future crop protection.
To increase dsRNA delivery efficiency, virus-induced gene silencing (VIGS) and host-induced gene silencing (HIGS) methods have been developed (Figure 2). The VIGS approach employs a recombinant virus carrying RNA sequences homologous to the host genes, and viral infection could trigger the production of dsRNA intermediates into host tissue during viral replication, resulting in RNA silencing and suppression of the corresponding genes of the host plant [66,70,71]. For instance, recombinant barley stripe mosaic virus (BSMV) strains carrying antisense sequences of wheat phytoene desaturase (TaPds) and auxin response factor 2-like protein (TaArf2-like) have been employed to suppress the corresponding transcription and significantly reduced Fusarium head blight (FHB) disease symptoms (infection with the fungus F. culmorum) in the spring wheat cultivar Apogee [72]. Although VIGS could be a potential approach for silencing the transcription of crucial genes associated with defense responses in host plants, this approach will introduce viral infection and result in new viral diseases. Moreover, VIGS does not modify the genetic composition of host plants, and the VIGS approach is broadly applied to transiently silence target genes to determine their potential biological roles [73]. For instance, silencing the transcription of the gene Tel7E01T1020600.1 (probably representing the Fhb7 gene) in wheat leaves through the VIGS approach confirmed its potential functions associated with resistance to F. graminearum infections [57]. These results suggested that the VIGS approach is not a suitable strategy for stable and consistent improvements of wheat cultivars by silencing the interested traits associated with the wheat endogenous genes (Figure 2), while the HIGS methods could overcome the previously mentioned drawbacks referred to in the VIGS approach, and it could be considered the ideal method for stable and consistent improvements of the resistance of elite wheat cultivars to diverse biotic stresses.
The term HIGS was proposed by Nowara et al. (2010) when they employed BSMV carrying the antisense RNA of 1,3-β-glucanosyltransferase (BgGTF1 and BgGTF2) of the powdery mildew fungus Blumeria graminis (D.C. Speer) f. sp. hordei to determine whether silencing the obligate biotrophic genes associated with fungal virulence adversely affected fungal development and haustorium formation [71]. Therefore, the HIGS approach was developed from VIGS, which involves the production of silencing-inducing RNA molecules targeting the crucial genes of pathogens or pests in plants with transgenic integration or viral infection approaches [65,74]. When sufficient dsRNA molecules are delivered into the tissue of pathogens or pests, they trigger the silencing of RNA transcription of the corresponding genes and have adverse effects on the virulence or performance of pathogens or pests [66]. In recent decades, the HIGS approach has been used to knock down the transcription of critical pathogen or pest genes in numerous systems comprising pathogens or herbivorous pests and their host wheat plants, including chewing insect–wheat, sap-sucking insect–wheat, biotrophic fungus–wheat, hemibiotrophic fungus–wheat, and virus–wheat systems, resulting in the disruption of the effects of pathogens or herbivorous pests on transgenic wheat plants [65,75]. The genes conferring vegetative growth and virulence to fungi are considered promising targets for HIGS-mediated crop protection approaches. For instance, recombinant BSMV vectors carrying antisense sequences of virulence genes, transcription factors or effector proteins of Puccinia striiformis f. sp. tritici (Pst), including calcineurin homolog and its regulatory subunit (PsCNA1, PsCNB1, PsCNB), mitogen-activated protein kinase (PsMAPK1), cyclophilin (PsCYC1), protein kinase A catalytic subunit (PsCPK1), and glycine-serine-rich effector (PstGSRE1), have been employed to suppress the corresponding transcription and reduce disease symptoms [72,74,76,77]. In addition, dsRNAs targeting the secreted lipase (Fgl1), mitogen-activated protein (MAP) kinase (Fmk1), and β-1,3-glucan synthase (Gls1) of F. culmorum were produced in the wheat cultivar Apogee with a similar method and severely affected the vegetative growth and virulence of F. culmorum and then reduced FHB disease symptoms [72]. To develop durable and consistent resistance to these fungi, RNAi constructs targeting the effector gene Avra10 of B. graminis were transferred into barley cultivar Pallas plants (without the Mla10 resistance gene) with PBMT and significantly reduced haustorium formation [71]. More recently, HIGS-mediated simultaneous silencing of the three virulence effector genes of B. graminis was established in the wheat cultivar Chancellor with PBMT, resulting in reduced fungal virulence [78]. Meanwhile, transgenic wheat plants expressing dsRNA targeting the crucial genes of F. graminearum significantly affected the development and virulence of fungi, and the transgenic seedlings exhibited stable and consistent resistance to FHB [79].
In addition, accumulating experimental evidence has revealed that transgene-mediated HIGS silencing of the important genes associated with the performance of aphids in wheat plants is a potential tool for controlling the associated damage. For instance, transgene-mediated HIGS silencing of the detoxifizyme gene carboxylesterase E4 (CbE E4) of S. avenae in wheat cultivar Liaochun 10 severely impaired herbivore tolerance to phoxim chemicals [80]. In addition, when S. avenae was fed on transgenic cereal plants, the expression of silencing-inducing RNA molecules with HIGS, such as those targeting the structural sheath protein (shp), effector zinc finger protein (SaZFP), chitin synthase 1 (CHS1), or lipase maturation factor 2-like genes, had severe adverse effects on the performance of aphids [81,82,83,84]. Furthermore, rapid advances in genomic or RNA-sequencing and analysis tools have facilitated the expression of dsRNAs targeting more genes involved in the growth, survival, or development of different biotic stresses in transgenic wheat plants.
Taken together, transient or constitutive cultivar improvement mediated by gene silencing is an effective strategy to combat fungal diseases and attacks by herbivorous insects [66]. However, there are still significant challenges associated with using gene silencing for plant protection that need to be addressed before practical application of this method in agriculture, such as target selection, the stability of dsRNA, dsRNA exchange issues, off-target effects, RNAi resistance, and food safety (Figure 2).
6. Genome Editing Involved in Crop Breeding Forward to Biotic Stress Resistance
As described previously, virus- or transgene-mediated gene silencing has emerged in recent decades as a promising approach to understand the potential roles of interesting genes, including biotic stress resistance or susceptible genes (Figure 2). Recent developments in wheat genome editing have enabled the precise modification of gene sequences to fully understand the potential function, molecular mechanisms, and regulation of candidate genes and even to develop valuable new traits in crops [85]. For instance, mutation of all three MLO (TaMLO-A1, TaMLO-B1, and TaMLO-D1) alleles conferred susceptibility to powdery mildew with a transcription activator-like effector nuclease (TALEN)-mediated editing system in bread wheat, and the plants exhibited strong resistance to powdery mildew [86]. Among these available sequence-specific editing tools, the clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated endonuclease 9 (Cas9, derived from Streptococcus pyogenes) system has gained wide acceptance for genome editing, where multiple genes can be directly targeted by single guide RNA (sgRNA) through nucleotide base pairing instead of DNA-binding proteins [87]. Since it emerged in 2012, the CRISPR/Cas9 system has received widespread attention and has been developed for a broad range of applications [88]. The CRISPR/Cas9-mediated genome editing system generally involves two steps: the specifically designed sgRNA directs the Cas9 endonuclease to produce double-strand breaks (DSBs) in the target locus of the genome, and cellular repair mechanisms, including homology-directed repair (HDR) and nonhomologous end-joining (NHEJ), are employed to repair these DSBs [87,89,90]. These repair mechanisms can generate insertions, deletions, or knock-in substitutions in the target locus, which may interrupt, eliminate, or correct the defects in genomes [87,88]. For instance, genome editing through HDR of CRISPR/Cas9 could repair DSBs with the desired DNA fragment, while the NHEJ-mediated cellular DNA repair process often randomly introduces deletions or insertions (InDels) [85,91]. In addition, Shan et al. (2014) described a stepwise protocol for CRISPR/Cas9 system-based editing of the rice and wheat genomes, which significantly facilitated more accurate targeting in mutagenesis, characterization of gene function, and finely tuned regulation to improve target traits in crops [92]. Due to its high specificity, simplicity, flexibility, and versatility for modifying endogenous genes, in the last decade, this system has been widely used for predictable and precise genome modification in cereal and horticultural crops, such as rice, maize, wheat, barley, sorghum, soybean, tomato, potato, and cotton [85,93,94]. It has been shown that the CRISPR/Cas9 editing system currently focuses more on target traits related to yield, quality, and tolerance to either biotic or abiotic stresses in different crops. For instance, the TaMLO-A1 allele was mutated with the CRISPR/Cas9 system, and the mutation frequency was similar to that obtained using the TALEN system [86]. More recently, three homoeoalleles of the transcription factor TaWRKY19 were mutated with the CRISPR/Cas9 system, resulting in transcriptional repression of the TaNOX10 gene and enhancing host plant ROS accumulation; thereafter, the mutated wheat seedlings exhibited strong resistance to Pst [95]. Therefore, a multiple gene editing-based system could be a novel promising tool to improve varieties with desirable traits in hexaploid wheat. This conclusion is supported by the experimental evidence obtained from other crops. For instance, six important loci associated with fruit number, size, shape, nutrient content, and plant architecture were simultaneously edited by the CRISPR/Cas9 system via a single transformation in wild tomato (Solanum pimpinellifolium), resulting in notably increased fruit size, fruit number, and lycopene levels in comparison to the wild tomato species [96].
Although the CRISPR/Cas9 editing system could provide potential opportunities to improve wheat crops, wheat genome editing is challenging (Figure 2). First, common wheat is a hexaploidy crop with high repetitive sequence content and three homologous chromosomes, which severely influence editing efficiency. The mutagenesis frequencies obtained using the CRISPR/Cas9 editing system in wheat showed a mutation rate typically less than 10%, which is lower than that in other crops [94,97]. Recent advances made by combining the alternative wheat ubiquitin promoter TaU3 with different editing systems, such as CRISPR/Cpf1 (derived from Prevotella and Francisella 1) and CRISPR/xCas9 (variant of SpCas9), to edit the economically important endogenous genes of wheat significantly increased the mutagenesis frequencies compared with the CRISPR/Cas9 editing system [97]. Meanwhile, either application of the TaU3 promoter or the Cestrum yellow leaf curling virus (CmYLCV) promoter into the CRISPR/Cas9 editing system could allow efficient editing of multiple genes in wheat [98]. Second, similar to transgenic breeding, the frequency of genome editing depends most on the system of genetic transformation and plant regeneration [94]. Currently, the PBMT and ATMT are two main approaches for transporting sgRNA into wheat callus cells during genome editing (Figure 2); thus, they are the major obstacles to gene editing processes [92,97]. Third, as described previously, both gene silencing and editing are subject to off-target effects [94]. Although the development of the CRISPR/Cas9 editing system has significantly reduced off-target effects, it is better for the accurate prediction of potential off-target sites. Recently, Cram et al. (2019) designed a web-based RNA bioinformatics portal (wheatCRISPR) for the design of highly specific sgRNAs and increased the efficiency of gene editing in the wheat genome [99]. Moreover, based on the current available literature, genome editing in most plants is performed by the CRISPR/Cas9-mediated NHEJ repair system. Although some studies have reported that CRISPR/Cas9-mediated HDR could repair DSBs with small precise changes (such as single-base substitution or few-base insertions) in a multitude of plant species, it is difficult to knock in long sequences precisely. The major challenge is the simultaneous delivery of sgRNA and the HDR donor carrying long homologous arms flanking each side and the desired sequences [94]. Therefore, new developments in sgRNA length manipulation would be very important for precisely changing DSBs with desired sequences.
As described previously, the advances obtained with GWASs and SNPs associated with desired traits, such as resistance to biotic and abiotic stress, provide a multitude of targets for genome editing. However, the CRISPR/Cas9 editing system is proficient in stochastic disruption of genes rather than point mutation or correction in the target locus, and the HDR-mediated cellular DNA repair process remains inefficient (typically ~0.1 to 5%). The SNP-CRISPR system established by Liu’s group could facilitate the development of new crop cultivars from wild species by adopting the CRISPR/Cas9 system [100]. These advances provide the potential to effectively improve current wheat cultivars with desirable agronomic traits through point mutations or corrections involved in genome editing strategies.
Overall, although the challenges associated with genome editing will remain in the following years, substantial efforts will continue to be focused on solving these problems. For instance, more promoters and Cas nucleases for use in the CRISPR/Cas toolbox and CRISPR epigenetic engineering tools could be engineered, and the optimization of sgRNA delivery strategies could improve the mutagenesis frequencies of these toolboxes [101]. Thus, CRISPR/Cas9 system-mediated genome editing or cultivar improvement would be the most promising methods for wheat molecular breeding in the future.
7. Concluding Remarks and Future Perspectives
The present review illustrates and summarizes the concrete advances achieved by basic research and new technologies in crop improvement in recent decades. As described previously, either crosspollination or genetic engineering approaches will be employed to develop novel wheat cultivars by introducing desirable traits and reducing undesirable traits. In particular, the achievements in methodologies and bioinformatics tools related to HTS, multiomics analysis, genome editing, mining of genetic diversity, and gene silencing have provided broad insights into crop improvement [102]. The combined use of these measures would greatly facilitate success in understanding the potential functions and regulation of interesting genes, thereby accelerating the process of breeding superior wheat cultivars with desirable traits, such as high yield, good flavor, and high nutrient levels in cereal grains, and tolerance to biotic and abiotic stresses [103]. For instance, the combined use of VIGS and CRISPR/Cas9 approaches targeting the wheat endogenous genes associated with being susceptible to biotic stresses would be an effective strategy for developing wheat resistant cultivars [104]. However, there are several obstacles that severely limit wheat improvement programs. First, the efficiencies of either silencing or editing in wheat improvement are still insufficient for developing elite wheat cultivars with desired traits because wheat has the largest genome. Fortunately, the rapidly developing toolbox for genome editing and next-generation sequencing technology will significantly improve the performance of these approaches and accelerate the process of wheat molecular breeding [101]. Second, most of the available resistant wheat cultivars are effective against one species of pathogen or herbivorous insect but not against others [105,106]. Adverse conditions, such as those created by global warming and extreme weather, further aggravate the damage to cereal crops by increasing the populations of various pests and the incidence of pandemics. Moreover, diverse attackers are more likely to colonize different or similar niches of the same plant to obtain nutrients, probably because their feeding or infestation could positively induce biological and physiological alterations [107]. The crosstalk between phytohormone signaling pathways induced by multiple pests and pathogens would fine-tune the plant defense responses and further aggravate the challenges of attackers [108]. Therefore, pyramiding breeding with more resistance genes associated with different attackers in elite cultivars will be an essential approach for breeding superior cultivars that can withstand attacks from multiple attackers [29]. Third, sustainable development in agriculture raises important questions for adopting the available wheat genotypes to develop novel wheat cultivars. The germplasm is fundamental for breeding programs, and substantial efforts have been made to identify molecular markers, including PCR-based markers and SNP or InDel tags. With the assistance of HTS, an increasing number of molecular markers will be discovered, which will provide targets for cultivar improvement. Notably, integrating multiomics analysis and genome editing into cultivar improvements could be an environmentally friendly and economical approach for wheat breeding and will allow the demands for sustainable development in agriculture to be fully met.
In summary, many questions regarding gene silencing and editing associated with wheat molecular breeding remain open (Figure 2). However, advances and high-value collaborative efforts in these fields will significantly improve mutagenesis frequencies and overcome the drawbacks of these approaches, such as off-target effects and RNAi resistance. Thus, the technical barriers of these approaches will be overcome further to facilitate a wide range of applications for breeding novel cultivars and obtaining cultivars with desired traits for agricultural production.
Author Contributions
K.L.: Conceptualization, Visualization, Writing—review & editing, Supervision, Funding acquisition. D.H.: Writing—original draft, Investigation, Data curation. J.G.: Writing—original draft, Data curation. G.L.: Writing—review & editing. B.L.: Visualization. X.C.: Data curation. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China, grant number 32260717, Natural Science Foundation of Shaanxi Province, China, grant number 2021JQ-619, and the Research Fund for the Doctoral Start-up Foundation of Yan’an University, grant number YDBK2019-65.
Data Availability Statement
Not applicable.
Acknowledgments
We apologize to our colleagues whose work could not be included because of space constraints. We would like to thank the reviewers for their critical comments on earlier versions of this manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Bassi, F.M.; Bentley, A.R.; Charmet, G.; Ortiz, R.; Crossa, J. Breeding schemes for the implementation of genomic selection in wheat (Triticum spp.). Plant Sci. 2016, 242, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Abdul Fiyaz, R.; Ajay, B.C.; Ramya, K.T.; Aravind Kumar, J.; Sundaram, R.M.; Subba Rao, L.V. Speed Breeding: Methods and Applications. In Accelerated Plant Breeding, Volume 1: Cereal Crops; Gosal, S.S., Wani, S.H., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 31–49. [Google Scholar] [CrossRef]
- De Zutter, N.; Audenaert, K.; Ameye, M.; De Boevre, M.; De Saeger, S.; Haesaert, G.; Smagghe, G. The plant response induced in wheat ears by a combined attack of Sitobion avenae aphids and Fusarium graminearum boosts fungal infection and deoxynivalenol production. Mol. Plant Pathol. 2017, 18, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Drakulic, J.; Caulfield, J.; Woodcock, C.; Jones, S.P.T.; Linforth, R.; Bruce, T.J.A.; Ray, R.V. Sharing a host plant (wheat [Triticum aestivum]) increases the fitness of Fusarium graminearum and the severity of Fusarium head blight but reduces the fitness of grain aphids (Sitobion avenae). Appl. Environ. Microbiol. 2015, 81, 3492–3501. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.P.; Paul, V.L.; Slater, R.; Warren, A.; Denholm, I.; Field, L.M.; Williamson, M.S. A mutation (L1014F) in the voltage-gated sodium channel of the grain aphid, Sitobion avenae, is associated with resistance to pyrethroid insecticides. Pest. Manag. Sci. 2014, 70, 1249–1253. [Google Scholar] [CrossRef]
- Jones, H.D. Transformation and Transgene Expression. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Brian, T., Murray, B.G., Murphy, D.J., Eds.; Academic Press: Oxford, UK, 2017; pp. 248–254. [Google Scholar] [CrossRef]
- Dai, S.; Zheng, P.; Marmey, P.; Zhang, S.; Tian, W.; Chen, S.; Beachy, R.N.; Fauquet, C. Comparative analysis of transgenic rice plants obtained by Agrobacterium-mediated transformation and particle bombardment. Mol. Breed. 2001, 7, 25–33. [Google Scholar] [CrossRef]
- Hu, T.; Metz, S.; Chay, C.; Zhou, H.P.; Biest, N.; Chen, G.; Cheng, M.; Feng, X.; Radionenko, M.; Lu, F.; et al. Agrobacterium-mediated large-scale transformation of wheat (Triticum aestivum L.) using glyphosate selection. Plant Cell Rep. 2003, 21, 1010–1019. [Google Scholar] [CrossRef]
- Wu, H.; Sparks, C.; Amoah, B.; Jones, H.D. Factors influencing successful Agrobacterium-mediated genetic transformation of wheat. Plant Cell Rep. 2003, 21, 659–668. [Google Scholar] [CrossRef]
- Zhao, T.; Zhao, S.; Chen, H.; Zhao, Q.; Hu, Z.; Hou, B.; Xia, G. Transgenic wheat progeny resistant to powdery mildew generated by Agrobacterium inoculum to the basal portion of wheat seedling. Plant Cell Rep. 2006, 25, 1199–1204. [Google Scholar] [CrossRef]
- Weir, B.; Gu, X.; Wang, M.; Upadhyaya, N.; Elliott, A.R.; Brettell, R.I.S. Agrobacterium tumefaciens-mediated transformation of wheat using suspension cells as a model system and green fluorescent protein as a visual marker. Funct. Plant Biol. 2001, 28, 807–818. [Google Scholar] [CrossRef]
- Chen, W.P.; Chen, P.D.; Liu, D.J.; Kynast, R.; Friebe, B.; Velazhahan, R.; Muthukrishnan, S.; Gill, B.S. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor. Appl. Genet 1999, 99, 755–760. [Google Scholar] [CrossRef]
- Shin, S.; Mackintosh, C.A.; Lewis, J.; Heinen, S.J.; Radmer, L.; Dill-Macky, R.; Baldridge, G.D.; Zeyen, R.J.; Muehlbauer, G.J. Transgenic wheat expressing a barley class II chitinase gene has enhanced resistance against Fusarium graminearum. J. Exp. Bot. 2008, 59, 2371–2378. [Google Scholar] [CrossRef]
- Bliffeld, M.; Mundy, J.; Potrykus, I.; Fütterer, J. Genetic engineering of wheat for increased resistance to powdery mildew disease. Theor. Appl. Genet. 1999, 98, 1079–1086. [Google Scholar] [CrossRef]
- Ward, E.R.; George, B.P.; Moyer, M.B.; Williams, S.C.; Dincher, S.S.; Kevin, C.S.; James, J.B.; Hope, T.T.; Ahl-Goy, P.; Meins, F.; et al. Differential regulation of β-1,3-glucanase messenger RNAs in response to pathogen infection. Plant Physiol. 1991, 96, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Aradottir, G.I.; Crespo-Herrera, L. Host plant resistance in wheat to barley yellow dwarf viruses and their aphid vectors: A review. Curr. Opin. Insect Sci. 2021, 45, 59–68. [Google Scholar] [CrossRef]
- Yu, Y.; Wei, Z. Increased oriental armyworm and aphid resistance in transgenic wheat stably expressing Bacillus thuringiensis (Bt) endotoxin and Pinellia ternate agglutinin (PTA). Plant Cell Tiss. Org. 2008, 94, 33–44. [Google Scholar] [CrossRef]
- Michiels, K.; Van Damme, E.J.; Smagghe, G. Plant-insect interactions: What can we learn from plant lectins? Arch. Insect Biochem. Physiol. 2010, 73, 193–212. [Google Scholar] [CrossRef]
- Christou, P.; Capell, T.; Kohli, A.; Gatehouse, J.A.; Gatehouse, A.M.R. Recent developments and future prospects in insect pest control in transgenic crops. Trends Plant Sci. 2006, 11, 302–308. [Google Scholar] [CrossRef]
- Nayak, S.N.; Singh, V.K.; Varshney, R.K. Marker-Assisted Selection. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Thomas, B., Murray, B.G., Murphy, D.J., Eds.; Academic Press: Oxford, UK, 2017; pp. 183–197. [Google Scholar] [CrossRef]
- Gupta, P.K.; Roy, J.K.; Prasad, M. Single nucleotide polymorphisms: A new paradigm for molecular marker technology and DNA polymorphism detection with emphasis on their use in plants. Curr. Sci. 2001, 80, 524–535. [Google Scholar]
- Kassa, M.T.; You, F.M.; Fetch, T.G.; Fobert, P.; Sharpe, A.; Pozniak, C.J.; Menzies, J.G.; Jordan, M.C.; Humphreys, G.; Zhu, T.; et al. Genetic mapping of SrCad and SNP marker development for marker-assisted selection of Ug99 stem rust resistance in wheat. Theor. Appl. Genet. 2016, 129, 1373–1382. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, Q.; Wang, Q.; Liu, S.; Yu, S.; Mu, J.; Huang, S.; Sela, H.; Distelfeld, A.; Huang, L.; et al. SNP-based pool genotyping and haplotype analysis accelerate fine-mapping of the wheat genomic region containing stripe rust resistance gene Yr26. Theor. Appl. Genet. 2018, 131, 1481–1496. [Google Scholar] [CrossRef]
- Singla, J.; Lüthi, L.; Wicker, T.; Bansal, U.; Krattinger, S.G.; Keller, B. Characterization of Lr75: A partial, broad-spectrum leaf rust resistance gene in wheat. Theor. Appl. Genet. 2017, 130, 1–12. [Google Scholar] [CrossRef]
- Zou, S.; Wang, H.; Li, Y.; Kong, Z.; Tang, D. The NB-LRR gene Pm60 confers powdery mildew resistance in wheat. New Phytol. 2018, 218, 298–309. [Google Scholar] [CrossRef]
- Liu, X.L.; Yang, X.F.; Wang, C.Y.; Wang, Y.J.; Zhang, H.; Ji, W.Q. Molecular mapping of resistance gene to English grain aphid (Sitobion avenae F.) in Triticum durum wheat line C273. Theor. Appl. Genet. 2012, 124, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Luo, K.; Wang, L.; Zhao, H.; Zhang, G. Molecular mapping of resistance gene to the English grain aphid, Sitobion avenae, in a Chinese wheat line XN98-10-35. Mol. Breed. 2015, 35, 203. [Google Scholar] [CrossRef]
- Luo, K.; Zhao, H.; Wang, X.; Kang, Z. Prevalent Pest Management Strategies for Grain Aphids: Opportunities and Challenges. Front. Plant Sci. 2022, 12, 3252. [Google Scholar] [CrossRef]
- Lu, H.; Rudd, J.C.; Burd, J.D.; Weng, Y. Molecular mapping of greenbug resistance genes Gb2 and Gb6 in T1AL.1RS wheat-rye translocations. Plant Breeding 2010, 129, 472–476. [Google Scholar] [CrossRef]
- Bernardo, R. Molecular markers and selection for complex traits in plants: Learning from the last 20 years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef]
- Young, P.R.; Vivier, M.A. 10—Genetics and genomic approaches to improve grape quality for winemaking. In Managing Wine Quality; Reynolds, A.G., Ed.; Woodhead Publishing: Sawston, UK, 2010; pp. 316–364. [Google Scholar] [CrossRef]
- Crespo-Herrera, L.A.; Akhunov, E.; Garkava-Gustavsson, L.; Jordan, K.W.; Smith, C.M.; Singh, R.P.; Åhman, I. Mapping resistance to the bird cherry-oat aphid and the greenbug in wheat using sequence-based genotyping. Theor. Appl. Genet. 2014, 127, 1963–1973. [Google Scholar] [CrossRef]
- Miedaner, T.; Wilde, F.; Steiner, B.; Buerstmayr, H.; Korzun, V.; Ebmeyer, E. Stacking quantitative trait loci (QTL) for Fusarium head blight resistance from non-adapted sources in an European elite spring wheat background and assessing their effects on deoxynivalenol (DON) content and disease severity. Theor. Appl. Genet. 2006, 112, 562–569. [Google Scholar] [CrossRef]
- Xue, S.; Li, G.; Jia, H.; Lin, F.; Cao, Y.; Xu, F.; Tang, M.; Wang, Y.; Wu, X.; Zhang, Z.; et al. Marker-assisted development and evaluation of near-isogenic lines for scab resistance QTLs of wheat. Mol. Breed. 2010, 25, 397–405. [Google Scholar] [CrossRef]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotechnol. J. 2020, 18, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Joukhadar, R.; El-Bouhssini, M.; Jighly, A.; Ogbonnaya, F.C. Genome-wide association mapping for five major pest resistances in wheat. Mol. Breed. 2013, 32, 943–960. [Google Scholar] [CrossRef]
- Eshed, Y.; Zamir, D. Less-than-additive epistatic interactions of quantitative trait loci in tomato. Genetics 1996, 143, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Marcel, T.C.; Aghnoum, R.; Durand, J.; Varshney, R.K.; Niks, R.E. Dissection of the Barley 2L1.0 Region Carrying the ‘Laevigatum’ Quantitative Resistance Gene to Leaf Rust Using Near-Isogenic Lines (NIL) and subNIL. Mol. Plant Microbe. Interact. 2007, 20, 1604–1615. [Google Scholar] [CrossRef]
- Wan, W.; Xiao, J.; Li, M.; Tang, X.; Wen, M.; Cheruiyot, A.K.; Li, Y.; Wang, H.; Wang, X. Fine mapping of wheat powdery mildew resistance gene Pm6 using 2B/2G homoeologous recombinants induced by the ph1b mutant. Theor. Appl. Genet. 2020, 133, 1265–1275. [Google Scholar] [CrossRef]
- Huang, L.; Brooks, S.A.; Li, W.; Fellers, J.P.; Trick, H.N.; Gill, B.S. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics 2003, 164, 655–664. [Google Scholar] [CrossRef]
- Cloutier, S.; McCallum, B.D.; Loutre, C.; Banks, T.W.; Wicker, T.; Feuillet, C.; Keller, B.; Jordan, M.C. Leaf rust resistance gene Lr1, isolated from bread wheat (Triticum aestivum L.) is a member of the large psr567 gene family. Plant Mol. Biol. 2007, 65, 93–106. [Google Scholar] [CrossRef]
- Ling, H.Q.; Zhu, Y.; Keller, B. High-resolution mapping of the leaf rust disease resistance gene Lr1 in wheat and characterization of BAC clones from the Lr1 locus. Theor. Appl. Genet 2003, 106, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, D.; Zeng, Q.; Duan, Y.; Yuan, F.; Shi, J.; Wang, Q.; Wu, J.; Huang, L.; Kang, Z.; et al. Fine mapping of wheat stripe rust resistance gene Yr26 based on collinearity of wheat with Brachypodium distachyon and rice. PLoS ONE 2013, 8, e57885. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, L.; Holdgate, S.; James, L.; Thomas, J.; Mackay, I.J.; Cockram, J. The evolving battle between yellow rust and wheat: Implications for global food security. Theor. Appl. Genet. 2022, 135, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, Y.; Li, Y.; Fahima, T.; Shen, Q.; Xie, C. Introgression of the powdery mildew resistance genes Pm60 and Pm60b from Triticum urartu to common wheat using Durum as a ‘Bridge’. Pathogens 2021, 11, 25. [Google Scholar] [CrossRef]
- Lukasik, E.; Takken, F.L. STANDing strong, resistance proteins instigators of plant defence. Curr. Opin. Plant Biol. 2009, 12, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Xue, J.; Wang, Q.; Wang, B.; Chen, J. Revisiting the origin of plant NBS-LRR genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Elzinga, D.A.; Jander, G. The role of protein effectors in plant-aphid interactions. Curr. Opin. Plant Biol. 2013, 16, 451–456. [Google Scholar] [CrossRef]
- Jaouannet, M.; Rodriguez, P.A.; Thorpe, P.; Lenoir, C.J.G.; MacLeod, R.; Escudero-Martinez, C.; Bos, J.I.B. Plant immunity in plant-aphid interactions. Front. Plant Sci. 2014, 5, 663. [Google Scholar] [CrossRef]
- Xu, H.; Qian, L.; Wang, X.; Shao, R.; Hong, Y.; Liu, S.; Wang, X. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef]
- Rossi, M.; Goggin, F.L.; Milligan, S.B.; Kaloshian, I.; Ullman, D.E.; Williamson, V.M. The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc. Natl. Acad. Sci. USA 1998, 95, 9750–9754. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.M.; Chuang, W. Plant resistance to aphid feeding: Behavioral, physiological, genetic and molecular cues regulate aphid host selection and feeding. Pest. Manag. Sci. 2014, 70, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Zhang, G.; Wang, C.; Ouellet, T.; Wu, J.; Zhu, Q.; Zhao, H. Candidate genes expressed in tolerant common wheat with resistant to English grain aphid. J. Econ. Entomol. 2014, 107, 1977–1984. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Yao, X.; Luo, C.; Hu, X.; Wang, C.; Wang, Y.; Hu, Z.; Zhang, G.; Zhao, H. Biological and morphological features associated with English grain aphid and bird cherry-oat aphid tolerance in winter wheat line XN98-10-35. J. Plant Growth Regul. 2019, 38, 46–54. [Google Scholar] [CrossRef]
- Stein, N.; Feuillet, C.; Wicker, T.; Schlagenhauf, E.; Keller, B. Subgenome chromosome walking in wheat: A 450-kb physical contig in Triticum monococcum L. spans the Lr10 resistance locus in hexaploid wheat (Triticum aestivum L.). Proc. Natl. Acad. Sci. USA 2000, 97, 13436–13441. [Google Scholar] [CrossRef]
- Wang, H.Y.; Liu, D.Q.; Yang, W.X. A wheat disease resistance gene analog of the NBS-LRR class: Identification and analysis. J. Plant Dis. Protect. 2011, 118, 63–68. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Borisjuk, N.; Kishchenko, O.; Eliby, S.; Schramm, C.; Anderson, P.; Jatayev, S.; Kurishbayev, A.; Shavrukov, Y.; Kirti, P.B. Genetic modification for wheat improvement: From transgenesis to genome editing. Biomed. Res. Int. 2019, 2019, 6216304. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, R.; Matzke, M.; Matzke, A.J.M. Plants, RNAi, and the Nobel Prize. Science 2006, 314, 1242–1243. [Google Scholar] [CrossRef]
- Qi, T.; Guo, J.; Peng, H.; Liu, P.; Kang, Z.; Guo, J. Host-induced gene silencing: A powerful strategy to control diseases of wheat and barley. Int. J. Mol. Sci. 2019, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Geng, S.; Li, A.; Mao, Y.; Mao, L. RNAi technology for plant protection and its application in wheat. aBIOTECH 2021, 2, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ang, L.; Weilin, Z. Current understanding of the molecular players involved in resistance to rice planthoppers. Pest. Manag. Sci. 2019, 75, 2566–2574. [Google Scholar] [CrossRef]
- Chung, S.H.; Jing, X.; Luo, Y.; Douglas, A.E. Targeting symbiosis-related insect genes by RNAi in the pea aphid-Buchnera symbiosis. Insect Biochem. Mol. Biol. 2018, 95, 55–63. [Google Scholar] [CrossRef]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Nowara, D.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.X.; Kumlehn, J.; Schweizer, P. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef]
- Chen, W.; Kastner, C.; Nowara, D.; Oliveira-Garcia, E.; Rutten, T.; Zhao, Y.; Deising, H.B.; Kumlehn, J.; Schweizer, P. Host-induced silencing of Fusarium culmorum genes protects wheat from infection. J. Exp. Bot. 2016, 67, 4979–4991. [Google Scholar] [CrossRef]
- Benedito, V.A.; Visser, P.B.; Angenent, G.C.; Krens, F.A. The potential of virus-induced gene silencing for speeding up functional characterization of plant genes. Genet. Mol. Res. 2004, 3, 323–341. [Google Scholar]
- Qi, T.; Zhu, X.; Tan, C.; Liu, P.; Guo, J.; Kang, Z.; Guo, J. Host-induced gene silencing of an important pathogenicity factor PsCPK1 in Puccinia striiformis f. sp. tritici enhances resistance of wheat to stripe rust. Plant Biotechnol. J. 2018, 16, 797–807. [Google Scholar] [CrossRef]
- Koch, A.; Wassenegger, M. Host-induced gene silencing-mechanisms and applications. New Phytol. 2021, 231, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Panwar, V.; McCallum, B.; Bakkeren, G. Host-induced gene silencing of wheat leaf rust fungus Puccinia triticina pathogenicity genes mediated by the Barley stripe mosaic virus. Plant Mol. Biol. 2013, 81, 595–608. [Google Scholar] [CrossRef]
- Yin, C.; Jurgenson, J.E.; Hulbert, S.H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Mol. Plant Microbe Interact. 2010, 24, 554–561. [Google Scholar] [CrossRef]
- Schaefer, L.K.; Parlange, F.; Buchmann, G.; Jung, E.; Wehrli, A.; Herren, G.; Müller, M.C.; Stehlin, J.; Schmid, R.; Wicker, T.; et al. Cross-kingdom RNAi of pathogen effectors leads to quantitative adult plant resistance in wheat. Front. Plant Sci. 2020, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Song, X.; Li, H.; Cao, L.; Sun, K.; Qiu, X.; Xu, Y.; Yang, P.; Huang, T.; Zhang, J.; et al. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Duan, X.; Lv, Y.; Zhang, X.; Nie, Z.; Xie, C.; Ni, Z.; Liang, R. Silencing of an aphid carboxylesterase gene by use of plant-mediated RNAi impairs Sitobion avenae tolerance of phoxim insecticides. Transgenic Res. 2014, 23, 389–396. [Google Scholar] [CrossRef]
- Abdellatef, E.; Will, T.; Koch, A.; Imani, J.; Vilcinskas, A.; Kogel, K. Silencing the expression of the salivary sheath protein causes transgenerational feeding suppression in the aphid Sitobion avenae. Plant Biotechnol. J. 2015, 13, 849–857. [Google Scholar] [CrossRef]
- Zhao, Y.; Sui, X.; Xu, L.; Liu, G.; Lu, L.; You, M.; Xie, C.; Li, B.; Ni, Z.; Liang, R. Plant-mediated RNAi of grain aphid CHS1 gene confers common wheat resistance against aphids. Pest. Manag. Sci. 2018, 74, 2754–2760. [Google Scholar] [CrossRef]
- Xu, L.; Hou, Q.; Zhao, Y.; Lu, L.; Li, B.; Ni, Z.; Liang, R. Silencing of a lipase maturation factor 2-like gene by wheat-mediated RNAi reduces the survivability and reproductive capacity of the grain aphid, Sitobion avenae. Arch. Insect Biochem. 2017, 95, e21392. [Google Scholar] [CrossRef]
- Sun, Y.; Sparks, C.; Jones, H.; Riley, M.; Francis, F.; Du, W.; Xia, L. Silencing an essential gene involved in infestation and digestion in grain aphid through plant-mediated RNA interference generates aphid-resistant wheat plants. Plant Biotechnol. J. 2019, 17, 852–854. [Google Scholar] [CrossRef]
- Kim, D.; Alptekin, B.; Budak, H. CRISPR/Cas9 genome editing in wheat. Funct. Integr. Genomics 2018, 18, 31–41. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Schaart, J.G.; van de Wiel, C.C.M.; Smulders, M.J.M. Genome editing of polyploid crops: Prospects, achievements and bottlenecks. Transgenic Res. 2021, 30, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, D.; Ramirez-Solis, R.; Garza-Elizondo, M.A.; de Lourdes Garza-Rodriguez, M.D.L.; Barrera-Saldana, H.A. Genome editing: A perspective on the application of CRISPR/Cas9 to study human diseases (Review). Int. J. Mol. Med. 2019, 43, 1559–1574. [Google Scholar] [CrossRef] [PubMed]
- Symington, L.S.; Gautier, J. Double-strand break end resection and repair pathway choice. Annu. Rev. Genet. 2011, 45, 247–271. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Schaefer, K.A.; Wu, W.; Colgan, D.F.; Tsang, S.H.; Bassuk, A.G.; Mahajan, V.B. Unexpected mutations after CRISPR-Cas9 editing in vivo. Nat. Methods 2017, 14, 547–548. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Gao, C. Genome editing in rice and wheat using the CRISPR/Cas system. Nat. Protoc. 2014, 9, 2395–2410. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; He, G.; Ma, L.; Deng, X.W. CRISPR/Cas9-mediated disruption of TaNP1 genes results in complete male sterility in bread wheat. J. Genet. Genom. 2020, 47, 263–272. [Google Scholar] [CrossRef]
- Song, G.; Jia, M.; Chen, K.; Kong, X.; Khattak, B.; Xie, C.; Li, A.; Mao, L. CRISPR/Cas9: A powerful tool for crop genome editing. Crop J. 2016, 4, 75–82. [Google Scholar] [CrossRef]
- Wang, N.; Fan, X.; He, M.; Hu, Z.; Tang, C.; Zhang, S.; Lin, D.; Gan, P.; Wang, J.; Huang, X.; et al. Transcriptional repression of TaNOX10 by TaWRKY19 compromises ROS generation and enhances wheat susceptibility to stripe rust. Plant Cell 2022, 34, 1784–1803. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef]
- Liu, H.; Wang, K.; Jia, Z.; Gong, Q.; Lin, Z.; Du, L.; Pei, X.; Ye, X. Efficient induction of haploid plants in wheat by editing of TaMTL using an optimized Agrobacterium-mediated CRISPR system. J. Exp. Bot. 2020, 71, 1337–1349. [Google Scholar] [CrossRef]
- Li, J.; Zhang, S.; Zhang, R.; Gao, J.; Qi, Y.; Song, G.; Li, W.; Li, Y.; Li, G. Efficient multiplex genome editing by CRISPR/Cas9 in common wheat. Plant Biotechnol. J. 2021, 19, 427–429. [Google Scholar] [CrossRef]
- Cram, D.; Kulkarni, M.; Buchwaldt, M.; Rajagopalan, N.; Bhowmik, P.; Rozwadowski, K.; Parkin, I.A.P.; Sharpe, A.G.; Kagale, S. WheatCRISPR: A web-based guide RNA design tool for CRISPR/Cas9-mediated genome editing in wheat. BMC Plant Biol. 2019, 19, 474. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Lin, Q.; Jin, S.; Gao, C. The CRISPR-Cas toolbox and gene editing technologies. Mol. Cell 2022, 82, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, W.; Fernie, A.R.; Wen, W. Combining novel technologies with interdisciplinary basic research to enhance horticultural crops. Plant J. 2022, 109, 35–46. [Google Scholar] [CrossRef]
- Chen, J.; Hu, X.; Shi, T.; Yin, H.; Sun, D.; Hao, Y.; Xia, X.; Luo, J.; Fernie, A.R.; He, Z.; et al. Metabolite-based genome-wide association study enables dissection of the flavonoid decoration pathway of wheat kernels. Plant Biotechnol. J. 2020, 18, 1722–1735. [Google Scholar] [CrossRef]
- Brauer, E.K.; Balcerzak, M.; Rocheleau, H.; Leung, W.; Schernthaner, J.; Subramaniam, R.; Ouellet, T. Genome editing of a deoxynivalenol-induced transcription factor confers resistance to Fusarium graminearum in wheat. Mol. Plant Microbe Interact. 2020, 33, 553–560. [Google Scholar] [CrossRef]
- Zhu, L.C.; Smith, C.M.; Fritz, A.; Boyko, E.; Voothuluru, P.; Gill, B.S. Inheritance and molecular mapping of new greenbug resistance genes in wheat germplasms derived from Aegilops tauschii. Theor. Appl. Genet. 2005, 111, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, S.; Dedryver, C.A. Reduced BYDV-PAV transmission by the grain aphid in a Triticum monococcum line. Eur. J. Plant Pathol. 2009, 123, 281–289. [Google Scholar] [CrossRef]
- Ni, X.; Quisenberry, S.S. Diuraphis noxia and Rhopalosiphum padi (Hemiptera: Aphididae) interactions and their injury on resistant and susceptible cereal seedlings. J. Econ. Entomol. 2006, 99, 551–558. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Luo, K.; Ouellet, T.; Zhao, H.; Wang, X.; Kang, Z. Wheat–Fusarium graminearum Interactions Under Sitobion avenae Influence: From Nutrients and Hormone Signals. Front. Nutr. 2021, 8, 703293. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).