Abstract
Although nitrogen (N) is the most limiting nutrient for agricultural production, its overuse is associated with environmental pollution, increased concentration of greenhouse gases, and several human and animal health implications. These implications are greatly affected by biochemical transformations and losses of N such as volatilization, leaching, runoff, and denitrification. Half of the globally produced N fertilizers are used to grow three major cereals—rice, wheat, and maize—and their current level of N recovery is approximately 30–50%. The continuously increasing application of N fertilizers, despite lower recovery of cereals, can further intensify the environmental and health implications of leftover N. To address these implications, the improvement in N use efficiency (NUE) by adopting efficient agronomic practices and modern breeding and biotechnological tools for developing N efficient cultivars requires immediate attention. Conventional and marker-assisted selection methods can be used to map quantitative trait loci, and their introgression in elite germplasm leads to the creation of cultivars with better NUE. Moreover, gene-editing technology gives the opportunity to develop high-yielding cultivars with improved N utilization capacity. The most reliable and cheap methods include agronomic practices such as site-specific N management, enhanced use efficiency fertilizers, resource conservation practices, precision farming, and nano-fertilizers that can help farmers to reduce the environmental losses of N from the soil–plant system, thus improving NUE. Our review illuminates insights into recent advances in local and scientific soil and crop management technologies, along with conventional and modern breeding technologies on how to increase NUE that can help reduce linked N pollution and health implications.
1. Introduction
The future food demand requires an increase of about 60% in agricultural production to accomplish the great challenge of feeding the burgeoning human population, which is predicted increase from 7.7 billion in 2019 to 9.7 billion by 2050 [1]. Among the essential nutrients of plants, N primarily affects the physiological, biochemical, and molecular responses, metabolism, and resource allocation of the plant and is thus considered the most limiting nutrient for crop production [2,3,4,5,6]. The synthetic N-fertilizer is a crucial input for the agricultural sector, accounting for almost half of the N sources for global food production, and thus, it is critical for fulfilling people’s food security, especially in developing countries. However, intensive and imbalanced use of N fertilizers is linked with losses of N from soil–plant systems, accounting for low NUE, higher cost of investment, and greater food insecurity [7]. This lost N leads to several environmental consequences such as eutrophication, increased GHG emission, and pollution of surface and underground water resources [8,9]. The consumption of nitrate-enriched water is linked with human health issues such as methemoglobinemia and cancer. Moreover, the higher nitrate content in runoff water might affect animals, which can cause direct poisoning in aquatic organisms and livestock [10].
Due to their significant contribution to the supply of proteins and calories (directly through grains and indirectly through livestock products) [11,12,13,14,15], cereal crops play a crucial role in the nutrition and food security of humans and animals, especially in developing countries. Among them, maize, rice, and wheat account for almost 90% of cereal production worldwide and are considered most imperative from the human nutrition point of view [16,17,18,19]. Around half of the world’s population is directly or indirectly dependent on N fertilizers for food supply [20]. Due to the extended role of synthetic N fertilizers in agriculture, nearly 120 Tg of reactive N is produced in industries annually [21]. Almost 50% is used to fertilize three dominant cereal crops, viz., maize, wheat, and rice (Figure 1). However, less than 40% of the applied N is taken up by the crops. At the same time, the leftover part enters the environment through various mechanisms, causing a wide range of agronomic, environmental, and health implications [22]. Thus, looking at N losses’ agronomic, environmental, and human health implications, some innovations are needed to improve NUE, especially in cereals. In this regard, the adoption/use of improved agronomic approaches such as site-specific nutrient management (SSNM), resource conservation practices, precision farming, and enhanced use of efficient fertilizers (EUEFs) and nano-fertilizers can help the farmers to reduce the environmental losses of N from the soil–plant system, thus improving NUE at the farm level [23].
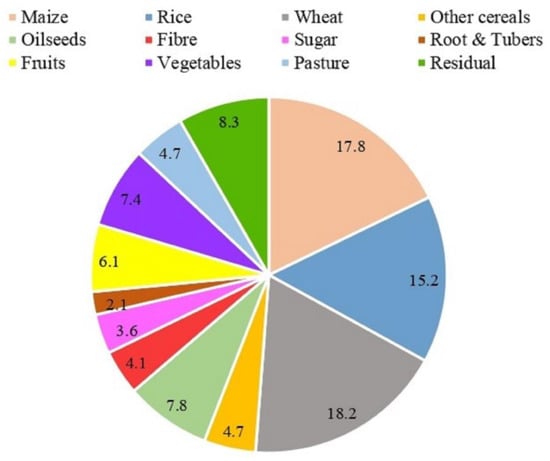
Figure 1.
Crop-wise N fertilizer consumption (%).
Further, conventional and molecular breeding with the aid of different biotechnological tools such as transgenics can also add to developing highly N-efficient cultivars [24]. However, the failure to achieve the expected success in developing N-efficient genotypes in cereals can be attributed to the lack of coordinated efforts by plant breeders, biotechnologists, and physiologists. Therefore, to increase success in creating N-efficient cultivars, it is imperative to apply comprehensive multidisciplinary strategies, including breeding, physiological, agronomical, and biotechnological approaches. This review is an attempt to provide insight into the N cycling in the biosphere and its various loss pathways from the soil–plant systems and associated consequences of N losses. We also discuss the crop management strategies for the management of N in soil and conventional and non-conventional breeding technologies to enhance NUE of cereals that can help reduce the linked N pollution in water bodies and health implications.
2. Nitrogen Route in Soil–Plant Systems
Biogeochemical cycling of N includes various pools, forms, and environmental processes of N in terrestrial and aquatic ecosystems. Several forms of N, such as N2, N2O, NO−, NO2−, NO3−, and NH3 (reduced form), are present in the soil and atmosphere (biosphere). The N cycling encompasses nitrification, denitrification, mineralization, immobilization, volatilization, precipitation, and fixation processes for the movement of N from one compartment to another. The N cycle is further divided into two cycles that include external and internal cycles. The external cycle comprises processes of N contribution in ecosystems such as N2 fixation, NH4+ and NO3− in rainwater, and the use of N fertilizers as organic and mineral forms, which can cause N loss from cropping systems, i.e., denitrification, NO3− leaching, and NH4+ volatilization.
On the contrary, the internal cycle includes processes of N transformation (conversion of the form) or assimilation into the plant body, root turnover, mineralization, and microbial immobilization [25]. N externally applied to soil is managed by plants in two ways, i.e., uptake (as nitrate and ammonium) of N by the plant and assimilation (amino acids) of N in the plant to form essential amino acids and other metabolites [26]. Available N released by applied fertilizer or organic N of soil is highly sensitive to its losses by the soil–plant system as denitrification, leaching, runoff, and volatilization. Therefore, N optimization is of great concern for economic and ecological reasons [27]. The N cycle is started by the fixation of atmospheric N (inert) and the generation of bio-available N (reactive) compounds, i.e., nitrate, ammonium, and other oxides. This cycle is anthropogenically induced by fertilizer input and fossil fuel burning. In agricultural practice, nitrate primarily arises from fertilizers, cattle manure, crop residues, and soil organic matter. Heterotrophic micro-organisms play a key role in converting organic substrates initially to ammonium and finally to nitrite by nitrifying bacteria. In native ecosystems, N comes from lightening, biological fixation, and atmospheric deposition (dry and wet deposition) [28,29,30,31]. Denitrification is the microbial process that eliminates nitrate and reduces it to nitrite, nitric oxide, and nitrous oxide under anaerobic conditions. The addition, removal, and succeeding reentry of N between different compartments of the soil and atmosphere to complete the N cycle are depicted in Figure 2 [32]. The quantity of inorganic N in the soil can be summarized by the equation given below of the N balance, which is dependent on the N cycle [33,34,35,36].
where P represents N deposition through precipitation, B represents biologically fixed N, F represents fertilizer N, U represents N from urine and manure, M represents N from mineralization, PL represents plant N uptake, G represents N gaseous losses, I represents N immobilization, L represents N losses through leaching and, E represents N losses through erosion and surface runoff.
N balance (N) = NP + NB + NF + NU + NM − NPL − NG − NI − NL − NE

Figure 2.
Global N cycle in relation to the soil–plant system.
3. Global N Consumption and Status of NUE
Global fertilizer demand reached 200.92 million metric tonnes (MT) in 2022, out of which more than 111.59 MT was shared by N alone, which was predicted by the Food and Agriculture Organization of the United Nations, Rome, 2019 [37] (Figure 3). Further FAO (2019) predicted that global total ammonia production capacity would reach 190.39 million tonnes in 2022, out of which the total supply might be 163.22 million tonnes. Globally, the most N-consuming countries are China, India, and the USA accounting for a 53.16% share, and four other countries, namely Brazil, Indonesia, Pakistan, and Canada, account for 13% of global N consumption in 2019 [37].
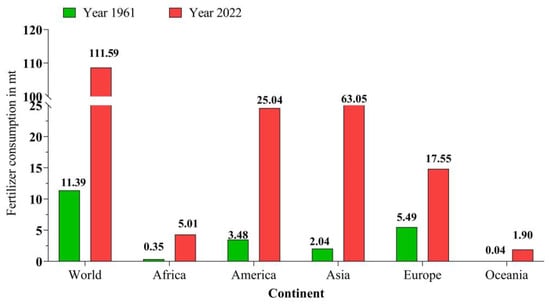
Figure 3.
Global synthetic N fertilizer consumption.
N is the sine qua non component of the present-day agriculture system as it is required in huge quantities for the plant’s growth and provides plant protein. These essential proteins (enzymes) convert light energy into carbohydrates, which is the primary energy source. More than 100 Tg N year−1 of reactive N is produced industrially by the Haber–Bosch process using fossil fuels as energy sources. Of this, half of the total production of N fertilizer is applied to three main cereals, i.e., wheat (18.2%), followed by maize (17.8%), and rice (15.2%), which deliver the bulk of human food calories and proteins (Figure 1). Other than cereal, the global application of N fertilizer to fruits and vegetables (13.5%), oil crops (7.8%), sugar (3.6%), fiber (4.1%), roots and tubers (2.1%), grassland (4.7%), and other agricultural and non-agricultural plant species (8.3%) (IFA 2017) (Figure 1).
Without the application of N fertilizer, food production would not be sufficient to feed even 50% of the current global population. Crop production must be increased to meet the requirement of an additional population of 2 to 3 billion by 2050. A total of 1551 Tg N was harvested from cereals, which was supplied by N fertilizer (48%) and soil (4%). Approximately 737 Tg of crop N (48%), equal to 25, 29, and 38 kg ha−1 year−1 for wheat, maize and rice, respectively, is imparted by sources other than fertilizer (soil N). In this N, the major source is non-symbiotic N fixation, i.e., about 370 Tg of total crop N (24%), equal to 13 kg ha−1 year−1 for maize or wheat and 22 kg ha−1 year−1 for rice [38]. Compost and atmospheric deposition are other sources of N, contributing 217 (14%) and 96 Tg (6%), respectively. Crop residues and seeds also add slightly to the nitrogen status of the soil [38].
Estimates of fertilizer NUE are highly variable, as they depend on the experimental methods and formulae used to determine it. Variations among studies in climatic, edaphic, crop, and soil management parameters, which are addressed in-depth in the review’s final section, are primarily responsible for the wide range of values (Figure 4). Moreover, total N in the grain or aboveground biomass can be responsible for estimates of fertilizer N recovery. Fertilizer N accumulated in roots is mostly not included as a part of fertilizer N recovered by the crop due to the difficulty in measuring the remains in the soil. Additionally, the uptake of fertilizer N by the crop following the fertilized crop is rarely added in the studies of fertilizer-use efficiency. Both conditions contribute to the determination of true fertilizer NUE. A summary of 93 published studies over different locations worldwide estimates the central tendency for NUE expressions for maize, wheat, and rice (Table 1) [39]. The average agronomic efficiency of N (AEN) among the cereals ranged from 18 kg grain per kg N in maize to 24 kg grain per kg N in rice. The partial factor productivity of N (PFPN, total grain yield at a certain level of N applied) differed markedly among maize, rice, and wheat (Table 1). Similar PFPN values of 62 to 70 in maize and rice and 44 in wheat were found. The values of AEN and PPFN also varied among regions and were highest in Asia and lowest in Africa. The huge variance in PFPN indicates that corn and rice produce more output than wheat with the same amount of N. The maize (65%) and wheat (57%) showed higher apparent recovery efficiency of N (AREN) over rice (46%). Likewise, PEN ranged from 29 to 53 kg grain per kg of N, with the highest in rice (Table 1). Among the world’s regions, Asia had higher physiological efficiency of N (PEN) of 47 (kg grain per kg of N) compared with 23 to 28 kg grain per kg of N in other regions. In a recent review article, it was reported the current level of N recovery for the three major cereals (rice, wheat, and maize) is approximately 30–50% [40].

Figure 4.
Nitrogen inputs in ten major river systems in the world [41,42,43].

Table 1.
Common NUE indices and their values for N fertilizer for maize, wheat, and rice.
4. NUE Concept and Components
Generally speaking, NUE is a representation of how effectively plants utilize the mineral N that is available. Its definition is the increase in economic performance per unit of N applied, absorbed, or utilized by the plant [44]. The determination of NUE is an important approach to evaluating the fate of applied chemical fertilizers and their role in improving crop yields. Nitrogen use efficiency (NUE) is an established metric used to benchmark N management. There are numerous approaches to calculating NUE, but it is difficult to find an authoritative resource that collates the various NUE indices and systematically identifies their assets and shortcomings (Congreves et al. 2021). However, according to one definition of NUE, it is a dynamic and complex trait and has been defined in several ways in the literature, although most of them denote the ability of a system to convert inputs into outputs [45]. The detailed information on commonly used terms of NUE along with their typical uses are summarized here (Table 2)
Table 2.
Different commonly used indices of nitrogen use efficiency, their assessment using the difference method, and their basic uses.
5. Environmental N Losses
Certain species of reactive nitrogen (Nr) such as N2O, NO3−, NO2−, NH3, and NH4+ cause a cascading effect on the environment and global ecosystem services [47]. The anthropogenic activities such as fossil fuel combustion and intensive application in the agricultural sector are linked with large-scale losses of N to the environment through various routes such as soil erosion, surface runoff, leaching, denitrification, and ammonia volatilization, which are summarized in Figure 2.
5.1. Soil Erosion and Surface Runoff
N adsorbed on the soil colloids are lost by water and wind agents. In arid and semiarid regions, the N losses are caused mainly by wind erosion, while water-erosion-mediated N loss is dominant in humid and sub-humid climatic regions. Soil-dissolved N from surface-applied nitrate and adsorbed soil N can be lost after heavy rain through runoff [48]. The quantity of N lost through runoff and soil erosion depends on the duration and intensity of rainfall. These losses were also greatly affected by management practices and can be minimized with some improved agronomic control measures. Large amounts of nitrogen are removed through eroding soil, which normally removes about three times as much nitrogen as is still present in the soil. The average amount of nitrogen in a tonne of fertile surface soil is 1–6 kg, but the average amount of nitrogen in a tonne of soil on eroded land is 0.1–0.5 kg. The average yearly total nitrogen loss was 10.2 kg ha−1 year−1, according to reports [49].
5.2. Ammonia Volatilization
Ammonium volatilization refers to the conversion of NH4+-N into NH3 gas and its loss into the atmosphere. It is a significant route of N loss, especially in soils with high pH (saline and alkaline). Besides the soil reaction and type of N fertilizers, soil and atmospheric temperature and moisture status of the soil also greatly affect the quantity of the NH3 (gas) released into the atmosphere [50,51]. IFA/FAO (2001) estimated that the loss of NH3 through the volatilization process from fertilized grassland, upland crops, and fertilized rice (13, 18, and 20%, respectively) in developing countries was higher than that of developed countries (6, 8 and 3%, respectively). The comparatively lower price of fertilizer N compared with the worth of crop product lost in developing countries has also led farmers to misuse/overdose of N-fertilizer, leading to lower N use efficiencies (Figure 5). The economic implications of NH3 emission from anthropogenic activities are particularly threatening. It emits about 7.6 million kg NH3 ha−1 from the crop and livestock sector, accounting for more than 90% of the total emission [52].
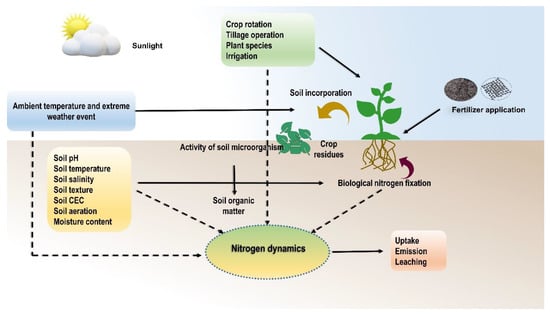
Figure 5.
Interactions between nitrogen dynamics and environmental factors.
5.3. Denitrification
Microbially mediated reduction of the nitrate form of N to a variety of gaseous forms of N (NO, N2O, and N2) under anaerobic conditions is termed denitrification [53]. Such activity is most commonly observed under a redox state where lesser O2 levels result in an increased microbial population that converts nitrate (NO3−) to nitrogen (N2) and nitrous oxide (N2O), and finally, this reduced form of N is lost to the atmosphere (Figure 6). Besides the soil moisture status, the denitrification process is greatly affected by the concentration of ammonical and nitrate N, carbon content, and soil and atmospheric temperature, texture, and drainage chrematistics of the soil [54,55]. Such N loss is responsible for 10–15% of applied N, depending on the edaphic and climatic conditions of the region. Among these aforementioned gases, N lost in the form of N2O is considered one of the most hazardous, as it is a greenhouse gas. Nitrous oxide (N2O) has an extended lifetime (120 years) and a higher potential for heat-trapping compared with CO2. For atmospheric N2O emissions, the major anthropogenic sources are agricultural activities, especially N fertilizer and manure application, livestock farming, fossil fuel combustion from fertilizer, and pesticide industries [56].

Figure 6.
Factors affecting N dynamics in soil.
5.4. Nitrate Leaching
Loss of dissolved nitrate (NO3−) with water beyond the root zone of the plants through the soil profile is referred to as leaching loss of N. The enormity of N loss through leaching is due to several factors such as agro-climatic situation, soil characteristics, management practices, and methods/types of N fertilizer used [57]. Leaching is the most commonly reported means of N loss in sandy soils with light texture due to their high percolation rates; hence, the application of the higher dose of fertilizer results in the leaching loss of NO3− to the water table, which causes water pollution [58]. Leaching also occurs from well-irrigated areas where the water table is shallow, and generally, N fertilizer is used in the form of nitrate. The wet soil due to irrigation generally has higher NO3− leaching potential and export; however, the ranges are wide in different ecologies. As a result of reduced percolation beyond the root zone, paddy cultivation under lowland settings with fine-textured soils reduces N losses by leaching. Additionally, the mobility of soil macrofauna and plant roots facilitates rapid nitrate transport in soil [59]. The leaching and runoff losses of nitrate deteriorate the quality of both ground and surface water bodies, leading to eutrophication and algal bloom, and are also linked with human health implications discussed in Section 6.2 [60]. According to earlier studies, up to 80% of applied nitrogen may be lost as NO3 runoff, depending on the type of soil [61]. Due to increased precipitation and slower plant nitrate uptake, colder temperatures significantly contribute to nitrate leaching [62]. According to earlier research on nitrate loss in soil, grazing land lost more than 100 kg N of nitrate per year after tillage through the soil profile over the following winter [62,63].
6. Environmental Consequences of N Losses
N cycling is a dynamic route in nature, and a range of biochemical transformation processes within the soil–plant system are responsible for changes in all applied and organic forms of N (Figure 2). Some of these processes (leaching, volatilization, denitrification, etc.) are responsible for N movement to the water bodies and atmosphere from soil–plant systems, consequently leading to severe environmental concerns. The major environmental consequences associated with lower NUE of fertilizer N are discussed here.
6.1. Groundwater Contamination and Nitrate Pollution
N that has gone beyond the root zone of the plant system via leaching can cause groundwater pollution. The leaching loss of N in NO3− indicates soil fertility degradation, environmental threats, and worsened public health [64]. It is a global phenomenon that occurs from various sources and causes surface and groundwater contamination, especially eutrophication, leading to the proliferation of algae blooms and biodiversity loss [65]. The utilization of drainage water from irrigated lands is the defining characteristic of nitrate leaching. This problem is especially prevalent in regions where crops with high water and nitrogen requirements are grown. It is more obvious for groundwater, where both communities and irrigation districts use subsurface water [66]. Additionally, nitrate leaching is an alarming issue for public health hazards as it creates two medical conditions, methemoglobinemia (or the ‘blue-baby syndrome’) in infants and stomach cancer in adults. According to the recommendation of the World Health Organization (WHO), drinking water should have a nitrate concentration of less than 10 ppm. Many of the world’s freshwater water resources exceed the safe limit of nitrate and were found to be unsuitable for use for drinking purposes [67].
6.2. Eutrophication
Eutrophication is the phenomenon of nutrient enrichment in water bodies due to the excessive use of fertilizers, particularly N and phosphorus. It has the presence of sufficient levels of planktonic, algae, and/or water weeds to impair the water for use in the domestic water supply, for recreation, and for fisheries (reduction in cold water fisheries and fish kills) [68]. Effluents containing higher nutrient levels from agro-fertilizer and other industries, municipal and domestic, including black water contamination and urban runoff, accelerate the eutrophication problems in many parts of the world. The consequences of eutrophication in water bodies are the reduction in submerged aquatic vegetation, oxygen, and biodiversity; the spread of some harmful algal blooms; imbalances in the food webs and bio geo-chemical cycling; and fish kills [69]. Eutrophication can lead to oxygen depletion, producing various chemicals, i.e., unionized ammonia and the formation of hydrogen sulfides that are both directly and indirectly toxic to aquatic organisms and humans. N from soil and fertilizers on agricultural land and its loss by runoff might be regarded as the leading cause of eutrophication. Estimates showed that the agricultural land of Africa and South America had a lower contribution in N flow into their rivers through their nutrient-depleted ecosystems (Figure 4). The agricultural ecosystem comes under the world’s major rivers, such as the Mississippi, Rhine, Po, and Ganges, which exhibit extreme use and higher N loss.
6.3. N Deposition
Ammonia released to the atmosphere from soil with inorganic N applied, as well as manure and other anthropogenic activities, can return to the geo surface along with SO2. The intensification of N inputs through atmospheric deposition denotes a possible threat to all ecosystems, but forests and aquatic systems are the most sensitive ecosystems [70]. Atmospheric N deposition is the third major reason behind the loss of worldwide biodiversity with rates that are more than doubled over the past century. Total microbial biomass, carbon, and respiration are lowered due to the addition of N [71]. Such negative effects are boosted with the increasing N application rate. The fungal-to-bacterial ratio, relative abundances of arbuscular mycorrhizal fungi, and Gram-negative bacteria are also reduced by N addition; however, Gram-positive bacteria are enhanced. The problem of global warming is also enhanced due to greenhouse gas (GHG) emissions mainly because of higher N deposition [72].
6.4. GHG Emission
Out of total anthropogenic emissions, the global GHG emissions from the agriculture sector account for 14–17% as a direct source (cultivation of crops, livestock, and deforestation). Further, 3–6% may be accounted for in terms of global GHG emissions if indirect sources such as production, packaging, storage, and transportation of agricultural inputs and farm products are considered for emissions [73]. Among the direct agricultural emissions, N2O, ruminant CH4, biomass burning, CH4 from rice production, and manure management account for 38, 32, 12, 11, and 7%, respectively [74]. Due to the increasing trend of using energy-intensive agricultural practices/farm mechanization, global GHGs emission from the agricultural sector is increasing rapidly [75]. Nitrous oxide (N2O) alone emitted from synthetic fertilizers, manures, and crop residues contribute more than 40% of total agricultural emissions. The emitted N2O accounts for the depletion of the ozone layer by reacting with stratospheric O2 and forming nitric acid (IPCC, 2007). N fertilizers massively contribute to the global GHG emissions among all synthetic fertilizers as they produce a large emission scale during production, transportation, storage, and consumption [76].
7. Factors Affecting N Dynamics in Soil–Plant Systems
Globally, there is a huge disparity between N demand and supply in cultivated agroecosystems [77]. N deficiency in soils limits crop growth and development; on the other hand, an excessive amount causes water pollution (eutrophication) and global warming (greenhouse gas emission) [78]. Therefore, efficient and precise management of N is crucial for crop production, profitability, and minimizing environmental losses. Generally, the NUE of crops depends on both biotic and abiotic factors, including edaphic (pH, soil moisture, and temperature), climatic (ambient temperature and rainfall), management (dose of N, application time, placement, type of fertilizer, and irrigation), and biological factors and also varies from crop to crop (varietal differences, genotypes, root development, and rootstock) as indicated in Figure 5 and classified in Figure 6.
7.1. Physico-Chemical Factors
These abiotic factors, including physical and chemical interventions, affect the soil’s nitrogen dynamics and efficiency. These factors could be induced or of natural origin in agroecosystems such as soil pH, soil texture, aeration, irrigation, moisture content, cation exchange capacity, soil salinity, soil temperature, and application of fertilizers and other harmful elements [79,80,81,82].
7.1.1. Soil pH
Soil pH plays an important role in plant-available N dynamics in soil. Both higher and low pH conditions adversely affect the N availability in soil. At neutral pH (pH 6–7) or in equilibrium conditions, the biological conversion of ammonium to nitrate is fast, and nitrate is easily available to plants. However, in low-pH conditions (pH < 6), nitrification is slow, and crops with the capacity to uptake ammonium may have an advantage over those with the capacity to uptake nitrate. Higher pH increases the chances of volatilization and leaching losses. Calcareous soils, (pH > 7) can lose significant amounts of ammonia gas. The ammonium-containing fertilizer reduces soil pH while nitrate-containing fertilizers increase it. When applied to the soil, ammonium-containing fertilizers such as urea produce H+ ions through their hydrolysis, resulting in a sharp increase in pH, resulting in substantial N loss through volatilization [83].
7.1.2. Soil Temperature
Both soils and ambient temperature can affect the rate of ammonia transportation between soil solution and atmosphere and the hydrolysis of urea in soil [84]. The seasonal and diurnal temperature condition also varies the N emission from soil. Higher soil and atmospheric temperature can enhance the emission, and low temperature reduces the N emission from soil. The overall amount of nitrogen lost through ammonia volatilization may not change much between high and low temperatures, according to some reports, as volatilization may last longer in zones with a cold environment [85]. Low-temperature soils contain relatively higher ammonium than nitrate, while the reverse trend is found for high-temperature soils. The N mineralization and nitrification are generally increased with increasing temperature. Hence, the overall availability of N to crops can be increased with temperature to the optimum level [86].
7.1.3. Soil Texture
The soil texture can affect the rate of mineralization processes, especially crop-residual N [87]. The mineralization of crop-residual N is faster in sandy soils as compared to loamy or clay soils. The lower rate of mineralization under clay soils is mainly because of higher physical protection of soil organic matter and microbial biomass carbon, which reduces microbial activities and thus N mineralization rate [88]. Further, SOC becomes humified, chemically stabilized, and adsorbed onto negatively charged clay minerals with high surface area in clay-dominated soils. Apart from this, clay concentration may alter soil moisture, affecting both the decomposition of SOC and N in clay-rich soils.
7.1.4. Cation Exchange Capacity (CEC)
The potential of soil to hold exchangeable cations (positively charged ions) is termed CEC. It affects the soil’s ability to hold onto essential plant nutrients and provides a buffer against soil acidification. The ammonium ions from any fertilizers will be held on the soil cation exchange site through electrostatic attraction for a short time until they are converted to the nitrate form. Therefore, CEC plays an important role in N dynamics as it helps retain ammonium ions in soil and decreases the concentration of ammonium in soil solutions, thus reducing N losses. The CEC can also act as a buffer to the soil and resist the sudden change in soil reaction. The CEC can affect the ammonia volatilization capacity of soil by resisting changes in soil pH during urea hydrolysis. During urea hydrolysis, a smaller increase in soil pH would be expected in clay soil than in sandy soil [89]. Therefore, due to higher CEC, the volatilization capacity is lower in clay soil than that in sandy soil [90].
7.1.5. Soil Aeration, Irrigation, and Moisture Content
In soil, macropores are filled with water, causing limited gas diffusion and reduced oxygen content of the soil air for a certain period after rainfall or irrigation. The aeration of root zones can improve the NUE of crops [91]. Low oxygen pressure affects soil mechanisms such as microbial activity, soil reactions, and functions of plant roots. Soil oxygen shortage directly impacts microorganisms participating in nitrogen-related processes such as decomposition and nitrogen mineralization. There is a potential for nitrate leaching from the soil and reduced availability to plants in conditions of excessive irrigation and heavy rainfall.
7.1.6. Topography of Land
The land’s topography is one of the dominant factors involving landscape-scale variation in N cycling, especially under tropical climatic conditions [92]. At the local landscape scale, the topography is regarded as a major influencing factor on soil N dynamics, affecting the distribution of soil N availability through microclimate, vegetation, and erosion [93]. On steep topography, heavy rainfall can cause significant hydrologic transport, resulting in the redistribution of N towards downslope lands [94]. The rate of N losses may depend upon the soil type, which is closely linked to the local or regional topography of the land. Therefore, the N cycle is likely to be regulated by interactions between topographically determined climate, soil, and plant species composition of the local and regional land resources.
7.1.7. Amount and Type N Fertilizers
Soil organic matter (SOM) is the major source of N mineralization and plant-available N in the soil. Long-term manure application is considered an efficient management practice to increase soil organic carbon (SOC) accumulation and N mineralization capacity [95]. Nitrate-based fertilizers have the least while ammonium-based products have the greatest potential to acidify the soil. Phosphatic fertilizers also have a small potential for acidification as compared to N fertilizers, due to the lower amounts of this nutrient used. Temporal fluctuations in soil N transformation may be correlated with seasonal patterns of environmental conditions [96]. The large portion of added N through fertilizers resides in the soil and continuously affects N dynamics in the soil after the crop is harvested. However, the long-term effects of added N may be positive or negative. The authors of [97] observed that soils treated with fertilizer N mineralized more N than unfertilized soils, and the N mineralized improved with progressing nitrogen dosing. The higher concentration of ammonium ion in soil solution increases soil acidity and enhances ammonia emission from the soil. Hence, the application of nitrogen fertilizers such as urea can significantly increase the ammonia emission rate [90]. The application of a lower dose of nitrogen improves the soil’s enzymatic and microbial activities, while a higher dose weakens and produces toxic effects [98,99,100].
7.2. Biological and Management Factors
The biotic and management factors could greatly affect the nitrogen dynamics and use efficiency in the soil. These factors could also be human-induced or naturally originated in agroecosystems such as microbial activities, incorporation of crop residues, crop rotation, tillage operations, and species of plants.
7.2.1. Soil Organic Matter, Organic Amendments, Crop Residues, and Tillage Operations
Soil organic matter (SOM) plays an essential role in agriculture by holding and providing plant nutrients and improving overall soil health [101]. The application of organic amendments to soils has improved water-holding capacity, reduced bulk density, improved soil fertility, and improved organic N in soils [102]. Most of the organic matter amendments (OMAs) are applied to the soil surface and incorporated and retained in soils over a long time period. Various mechanisms can act to sustain added N, including physical protection via soil aggregation by binding agents produced by soil microbes during the course of organic matter decomposition. The application of compost can affect the N dynamics either positively or negatively based on the type of soil, as shown in Table 3. The incorporation of crop residues modifies the soil environment, affecting the soil’s microbial activity, and following the nutrient cycle [103]. The mixed applications of crop residues and mineral fertilizers may solve the practical restriction of input availability, but it may also help in improving fertilizer/water use efficiency, physical conditions of soils, crop productivity, crop N synchrony, and N loss reduction through interactive effects between both types of inputs [104].
Table 3.
Studies comparing sensor-based approaches with other N-management strategies (modified from Colaço and Bramley, 2018).
The crop residue input can improve soil N dynamics through the enhancement of WUE and NUE, as well as the input of the substrate of SOM. The microbial and enzymatic activities are higher under crop residue application and zero tillage conditions as compared to conventional tillage without residue application. N2O emission is dominantly affected by several intercultural and tillage operations. N2O emission from No-tilled soils was found to be higher as compared to the tilled soils [119]. The improved crop residue management with N fertilizer or legumes residue considerably enhances the N economy of cereal cropping systems and enhances crop productivity in soils with low N content in a short time. Soil N loss may be reduced by applying effective legume crops, which can provide adequate biological nitrogen fixation input to improve soil N by improved recycling of N through plant residues. The leguminous green manures also play an important role in maintaining NO3 in soil [120].
7.2.2. Microbial Activity in the Soil
Soil N mineralization is a biological process stimulated by microbial activity. Microbially driven N mineralization and transformation are crucial to plant growth and development in the soil. The application of higher rates of N fertilizers than the recommended dose to the crops has not only decreased NUE but has also resulted in long-term environmental degradation problems, i.e., acidification of soil [121]; GHG emissions, especially N2O; eutrophication; and reduced microbial and enzymatic activity in the soil. The rate-limiting step of microbial decomposition of organic matter in the soil nitrogen cycle is the depolymerization of proteins to oligopeptides and amino acids by extracellular proteases, rather than the succeeding mineralization of amino acids to ammonium [122].
N deposition usually stimulates aboveground productivity and affects microbial activities in soil. According to [123], N deposition can negatively affect soil microbial respiration, which has been described by enhanced soil acidification and reduced microbial biomass or hindered oxidative enzyme activities. Zhu et al. [124] reported the positive correlations between the NO3-N immobilization rate, heterotrophic nitrification rate, and fungal biomass in forest soils. The heterotrophic nitrification and immobilization of NO3-N may be significant N transformation pathways influencing agricultural productivity. The nitrogen-fixing bacteria account for about 90% of symbiotic’s N fixation, and the free-living N-fixing bacteria fix only 1–5% of N in the soil. However, the links and interactions between microbial activity and N dynamics in the soil remain unclear.
7.2.3. Plant Species
The species of plant could affect the N mineralization rates in soils [125]. For example, in the subtropical region of eastern China, evergreen broadleaf forests manifest higher nitrogen mineralization rates than fir plantations and secondary shrubs. In soil, the plant species can also affect the soil N cycle. For instance, NO3− production via oxidation of organic N was three times quicker in the soil of Scots pine (Pinus sylvestris L.) than in that of pedunculate oak [126]. During the early successional process, in the shrub associations and coniferous forests, the concentrations of soil mineral N and net N mineralization were significantly lower as compared to post-successionally mixed and broadleaf forests [127]. The understanding of nitrogen mineralization and soil N cycle correlated with the variations between plant species.
8. Approaches for Improving NUE
Improved N management is an important aspect of sustainable crop production which provides the optimal crop yield with the least adverse effects on water and air resources while maintaining and/or improving soil health. The proper intermix of improved fertilizer, soil, water, and crop management practices will maximize the uptake of N by crops, minimize losses of N, and optimize the supply of N from soil and other indigenous sources, resulting in maximizing the N use efficiency. Tailoring N-efficient genotypes with improved agronomic production technologies is required to improve the NUE under the changing climate scenario. Several improved N management approaches have been developed to enhance the N use efficiencies in cereal crops, which are discussed below.
8.1. Genetic Approaches
The main aim of plant breeding is to generate cultivars with desirable characteristics. Plant breeders develop and manipulate the genome of strategic crops to obtain their long-needed engineered cultivars. Plant breeding plays a key role in developing suitable cultivars for sustainable agriculture along with food and nutritional security. The agriculture-based scientific community throughout the globe is observing the effect of a ballooning population on the limited natural (land, water, and soil) resources. Since the mid-twentieth century, plant breeders have developed valuable unique (true-to-type) varieties to balance the effects of demand and supply. As the world economy is demand-driven, the development of varieties with conventional and/or modern breeding approaches is the need of the hour. The expression of NUE is a quantitative trait regulated by a large number of genes [128]. For a deeper understanding of quantitative traits, QTL mapping is an invaluable resource. Many factors, including parental suitability, population size, testing in multiple locations, and genome coverage, contribute to successful QTL mapping for complex traits such as NUE. Variations in the environment can have an impact on QTL. The expression of constitutive QTL is not affected by changes in environmental conditions, while the expression of adaptive QTL is altered by environmental changes. Quantitative trait linkage (QTL) analysis can help us learn more about the connections between traits. Indications of genetic and functional linkage between phenotypically correlated traits are strengthened when the QTL associated with these traits are found to be located in the same genomic regions [128,129].
Finding SNPs, candidate genes, and regulatory pathways that are low-cost, user-friendly, widely distributed, co-dominant, trait-associated, and regulatory could be a major step forward in the race to improve wheat quality around the world. High-throughput genotyping techniques have the potential to increase marker density, which could increase the reliability of identified QTL for nitrogen uptake and utilization-related traits. There are a number of promising approaches to enhancing NUE, such as those that emphasize root architecture or senescence and remobilization. Part of the future genetic gain in cereal crops could depend on our ability to identify major and stable QTL-controlling NUE-related traits and use this information and knowledge in crop-improvement breeding programs [130]. Further research is warranted to quantify differences in feedback effects on the responsiveness of the root: shoot ratio to low N status, as the extent of genetic diversity on this adjustment, is largely unknown at present. Therefore, recent technological advancements have provided a wide range of possibilities and improvements in plant genetics and breeding.
8.2. Conventional Breeding
Over the last two centuries, crop development has been changed to a large extent due to several signs of progress in contemporary genetics. Conventional breeding approaches are dependent on two factors: (1) genetic diversity present in the wild, landraces, and closely related genotypes and (2) advancements in breeding methods through Mendelian and quantitative approaches for elite selection. In most cases, screening existing diversity and selection depends on phenotypic characters. The particular phenotype is highly based on the given genotype and environmental conditions, making it variable and unpredictable. This is the main demerit of classical breeding as it incorporates several stages such as selection, crossing, and then testing under natural environmental conditions.
Indirect genotype selection in the environment may lead to better outcomes. However, the selection environment should be representative of the final growing conditions for the genotypes. Results from NUE tests that show some genotypes perform better under low-N conditions suggest that they may have an adaptive mechanism for this. Long-duration genotypes require more nitrogen (N) for growth and development and, ultimately, for prolonged production of higher yields, so this factor must also be taken into account when selecting the genotype. Environmental nitrogen use efficiency (NUE) studies on crop plants are the standard for selecting NUE-boosting cultivars during the breeding process [131]. Using heterosis to confer NUE in crop plants may be useful for plant breeders. Mid-parent heterosis was found to range from 3.5 percent to 15.0 percent across European studies on average. Hybrids were also said to be more stable and resistant to abiotic stress than pure lines. The hybrids either increased protein content in the grain without changing yield or maintained protein content in the grain while increasing yield. These results suggest that hybrids have a higher nutrient uptake and utilization compared to pure lines. Some research has found that low N levels are associated with a higher frequency of best parent heterosis than high N levels [131,132].
Conventional breeding is very laborious and time- and resource-consuming. To discover the trait of interest and their introgression, a large sample size is required. In addition, breeders must employ a vast array of genome-wide diversity from the available landraces in order to screen and select plants with the desired genetic background. Therefore, advanced breeding techniques provide greater opportunities for genome editing for rapid trait improvement.
8.3. Molecular Breeding Approaches
Molecular breeding refers to the development of a new genotype using molecular information. The NUE of crops grown in the 21st century is low due to the extensive breeding of germplasm with a narrow genetic base just targeting yield-related traits. Assessments are generally carried out with adequate N levels commonly based upon preceding observations and response of chosen varieties [133] (Figure 7). Plant roots play a crucial role in improving NUE; thus, understanding root architecture at a deeper level will further improve our understanding. However, the limited availability of high-throughput and extensive phenotyping tools for plant root architecture has hindered our understanding of genes related to NUE and further using them for advanced breeding programs [134].

Figure 7.
Molecular breeding for enhancing NUE in cereals.
The development of next-generation sequencing (NGS) technologies has allowed for the rapid, cheap, and precise sequencing of both transcriptomes and genomes. The advent of NGS has made it possible to obtain the transcription profiles, genomic data, and mutations such as single-nucleotide polymorphisms (SNPs) of multiple species, despite having only a small number of markers to work with. Genome-wide association studies (MASs) of novel agricultural cultivars require genetic markers, and these data, when combined with biochemical, metabolomic, proteomic, and physiological information, will make it easier to identify the genes responsible for specific phenotypic traits [135]. Genome-wide association studies (GWASs) make use of the data gleaned from reference genomes to map QTL in plants, develop genomic selection programs, and discover interesting mutations in populations of mutated individuals. GWASs are proving to be a useful tool for dissecting the genetic basis of complex biochemical/physiological traits in plants, such as NUE. They can take advantage of the inherent variation as well as the variation created by breeding populations of RIL (recombinant inbred line) offspring with appropriate parents [135].
Moreover, NUE can be enhanced by improving root architecture traits such as by altering root length, density, radius, surface area, and number, etc. Similarly, barley genotypes with higher N uptake affinity show increased root dry weight and volume when measured at the seedling stage [136].
8.4. Quantitative Trait Loci (QTL-Based Approaches)
The improvement in the NUE of crop plants has benefited greatly from recent developments in QTL identification. Finding the important chromosomal areas that segregate NUE can be accomplished with the use of QTL discovery and gene mapping. Under normal and low-N stress situations, QTL underlying grain yield and related attributes may expedite the emergence of NUE variants in crops [137]. The major checkpoints in plants regulating the NUE are some strategic positions along the N fixation pathway such as NR, NiR, GS, and GOGAT enzymes, respectively [138]. A considerable amount of work has been carried out in the past decade on the control of inorganic N assimilation and the co-relation with carbon (C) metabolism at physiological, cellular, and molecular levels. The availability of huge transcriptomic and other omics data at the organism level has deepened our knowledge of the regulation of N absorption in both controlled and changing climatic conditions at the field level. The discovery of prospective candidate genes will act as one of the parameters in NUE regulation for monitoring genotypes for their N sensitivity. QTLs can be used to detect and understand the inheritance of genes involved in nutrient use efficiency (NUE), root growth, development, and structure [135]. Genome-wide association studies (GWAS) and quantitative trait loci (QTL) mapping have helped disentangle the genetics of complex quantitative traits associated with NUE. NUE is regulated by a large number of quantitative trait loci (QTLs), which have been identified in a wide variety of crops, including rice (Tong et al., 2011; Wei et al., 2012), barley (Kindu et al., 2014), maize (Liu et al., 2012), and many more. To facilitate genotype selection in early generations, marker-assisted selection (MAS) makes use of molecular markers associated with a target trait. It makes using traits in breeding programs easier, and the resulting agricultural varieties are better. As a result of their close proximity to the genes responsible for nutrient use efficiency on the chromosomes, a marker-assisted selection (MAS) framework of molecular breeding could use this information to their advantage. QTL mapping on chromosomes is typically laborious and time-consuming, but this process can be sped up with the availability of collections of near-isogenic lines (NILs), which are created to confirm a QTL for the desired trait. Several crops’ breeding programs have made use of NILs for the development of new cultivars and for genetic studies [135].
The chromosomal positions of unidentified genes that affect the quantitative variation in complex characteristics such as NUE can be identified by quantitative trait locus (QTL) mapping. Several investigations have been done to map QTL for NUE in cereals [139,140]. However, this type of genotype has vital effects on the NUE of a plant; for instance, rice N-efficient genotypes IR3932-182-2-3-3-2, IR54853-B-B-318, and IR29723-88-2-3-3 produced a greater yield in comparison to the N inefficient genotypes. On average, a 10% higher yield was observed in an elite rice cultivar, i.e., green super rice (GSR) under field trials with low N input [141]. Furthermore, the QTLs for 22 vital traits were identified in rice with 32 SSSLs through a random block design in different cropping seasons [142]. However, to date, limited scientific research and investigations have been undertaken on specific breeding for N-input systems [143]. Additionally, the functional validation of genes and associated families related to N uptake under control, limiting, and non-limiting experimental conditions needs to be explored. Therefore, molecular breeding of crops for higher NUE increased grain yield over a period of time and has been well documented by many researchers [40]. This knowledge could be further enriched by developing a deeper understanding of genotyping based on sequencing, breeding by modern molecular approaches, next-generation sequencing, high-density and single-nucleotide polymorphism linkage maps, etc. [144].
8.5. Marker-Assisted Selection (MAS-Based Approaches)
Omics approaches provide new possibilities resulting from the next-generation sequencing approaches in which whole-genome sequencing of any plant genotype is possible in a short period of time at very nominal charges. This genomic and transcriptomic-based sequence information provides plant breeders with information about genes, their location, sequence information, and a huge number of DNA markers for generating high-density genetic maps and marker-assisted selection (MAS). It relies greatly on the linkage amongst markers, gene(s), and QTL of interest [145]. This allows the introgression of single or several gene(s) or QTLs from one or more genotypes into the recurrent parent genome. In addition, it permits gene pyramiding, i.e., the introduction of a single or several genes of interest into the particular genomic background [146]. Further, MAS allows for a ‘breeding by design’ strategy, which forecasts the result of a set of crosses as stated by DNA-based marker information. Another application of MAS is ‘genomic selection’ (GS), which relies on the concurrent analysis of outcomes on the phenotype of all existing loci, haplotypes, and markers. GS needs accessibility of phenotypic and genotypic information and the formation of a genomic model so that the alterations in the phenotype are elucidated by the markers examined [147].
8.6. Genetic Engineering
Genetic engineering has been applied in several crops to develop genetically modified organisms (GMOs) by forward or reverse genetics approaches. Genes have been integrated from other organisms/plants into different plant species to obtain the desired characteristics. Genes-silencing approaches hamper gene expression by inhibiting translation through mRNA targeting [148]. Genes associated with N metabolic pathways such as nitrate transporters, ammonium transporters, and associated transcription factors have been engineered into cereals to enhance NUE [149]. Commonly, constitutive promoters such as CaMV35S or specific promoters such as OsNAR2.1 have been used to augment NUE in transgenic lines [150]. Additionally, the OsNRT1.1B allele of indica rice has been genetically engineered into japonica rice to enhance NUE [151]. Likewise, [152] introduced the ZmDof1 gene from Zea mays to rice and found higher carbon and nitrogen fixation under conditions of lower nitrogen. Further, the overexpression of the ASN1 gene in Arabidopsis improves the nitrogen status of seeds [153]. The constitutive expression of a cytoplasmic glutamine synthetase gene (GS1) in tobacco from alfalfa resulted in a higher photosynthetic rate in areas with low N input [154]. These results indicate that transgenic tobacco lines are more efficient in utilizing N under limiting conditions. Similarly, GS1 gene over-expression in poplar also resulted in 41% higher growth rate in transgenic lines.
The CRISPR-Cas9 genome-editing tool has changed the era of genetically engineered plants to develop elite crop varieties [36,155,156,157]. The CRISPR system is defined as Clustered Regularly Interspaced Short Palindromic Repeats, a type of immune defense system naturally found in prokaryotes. The CRISPR nuclease system has revolutionized gene modification for the maneuvering of the genome of a living being [158]. The CRISPR–Cas9 system can be utilized more efficiently to improve the genomes of cereals to improve their NUE. The CRISPR–Cas9 system has proven applications in several research areas such as biotic and abiotic stresses and improving a particular crop’s quality [159]. Due to the application of CRISPR/Cas9, the expression of symbiotic N fixation genes in Lotus japonicus was reduced, and the possibility of genome editing in legumes was also explored. Furthermore, single base modification in OsNRT1.1B (a nitrate transporter gene) has been carried out by a fusion of nCas9 and human APOBEC3A nuclease and has been reported to enhance NUE in rice [160]. Sathee et al. [161] also reviewed several studies to target NUE in cereals using CRISPR/Cas9. In this study, various genome editing techniques for utilizing essential negative and positive regulators were discussed. CRISPR variants that achieve transcription activation or interference with dCas9 are known as CRISPR activation (CRISPRa) and CRISPR interference (CRISPRi). Plants can create targeted DNA methylation and gene activation using the dCas9-SunTag technology. Genome editing can target negative regulators of nutrient signaling, which can enhance food absorption and stress signaling in resource-constrained environments. A successful method to produce precise alterations is promoter engineering using CRISPR/dead (d) Cas9 (dCas9) cytosine and adenine base editing, as well as prime editing. The additional benefit of using transcriptional activators and/or repressors to specifically overexpress desired genes is another benefit of the CRISPR/dCas9 system.
Similarly, numerous N transporter genes have been genetically engineered in rice to improve its NUE, for instance, low-affinity nitrate transporter NRT1.1b [162], high-affinity nitrate transporter NAR2.1, NRT2.3a [163], and ammonium transporter AMT1.1 [164]. Modern genetic tools and techniques have helped improve the NUE of cereals and other strategic crops. Therefore, there is a call for advancing from laborious and time-consuming conventional breeding approaches to more effective modern breeding tools, viz., omics and CRISPR/Cas9. Nonetheless, robust genetic engineering target multiplexing, precise editing, and efficient transformation and regeneration would bring another agricultural revolution. As NUE is a complex and multigenic trait, regulated by various sets of genes, its improvement in cereals to enhance yield is a pressing priority.
9. Agronomic Approaches
To achieve the goal of sustainable crop production along with higher NUE, N needs to be managed so that its losses remain at a minimal level. Some improved agronomic strategies of N management, which involve modifications of soil, plant, and fertilizer variables in favor of maximum economic crop yields and NUE, are discussed in this section. Therefore, by combining several breeding and agronomic approaches, a balanced way to improve the NUE of cereals and maintain soil fertility and grains with higher yield and nutrient traits is possible. Hence, a holistic approach is needed for identifying and developing cultivars with enhanced N uptake and translocation efficiency.
9.1. Site-Specific N Management (SSNM)
Agricultural land, especially in developing countries, is typically marginal and small, with high spatial and temporal variability linked with differences in soil fertility, the nature of fertilizers, and its application methods and the diversified use of other local resources available to farmers. N application through blanket recommendations occasionally permits farmers to use N fertilizers in excess of the permitted amounts to achieve extreme yields, resulting in a lower NUE. It is essential to strike a balance between a crop’s nitrogen needs and soil nitrogen stores to achieve sustainable agricultural yields with high NUE and minimal soil and air pollution [165]. SSNM is a set of N-management principles that aims to optimize the supply of soil N over time and space to match the requirements of crops. The major emphasis of SSNM is to feed the N to crops at an optimal amount and time to achieve higher yield and NUE. It optimizes the fertilizer use to bridge the deficit between the N requirements of a crop and N supplied from natural sources [166]. It could help reduce the cost of cultivation and environmental pollution and improve the crop yield over blanket N application (Sapkota et al. 2021). The site-specific nutrient management (SSNM) strategy, which is based on soil-nutrient supply and nutrient-uptake need for a defined yield, has recently demonstrated the potential to increase farm profit by ensuring that the right amount of nutrients is supplied to a variety of crops [167]. The SSNM method is a useful tool for delivering nutrients to crops to increase crop output, in addition to aiming to reducing fertilizer consumption. It also attempts to improve nutrient use efficiency, which will result in greater net returns per unit of fertilizer invested. In contrast to BSR, which is still widely used in a number of developing nations, SSNM guarantees enhanced production and soil health over the long term (Timsina et al., 2013). Over the last decade, some attempts have been made to carry out SSNM in major cereals, i.e., rice, maize, and wheat, using LCC, chlorophyll meter, and nutrient expert (NE) tools, which are discussed as follows.
9.2. Chlorophyll Meter
Crop N status can be estimated using a chlorophyll meter, as most N in the plant system is contained in the rubisco enzyme found in chloroplasts and chlorophyll proteins, and there is a close relationship between leaf N and leaf chlorophyll content. The chlorophyll meter, popularly known as the SPAD meter (soil–plant analysis development), can quickly and reliably provide an estimate of leaf N status of the crop by measuring chlorophyll content. It was also recommended to assess the effectiveness of N applied late in standing crops to increase grain yield and protein content [168]. Rhezali and El-Aissaoui (2021) observed a significant linear correlation between absolute SPAD values and leaf N concentrations of corn.
The Minolta SPAD-502 has been commonly used to estimate corn N status and refine in-season N management [169]. Ali et al. [170] worked out a critical SPAD value of 37 for fertilizer N application using the Cate Nelson procedure and observed that the application of 30 kg N/ha whenever the SPAD meter reading fell below a critical value gave the optimum rice yield with higher NUE over the blanket recommendation for N fertilizers. A threshold SPAD value of 37 for tailoring fertilizer N application to wheat in the eastern Indo-Gangetic plain was worked out by [171]. Singh et al. [172] used the SPAD meter for guiding the application of fertilizer N at the MT stage of irrigated wheat and observed that wheat responded to the top dressing of 30 kg N/ha when the SPAD reading fell below 44 at the maximum tillering stage with the 20% yield increase at an SPAD value of 42 or less.
9.3. Leaf Color Chart (LCC)
The cost of a chlorophyll meter is high, which is beyond the reach of poor Asian farmers. Thus, an inexpensive alternative tool, LCC, was developed from a Japanese prototype by the International Rice Research Institute (IRRI) in the Philippines to monitor relative leaf greenness as an indicator of leaf N status [173] and can help farmers in efficient N management [174]. Like the chlorophyll meter, LCC also needs the critical color shades threshold to be determined for guiding the N application [175]. For example, [176] found the following threshold values: 3 for basmati rice, 5 for hybrid maize, and 4 for wheat. LCC has been successfully used for N management in different crops and found to be effective in avoiding excess use of N fertilizer, improved productivity, and NUEs by enhancing the congruence between the time of fertilizer application and the need of the plant [177,178,179,180] (Table 4). Leaf color charts are effective and low-cost tools that help farmers improve their N management, and efforts are underway to stimulate the technology at a wider scale among Asian rice farmers.
Table 4.
Effect of leaf color chart (LCC)-based N management on grain yield, nitrogen uptake, and NUE.
9.4. Nutrient Expert (NE)
Nutrient Expert (NE) is a recently developed decision-support system software based on precise nutrient management technology that helps improve crop yields, environmental quality, and overall agricultural sustainability.
Table 5.
Effect of Nutrient Expert (NE)-based N-management practices on grain yield of different cereals.
Table 5.
Effect of Nutrient Expert (NE)-based N-management practices on grain yield of different cereals.
| Crop | N Management Practice | Yield (t/ha) | % Increase in Yield with NE | Place | Source |
|---|---|---|---|---|---|
| Rice | NE | 5.46 | - | Morang (Nepal) | [184] |
| GR | 4.79 | 13.99 | |||
| FFP | 4.43 | 23.25 | |||
| NE–SSNM (LCC) | 6.67 | - | Pantnagar (India) | [185] | |
| FFP | 6.06 | 10.07 | |||
| NE | 5.14 | - | Lamjung campus (Nepal) | [186] | |
| GR | 4.24 | 27.86 | |||
| FFP | 4.02 | 5.47 | |||
| NE | 5.79 | - | Lamjung (Nepal) | [187] | |
| FFP | 4.21 | 37.53 | |||
| Maize | NE | 8.4 | - | Indonesia | [188] |
| FFP | 7.5 | 12.00 | |||
| NE | 9.1 | - | Philippines | [188] | |
| FFP | 7.5 | 21.33 | |||
| NE *1 | 7.94 | - | Southern India | [189] | |
| FP *1 | 6.87 | 15.57 | |||
| NE *2 | 9.11 | - | Southern India | [189] | |
| FP *2 | 7.98 | 14.16 | |||
| NE | 8.4 | - | India | [190] | |
| FFP | 7.8 | 7.69 | |||
| NE | 10.10 | - | China | [191] | |
| FP | 9.90 | 2.02 | |||
| NE | 12.30 | - | Northeast China | [192] | |
| FFP | 11.40 | 7.89 | |||
| SSNM-NE | 5.83 | - | Pantnagar (India) | [181] | |
| FFP | 4.77 | 22.22 | |||
| NE | 8.06 | - | Morang (Nepal) | [193] | |
| FFP | 4.53 | 77.92 | |||
| NE | 9.22 | - | Jhapa (Nepal) | [193] | |
| FFP | 4.94 | 86.64 | |||
| Wheat | NE–SSNM (LCC) | 4.32 | - | Pantnagar (India) | [185] |
| FFP | 3.02 | 43.05 | |||
| SSNM-NE | 4.55 | - | Pantnagar (India) | [181] | |
| FFP | 4.22 | 7.81 | |||
| NE | 4.02 | - | Morang (Nepal) | [193] | |
| FFP | 2.06 | 95.15 | |||
| NE | 4.71 | - | Jhapa (Nepal) | [193] | |
| FFP | 3.00 | 57.00 | |||
| NE | 7.17 | - | China | [194] | |
| FP | 6.88 | 4.22 | |||
| NE | 8.1 | - | North-central China | [195] | |
| FP | 7.9 | 2.53 |
FFP—Farmers Fertilizer Practices; FP—Farmers Practices; NE—Nutrient Expert; SSNM—Site-specific nitrogen management; LCC—Leaf color chart; RF—Recommended fertilizer; GR—Government recommendations; PFP—Partial factor productivity; *1 indicates the data for rice cv. Hybrid PHB-71 during 2002; *2 indicates the data for maize cv. 30V92.
NE is an emerging N management diagnostic tool that applies N input variables at the right place and time with the right amount per crop demand [196]. NE has been tested for N management in rice, wheat, and maize. Results show that it avoids excessive use of N fertilizer and improves productivity and NUEs compared to farmer practice by matching N fertilizer application time and plant needs [197]. Similarly, [198] reported 17, 56, and 58% reductions in NPK fertilizer use in maize when applied with NE recommendations compared to FP. Several other researchers also significantly improved the yield of different cereals with NE-based N application (Table 5).
9.5. Enhanced-Efficiency Nitrogen Fertilizers (EENFs)
NUE can be improved by the use of EENFs, which are developed to reduce N losses. EENFs reduce ammonia volatilization and nitrification–denitrification losses by reducing or controlling the rate of N release from fertilizer (slow-release fertilizers) by inhibiting the nitrification process (nitrification inhibitors) and urease activity (urease inhibitors) and ultimately help synchronize the N supply as per the crop demand [199].
9.6. Slow Release Fertilizer
High N application leads to low NUE due to various N losses. Multiple top dressings of N fertilizers at different crop growth stages can be performed to match plant demand to improve grain yield and NUE. However, such multiple top dressings incur additional labor costs. In this regard, slow-/controlled-release fertilizer holds a great promise, which closely matches the crop N requirement with a single application, thereby saving labor costs. Slow-release fertilizers include those formed by urea and aldehydes’ condensation products (such as IBDU—isobutylidene diurea) and coated or encapsulated fertilizers’ sulfur-coated urea and polymer-coated urea [200]. Different N losses can be minimized due to the application of controlled-release N fertilizers owing to delayed N-releasing patterns can synchronize with the plant demand. Many studies indicate that slow-release fertilizers can improve the grain yield and NUE of major cereals [201]. Reference [202] noted that the ratio of NO3−/NH4+, an indicator for nitrification, was lower with the application of polyurethane-coated urea than urea by itself, indicating a slower nitrification rate. A lower NO3−/NH4+ ratio is beneficial for higher N uptake, growth, and yield [203].
9.7. Nitrification Inhibitors
Nitrate leaching and denitrification losses from fertilizer N can be reduced by maintaining N in NH4+ form for a longer period. This can be possible by inhibiting nitrification (NH4+-N to NO3−-N) through the addition of nitrification inhibitors and conventionally used N fertilizer [204]. Though a lot of NIs (nitrapyrin and dicyandiamide (DCD)) have been developed, they are less popular among resource-poor farmers in Asian countries due to their high cost and limited availability [205]. In this regard, neem-coated urea (NCU) is a cheaper alternative for resource-poor farmers. The use of neem-coated urea reduced the nitrate concentration. It enhanced the ammonium ion concentration in soil [206], which can be of great use in the case of transplanted rice in low-land ecosystems, where the losses due to denitrification and nitrate leaching are substantial. Aboveground dry matter accumulation in rice due to NCU is relatively low at early stages, but at later stages, it supersedes PU, which might be due to the conservation of nitrogen, which became available at later stages with the use of NCU synchronized with the plant demand, enhancing the grain and straw N content [207] over PU. Several neem-based NIs (NCU, neem-cake-coated urea, and neem-extract-coated urea) have been reported to increase enhanced yield and profitability, N uptake, and NUEs in rice with the use of NCU over PU. However, the effect was more pronounced in sandy loam soil than in clayey loam soil [208].
9.8. Urease Inhibitors
Urea is the most important N fertilizer worldwide (IFA 2017) because of its higher N concentration and lower cost of production in comparison to other N sources [209]. After urea is applied, it undergoes urea hydrolysis through urease activity, which increases the pH of the soil surrounding the urea granules and causes loss of ammonia (NH3) to the atmosphere through volatilization, which is a serious environmental issue. N lost through NH3 volatilization can reach up to 40% of applied N fertilizer or even more under hot and humid conditions [210,211]. The amount of N lost as NH3 depends upon soil and environmental conditions (texture of soil, soil moisture content, and temperature) [211] and N application rate.
Compounds such as hydroquinone, some benzoquinones [212], phenolic aldehydes, and benzoylthioureas have inhibitory effects on the activity of urease, but the most widely used compound is N-(n-butyl) thiophosphoric triamide (NBPT) [211]. The meta-analysis of [213] showed that NH3 loss was only 15% when NBPT was used along with urea, but it was 31% with untreated urea. Cantarella et al. [211] compiled the results of different studies and found that the mean increment in yield with the use of the urease inhibitor (UI) NBPT along with urea was −0.8% in sugarcane, 1.8% in cotton, 4.1% in corn, 5.2% in bean, 6% in barley, 7.6% in rice, 8.4% in pasture, and 10.2% in wheat compared to that of untreated urea, with the mean reduction in NH3 loss up to 53.2%. Though urease inhibitors have been proved to reduce NH3 volatilization, their limited shelf life hinders effective inhibition over a longer period; hence, there is a need to improve the shelf life of UIs [211].
9.9. Nano-Fertilizers
Nanotechnology is a novel approach that implements a similar method as the formulation or polymer coating of urea with urease-regulating nutrient elements but at different scales of particle size. Here, the sizes of polymer coating materials or nutrient elements (≥1000 nm) are reduced by 1000 to 10 times, resulting in a corresponding material size of 1 nm to 100 nm (nano-scale) [214]. Nano-scale additives or nanofilms and nanopolymers of other nutrients were also utilized for modifications of N fertilizers for controlling/slowing the N release, improving crop productivity, N uptake, and NUE. The development of fertilizer products utilizing a nanoscale process is called nano-fertilizer or nano-enabled fertilizer, which in most cases has been found to reduce the N losses and increase NUE and productivity of crops over conventional fertilizers [215].
A slow-release nano N-fertilizer developed using nano-encapsulation of urea with the use of hydroxyapatite (HAP), a naturally occurring P-mineral and ground wood material of Gliricidia sepium, resulted in a 44% reduction in the N release rate after 60 days compared to conventional urea (CU). This slow-release action was due to the interaction of urea with the nano-HAP by its amine and carboxyl groups, creating moderately strong bonds [216]. This nano-encapsulated fertilizer increased the grain yield of rice and its leaf N by 7 and 6%, respectively, with 50% less N in the urea–HAP formulation than CU [217].
Using polycaprolactone, polyacrylamide hydrogel, and montmorillonite clay, urea nanocomposites were created that reduced N release by 25%, 20%, and 21%, respectively, compared to CU. Moreover, these compounds have been demonstrated to dramatically reduce N2O emissions in wheat fields [218]. Likewise, nano-carbon, urea formaldehyde composites, and zeolite nano-composite have been tested for their efficiency in slowing down the release and transformation of N from urea. Results showed significant reductions in total N release or amount of NH4+ or NO3− in soil incubation studies [219].
9.10. Precision Farming
Precision farming is the use of information and technology to identify, analyze, and demonstrate the variability in time and space dimensions in every aspect of agricultural operations for efficient management of resources with an aim to conserve environmental resources, as well as to achieve maximum production. The in-field variation with respect to the available N is quantified with the help of the global positioning system (GPS), the geographic information system (GIS), and remote sensing techniques, and the exact amount of fertilizer N is applied at the right time and right place through a variable rate applicator [220] so that precise and timely application of N as per the crop need across the landscape can be achieved [221]. The entire agricultural field is divided into different zones, and fertilizer and other inputs are applied as per the site-specific input needs. However, it requires a thorough understanding of the delineation process, the spatial and temporal variability in the field and crop for a particular input, and corresponding quantified needs and their ultimate impact on crop yield [222]. Moreover, Grell et al. (2021) showed the instantaneous prediction of NO3 levels in the soil using the leading machine model. In the aforementioned study, they utilized the point-of-use method of measurement of NH4+, soil electrical conductivity, pH, easily accessible weather, and timing data (R2 = 0.70). Therefore, the method used for enough precision and application of nitrogenous soil nutrients may be identified and forecasted, allowing fertilization schedules to be adjusted for crop needs and preventing overfertilization while enhancing crop yields.
Electronic sensors are commonly used to diagnose the site-specific variability in crop N demand and soil supply, utilizing the various crop and soil parameters [223]. Similarly, non-destructive proximal canopy reflectance sensors have emerged as the tools for SSNM with the most potential [224]. Further, sophisticated N management through local or remote N sensors can be carried out to assess the supplemental N requirement of crops. Typically, sensor-based N applications are top-dressed during mid-season and can synchronize with N uptake by the crop. Further, sensor-based N rates are always lower than the fixed-stage split N application. It is obvious that the split application of N fertilizer will increase the NUE over pre-plant application whether it is applied with the use of a sensor or not. Sensor-guided N application reduces the environmental impacts associated with N fertilizers but is mostly not profitable compared with the prevailing practices.
9.11. Improved Method of N Application
Methods of fertilizer N application affect N losses and thereby crop performance. For example, broadcasting of N fertilizer involves greater N loss in rice through ammonia volatilization [225] and reduces NUE. On the other hand, the deep placement of N fertilizers such as urea or urea super granule (USG) can enhance the grain yield and N recovery compared to broadcast N application. Higher NUE with the placement of USG than urea is ascribed to the reduction in N losses (NH3 volatilization, nitrification–denitrification), regulated N supply to the crop, and its placement below the oxidized zone [226].
Despite the proven advantages of deep placement of USG, it is not commonly used by farmers, primarily due to its unavailability and the labor required for its placement [227]. Band placement of N fertilizer is another option that can reduce NH3 volatilization losses and improve NUE compared to soil surface application, especially in this method’s water-scarce conditions [228]. Further, the placement of urea results in higher NH4+ concentration in the fertilizer zone, which has toxic effects on nitrifiers, causing the inhibition of the nitrification process, thereby minimizing the leaching loss. Fertilizers are applied directly to the zone of maximum root activity through emitters in the case of drip fertigation, which can enhance the fertilizer use efficiency compared to other fertilizer application methods (broadcasting/furrow placement). Better water economy, along with higher yields, is also reported for drip fertigation. Therefore, improvement in water and NUE can be achieved through the adoption of drip fertigation [229,230,231].
9.12. Integrated/Balanced N Management
Continuous and imbalanced use of chemical N fertilizers results in an imbalance in the soil’s N status, reducing crop yield and soil quality and accelerating deficiencies of other nutrients [232]. In the past, it was evident that an increase in food grain production over the years was associated with the application of proportionately higher amounts of nutrients, implying lower nutrient use efficiency of applied fertilizers [233]. Further, higher doses of fertilizers (especially N) are associated with environmental problems such as nitrate contamination in groundwater, eutrophication, and increased greenhouse gas emissions [234]. In some cases, nutrient application imbalance is observed, leading to nutrient mining, reducing productivity and NUE [235]. Hence, the balanced fertilization of macro and micro-nutrients is the prerequisite for the realization of higher productivity [236]. Integrated nutrient management (INM) is the key to reducing fertilizer-derived environmental problems with the enhancement of yield [237] and nutrient-use efficiency [233].
Optimum nutrient availability in plants is the pre-requisite for yield enhancement, which can be met by maintaining good soil fertility. Improving NUE can be achieved by increasing the yield while reducing the losses to the environment by managing the optimum levels of nutrients in the root zone of crops to meet their biological potential. INM practices reduce the losses of N through runoff, volatilization, leaching, and emissions and result in higher NUE [233]. The main principles of INM are optimizing the nutrient inputs considering all the possible sources, matching the soil nutrient supply with that of the crop over space and time, and minimizing the N losses while improving the crop yields [238]. The application of mineral fertilizer and animal manure has been reported to result in a higher and more stable yield in wheat and maize than the application of mineral fertilizer alone with higher mean NUE in a long-term experiment in China [239]. The NUE increased when one-third of the recommended dose of N was supplemented through organics (paddy straw, green gram residues, and sesbania). The sustainability of crop production and soil health could be maintained or improved by the integrated use of organic and inorganic N fertilizers [240].
9.13. Improved Water Management
N and water management have an interactive effect [241] regarding N accumulation, translocation, and partitioning. Excess fertilizer N and irrigation application cause greater N movement along the moving water beyond the root zone, increasing the N losses via leaching [242]. Surface irrigation methods involve water losses that also lead to N losses via seepage or percolation [243]. However, drip irrigation can reduce surface run-off, evaporation, and deep percolation losses while placing water precisely to maintain the optimum level of available soil moisture in the root zone of crops. AEN and PEN increase with an increase in water availability. For example, AEN and PEN values for severe water stress (35± 5% field capacity, FC), mild water stress (55± 5% FC), and adequate water conditions (75± 5% FC) were 4.1, 6.7, and 7.3 kg/kg N and 15.9, 22.6 and 24.7 kg/kg N, respectively [241]. Under low water availability, the release of controlled-release urea was restricted, possibly due to mineral N availability and root activity, resulting in a lower grain yield. Furthermore, the N uptake was reduced under low water availability, causing more fertilizer to be left in the soil rather than absorbed by the crop [244]. With the scarcity of water, subsurface drip irrigation is receiving more attention, especially for row crops. Subsurface drip irrigation reduces the net irrigation requirement with higher yields and NUEs. The reduction in net irrigation can reduce different N losses (deep percolation and runoff) [245] and precise application of N fertilizers as per the time and dose and also due to the uniform application of fertilizer N directly to the crop root zone.
9.14. Sustainable Crop Rotation
Crop rotation is very important for maintaining soil fertility and reducing the incidence of pests and soil pollution. Changing crops in rotation results in changes in soil fertility due to atmospheric N fixation, the application of variable rates of chemical fertilizers, and the incorporation of crop residues [246]. Further, diversifying the rotation has been associated with an increase in organic matter, soil quality, aggregate stability, decreased soil erosion, and reduced emission of GHGs. Crops grown in rotation give greater yield and allow the economic dose of N to be minimized due to the diversification, which results in higher NUE than monoculture [247]. For example, a rice–wheat system significantly improved the REN in rice (50.4%) than a rice–fallow system (31.9%) at 142.5 kg ha−1 of N application. The continuous growth of cereal leads to a reduction in yields and enhances the external application of nutrients. Introducing a legume crop to a cereal-based cropping system can maintain soil fertility and can give stable yields. The authors of [248] reported that the substitution of a rice–wheat system for a rice–potato–green gram system improved the system’s productivity, profitability, and soil health. Including legumes in cropping systems, especially for green manuring, fodder, or grain purposes is an assured agro-technique that improves NUE.
9.15. Crop Residue Management
The intensification of cropping systems to meet estimated food requirements and food security has created a problem in terms of residue management. For instance, in intensive rice-based cropping systems (e.g., double rice cropping systems in China and rice-wheat cropping systems in India), the sowing of the next crop after the wet season rice is generally delayed due to improper residue management because of a lack of alternative uses of crop residues, resulting in burning of residues for sowing of the second crop (rice or wheat) [249,250]. The burning of residues has multiple consequences, such as atmospheric pollution, loss of biodiversity, human health hazards, and GHG emissions, and decreases the return of nutrients such as N, P, and S. The incorporation of crop residues not only leads to the addition of nutrients to soil but also improves the soil properties, increasing productivity and the use efficiency of applied fertilizers [251]. However, a higher C:N ratio of cereal crop residue may lead to the immobilization of fertilizer N in the soil if incorporated into the soil, and the crop may experience temporal N deficiency.
This can be minimized by applying an additional quantity of fertilizer N or by composting crop residues and applying the compost to the soil. Wheat straw is generally used as animal fodder in India and is collected from fields, but rice straw is of limited use, and its management is problematic [252]. Due to labor scarcity, farmers use combined harvesting for the timely sowing of wheat crops. However, loose rice straw after combined harvesting hinders the operation of zero-till seed drill, and hence farmers often burn rice straw. The addition of crop residues to the soil surface or their incorporation enhances the productivity, NUE, and soil health under major cereals such as rice, maize, and wheat [253,254,255]. In a meta-analysis, [256] noted that in situ crop residue retention led to a 5.2% increment in rice grain yield in China. Further, the crop residue retention gave similar yields with 29.4, 8.3, and 21.9% reduced application of N, P, and K fertilizers, respectively, suggesting that a portion of the recommended dose of fertilizer can be saved due to residue retention.
10. Conclusions and Future Prospects
Although N is the most limiting nutrient for agricultural production, its overuse, especially in the form of synthetic N fertilizers, is associated with high losses that pollute the environment and increase the concentration of greenhouse gases, and it is also responsible for human and animal health implications. Therefore, N use efficiency must be improved to supply sufficient N to meet increased food demand and maintain a safe environment. The competent utilization of known N efficient mechanisms and candidate genes can provide new clues for tailoring genotypes that generate optimum yields, thus improving the NUE in light of the forecasted climate change scenarios. Genetic engineering and biotechnological tools should also be integrated into the breeding programs to target particular candidate genes responsible for higher N efficacy. Therefore, genetic engineering and molecular breeding tools (QTLs/candidate genes and MAS) should be combined with modern breeding strategies to achieve genotypes with enhanced N recovery. The adoption of some of the recent agronomic management strategies such as site-specific N management, fertilizers with enhanced use-efficiency, resource-conservation practices, precision farming, and nano-fertilizers can help farmers reduce the environmental losses of N from soil–plant systems, thus improving NUE at the farm level. Therefore, by combining several breeding and agronomic approaches, a balanced way to improve the NUE of cereals, along with maintaining soil fertility and grains with higher yield and nutrient traits, is possible. Hence, a holistic approach is needed to identify and develop cultivars with enhanced N uptake and translocation efficiency to sustain cereal production under climate change scenarios.
Since NUE is a complicated multigenic trait, improving it is challenging. Since CRISPR/Cas9 has mostly been used to study genes’ negative regulators, it is anticipated that other genes of this type will be discovered in the future. For NUE research, it is crucial to advance our understanding of genes, transcription factors, and microRNAs, as well as to clarify the underlying molecular and physiological mechanisms of N routes. The key challenge for breeders and agronomists is to enhance the uptake and utilization of N by the root systems of these targeted cereals to improve NUE, especially under the climate change scenario. Improved agronomic technologies/practices will contribute significantly in this regard; however, breeding approaches have not been fully explored, especially in the case of root traits, one of the most important traits associated with NUE. Therefore, better exploration of root and phenotypic characteristics employing modern genetic approaches can provide scope for NUE improvement. The most useful resource for improving NUE is the utilization of existing genetic variation among germplasm. Unfortunately, limited genetic variation is available in modern germplasm pools, the exploration of which has been limited. Therefore, there is a huge scope to collect the diverse germplasm with higher genetic variation to enrich the genetic pools and explore and identify alleles with higher NUE traits.
Author Contributions
M.R.Y., S.K. (Sandeep Kumar 1), M.K.L. and D.K.: Conceptualization and writing—original draft preparation; R.K., R.K.Y., S.K. (Sandeep Kumar 1), G.N., J.S., P.U., N.K.M., T.M, A.P.G. and V.P.K.: Writing—original draft preparation, figures and tables; M.R.Y., S.K. (Sandeep Kumar 2), M.K.L., V.D.R., P.K.J. and T.M.: Supervision and writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by the Ministry of Science and Higher Education of the Russian Federation (no. FENW-2023-0008) and by the Strategic Academic Leadership Program of the Southern Federal University ("Priority 2030").
Data Availability Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the review.
Acknowledgments
The authors would like to acknowledge the Director of RARI for support and guidance. Authors would also thanks Russian Phytopathology Research Institute, Moscow, Russia.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Ahmad, T.; Saboor, A.; Baig, I.A.; Afzal, A. Socio-Economic Policy Imperatives for Sustainable Food System in Pakistan. J. South Asian Stud. 2021, 9, 113–131. [Google Scholar] [CrossRef]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of Nitrogen in Agriculture and Environment: Agronomic, Eco-Physiological and Molecular Approaches to Improve Nitrogen Use Efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Khan, L.U.; Altaf, M.M.; Jahan, M.S.; Nawaz, M.A.; Naz, S.; Shahid, S.; Lal, M.K.; et al. Protective Mechanisms of Melatonin Against Vanadium Phytotoxicity in Tomato Seedlings: Insights into Nutritional Status, Photosynthesis, Root Architecture System, and Antioxidant Machinery. J. Plant Growth Regul. 2022, 41, 3300–3316. [Google Scholar] [CrossRef]
- Lal, M.K.; Singh, B.; Tiwari, R.K.; Kumar, S.; Gopalakrishnan, S.; Gaikwad, K.; Kumar, A.; Paul, V.; Singh, M.P. Interactive Effect of Retrogradation and Addition of Pulses, Cooking Oil on Predicted Glycemic Index and Resistant Starch of Potato. Starch/Staerke 2022, 74, 2100221. [Google Scholar] [CrossRef]
- Raigond, P.; Parmar, V.; Thakur, A.; Lal, M.K.; Changan, S.S.; Kumar, D.; Dutt, S.; Singh, B. Composition of Different Carbohydrate Fractions in Potatoes: Effect of Cooking and Cooling. Starch/Staerke 2021, 73, 2100015. [Google Scholar] [CrossRef]
- Lal, M.K.; Tiwari, R.K.; Kumar, R.; Naga, K.C.; Kumar, A.; Singh, B.; Raigond, P.; Dutt, S.; Chourasia, K.N.; Kumar, D.; et al. Effect of Potato Apical Leaf Curl Disease on Glycemic Index and Resistant Starch of Potato (Solanum tuberosum L.) Tubers. Food Chem. 2021, 359, 129939. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.O.; White, J.C.; Franklin, F.A.; Melse-Boonstra, A.; Koele, N.; Pandey, R.; Rodenburg, J.; Senthilkumar, K.; Demokritou, P.; et al. Safeguarding Human and Planetary Health Demands a Fertilizer Sector Transformation. Plants People Planet 2020, 2, 302–309. [Google Scholar] [CrossRef]
- Rashmi, I.; Roy, T.; Kartika, K.S.; Pal, R.; Coumar, V.; Kala, S.; Shinoji, K.C. Organic and Inorganic Fertilizer Contaminants in Agriculture: Impact on Soil and Water Resources. In Contaminants in Agriculture: Sources, Impacts and Management; Springer: Cham, Switzerland, 2020; pp. 3–41. [Google Scholar] [CrossRef]
- A., A.; K.R., R.; Mathew, J. Fate of the Conventional Fertilizers in Environment. In Controlled Release Fertilizers for Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2021; pp. 25–39. [Google Scholar] [CrossRef]
- Mushtaq, N.; Singh, D.V.; Bhat, R.A.; Dervash, M.A.; Hameed, O. bin Freshwater Contamination: Sources and Hazards to Aquatic Biota. In Fresh Water Pollution Dynamics and Remediation; Springer: Singapore, 2020; pp. 27–50. [Google Scholar] [CrossRef]
- Lal, M.K.; Singh, B.; Sharma, S.; Singh, M.P.; Kumar, A. Glycemic Index of Starchy Crops and Factors Affecting Its Digestibility: A Review. Trends Food Sci. Technol. 2021, 111, 741–755. [Google Scholar] [CrossRef]
- Devi, R.; Behera, B.; Raza, M.B.; Mangal, V.; Altaf, M.A.; Kumar, R.; Kumar, A.; Tiwari, R.K.; Lal, M.K.; Singh, B. An Insight into Microbes Mediated Heavy Metal Detoxification in Plants: A Review. J. Soil Sci. Plant Nutr. 2022, 22, 914–936. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Mangal, V.; Altaf, M.A.; Sharma, S.; Singh, B.; Kumar, M. Insight into Melatonin-Mediated Response and Signaling in the Regulation of Plant Defense under Biotic Stress. Plant Mol. Biol. 2022, 109, 385–399. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Bashyal, B.M.; Shanmugam, V.; Lal, M.K.; Kumar, R.; Sharma, S.; Vinod; Gaikwad, K.; Singh, B.; Aggarwal, R. Impact of Fusarium Dry Rot on Physicochemical Attributes of Potato Tubers during Postharvest Storage. Postharvest. Biol. Technol. 2021, 181, 111638. [Google Scholar] [CrossRef]
- Kumar, A.; Nayak, S.; Ngangkham, U.; Sah, R.P.; Lal, M.K.; Azharudheen, T.P.; Behera, S.; Swain, P.; Behera, L.; Sharma, S. A Single Nucleotide Substitution in the SPDT Transporter Gene Reduced Phytic Acid and Increased Mineral Bioavailability from Rice Grain (Oryza sativa L.). J. Food Biochem. 2021, 45, e13822. [Google Scholar] [CrossRef] [PubMed]
- Nayak, L.; Panda, D.; Dash, G.K.; Lal, M.K.; Swain, P.; Baig, M.J.; Kumar, A. A Chloroplast Glycolate Catabolic Pathway Bypassing the Endogenous Photorespiratory Cycle Enhances Photosynthesis, Biomass and Yield in Rice (Oryza sativa L.). Plant Sci. 2022, 314, 111103. [Google Scholar] [CrossRef] [PubMed]
- Lal, M.K.; Tiwari, R.K.; Gahlaut, V.; Mangal, V.; Kumar, A.; Singh, M.P.; Paul, V.; Kumar, S.; Singh, B.; Zinta, G. Physiological and Molecular Insights on Wheat Responses to Heat Stress. Plant Cell Rep. 2022, 41, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Lal, M.K.; Dutt, S.; Raigond, P.; Changan, S.S.; Tiwari, R.K.; Chourasia, K.N.; Mangal, V.; Singh, B. Functional Fermented Probiotics, Prebiotics, and Synbiotics from Non-Dairy Products: A Perspective from Nutraceutical. Mol. Nutr. Food Res. 2022, 66, 2101059. [Google Scholar] [CrossRef]
- Lal, M.K.; Vengavasi, K.; Pandey, R. Interactive Effects of Low Phosphorus and Elevated CO2 on Root Exudation and Nutrient Uptake in Wheat Is Modified under Sulphur Nutrition. Plant Physiol. Rep. 2019, 24, 63–73. [Google Scholar] [CrossRef]
- Yadav, M.R.; Kumar, R.; Parihar, C.M.; Yadav, R.K.; Jat, S.L.; Ram, H.; Meena, R.K.; Singh, M.; Birbal; Verma, A.P.; et al. Strategies for Improving Nitrogen Use Efficiency: A Review. Agric. Rev. 2017, 38, 29–40. [Google Scholar] [CrossRef]
- Siddiquee, S.; Rovina, K.; Azad, S.A.; Naher, L.; Suryani, S.; Chaikaew, P. Heavy Metal Contaminants Removal from Wastewater Using the Potential Filamentous Fungi Biomass: A Review. J. Microb. Biochem. Technol. 2015, 7, 384–393. [Google Scholar] [CrossRef]
- Biswas, S.S.; Ghosh, A.; Singhal, S.K.; Biswas, D.R.; Roy, T.; Sarkar, A.; Das, D. Phosphorus Enriched Organic Amendments Can Increase Nitrogen Use Efficiency in Wheat. Commun. Soil Sci. Plant Anal. 2019, 50, 1178–1191. [Google Scholar] [CrossRef]
- Chivenge, P.; Sharma, S.; Bunquin, M.A.; Hellin, J. Improving Nitrogen Use Efficiency—A Key for Sustainable Rice Production Systems. Front. Sustain. Food Syst. 2021, 5, 400. [Google Scholar] [CrossRef]
- Neeraja, C.N.; Voleti, S.R.; Desiraju, S.; Kuchi, S.; Bej, S.; Talapanti, K.; Puskur, R.R. Molecular Breeding for Improving Nitrogen Use Efficiency in Rice: Progress and Perspectives. In Molecular Breeding for Rice Abiotic Stress Tolerance and Nutritional Quality; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; pp. 234–248. [Google Scholar] [CrossRef]
- Metting, F.B. Algae and Cyanobacteria. Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties; Soil Science Society of America, Inc.: Madison, WI, USA, 2018; pp. 427–458. [Google Scholar] [CrossRef]
- Farzadfar, S.; Knight, J.D.; Congreves, K.A. Soil Organic Nitrogen: An Overlooked but Potentially Significant Contribution to Crop Nutrition. Plant Soil 2021, 462, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.L.; Kidwell, K.K.; McCracken, V.A.; Bolton, R.P.; Allen, M. Economically Optimal Wheat Yield, Protein and Nitrogen Use Component Responses to Varying N Supply and Genotype. Front. Plant Sci. 2020, 10, 1790. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.; Priya, N.; Ramanathan, A.L. Impacts of Anthropogenic Perturbations on Reactive Nitrogen Dynamics in Mangrove Ecosystem: Climate Change Perspective. J. Clim. Chang. 2019, 5, 9–21. [Google Scholar] [CrossRef]
- More, S.J.; Bardhan, K.; Ravi, V.; Pasala, R.; Chaturvedi, A.K.; Lal, M.K.; Siddique, K.H.M. Morphophysiological Responses and Tolerance Mechanisms in Cassava (Manihot esculenta Crantz) Under Drought Stress. J. Soil Sci. Plant Nutr. 2023, 2023, 1–21. [Google Scholar] [CrossRef]
- Altaf, M.A.; Mandal, S.; Behera, B.; Mangal, V.; Naz, S.; Kumar, R.; Kumar, A.; Ghorai, M.; Singh, B.; Dey, A.; et al. Salinity Stress Tolerance in Solanaceous Crops: Current Understanding and Its Prospects in Genome Editing. J. Plant Growth Regul. 2022, 1–17. [Google Scholar] [CrossRef]
- Yadav, M.R.; Kumar, S.; Behera, B.; Yadav, V.P.; Khrub, A.S.; Yadav, L.R.; Gupta, K.C.; Meena, O.P.; Baloda, A.S.; Raza, M.B.; et al. Energy-Carbon Footprint, Productivity and Profitability of Barley Cultivars under Contrasting Tillage-Residue Managements in Semi-Arid Plains of North-West India. J. Soil Sci. Plant Nutr. 2022, 1–16. [Google Scholar] [CrossRef]
- St. Luce, M.; Whalen, J.K.; Ziadi, N.; Zebarth, B.J. Nitrogen Dynamics and Indices to Predict Soil Nitrogen Supply in Humid Temperate Soils. Adv. Agron. 2011, 112, 55–102. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C. Nitrate Leaching in Temperate Agroecosystems: Sources, Factors and Mitigating Strategies. Nutr. Cycl. Agroecosyst. 2002, 64, 237–256. [Google Scholar] [CrossRef]
- Sharma, E.; Lal, M.K.; Gulati, A. Targeted UHPLC-QTOF-IMS Based Metabolite Profiling for Bioactive Compounds in Rosa Webbiana Wallich Ex Royle: An Unexploited Native from Western Himalayas. Plant Physiol. Biochem. 2023, 195, 58–66. [Google Scholar] [CrossRef]
- Mukherjee, P.; Suriyakumar, P.; Vanchinathan, S.; Krishnan, V.; Lal, M.K.; Jha, P.K.; Chinnusamy, V.; Anand, A.; Prasad, P.V.V. Hydrogen Peroxide and GA3 Levels Regulate the High Night Temperature Response in Pistils of Wheat (Triticum aestivum L.). Antioxidants 2023, 12, 342. [Google Scholar] [CrossRef]
- Li, X.; Song, B.; Yin, D.; Lal, M.K.; Riaz, M.; Song, X.; Huang, W. Influence of Biochar on Soil Properties and Morphophysiology of Sugar Beet Under Fomesafen Residues. J. Soil Sci. Plant Nutr. 2023, 1–14. [Google Scholar] [CrossRef]
- FAOSTAT FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 14 March 2019).
- Ladha, J.K.; Tirol-Padre, A.; Reddy, C.K.; Cassman, K.G.; Verma, S.; Powlson, D.S.; Van Kessel, C.; De Richter, D.B.; Chakraborty, D.; Pathak, H. Global Nitrogen Budgets in Cereals: A 50-Year Assessment for Maize, Rice, and Wheat Production Systems. Sci. Rep. 2016, 6, 19355. [Google Scholar] [CrossRef] [PubMed]
- Ladha, J.K.; Pathak, H.; Krupnik, T.J.; Six, J.; van Kessel, C. Efficiency of Fertilizer Nitrogen in Cereal Production: Retrospects and Prospects. Adv. Agron. 2005, 87, 85–156. [Google Scholar] [CrossRef]
- Impa, S.M.; Raju, B.; Hein, N.T.; Sandhu, J.; Prasad, P.V.V.; Walia, H.; Jagadish, S.V.K. High Night Temperature Effects on Wheat and Rice: Current Status and Way Forward. Plant Cell Environ. 2021, 44, 2049–2065. [Google Scholar] [CrossRef]
- Wang, X.; Hu, M.; Ren, H.; Li, J.; Tong, C.; Musenze, R.S. Seasonal Variations of Nitrous Oxide Fluxes and Soil Denitrification Rates in Subtropical Freshwater and Brackish Tidal Marshes of the Min River Estuary. Sci. Total Environ. 2018, 616–617, 1404–1413. [Google Scholar] [CrossRef]
- Lemarchand, D.; Gaillardet, J.; Lewin; Allégre, C.J. The Influence of Rivers on Marine Boron Isotopes and Implications for Reconstructing Past Ocean PH. Nature 2000, 408, 951–954. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, J. Investigating the Effects of Point Source and Nonpoint Source Pollution on the Water Quality of the East River (Dongjiang) in South China. Ecol. Indic. 2013, 32, 294–304. [Google Scholar] [CrossRef]
- Norton, R.; Davidson, E.; TL, R. Nitrogen Use Efficiency and Nutrient Performance Indicators. Glob. Partnersh. Nutr. Manag. 2015, 14, 15. [Google Scholar]
- Fageria, N.K.; Baligar, V.C. Enhancing Nitrogen Use Efficiency in Crop Plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar] [CrossRef]
- Leip, A.; Britz, W.; Weiss, F.; de Vries, W. Farm, Land, and Soil Nitrogen Budgets for Agriculture in Europe Calculated with CAPRI. Environ. Pollut. 2011, 159, 3243–3253. [Google Scholar] [CrossRef]
- San Martín, W. Global Nitrogen in Sustainable Development: Four Challenges at the Interface of Science and Policy. In Life on Land; Springer: Cham, Switzerland, 2021; pp. 485–499. [Google Scholar] [CrossRef]
- Wu, L.; Peng, M.; Qiao, S.; Ma, X. Assessing Impacts of Rainfall Intensity and Slope on Dissolved and Adsorbed Nitrogen Loss under Bare Loessial Soil by Simulated Rainfalls. Catena 2018, 170, 51–63. [Google Scholar] [CrossRef]
- Mandal, S.; Thangarajan, R.; Bolan, N.S.; Sarkar, B.; Khan, N.; Ok, Y.S.; Naidu, R. Biochar-Induced Concomitant Decrease in Ammonia Volatilization and Increase in Nitrogen Use Efficiency by Wheat. Chemosphere 2016, 142, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, P.; Dong, S.; Zhang, J.; Zhao, B. Effects of Fertilizer Type and Rate on Summer Maize Grain Yield and Ammonia Volatilization Loss in Northern China. J. Soils Sediments 2019, 19, 2200–2211. [Google Scholar] [CrossRef]
- Janz, B.; Havermann, F.; Lashermes, G.; Zuazo, P.; Engelsberger, F.; Torabi, S.M.; Butterbach-Bahl, K. Effects of Crop Residue Incorporation and Properties on Combined Soil Gaseous N2O, NO, and NH3 Emissions—A Laboratory-Based Measurement Approach. Sci. Total Environ. 2022, 807, 151051. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Baluska, F.; Fujita, M. Nitric Oxide-Induced Salt Stress Tolerance in Plants: ROS Metabolism, Signaling, and Molecular Interactions. Plant Biotechnol. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Rout, P.R.; Shahid, M.K.; Dash, R.R.; Bhunia, P.; Liu, D.; Varjani, S.; Zhang, T.C.; Surampalli, R.Y. Nutrient Removal from Domestic Wastewater: A Comprehensive Review on Conventional and Advanced Technologies. J. Environ. Manag. 2021, 296, 113246. [Google Scholar] [CrossRef]
- Liu, R.; Suter, H.; He, J.; Hayden, H.; Chen, D. Influence of Temperature and Moisture on the Relative Contributions of Heterotrophic and Autotrophic Nitrification to Gross Nitrification in an Acid Cropping Soil. J. Soils Sediments 2015, 15, 2304–2309. [Google Scholar] [CrossRef]
- Pramanick, B.; Kumar, M.; Naik, B.M.; Kumar, M.; Singh, S.K.; Maitra, S.; Naik, B.S.S.S.; Rajput, V.D.; Minkina, T. Long-Term Conservation Tillage and Precision Nutrient Management in Maize–Wheat Cropping System: Effect on Soil Properties, Crop Production, and Economics. Agronomy 2022, 12, 2766. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous Oxide Emissions from Soils: How Well Do We Understand the Processes and Their Controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Bibi, S.; Saifullah, S.; Naeem, A.; Dahlawi, S. Environmental Impacts of Nitrogen Use in Agriculture, Nitrate Leaching and Mitigation Strategies. In Soil Science: Agricultural and Environmental Prospectives; Springer: Cham, Switzerland, 2016; pp. 131–157. [Google Scholar] [CrossRef]
- Bijay-Singh; Craswell, E. Fertilizers and Nitrate Pollution of Surface and Ground Water: An Increasingly Pervasive Global Problem. SN Appl. Sci. 2021, 3, 518. [Google Scholar] [CrossRef]
- Silva, R.G.; Cameron, K.C.; Di, H.J.; Smith, N.P.; Buchan, G.D. Effect of Macropore Flow on the Transport of Surface-Applied Cow Urine through a Soil Profile. Aust. J. Soil Res. 2000, 38, 13–23. [Google Scholar] [CrossRef]
- Brender, J.D.; Weyer, P.J.; Romitti, P.A.; Mohanty, B.P.; Shinde, M.U.; Vuong, A.M.; Sharkey, J.R.; Dwivedi, D.; Horel, S.A.; Kantamneni, J.; et al. Prenatal Nitrate Intake from Drinking Water and Selected Birth Defects in Offspring of Participants in the National Birth Defects Prevention Study. Environ. Health Perspect. 2013, 121, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Watt, W.; Tulinsky, A.; Swenson, R.P.; Watenpaugh, K.D. Comparison of the Crystal Structures of a Flavodoxin in Its Three Oxidation States at Cryogenic Temperatures. J. Mol. Biol. 1991, 218, 195–208. [Google Scholar] [CrossRef]
- Mahmud, K.; Panday, D.; Mergoum, A.; Missaoui, A. Nitrogen Losses and Potential Mitigation Strategies for a Sustainable Agroecosystem. Sustainability 2021, 13, 2400. [Google Scholar] [CrossRef]
- Singh, S.K.; Patra, A.; Chand, R.; Jatav, H.S.; Luo, Y.; Rajput, V.D.; Sehar, S.; Attar, S.K.; Khan, M.A.; Jatav, S.S.; et al. Surface Seeding of Wheat: A Sustainable Way towards Climate Resilience Agriculture. Sustainability 2022, 14, 7460. [Google Scholar] [CrossRef]
- Zhang, M.; Tian, Y.; Zhao, M.; Yin, B.; Zhu, Z. The Assessment of Nitrate Leaching in a Rice–Wheat Rotation System Using an Improved Agronomic Practice Aimed to Increase Rice Crop Yields. Agric. Ecosyst. Environ. 2017, 241, 100–109. [Google Scholar] [CrossRef]
- de Vries, W. Impacts of Nitrogen Emissions on Ecosystems and Human Health: A Mini Review. Curr. Opin. Environ. Sci. Health 2021, 21, 100249. [Google Scholar] [CrossRef]
- Pratt, P.F. Nitrogen Use and Nitrate Leaching in Irrigated Agriculture. In Nitrogen in Crop Production; American Society of Agronomy, Inc.; Crop Science Society of America, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 2015; pp. 319–333. [Google Scholar] [CrossRef]
- Nakagawa, K.; Amano, H.; Asakura, H.; Berndtsson, R. Spatial Trends of Nitrate Pollution and Groundwater Chemistry in Shimabara, Nagasaki, Japan. Environ. Earth Sci. 2016, 75, 234. [Google Scholar] [CrossRef]
- Kováčová, V. Assessment of Surface Water Eutrophication at ŽitnÝ Ostrov Region. Acta Hydrol. Slovaca 2020, 21, 65–73. [Google Scholar] [CrossRef]
- Ferreira, J.G.; Andersen, J.H.; Borja, A.; Bricker, S.B.; Camp, J.; Cardoso da Silva, M.; Garcés, E.; Heiskanen, A.S.; Humborg, C.; Ignatiades, L.; et al. Overview of Eutrophication Indicators to Assess Environmental Status within the European Marine Strategy Framework Directive. Estuar. Coast. Shelf Sci. 2011, 93, 117–131. [Google Scholar] [CrossRef]
- Zhang, C.H.; Guo, H.R.; Huang, H.; Ma, T.Y.; Song, W.; Chen, C.J.; Liu, X.Y. Atmospheric Nitrogen Deposition and Its Responses to Anthropogenic Emissions in a Global Hotspot Region. Atmos. Res. 2021, 248, 105137. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.; Kong, X.; Hou, D.; Gu, J.; Liu, L.; Wang, Z.; Yang, J. Progressive Integrative Crop Managements Increase Grain Yield, Nitrogen Use Efficiency and Irrigation Water Productivity in Rice. Field Crops Res. 2018, 215, 1–11. [Google Scholar] [CrossRef]
- Gong, Y.; Wu, J.; Vogt, J.; Le, T.B. Warming Reduces the Increase in N2O Emission under Nitrogen Fertilization in a Boreal Peatland. Sci. Total Environ. 2019, 664, 72–78. [Google Scholar] [CrossRef]
- Vermeulen, I.; Block, C.; Vandecasteele, C. Estimation of Fuel-Nitrogen Oxide Emissions from the Element Composition of the Solid or Waste Fuel. Fuel 2012, 94, 75–80. [Google Scholar] [CrossRef]
- Bellarby, J.; Foereid, B.; Hastings, A.F.S.J.; Smith, P. Cool Farming: Climate Impacts of Agriculture and Mitigation Potential; Greenpeace International: Amsterdam, The Netherlands, 2008; 44p. [Google Scholar]
- Mushtaq, S.; Maraseni, T.N.; Reardon-Smith, K. Climate Change and Water Security: Estimating the Greenhouse Gas Costs of Achieving Water Security through Investments in Modern Irrigation Technology. Agric. Syst. 2013, 117, 78–89. [Google Scholar] [CrossRef]
- Yan, M.; Luo, T.; Bian, R.; Cheng, K.; Pan, G.; Rees, R. A Comparative Study on Carbon Footprint of Rice Production between Household and Aggregated Farms from Jiangxi, China. Environ. Monit. Assess 2015, 187, 332. [Google Scholar] [CrossRef] [PubMed]
- Maheswari, M.; Murthy, A.N.G.; Shanker, A.K. Nitrogen Nutrition in Crops and Its Importance in Crop Quality. In The Indian Nitrogen Assessment: Sources of Reactive Nitrogen, Environmental and Climate Effects, Management Options, and Policies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 175–186. [Google Scholar] [CrossRef]
- Kumar, A.; Mishra, S.; Bakshi, S.; Upadhyay, P.; Thakur, T.K. Response of Eutrophication and Water Quality Drivers on Greenhouse Gas Emissions in Lakes of China: A Critical Analysis. Ecohydrology 2023, 16, e2483. [Google Scholar] [CrossRef]
- Naz, S.; Bilal, A.; Saddiq, B.; Ejaz, S.; Ali, S.; Tul, S.; Haider, A.; Sardar, H.; Nasir, B.; Ahmad, I.; et al. Foliar Application of Salicylic Acid Improved Growth, Yield, Quality and Photosynthesis of Pea (Pisum sativum L.) by Improving Antioxidant Defense Mechanism under Saline Conditions. Sustainability 2022, 14, 14180. [Google Scholar] [CrossRef]
- Altaf, M.A.; Behera, B.; Mangal, V.; Singhal, R.K.; Kumar, R.; More, S.; Naz, S.; Mandal, S.; Dey, A.; Saqib, M.; et al. Tolerance and Adaptation Mechanism of Solanaceous Crops under Salinity Stress. Funct. Plant Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity Responses and Tolerance Mechanisms in Underground Vegetable Crops: An Integrative Review. Planta 2022, 255, 68. [Google Scholar] [CrossRef]
- Mangal, V.; Lal, M.K.; Tiwari, R.K.; Altaf, M.A.; Sood, S.; Kumar, D.; Bharadwaj, V.; Singh, B.; Singh, R.K.; Aftab, T. Molecular Insights into the Role of Reactive Oxygen, Nitrogen and Sulphur Species in Conferring Salinity Stress Tolerance in Plants. J. Plant Growth Regul. 2022, 42, 554–574. [Google Scholar] [CrossRef]
- Nadarajan, S.; Sukumaran, S. Chemistry and Toxicology behind Chemical Fertilizers. In Controlled Release Fertilizers for Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2021; pp. 195–229. [Google Scholar] [CrossRef]
- Siman, F.C.; Andrade, F.V.; Passos, R.R.; Littig, M.; de Oliveira Machado, F. Nitrogen Fertilizers and Volatilization of Ammonia, Carbonic Gas Emissions and Urease Activity. Commun. Soil Sci. Plant Anal. 2022, 53, 426–438. [Google Scholar] [CrossRef]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen Losses from the Soil/Plant System: A Review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Boczulak, S.A. The Effect of Soil Temperature on Soil Nitrogen form Availability and Nitrogen Uptake by Conifers of British Columbia. Ph.D. Dissertation, University of Victoria, Victoria, BC, Canada, 2013. [Google Scholar]
- Spackman, J.A.; Fernandez, F.G.; Coulter, J.A.; Kaiser, D.E.; Paiao, G. Soil Texture and Precipitation Influence Optimal Time of Nitrogen Fertilization for Corn. Agron. J. 2019, 111, 2018–2030. [Google Scholar] [CrossRef]
- Rakhsh, F.; Golchin, A.; Beheshti Al Agha, A.; Nelson, P.N. Mineralization of Organic Carbon and Formation of Microbial Biomass in Soil: Effects of Clay Content and Composition and the Mechanisms Involved. Soil Biol. Biochem. 2020, 151, 108036. [Google Scholar] [CrossRef]
- Whitehead, D.C.; Raistrick, N. The Volatilization of Ammonia from Cattle Urine Applied to Soils as Influenced by Soil Properties. Plant Soil 1993, 148, 43–51. [Google Scholar] [CrossRef]
- He, Z.L.; Calvert, D.v.; Alva, A.K.; Li, Y.C.; Banks, D.J. Clinoptilolite Zeolite and Cellulose Amendments to Reduce Ammonia Volatilization in a Calcareous Sandy Soil. Plant Soil 2002, 247, 253–260. [Google Scholar] [CrossRef]
- Chen, S.; Liu, S.; Zheng, X.; Yin, M.; Chu, G.; Xu, C.; Yan, J.; Chen, L.; Wang, D.; Zhang, X. Effect of Various Crop Rotations on Rice Yield and Nitrogen Use Efficiency in Paddy–Upland Systems in Southeastern China. Crop J. 2018, 6, 576–588. [Google Scholar] [CrossRef]
- Tokuchi, N.; Hirobe, M.; Kondo, K.; Arai, H.; Hobara, S.; Fukushima, K.; Matsuura, Y. Soil Nitrogen Dynamics in Larch Ecosystem. In Permafrost Ecosystems; Elsevier: Amsterdam, The Netherlands, 2010; pp. 229–243. [Google Scholar] [CrossRef]
- Seibert, J.; Stendahl, J.; Sørensen, R. Topographical Influences on Soil Properties in Boreal Forests. Geoderma 2007, 141, 139–148. [Google Scholar] [CrossRef]
- Chaves, J.; Neill, C.; Germer, S.; Gouveia Neto, S.; Krusche, A.V.; Castellanos Bonilla, A.; Elsenbeer, H. Nitrogen Transformations in Flowpaths Leading from Soils to Streams in Amazon Forest and Pasture. Ecosystems 2009, 12, 961–972. [Google Scholar] [CrossRef]
- Cai, A.; Xu, H.; Shao, X.; Zhu, P.; Zhang, W.; Xu, M.; Murphy, D.V. Carbon and Nitrogen Mineralization in Relation to Soil Particle-Size Fractions after 32 Years of Chemical and Manure Application in a Continuous Maize Cropping System. PLoS ONE 2016, 11, e0152521. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Li, L.; Han, X. Seasonal Variations in Nitrogen Mineralization under Three Land Use Types in a Grassland Landscape. Acta Oecologica 2008, 34, 322–330. [Google Scholar] [CrossRef]
- Jha, P.; Hati, K.M.; Dalal, R.C.; Dang, Y.P.; Kopittke, P.M.; McKenna, B.A.; Menzies, N.W. Effect of 50 Years of No-Tillage, Stubble Retention, and Nitrogen Fertilization on Soil Respiration, Easily Extractable Glomalin, and Nitrogen Mineralization. Agronomy 2022, 12, 151. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Lal, M.K.; Tiwari, R.K.; Shakoor, A. Melatonin Mitigates Cadmium Toxicity by Promoting Root Architecture and Mineral Homeostasis of Tomato Genotypes. J. Soil Sci. Plant Nutr. 2022, 22, 1112–1128. [Google Scholar] [CrossRef]
- Jaiswal, S.; Kumar, A.; Choudhury, B.U.; Thangam, R.; Lal, M.K.; Shettigar, N.; Kumar, R.; Verma, H.; Bhattacharjee, B.; Mishra, V.K. Development of Ammonium Nitrate Free Nutrient Media for Aluminium Toxicity Tolerance Screening of Rice Genotypes from North-Eastern India. J. Plant Nutr. 2022. [Google Scholar] [CrossRef]
- Behera, B.; Kancheti, M.; Raza, M.B.; Shiv, A.; Mangal, V.; Rathod, G.; Altaf, M.A.; Kumar, A.; Aftab, T.; Kumar, R.; et al. Mechanistic Insight on Boron-Mediated Toxicity in Plant Vis-a-Vis Its Mitigation Strategies: A Review. Int. J. Phytoremediation 2023, 25, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Amanullah; Khalid, S.; Imran; Khan, H.A.; Arif, M.; Altawaha, A.R.; Adnan, M.; Fahad, S.; Parmar, B. Organic Matter Management in Cereals Based System: Symbiosis for Improving Crop Productivity and Soil Health. In Sustainable Agriculture Reviews 29; Springer: Cham, Switzerland, 2019; pp. 67–92. [Google Scholar] [CrossRef]
- Ryals, R.; Silver, W.L. Effects of Organic Matter Amendments on Net Primary Productivity and Greenhouse Gas Emissions in Annual Grasslands. Ecol. Appl. 2013, 23, 46–59. [Google Scholar] [CrossRef]
- Fu, B.; Chen, L.; Huang, H.; Qu, P.; Wei, Z. Impacts of Crop Residues on Soil Health: A Review. Environ. Pollut. Bioavailab. 2021, 33, 164–173. [Google Scholar] [CrossRef]
- Gentile, R.; Vanlauwe, B.; van Kessel, C.; Six, J. Managing N Availability and Losses by Combining Fertilizer-N with Different Quality Residues in Kenya. Agric. Ecosyst. Environ. 2009, 131, 308–314. [Google Scholar] [CrossRef]
- Barker, D.W.; Sawyer, J.E. Using Active Canopy Sensing to Adjust Nitrogen Application Rate in Corn. Agron. J. 2012, 104, 926–933. [Google Scholar] [CrossRef]
- Holland, K.H.; Schepers, J.S. Derivation of a Variable Rate Nitrogen Application Model for In-Season Fertilization of Corn. Agron. J. 2010, 102, 1415–1424. [Google Scholar] [CrossRef]
- Thompson, L.J.; Ferguson, R.B.; Kitchen, N.; Frazen, D.W.; Mamo, M.; Yang, H.; Schepers, J.S. Model and Sensor-Based Recommendation Approaches for in-Season Nitrogen Management in Corn. Agron. J. 2015, 107, 2020–2030. [Google Scholar] [CrossRef]
- Stamatiadis, S.; Schepers, J.S.; Evangelou, E.; Tsadilas, C.; Glampedakis, A.; Glampedakis, M.; Dercas, N.; Spyropoulos, N.; Dalezios, N.R.; Eskridge, K. Variable-Rate Nitrogen Fertilization of Winter Wheat under High Spatial Resolution. Precis. Agric. 2018, 19, 570–587. [Google Scholar] [CrossRef]
- Samborski, S.M.; Gozdowski, D.; Stepień, M.; Walsh, O.S.; Leszczyńska, E. On-Farm Evaluation of an Active Optical Sensor Performance for Variable Nitrogen Application in Winter Wheat. Eur. J. Agron. 2016, 74, 56–67. [Google Scholar] [CrossRef]
- Lukina, E.V.; Freeman, K.W.; Wynn, K.J.; Thomason, W.E.; Mullen, R.W.; Stone, M.L.; Solie, J.B.; Klatt, A.R.; Johnson, G.v.; Elliott, R.L.; et al. NITROGEN FERTILIZATION OPTIMIZATION ALGORITHM BASED ON IN-SEASON ESTIMATES OF YIELD AND PLANT NITROGEN UPTAKE. J. Plant Nutr. 2007, 24, 885–898. [Google Scholar] [CrossRef]
- Cao, Q.; Miao, Y.; Li, F.; Gao, X.; Liu, B.; Lu, D.; Chen, X. Developing a New Crop Circle Active Canopy Sensor-Based Precision Nitrogen Management Strategy for Winter Wheat in North China Plain. Precis. Agric. 2017, 18, 2–18. [Google Scholar] [CrossRef]
- Bausch, W.C.; Diker, K. Innovative Remote Sensing Techniques to Increase Nitrogen Use Efficiency of Corn. Commun. Soil Sci. Plant Anal. 2001, 32, 1371–1390. [Google Scholar] [CrossRef]
- Raun, W.R.; Solie, J.B.; Stone, M.L.; Martin, K.L.; Freeman, K.W.; Mullen, R.W.; Zhang, H.; Schepers, J.S.; Johnson, G.V. Optical Sensor-Based Algorithm for Crop Nitrogen Fertilization. Commun. Soil Sci. Plant Anal. 2007, 36, 2759–2781. [Google Scholar] [CrossRef]
- Raun, W.R.; Solie, J.B.; Johnson, G.V.; Stone, M.L.; Mutten, R.W.; Freeman, K.W.; Thomason, W.E.; Lukina, E.v. Improving Nitrogen Use Efficiency in Cereal Grain Production with Optical Sensing and Variable Rate Application. Agron. J. 2002, 94, 815–820. [Google Scholar] [CrossRef]
- Yao, Y.; Miao, Y.; Huang, S.; Gao, L.; Ma, X.; Zhao, G.; Jiang, R.; Chen, X.; Zhang, F.; Yu, K.; et al. Active Canopy Sensor-Based Precision N Management Strategy for Rice. Agron. Sustain. Dev. 2012, 32, 925–933. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Majumdar, K.; Jat, M.L.; Kumar, A.; Bishnoi, D.K.; McDonald, A.J.; Pampolino, M. Precision Nutrient Management in Conservation Agriculture Based Wheat Production of Northwest India: Profitability, Nutrient Use Efficiency and Environmental Footprint. Field Crops Res. 2014, 155, 233–244. [Google Scholar] [CrossRef]
- Ortiz-Monasterio, J.I.; Raun, W. Paper Presented at International Workshop on Increasing Wheat Yield Potential, CIMMYT, Obregon, Mexico, 20-24 March 2006: Reduced Nitrogen and Improved Farm Income for Irrigated Spring Wheat in the Yaqui Valley, Mexico, Using Sensor Based Nitrogen Managem. J. Agric. Sci. 2007, 145, 215–222. [Google Scholar] [CrossRef]
- Butchee, K.S.; May, J.; Arnall, B. Sensor Based Nitrogen Management Reduced Nitrogen and Maintained Yield. Crop Manag. 2011, 10, 1–5. [Google Scholar] [CrossRef]
- Shakoor, A.; Shahbaz, M.; Farooq, T.H.; Sahar, N.E.; Shahzad, S.M.; Altaf, M.M.; Ashraf, M. A Global Meta-Analysis of Greenhouse Gases Emission and Crop Yield under No-Tillage as Compared to Conventional Tillage. Sci. Total Environ. 2021, 750, 142299. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Islam, A.M.; Azirun, S.M.; Boyce, A.N. Tropical Legume Crop Rotation and Nitrogen Fertilizer Effects on Agronomic and Nitrogen Efficiency of Rice. Sci. World J. 2014, 2014, 490841. [Google Scholar] [CrossRef]
- Yang, T.; Lupwayi, N.; Marc, S.A.; Siddique, K.H.M.; Bainard, L.D. Anthropogenic Drivers of Soil Microbial Communities and Impacts on Soil Biological Functions in Agroecosystems. Glob. Ecol. Conserv. 2021, 27, e01521. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Hämmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of Microbial Nitrogen Use Efficiency to Carbon:Nitrogen Imbalances Regulates Soil Nitrogen Cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef]
- Zhou, S.; Xiang, Y.; Tie, L.; Han, B.; Huang, C. Simulated Nitrogen Deposition Significantly Reduces Soil Respiration in an Evergreen Broadleaf Forest in Western China. PLoS ONE 2018, 13, e0204661. [Google Scholar] [CrossRef]
- Zhu, T.; Meng, T.; Zhang, J.; Yin, Y.; Cai, Z.; Yang, W.; Zhong, W. Nitrogen Mineralization, Immobilization Turnover, Heterotrophic Nitrification, and Microbial Groups in Acid Forest Soils of Subtropical China. Biol. Fertil. Soils 2013, 49, 323–331. [Google Scholar] [CrossRef]
- Lama, S.; Velescu, A.; Leimer, S.; Weigelt, A.; Chen, H.; Eisenhauer, N.; Scheu, S.; Oelmann, Y.; Wilcke, W. Plant Diversity Influenced Gross Nitrogen Mineralization, Microbial Ammonium Consumption and Gross Inorganic N Immobilization in a Grassland Experiment. Oecologia 2020, 193, 731–748. [Google Scholar] [CrossRef] [PubMed]
- Buckeridge, K.M.; Zufelt, E.; Chu, H.; Grogan, P. Soil Nitrogen Cycling Rates in Low Arctic Shrub Tundra Are Enhanced by Litter Feedbacks. Plant Soil 2010, 330, 407–421. [Google Scholar] [CrossRef]
- Ren, W.; Chen, F.S.; Hu, X.F.; Yu, M.Q.; Feng, X. Soil Nitrogen Transformations Varied with Plant Community under Nanchang Urban Forests in Mid-Subtropical Zone of China. J. For. Res. 2011, 22, 569–576. [Google Scholar] [CrossRef]
- Yang, X.; Xia, X.; Zhang, Z.; Nong, B.; Zeng, Y.; Xiong, F.; Wu, Y.; Gao, J.; Deng, G.; Li, D. QTL Mapping by Whole Genome Re-Sequencing and Analysis of Candidate Genes for Nitrogen Use Efficiency in Rice. Front. Plant Sci. 2017, 8, 1634. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, N.; Sethi, M.; Kumar, A.; Dang, D.; Singh, J.; Chhuneja, P. Biochemical and Genetic Approaches Improving Nitrogen Use Efficiency in Cereal Crops: A Review. Front. Plant Sci. 2021, 12, 757. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Hawkesford, M.J.; Barraclough, P.B.; Holdsworth, M.J.; Kerr, S.; Kightley, S.; Shewry, P.R. Identifying Traits to Improve the Nitrogen Economy of Wheat: Recent Advances and Future Prospects. Field Crops Res. 2009, 114, 329–342. [Google Scholar] [CrossRef]
- Syed, M.A.; Alam, M.A.; Hossain, A.; Islam, M.R.; Vemuri, H.; Jahan, N. Rice Breeding and Genomics Approaches for Improving Water and Nitrogen Use Efficiency. In Rice Research for Quality Improvement: Genomics and Genetic Engineering; Springer: Singapore, 2020; pp. 339–372. [Google Scholar] [CrossRef]
- Cormier, F.; Foulkes, J.; Hirel, B.; Gouache, D.; Moënne-Loccoz, Y.; le Gouis, J. Breeding for Increased Nitrogen-Use Efficiency: A Review for Wheat (T. aestivum L.). Plant Breed. 2016, 135, 255–278. [Google Scholar] [CrossRef]
- Kamprath, E.J.; Moll, R.H.; Rodriguez, N. Effects of Nitrogen Fertilization and Recurrent Selection on Performance of Hybrid Populations of Corn 1. Agron. J. 1982, 74, 955–958. [Google Scholar] [CrossRef]
- Fiorani, F.; Schurr, U. Future Scenarios for Plant Phenotyping. Annu. Rev. Plant Biol. 2013, 64, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, A.; Nocito, F.F.; Morgutti, S.; Sacchi, G.A. Plant Breeding for Improving Nutrient Uptake and Utilization Efficiency. In Advances in Research on Fertilization Management of Vegetable Crops; Springer: Cham, Switzerland, 2017; pp. 221–246. [Google Scholar] [CrossRef]
- Anbessa, Y.; Juskiw, P. Review: Strategies to Increase Nitrogen Use Efficiency of Spring Barley. Can. J. Plant Sci. 2012, 92, 617–625. [Google Scholar] [CrossRef]
- Ertiro, B.T.; Olsen, M.; Das, B.; Gowda, M.; Labuschagne, M. Genetic Dissection of Grain Yield and Agronomic Traits in Maize under Optimum and Low-Nitrogen Stressed Environments. Int. J. Mol. Sci. 2020, 21, 543. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Nie, X.L.; Xiao, X.G. Over-Expression of a Tobacco Nitrate Reductase Gene in Wheat (Triticum aestivum L.) Increases Seed Protein Content and Weight without Augmenting Nitrogen Supplying. PLoS ONE 2013, 8, e74678. [Google Scholar] [CrossRef]
- Ranjan, R.; Yadav, R. Targeting Nitrogen Use Efficiency for Sustained Production of Cereal Crops. J. Plant Nutr. 2019, 42, 1086–1113. [Google Scholar] [CrossRef]
- Zhou, Y.; Tao, Y.; Tang, D.; Wang, J.; Zhong, J.; Wang, Y.; Yuan, Q.; Yu, X.; Zhang, Y.; Wang, Y.; et al. Identification of QTL Associated with Nitrogen Uptake and Nitrogen Use Efficiency Using High Throughput Genotyped CSSLs in Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 1166. [Google Scholar] [CrossRef] [PubMed]
- Yorobe, J.M.; Ali, J.; Pede, V.O.; Rejesus, R.M.; Velarde, O.P.; Wang, H. Yield and Income Effects of Rice Varieties with Tolerance of Multiple Abiotic Stresses: The Case of Green Super Rice (GSR) and Flooding in the Philippines. Agric. Econ. 2016, 47, 261–271. [Google Scholar] [CrossRef]
- Ali, J.; Jewel, Z.A.; Mahender, A.; Anandan, A.; Hernandez, J.; Li, Z. Molecular Genetics and Breeding for Nutrient Use Efficiency in Rice. Int. J. Mol. Sci. 2018, 19, 1762. [Google Scholar] [CrossRef] [PubMed]
- Lammerts Van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The Need to Breed Crop Varieties Suitable for Organic Farming, Using Wheat, Tomato and Broccoli as Examples: A Review. NJAS-Wagening. J. Life Sci. 2011, 58, 193–205. [Google Scholar] [CrossRef]
- Yadav, M.R.; Choudhary, M.; Singh, J.; Lal, M.K.; Jha, P.K.; Udawat, P.; Gupta, N.K.; Rajput, V.D.; Garg, N.K.; Maheshwari, C.; et al. Impacts, Tolerance, Adaptation, and Mitigation of Heat Stress on Wheat under Changing Climates. Int. J. Mol. Sci. 2022, 23, 2838. [Google Scholar] [CrossRef] [PubMed]
- Metzker, M.L. Sequencing Technologies the next Generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sandhu, N.; Dixit, S.; Yadav, S.; Swamy, B.P.M.; Shamsudin, N.A.A. Marker-Assisted Selection Strategy to Pyramid Two or More QTLs for Quantitative Trait-Grain Yield under Drought. Rice 2018, 11, 35. [Google Scholar] [CrossRef]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H.; Smith, K.P.; Sorrells, M.E.; Jannink, J.L. Genomic Selection in Plant Breeding. Knowledge and Prospects. Adv. Agron. 2011, 110, 77–123. [Google Scholar] [CrossRef]
- Liang, C.; Wang, Y.; Zhu, Y.; Tang, J.; Hu, B.; Liu, L.; Ou, S.; Wu, H.; Sun, X.; Chu, J.; et al. OsNAP Connects Abscisic Acid and Leaf Senescence by Fine-Tuning Abscisic Acid Biosynthesis and Directly Targeting Senescence-Associated Genes in Rice. Proc. Natl. Acad. Sci. USA 2014, 111, 10013–10018. [Google Scholar] [CrossRef]
- Li, M.; Xu, J.; Gao, Z.; Tian, H.; Gao, Y.; Kariman, K. Genetically Modified Crops Are Superior in Their Nitrogen Use Efficiency-A Meta-Analysis of Three Major Cereals. Sci. Rep. 2020, 10, 8568. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, H.; Liu, X.; Liu, Y.; Gao, S.; Zhou, J. The Effect of N Fertilizer Placement on the Fate of Urea-15N and Yield of Winter Wheat in Southeast China. PLoS ONE 2016, 11, e0153701. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Zhang, Z.; Chai, X.; Wang, H.; Wang, Y.; et al. Variation in NRT1.1B Contributes to Nitrate-Use Divergence between Rice Subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef]
- Kurai, T.; Wakayama, M.; Abiko, T.; Yanagisawa, S.; Aoki, N.; Ohsugi, R. Introduction of the ZmDof1 Gene into Rice Enhances Carbon and Nitrogen Assimilation under Low-Nitrogen Conditions. Plant Biotechnol. J. 2011, 9, 826–837. [Google Scholar] [CrossRef]
- Lam, H.M.; Wong, P.; Chan, H.K.; Yam, K.M.; Chen, L.; Chow, C.M.; Coruzzi, G.M. Overexpression of the ASN1 Gene Enhances Nitrogen Status in Seeds of Arabidopsis. Plant Physiol. 2003, 132, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, S.I.; Allen, D.J.; Ortiz-Lopez, A.; Hernández, G. Over-Expression of Cytosolic Glutamine Synthetase Increases Photosynthesis and Growth at Low Nitrogen Concentrations. J. Exp. Bot. 2001, 52, 1071–1081. [Google Scholar] [CrossRef]
- Mandal, S.; Ghorai, M.; Anand, U.; Roy, D.; Kant, N.; Mishra, T.; Mane, A.B.; Jha, N.K.; Lal, M.K.; Tiwari, R.K.; et al. Cytokinins: A Genetic Target for Increasing Yield Potential in the CRISPR Era. Front. Genet. 2022, 13, 26. [Google Scholar] [CrossRef]
- Kumar, R.; Kaundal, P.; Tiwari, R.K.; Kumar Lal, M.; Kumari, H.; Kumar, R.; Chandra Naga, K.; Kumar, A.; Singh, B.; Sagar, V.; et al. Development of Reverse Transcription Recombinase Polymerase Amplification (RT-RPA): A Methodology for Quick Diagnosis of Potato Leafroll Viral Disease in Potato. Int. J. Mol. Sci. 2023, 24, 2511. [Google Scholar] [CrossRef]
- Choudhary, C.S.; Behera, B.; Raza, M.B.; Mrunalini, K.; Bhoi, T.K.; Lal, M.K.; Nongmaithem, D.; Pradhan, S.; Song, B.; Das, T.K. Mechanisms of Allelopathic Interactions for Sustainable Weed Management. Rhizosphere 2023, 25, 100667. [Google Scholar] [CrossRef]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life 2021, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Khatodia, S.; Bhatotia, K.; Passricha, N.; Khurana, S.M.P.; Tuteja, N. The CRISPR/Cas Genome-Editing Tool: Application in Improvement of Crops. Front. Plant Sci. 2016, 7, 506. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Song, Q.; Li, C.; Jin, S.; Zhang, D.; Wang, Y.; Qiu, J.L.; Gao, C. Efficient C-to-t Base Editing in Plants Using a Fusion of Ncas9 and Human Apobec3a. Nat. Biotechnol. 2018, 36, 950. [Google Scholar] [CrossRef] [PubMed]
- Sathee, L.; Jagadhesan, B.; Pandesha, P.H.; Barman, D.; Adavi, B.S.; Nagar, S.; Krishna, G.K.; Tripathi, S.; Jha, S.K.; Chinnusamy, V. Genome Editing Targets for Improving Nutrient Use Efficiency and Nutrient Stress Adaptation. Front. Genet. 2022, 14, 1427. [Google Scholar] [CrossRef]
- Kant, S. Understanding Nitrate Uptake, Signaling and Remobilisation for Improving Plant Nitrogen Use Efficiency. Semin Cell Dev Biol 2018, 74, 89–96. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Liu, S.; Fan, X.; Zhao, L.; Song, M.; Fan, X.; Xu, G. Co-Overexpression of OsNAR2.1 and OsNRT2.3a Increased Agronomic Nitrogen Use Efficiency in Transgenic Rice Plants. Front. Plant Sci. 2020, 11, 1245. [Google Scholar] [CrossRef] [PubMed]
- Ranathunge, K.; El-Kereamy, A.; Gidda, S.; Bi, Y.M.; Rothstein, S.J. AMT1;1 Transgenic Rice Plants with Enhanced NH4+ Permeability Show Superior Growth and Higher Yield under Optimal and Suboptimal NH4+ Conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef]
- Srivastava, A.K. Integrated Nutrient Management in Citrus. In Advances in Citrus Nutrition; Springer: Dordrecht, The Netherlands, 2012; pp. 369–389. ISBN 9789400741713. [Google Scholar]
- Cruz, J.L.; Filho, M.A.C.; Coelho, E.F.; Dos Santos, A.A. Salinidade Reduz a Fotossíntese e o Índice de Colheita Da Mandioca (Manihot esculenta Crantz). Acta Sci. Agron. 2017, 39, 545–555. [Google Scholar] [CrossRef]
- Singh, V.K.; Shukla, A.K.; Singh, M.P.; Majumdar, K.; Mishra, R.P.; Rani, M.; Singh, S.K. Effect of Site-Specific Nutrient Management on Yield, Profit and Apparent Nutrient Balance under Pre-Dominant Cropping Systems of Upper Gangetic Plains. Artic. Indian J. Agric. Sci. 2015, 85, 43–51. [Google Scholar]
- Singh, R.; Singh, T.; Soni, R.L. Enhancement in the Productivity of Maize (Zea mays L.) through Integrated Balanced Nutrient Management in Banswara District. Ann. Agric. Res. 2012, 33, 14–16. [Google Scholar]
- Bijay-Singh; Ali, A.M. Using Hand-Held Chlorophyll Meters and Canopy Reflectance Sensors for Fertilizer Nitrogen Management in Cereals in Small Farms in Developing Countries. Sensors 2020, 20, 1127. [Google Scholar] [CrossRef]
- Ali, A.M.; Thind, H.S.; Sharma, S.; Yadvinder-Singh. Site-Specific Nitrogen Management in Dry Direct-Seeded Rice Using Chlorophyll Meter and Leaf Colour Chart. Pedosphere 2015, 25, 72–81. [Google Scholar] [CrossRef]
- Maiti, D.; Das, D.K.; Karak, T.; Banerjee, M. Management of Nitrogen through the Use of Leaf Color Chart (LCC) and Soil Plant Analysis Development (SPAD) or Chlorophyll Meter in Rice under Irrigated Ecosystem. Sci. World J. 2004, 4, 838–846. [Google Scholar] [CrossRef]
- Singh, B.; Singh, Y.; Ladha, J.K.; Bronson, K.F.; Balasubramanian, V.; Singh, J.; Khind, C.S. Chlorophyll Meter- and Leaf Color Chart-Based Nitrogen Management for Rice and Wheat in Northwestern India. Agron. J. 2002, 94, 821–829. [Google Scholar] [CrossRef]
- Jat, R.A.; Wani, S.P.; Sahrawat, K.L.; Singh, P.; Dhaka, S.R.; Dhaka, B.L. Recent Approaches in Nitrogen Management for Sustainable Agricultural Production and Eco-Safety. Arch. Agron. Soil Sci. 2012, 58, 1033–1060. [Google Scholar] [CrossRef]
- Kumar, D.; Patel, R.A.; Ramani, V.P.; Rathod, S.V. Evaluating Precision Nitrogen Management Practices in Terms of Yield, Nitrogen Use Efficiency and Nitrogen Loss Reduction in Maize Crop Under Indian Conditions. Int. J. Plant Prod. 2021, 15, 243–260. [Google Scholar] [CrossRef]
- Singh, J.; Singh, V.; Kaur, S. Precision Nitrogen Management Improves Grain Yield, Nitrogen Use Efficiency and Reduces Nitrous Oxide Emission from Soil in Spring Maize. J. Plant Nutr. 2020, 43, 2311–2321. [Google Scholar] [CrossRef]
- Shukla, A.K.; Singh, V.K.; Dwivedi, B.S.; Sharma, S.K.; Singh, Y. Nitrogen Use Efficiencies Using Leaf Colour Chart in Rice (Oryza sativa)-Wheat (Triticum aestivum) Cropping System. Indian J. Agric. Sci. 2006, 76, 651–656. [Google Scholar]
- Yang, W.H.; Peng, S.; Huang, J.; Sanico, A.L.; Buresh, R.J.; Witt, C. Using Leaf Color Charts to Estimate Leaf Nitrogen Status of Rice. Agron. J. 2003, 95, 212–217. [Google Scholar] [CrossRef]
- Shukla, A.K.; Ladha, J.K.; Singh, V.K.; Dwivedi, B.S.; Balasubramanian, V.; Gupta, R.K.; Sharma, S.K.; Singh, Y.; Pathak, H.; Pandey, P.S.; et al. Calibrating the Leaf Color Chart for Nitrogen Management in Different Genotypes of Rice and Wheat in a Systems Perspective. Agron. J. 2004, 96, 1606–1621. [Google Scholar] [CrossRef]
- Gupta, R.K.; Singh, V.; Singh, Y.; Singh, B.; Thind, H.S.; Kumar, A.; Vashistha, M. Need-Based Fertilizer Nitrogen Management Using Leaf Colour Chart in Hybrid Rice (Oryza sativa). Indian J. Agric. Sci. 2011, 81, 1153–1157. [Google Scholar]
- Ahmad, N.A.; Muttalib, M.F.A.; Uda, M.N.A.; Arsat, Z.A.; Abdullah, F.; Hashim, M.K.R.; Azizan, F.A.; Jusoh, M.F.; Kamaruzaman, S.R.S.; Nordin, A.A. Measurement of Leaf Chlorophyll Content in Harumanis Mango Cultivated in a Greenhouse Using SPAD Meter. Mater Today Proc. 2023. [Google Scholar] [CrossRef]
- Singh, V.; Pant, A.K.; Bahtnagar, A.; Bhatt, M. Evaluation of Nutrient Expert Based Fertilizer Recommendation for Growth, Yield and Nutrient Uptake of Maize Hybrids and Soil Properties in Maize-Wheat Cropping System in Mollisol. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3539–3550. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, R.K.; Kaur, J.; Jat, M.L.; Martin, K.L.; Singh, Y.; Singh, V.; Chandna, P.; Choudhary, O.P.; Gupta, R.K.; et al. Assessment of the Nitrogen Management Strategy Using an Optical Sensor for Irrigated Wheat. Agron. Sustain. Dev. 2011, 31, 589–603. [Google Scholar] [CrossRef]
- Appels, R.; Eversole, K.; Feuillet, C.; Keller, B.; Rogers, J.; Stein, N.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; Poland, J.; et al. Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Gupta, G.; Shrestha, A.; Shrestha, A.; Amgain, L.P. Evaluation of Different Nutrient Management Practice in Yield and Growth in Rice in Morang District. Adv. Plants Agric. Res. 2016, 6, 187–191. [Google Scholar] [CrossRef]
- Siscar-Lee, J.J.H.; Juliano, B.O.; Qureshi, R.H.; Akbar, M. Effect of Saline Soil on Grain Quality of Rices Differing in Salinity Tolerance. Plant Foods Hum. Nutr. 1990, 40, 31–36. [Google Scholar] [CrossRef]
- Budhathoki, S.; Amgain, L.P.; Subedi, S.; Iqbal, M.; Shrestha, N.; Aryal, S. Assessing Growth, Productivity and Profitability of Drought Tolerant Rice Using Nutrient Expert—Rice and Other Precision Fertilizer Management Practices in Lamjung, Nepal. Acta. Sci. Agric. 2018, 2, 153–158. [Google Scholar]
- Shrestha, S.; Amgain, L.P.; Subedi, R.; Shrestha, P.; Shahi, S. Productivity and Profitability Analysis of Old Aged Hybrid Rice Seedling Using Nutrient-Rice Expert and Other Precision Nutrient Management Practices at Lamjung, Nepal. Int. J. Appl. Sci. Biotechnol. 2018, 6, 232–237. [Google Scholar] [CrossRef]
- Pampolino, M.F.; Witt, C.; Pasuquin, J.M.; Johnston, A.; Fisher, M.J. Development Approach and Evaluation of the Nutrient Expert Software for Nutrient Management in Cereal Crops. Comput. Electron. Agric. 2012, 88, 103–110. [Google Scholar] [CrossRef]
- Satyanarayana, T.; Majumdar, K.; Pampolino, M.; Johnston, A.M.; Jat, M.L.; Kuchanur, P.; Sreelatha, D.; Sekhar, J.C.; Kumar, Y.; Maheswaran, R.; et al. Nutrient ExpertTM: A Tool to Optimize Nutrient Use and Improve Productivity of Maize. Better Crops-South Asia 2013, 97, 21–24. [Google Scholar]
- Majumdar, K.; Satyanarayana, T.; Dutta, S.; Pampolino, M.; Jat, M.L.; Shahi, V.; Iftikar, W.; Govil, V.; Singh, V.K. On-Farm Performance of „Nutrient Expert‟ for Maize: Fertilizer Recommendation, Yield and Nutrient Use Efficiency. Better Crops South-Asia 2014, 8, 24–27. [Google Scholar]
- Xu, X.; He, P.; Pampolino, M.F.; Johnston, A.M.; Qiu, S.; Zhao, S.; Chuan, L.; Zhou, W. Fertilizer Recommendation for Maize in China Based on Yield Response and Agronomic Efficiency. Field Crops Res. 2014, 157, 27–34. [Google Scholar] [CrossRef]
- Xu, X.; He, P.; Pampolino, M.F.; Li, Y.; Liu, S.; Xie, J.; Hou, Y.; Zhou, W. Narrowing Yield Gaps and Increasing Nutrient Use Efficiencies Using the Nutrient Expert System for Maize in Northeast China. Field Crops Res. 2016, 194, 75–82. [Google Scholar] [CrossRef]
- Dahal, S.; Shrestha, A.; Dahal, S.; Amgain, L.P. Nutrient Expert Impact on Yield and Economic In Maize and Wheat. Int. J. Appl. Sci. Biotechnol. 2018, 6, 45–52. [Google Scholar] [CrossRef]
- Sun, Y.M.; Yang, Y.M.; Yang, J.F.; Huang, S.H.; Jia, L.L.; Han, B.W.; Liu, M.C. Evaluation of Nutrient Expert System in Improving Nutrient Use Efficiency and Environmental Benefits for Winter Wheat in the Hebei Plain. IOP Conf. Ser. Earth Environ. Sci. 2018, 185, 012011. [Google Scholar] [CrossRef]
- Zhang, J.J.; He, P.; Xu, X.P.; Ding, W.C.; Ullah, S.; Wang, Y.L.; Jia, L.L.; Cui, R.Z.; Wang, H.T.; Zhou, W. Nutrient Expert Improves Nitrogen Efficiency and Environmental Benefits for Winter Wheat in China. Agron. J. 2018, 110, 696–706. [Google Scholar] [CrossRef]
- Timsina, J.; Dutta, S.; Devkota, K.P.; Chakraborty, S.; Neupane, R.K.; Bishta, S.; Amgain, L.P.; Singh, V.K.; Islam, S.; Majumdar, K. Improved Nutrient Management in Cereals Using Nutrient Expert and Machine Learning Tools: Productivity, Profitability and Nutrient Use Efficiency. Agric. Syst. 2021, 192, 103181. [Google Scholar] [CrossRef]
- Rurinda, J.; Zingore, S.; Jibrin, J.M.; Balemi, T.; Masuki, K.; Andersson, J.A.; Pampolino, M.F.; Mohammed, I.; Mutegi, J.; Kamara, A.Y.; et al. Science-Based Decision Support for Formulating Crop Fertilizer Recommendations in Sub-Saharan Africa. Agric. Syst. 2020, 180, 102790. [Google Scholar] [CrossRef]
- Jat, M.L.; Satyanarayana, T.; Majumdar, K.; Parihar, C.M.; Jat, S.L.; Tetarwal, J.P.; Jat, R.K.; Saharawat, Y.S. Fertiliser best management practices for maize systems. Indian J. Fertil. 2013, 9, 80–94. [Google Scholar]
- Linquist, B.A.; Liu, L.; van Kessel, C.; van Groenigen, K.J. Enhanced Efficiency Nitrogen Fertilizers for Rice Systems: Meta-Analysis of Yield and Nitrogen Uptake. Field Crops Res. 2013, 154, 246–254. [Google Scholar] [CrossRef]
- Rajan, M.; Shahena, S.; Chandran, V.; Mathew, L. Controlled Release of Fertilizers—Concept, Reality, and Mechanism. In Controlled Release Fertilizers for Sustainable Agriculture; Elsevier: Amsterdam, The Netherlands, 2021; pp. 41–56. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, L.; Xu, Y.; Yang, Y.; Shi, R. Application of Controlled Release Urea Improved Grain Yield and Nitrogen Use Efficiency: A Meta-Analysis. PLoS ONE 2020, 15, e0241481. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; He, X.; Duan, H.; Han, L.; Huang, G. Evaluation of Controlled Release Urea on the Dynamics of Nitrate, Ammonium, and Its Nitrogen Release in Black Soils of Northeast China. Int. J. Environ. Res. Public Health 2018, 15, 119. [Google Scholar] [CrossRef] [PubMed]
- Lagum, A.A.; Elektorowicz, M. Modification of Nitrifying Microbial Community via DC Electrical Field Application. J. Environ. Chem. Eng. 2022, 10, 107743. [Google Scholar] [CrossRef]
- Meena, A.K.; Singh, D.K.; Pandey, P.C.; Nanda, G. Dynamics of Dry Matter and Nitrogen Distribution in Transplanted Rice on Mollisols. J. Plant Nutr. 2019, 42, 749–758. [Google Scholar] [CrossRef]
- Soares, J.R.; Souza, B.R.; Mazzetto, A.M.; Galdos, M.V.; Chadwick, D.R.; Campbell, E.E.; Jaiswal, D.; Oliveira, J.C.; Monteiro, L.A.; Vianna, M.S.; et al. Mitigation of Nitrous Oxide Emissions in Grazing Systems through Nitrification Inhibitors: A Meta-Analysis. Nutr. Cycl. Agroecosyst. 2023. [Google Scholar] [CrossRef]
- Sridharan, G.V.; Hazarika, D.; Bhat, M.R.; Suraksha, R.S.; Singh, S. Nitrification Inhibition Studies of Neem Coating on Urea Prills. Asian J. Chem. 2017, 29, 196–198. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R.; Kumar, H.; Yadav, R.; Shweta; Yadav, S.M.; Yadav, B.K. Impact of Coated Urea and Nitrogen Levels on Yield, n-Content and N-Uptake in Rice (Oryza sativa). Plant Arch. 2014, 14, 785–787. [Google Scholar]
- Thind, H.S.; Singh, B.; Pannu, R.P.S.; Singh, Y.; Singh, V.; Gupta, R.K.; Singh, G.; Kumar, A.; Vashistha, M. Managing Neem (Azadirachta Indica)-Coated Urea and Ordinary Urea in Wheat (Triticum aestivum) for Improving Nitrogen-Use Efficiency and High Yields. Indian J. Agric. Sci. 2010, 80, 960–964. [Google Scholar]
- Alexander, J.R.; Baker, J.M.; Gamble, J.D.; Venterea, R.T.; Spokas, K.A. Spatiotemporal Distribution of Roots in a Maize-Kura Clover Living Mulch System: Impact of Tillage and Fertilizer N Source. Soil Tillage Res. 2023, 227, 105590. [Google Scholar] [CrossRef]
- Cantarella, H.; Mattos, D.; Quaggio, J.A.; Rigolin, A.T. Fruit Yield of Valencia Sweet Orange Fertilized with Different N Sources and the Loss of Applied N. Nutr. Cycl. Agroecosyst. 2003, 67, 215–223. [Google Scholar] [CrossRef]
- Cantarella, H.; Otto, R.; Soares, J.R.; de Brito Silva, A.G. Agronomic Efficiency of NBPT as a Urease Inhibitor: A Review. J. Adv. Res. 2018, 13, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Modolo, L.V.; da-Silva, C.J.; Brandão, D.S.; Chaves, I.S. A Minireview on What We Have Learned about Urease Inhibitors of Agricultural Interest since Mid-2000s. J. Adv. Res. 2018, 13, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.G.B.; Sequeira, C.H.; Sermarini, R.A.; Otto, R. Urease Inhibitor NBPT on Ammonia Volatilization and Crop Productivity: A Meta-Analysis. Agron. J. 2017, 109, 1–13. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Bindraban, P.S. Nanofertilizers: New Products for the Industry? J. Agric. Food Chem. 2018, 66, 6462–6473. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.K.; Song, X.P.; Joshi, A.; Rajput, V.D.; Singh, M.; Sharma, A.; Singh, R.K.; Li, D.M.; Arora, J.; Minkina, T.; et al. Nanofertilizer Possibilities for Healthy Soil, Water, and Food in Future: An Overview. Front. Plant Sci. 2022. 13, 865048. [CrossRef]
- Kottegoda, N.; Munaweera, I.; Madusanka, N.; Karunaratne, V. A Green Slow-Release Fertilizer Composition Based on Urea-Modified Hydroxyapatite Nanoparticles Encapsulated Wood. Curr. Sci. 2011, 101, 73–78. [Google Scholar]
- Kottegoda, N.; Sandaruwan, C.; Priyadarshana, G.; Siriwardhana, A.; Rathnayake, U.A.; Berugoda Arachchige, D.M.; Kumarasinghe, A.R.; Dahanayake, D.; Karunaratne, V.; Amaratunga, G.A.J. Urea-Hydroxyapatite Nanohybrids for Slow Release of Nitrogen. ACS Nano 2017, 11, 1214–1221. [Google Scholar] [CrossRef]
- Pereira, L.G.; Chagas, M.F.; Dias, M.O.S.; Cavalett, O.; Bonomi, A. Life Cycle Assessment of Butanol Production in Sugarcane Biorefineries in Brazil. J. Clean. Prod. 2015, 96, 557–568. [Google Scholar] [CrossRef]
- Yamamoto, C.F.; Pereira, E.I.; Mattoso, L.H.C.; Matsunaka, T.; Ribeiro, C. Slow Release Fertilizers Based on Urea/Urea-Formaldehyde Polymer Nanocomposites. Chem. Eng. J. 2016, 287, 390–397. [Google Scholar] [CrossRef]
- Reddy, M.R.; Avinash, A.V.; Jaswal, A. Techniques for Improving Fertilizer Use Efficiency—A Review. Eur. J. Mol. Clin. Med. 2020, 7, 2725–2732. [Google Scholar]
- Ozdogan, M. The Spatial Distribution of Crop Types from MODIS Data: Temporal Unmixing Using Independent Component Analysis. Remote Sens. Environ. 2010, 114, 1190–1204. [Google Scholar] [CrossRef]
- Balafoutis, A.; Beck, B.; Fountas, S.; Vangeyte, J.; Van Der Wal, T.; Soto, I.; Gómez-Barbero, M.; Barnes, A.; Eory, V. Precision Agriculture Technologies Positively Contributing to Ghg Emissions Mitigation, Farm Productivity and Economics. Sustainability 2017, 9, 1339. [Google Scholar] [CrossRef]
- Hayat, S.; Yadav, S.; Wani, A.S.; Irfan, M.; Alyemini, M.N.; Ahmad, A. Impact of Sodium Nitroprusside on Nitrate Reductase, Proline Content, and Antioxidant System in Tomato under Salinity Stress. Hortic. Environ. Biotechnol. 2012, 53, 362–367. [Google Scholar] [CrossRef]
- Colaço, A.F.; Bramley, R.G.V. Do Crop Sensors Promote Improved Nitrogen Management in Grain Crops? Field Crops Res. 2018, 218, 126–140. [Google Scholar] [CrossRef]
- Ke, J.; He, R.; Hou, P.; Ding, C.; Ding, Y.; Wang, S.; Liu, Z.; Tang, S.; Ding, C.; Chen, L.; et al. Combined Controlled-Released Nitrogen Fertilizers and Deep Placement Effects of N Leaching, Rice Yield and N Recovery in Machine-Transplanted Rice. Agric. Ecosyst. Environ. 2018, 265, 402–412. [Google Scholar] [CrossRef]
- Koudjega, K.; Ablede, K.A.; Lawson, I.Y.D.; Abekoe, M.K.; Owusu-Bennoah, E.; Tsatsu, D.K. Reducing Ammonia Volatilization and Improving Nitrogen Use Efficiency of Rice at Different Depths of Urea Supergranule Application. Commun. Soil Sci. Plant Anal. 2019, 50, 974–986. [Google Scholar] [CrossRef]
- Bautista, E.U.; Koike, M.; Suministrado, D.C. PM—Power and Machinery: Mechanical Deep Placement of Nitrogen in Wetland Rice. J. Agric. Eng. Res. 2001, 78, 333–346. [Google Scholar] [CrossRef]
- Mehrabi, F.; Sepaskhah, A.R. Interaction Effects of Planting Method, Irrigation Regimes, and Nitrogen Application Rates on Yield, Water and Nitrogen Use Efficiencies of Winter Wheat (Triticum aestivum). Int. J. Plant Prod. 2018, 12, 265–283. [Google Scholar] [CrossRef]
- Zhang, X.; Meng, F.; Li, H.; Wang, L.; Wu, S.; Xiao, G.; Wu, W. Optimized Fertigation Maintains High Yield and Mitigates N2O and NO Emissions in an Intensified Wheat–Maize Cropping System. Agric. Water Manag. 2019, 211, 26–36. [Google Scholar] [CrossRef]
- Lal, M.K.; Tiwari, R.K.; Kumar, A.; Dey, A.; Kumar, R.; Kumar, D.; Jaiswal, A.; Changan, S.S.; Raigond, P.; Dutt, S.; et al. Mechanistic Concept of Physiological, Biochemical, and Molecular Responses of the Potato Crop to Heat and Drought Stress. Plants 2022, 11, 2857. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Lal, M.K.; Sahoo, S.K.; Dash, G.K.; Sahoo, U.; Behera, B.; Nayak, L.; Bagchi, T.B. Title: The Diversity of Phytic Acid Content and Grain Processing Play Decisive Role on Minerals Bioavailability in Rice. J. Food Compos. Anal. 2022, 115, 105032. [Google Scholar] [CrossRef]
- Hashim, M.; Dhar, S.; Vyas, A.K.; Pramesh, V.; Kumar, B. Integrated Nutrient Management in Maize (Zea Mays)-Wheat (Triticum aestivum) Cropping System. Indian J. Agron. 2015, 60, 352–359. [Google Scholar]
- Zhang, J.; Jiang, X.; Li, T.; Chang, T. Effect of Elevated Temperature Stress on the Production and Metabolism of Photosynthate in Tomato (Lycopersicon esculentum L.) Leaves. J. Hortic. Sci. Biotechnol. 2012, 87, 293–298. [Google Scholar] [CrossRef]
- Martínez-Dalmau, J.; Berbel, J.; Ordóñez-Fernández, R. Nitrogen Fertilization. A Review of the Risks Associated with the Inefficiency of Its Use and Policy Responses. Sustainability 2021, 13, 5625. [Google Scholar] [CrossRef]
- Singh, D.K.; Pandey, P.C.; Nanda, G.; Gupta, S. Long-Term Effects of Inorganic Fertilizer and Farmyard Manure Application on Productivity, Sustainability and Profitability of Rice-Wheat System in Mollisols. Arch. Agron. Soil Sci. 2019, 65, 139–151. [Google Scholar] [CrossRef]
- Jat, M.L.; Bijay-Singh; Gerard, B. Nutrient Management and Use Efficiency in Wheat Systems of South Asia. Adv. Agron. 2014, 125, 171–259. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, M.; Kumar, S.; Meena, R.K.; Yadav, M.R.; Makarana, G.; Kushwaha, M.; Dutta, S.; Kumar, R. Enhancement in Productivity and Quality of Fodder Maize Cultivars through Integrated Nutrient Management Strategies. Indian J. Agric. Sci. 2022, 92, 126–130. [Google Scholar] [CrossRef]
- Wu, W.; Ma, B. Integrated Nutrient Management (INM) for Sustaining Crop Productivity and Reducing Environmental Impact: A Review. Sci. Total Environ. 2015, 512–513, 415–427. [Google Scholar] [CrossRef]
- Duan, Y.; Xu, M.; Gao, S.; Yang, X.; Huang, S.; Liu, H.; Wang, B. Nitrogen Use Efficiency in a Wheat-Corn Cropping System from 15 Years of Manure and Fertilizer Applications. Field Crops Res. 2014, 157, 47–56. [Google Scholar] [CrossRef]
- Midya, A.; Saren, B.K.; Dey, J.K.; Maitra, S.; Praharaj, S.; Gaikwad, D.J.; Gaber, A.; Alhomrani, M.; Hossain, A. Crop Establishment Methods and Integrated Nutrient Management Improve: Part Ii. Nutrient Uptake and Use Efficiency and Soil Health in Rice (Oryza sativa L.) Field in the Lower Indo-Gangetic Plain, India. Agronomy 2021, 11, 1894. [Google Scholar] [CrossRef]
- Li, G.; Zhao, B.; Dong, S.; Zhang, J.; Liu, P.; Vyn, T.J. Impact of Controlled Release Urea on Maize Yield and Nitrogen Use Efficiency under Different Water Conditions. PLoS ONE 2017, 12, e0181774. [Google Scholar] [CrossRef] [PubMed]
- Aulakh, M.S.; Bijay-Singh. Nitrogen Losses and Fertilizer N Use Efficiency in Irrigated Porous Soils. Nutr. Cycl. Agroecosyst. 1996, 47, 197–212. [Google Scholar] [CrossRef]
- Jain, N.K.; Meena, H.N.; Bhaduri, D.; Yadav, R.S. Drip Fertigation and Irrigation Interval Effects on Growth, Productivity, Nutrient, and Water Economy in Summer Peanut. Commun. Soil Sci. Plant Anal. 2018, 49, 2406–2417. [Google Scholar] [CrossRef]
- Teixeira, E.I.; George, M.; Herreman, T.; Brown, H.; Fletcher, A.; Chakwizira, E.; de Ruiter, J.; Maley, S.; Noble, A. The Impact of Water and Nitrogen Limitation on Maize Biomass and Resource-Use Efficiencies for Radiation, Water and Nitrogen. Field Crops Res. 2014, 168, 109–118. [Google Scholar] [CrossRef]
- van Donk, S.J.; Shaver, T.M. Effects of Nitrogen Application Frequency via Subsurface Drip Irrigation on Corn Development and Grain Yield. J. Plant Nutr. 2016, 39, 1830–1839. [Google Scholar] [CrossRef]
- Chen, Y.M.; Zhang, J.Y.; Xu, X.; Qu, H.Y.; Hou, M.; Zhou, K.; Jiao, X.G.; Sui, Y.Y. Effects of Different Irrigation and Fertilization Practices on Nitrogen Leaching in Facility Vegetable Production in Northeastern China. Agric. Water Manag. 2018, 210, 165–170. [Google Scholar] [CrossRef]
- Renwick, L.L.R.; Bowles, T.M.; Deen, W.; Gaudin, A.C.M. Potential of Increased Temporal Crop Diversity to Improve Resource Use Efficiencies: Exploiting Water and Nitrogen Linkages. In Agroecosystem Diversity: Reconciling Contemporary Agriculture and Environmental Quality; Elsevier: Amsterdam, The Netherlands, 2018; pp. 55–73. [Google Scholar] [CrossRef]
- Banjara, T.R.; Bohra, J.S.; Kumar, S.; Singh, T.; Shori, A.; Prajapat, K. Sustainable Alternative Crop Rotations to the Irrigated Rice-Wheat Cropping System of Indo-Gangetic Plains of India. Arch. Agron. Soil Sci. 2021, 68, 1568–1585. [Google Scholar] [CrossRef]
- Xu, Y.; Nie, L.; Buresh, R.J.; Huang, J.; Cui, K.; Xu, B.; Gong, W.; Peng, S. Agronomic Performance of Late-Season Rice under Different Tillage, Straw, and Nitrogen Management. Field Crops Res. 2010, 115, 79–84. [Google Scholar] [CrossRef]
- Dwivedi, B.S.; Singh, V.K.; Meena, M.C. Efficient Nitrogen Management Under Predominant Cropping Systems of India. In The Indian Nitrogen Assessment: Sources of Reactive Nitrogen, Environmental and Climate Effects, Management Options, and Policies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 95–115. [Google Scholar] [CrossRef]
- Kumar, N.; Chaudhary, A.; Ahlawat, O.P.; Naorem, A.; Upadhyay, G.; Chhokar, R.S.; Gill, S.C.; Khippal, A.; Tripathi, S.C.; Singh, G.P. Crop Residue Management Challenges, Opportunities and Way Forward for Sustainable Food-Energy Security in India: A Review. Soil Tillage Res. 2023, 228, 105641. [Google Scholar] [CrossRef]
- Yadvinder-Singh; Singh, M.; Sidhu, H.S.; Humphreys, E.; Thind, H.S.; Jat, M.L.; Blackwell, J.; Singh, V. Nitrogen Management for Zero till Wheat with Surface Retention of Rice Residues in North-West India. Field Crops Res. 2014, 184, 183–191. [Google Scholar] [CrossRef]
- Vashisht, B.B.; Jalota, S.K.; Ramteke, P.; Kaur, R.; Jayeswal, D.K. Impact of Rice (O. sativa L.) Straw Incorporation Induced Changes in Soil Physical and Chemical Properties on Yield, Water and Nitrogen–Balance and –Use Efficiency of Wheat (T. aestivum L.) in Rice–Wheat Cropping System: Field and Simulation Studies. Agric. Syst. 2021, 194, 103279. [Google Scholar] [CrossRef]
- Qin, X.; Huang, T.; Lu, C.; Dang, P.; Zhang, M.; Guan, X.K.; Wen, P.F.; Wang, T.C.; Chen, Y.; Siddique, K.H.M. Benefits and Limitations of Straw Mulching and Incorporation on Maize Yield, Water Use Efficiency, and Nitrogen Use Efficiency. Agric. Water Manag. 2021, 256, 107128. [Google Scholar] [CrossRef]
- Lin, L.; Zheng, Z.; Hua, T.; Ashraf, U.; Hamoud, Y.A.; Alaa, A.A.; Xiangru, T.; Meiyang, D.; Zaiman, W.; Shenggang, P. Nitrogen Deep Placement Combined with Straw Mulch Cultivation Enhances Physiological Traits, Grain Yield and Nitrogen Use Efficiency in Mechanical Pot-Seedling Transplanting Rice. Rice Sci. 2022, 29, 89–100. [Google Scholar] [CrossRef]
- Huang, S.; Zeng, Y.; Wu, J.; Shi, Q.; Pan, X. Effect of Crop Residue Retention on Rice Yield in China: A Meta-Analysis. Field Crops Res. 2013, 154, 188–194. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).