Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth Conditions
2.2. Collection of Soil Samples
2.3. Rhizosphere Bacterial Genome Sequencing
2.4. Bioinformatical Analyses
3. Results
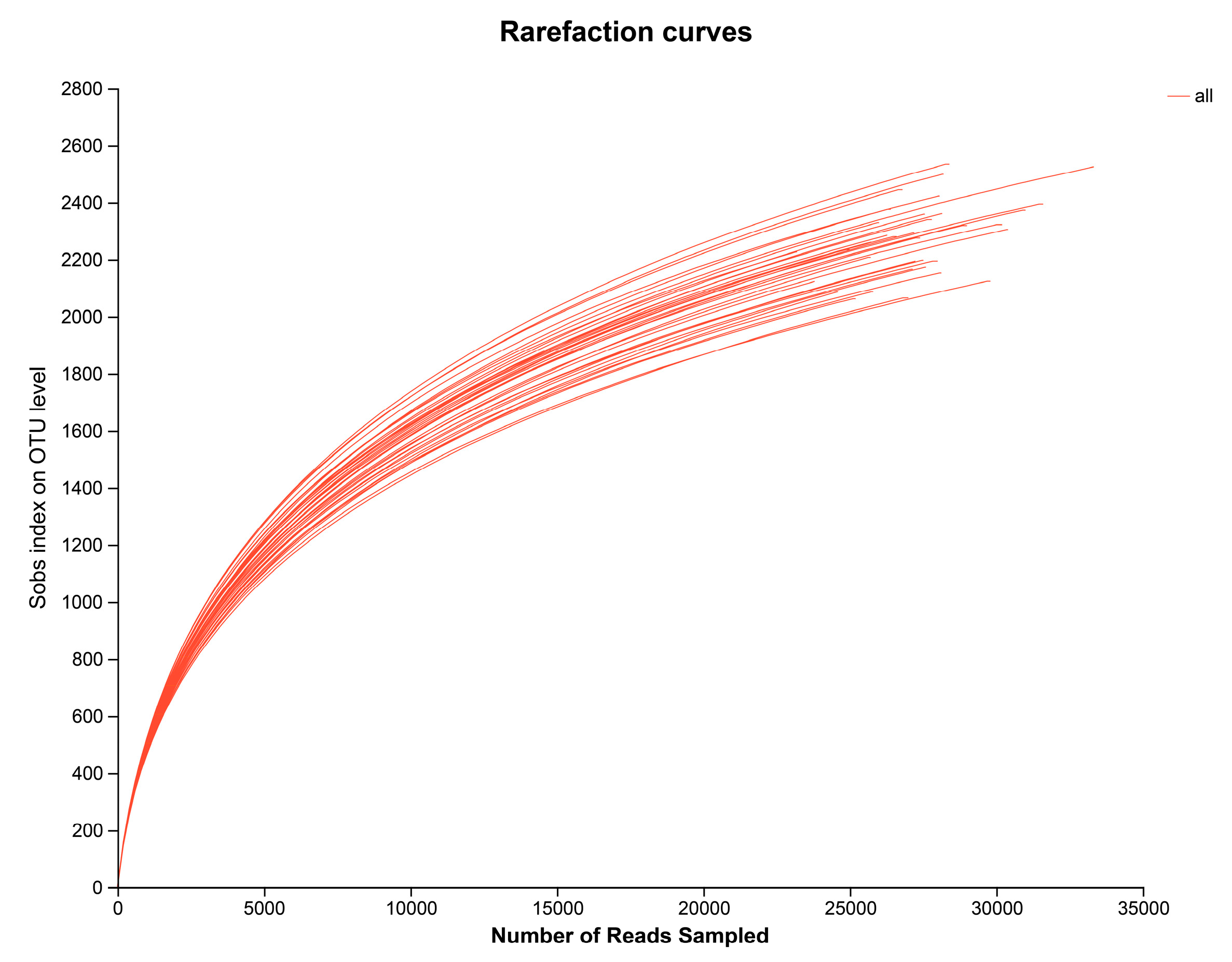
3.1. Quality Analysis of Barley Rhizosphere Bacteria Sequencing
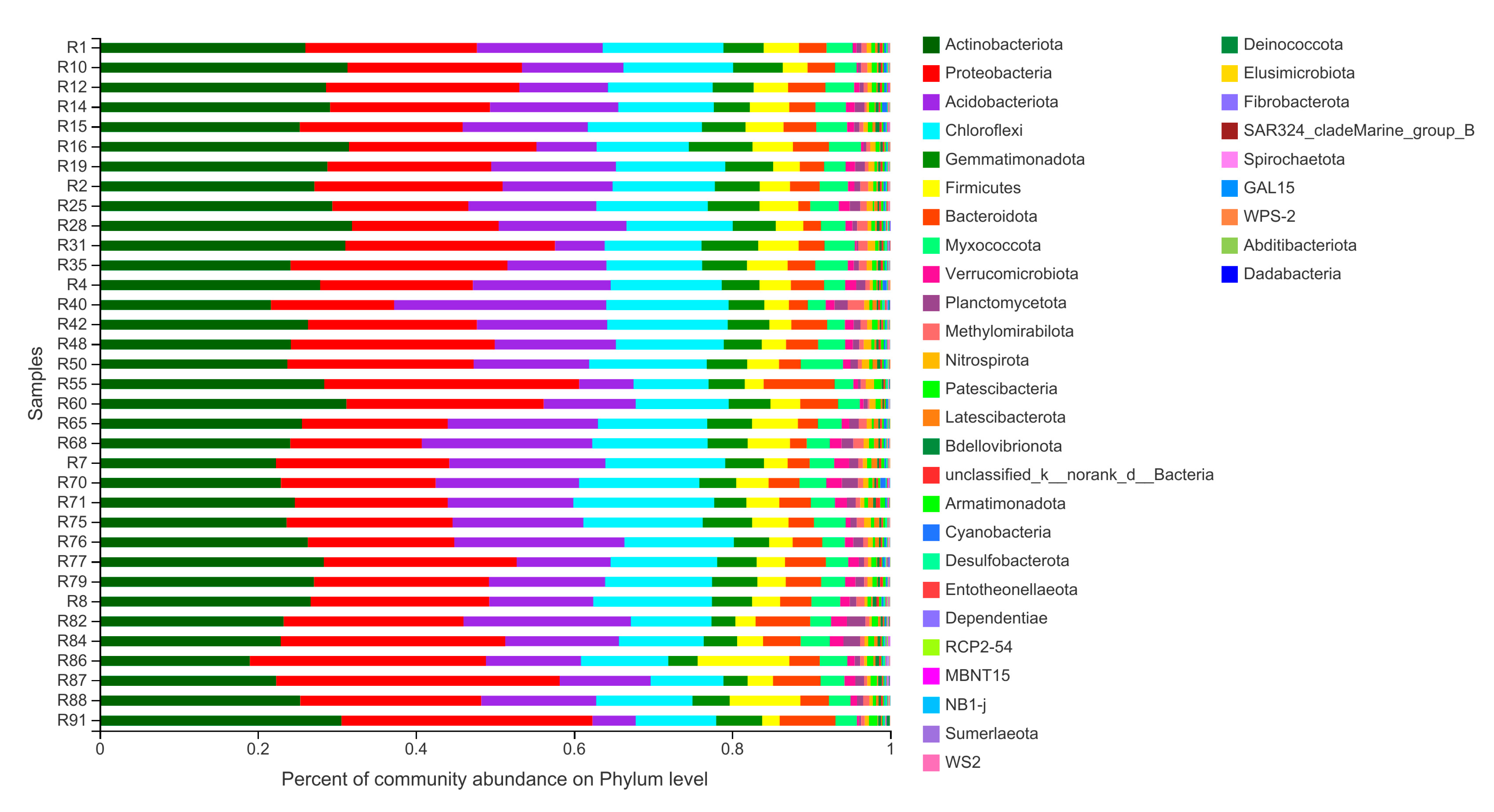
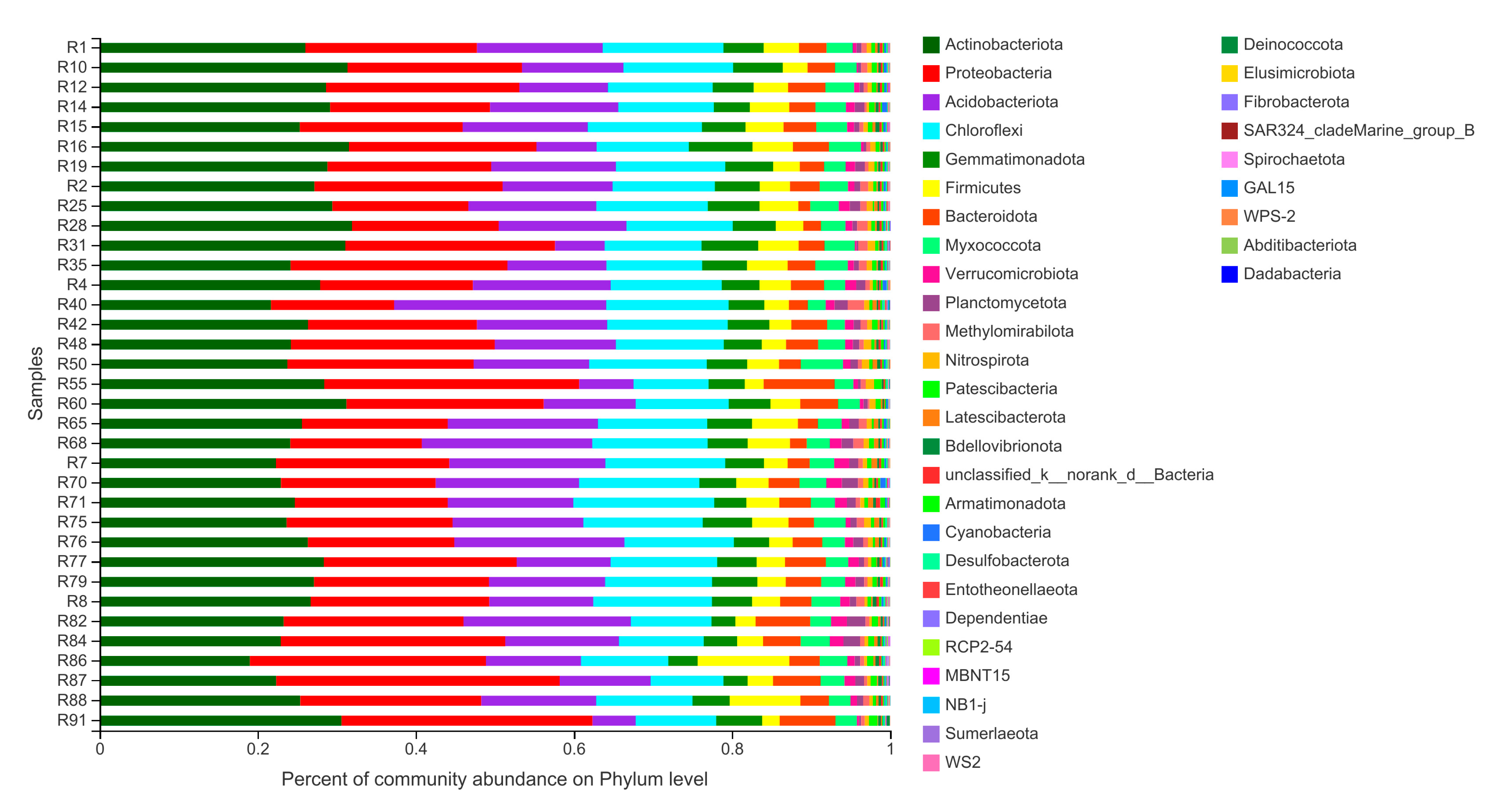
3.2. Bacterial Community Composition in Barley Rhizosphere at Phylum Level
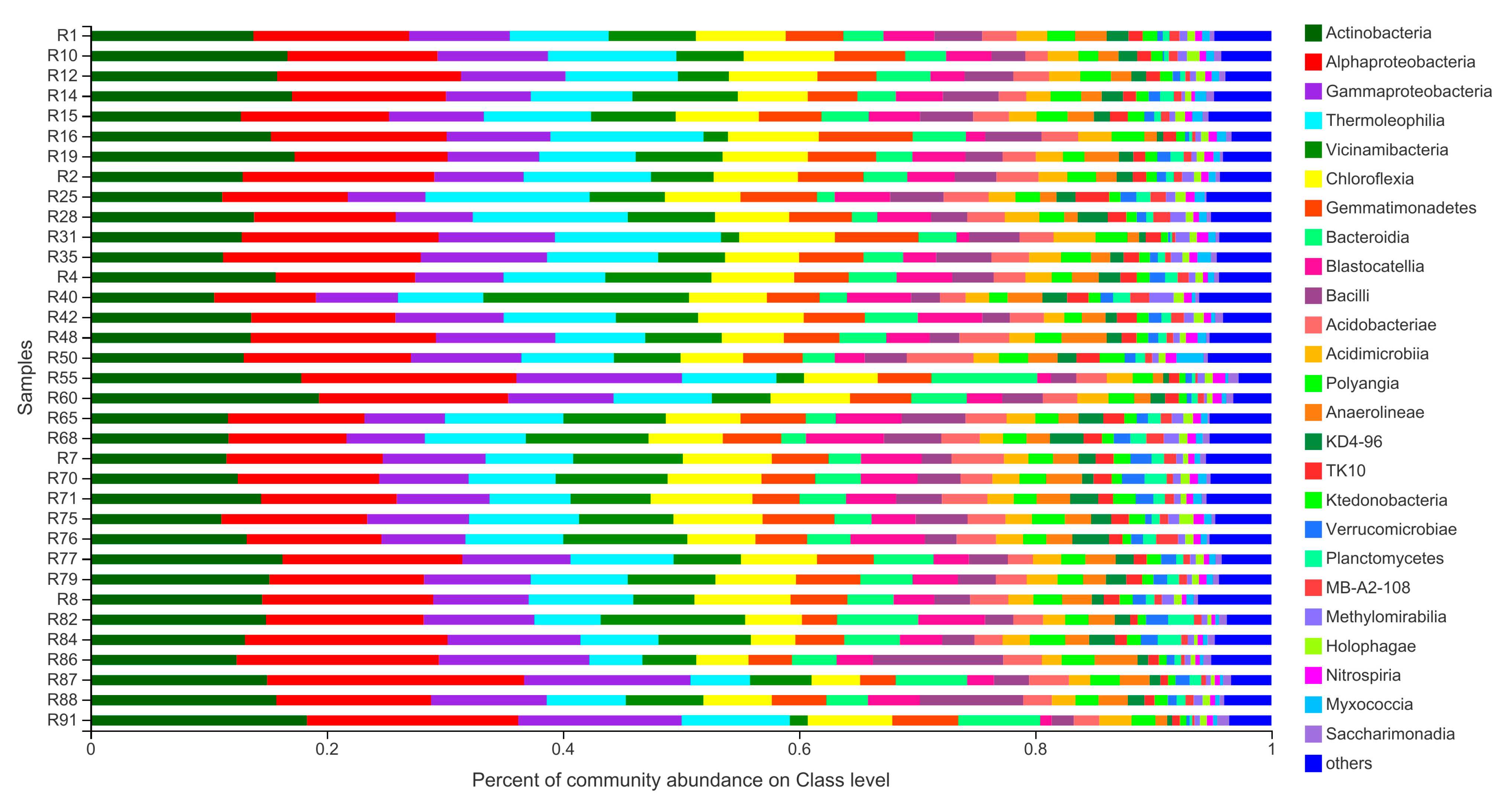
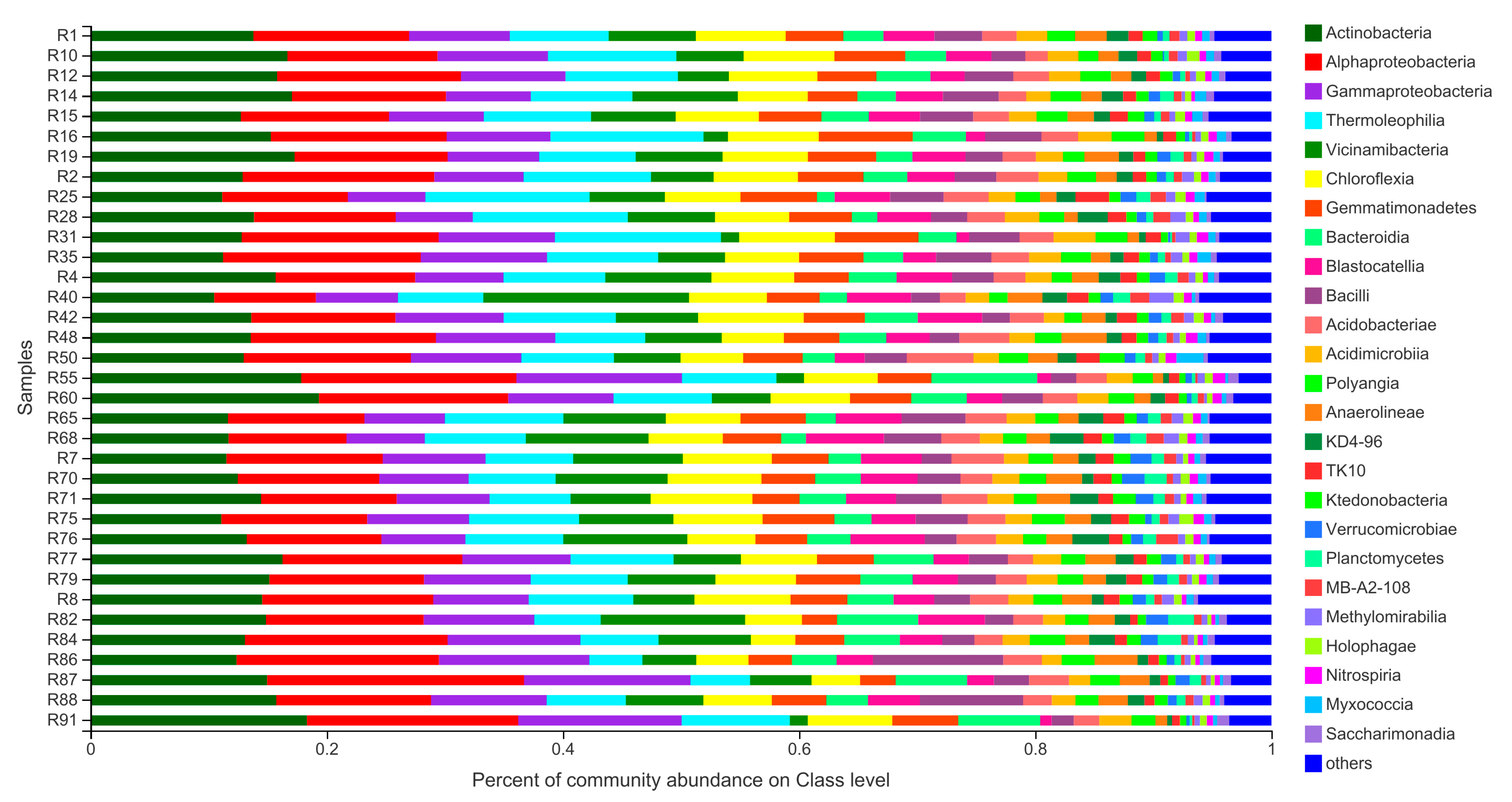
3.3. Community Composition of Barley Rhizosphere Bacterial Community at Class Level
3.4. The Bacterial Community Composition of Barley Rhizosphere at the Level of Order and Family
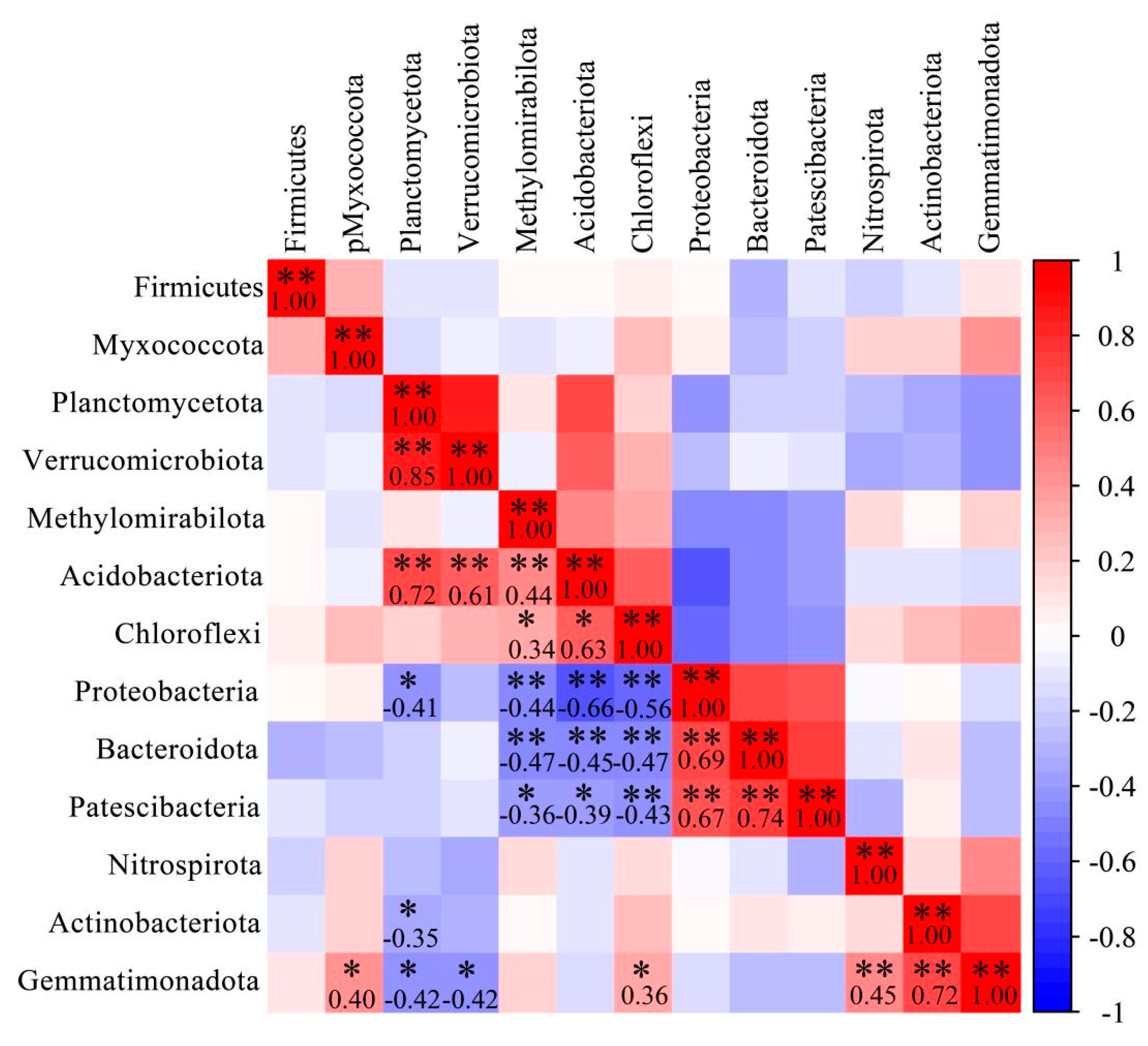
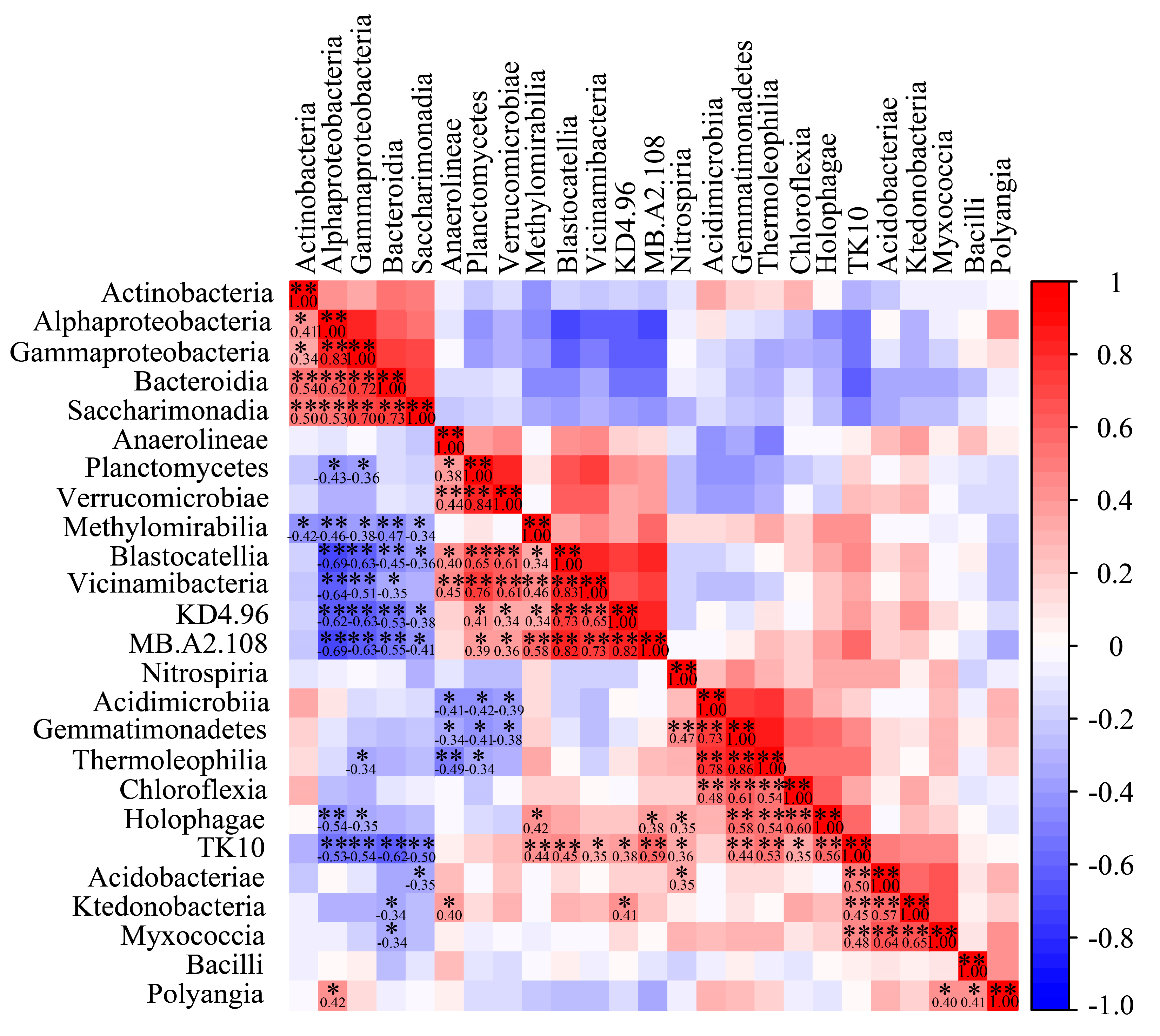
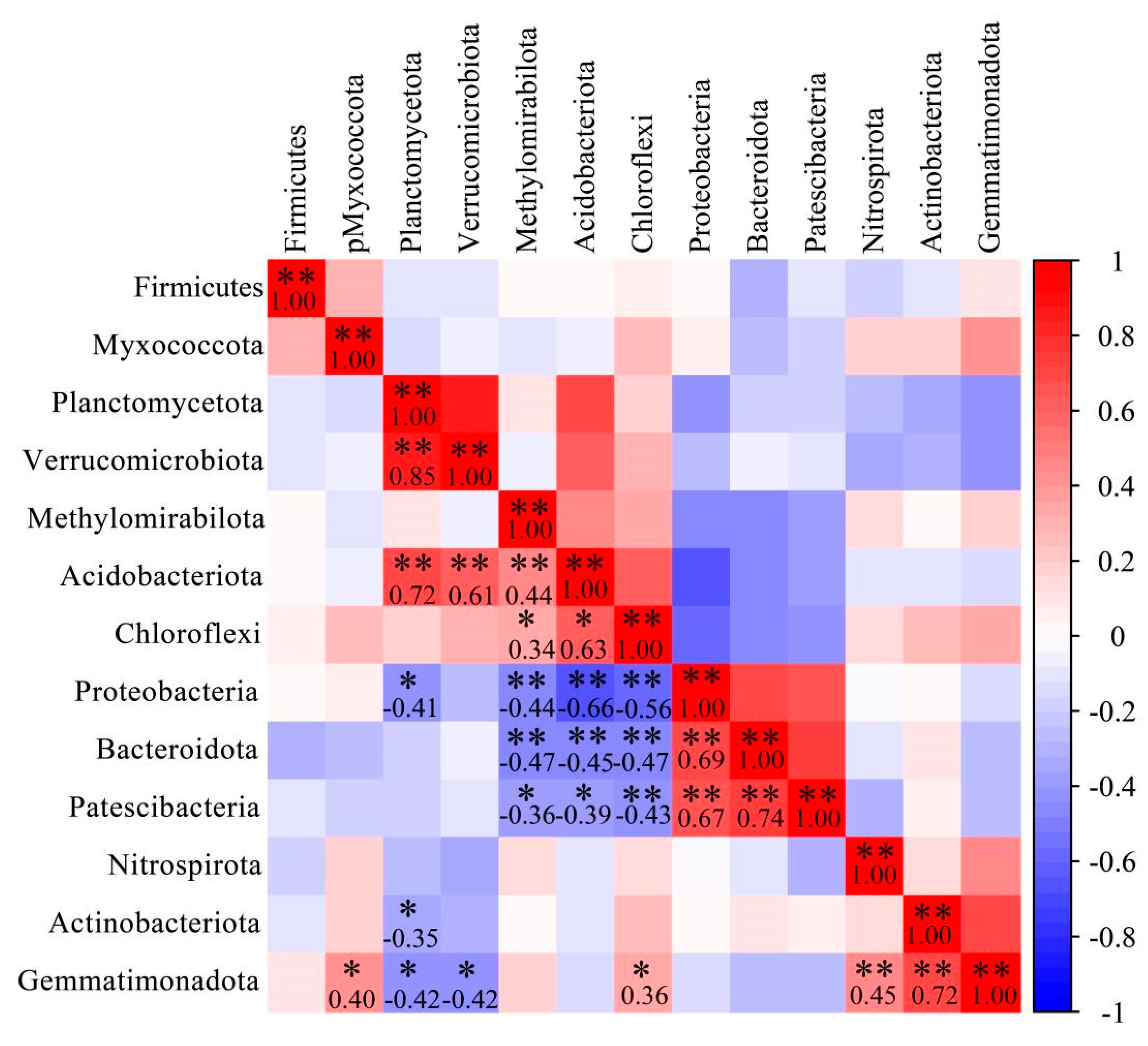
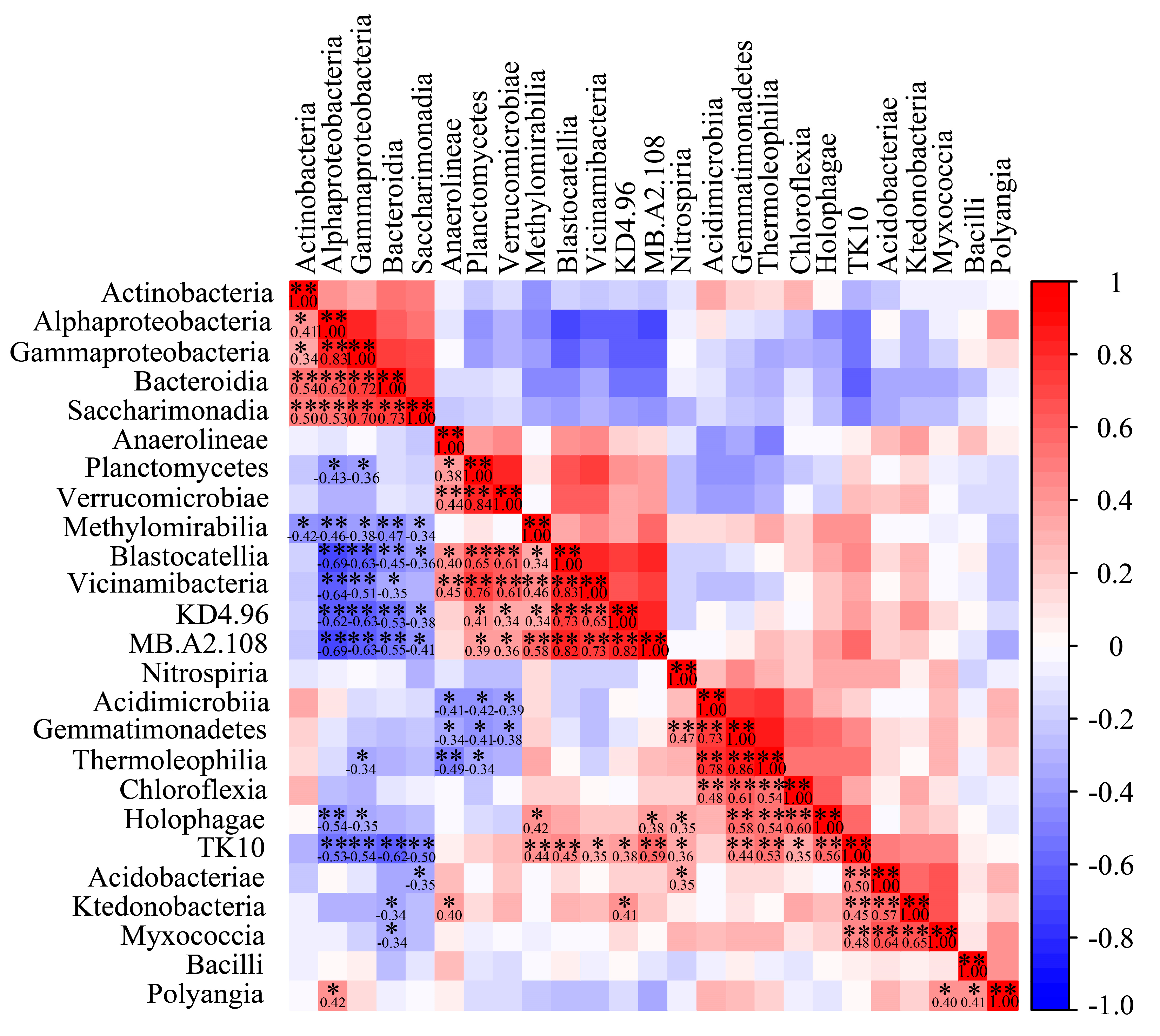
3.5. Correlation Analysis of Dominant Bacterial Community in Barley Rhizosphere
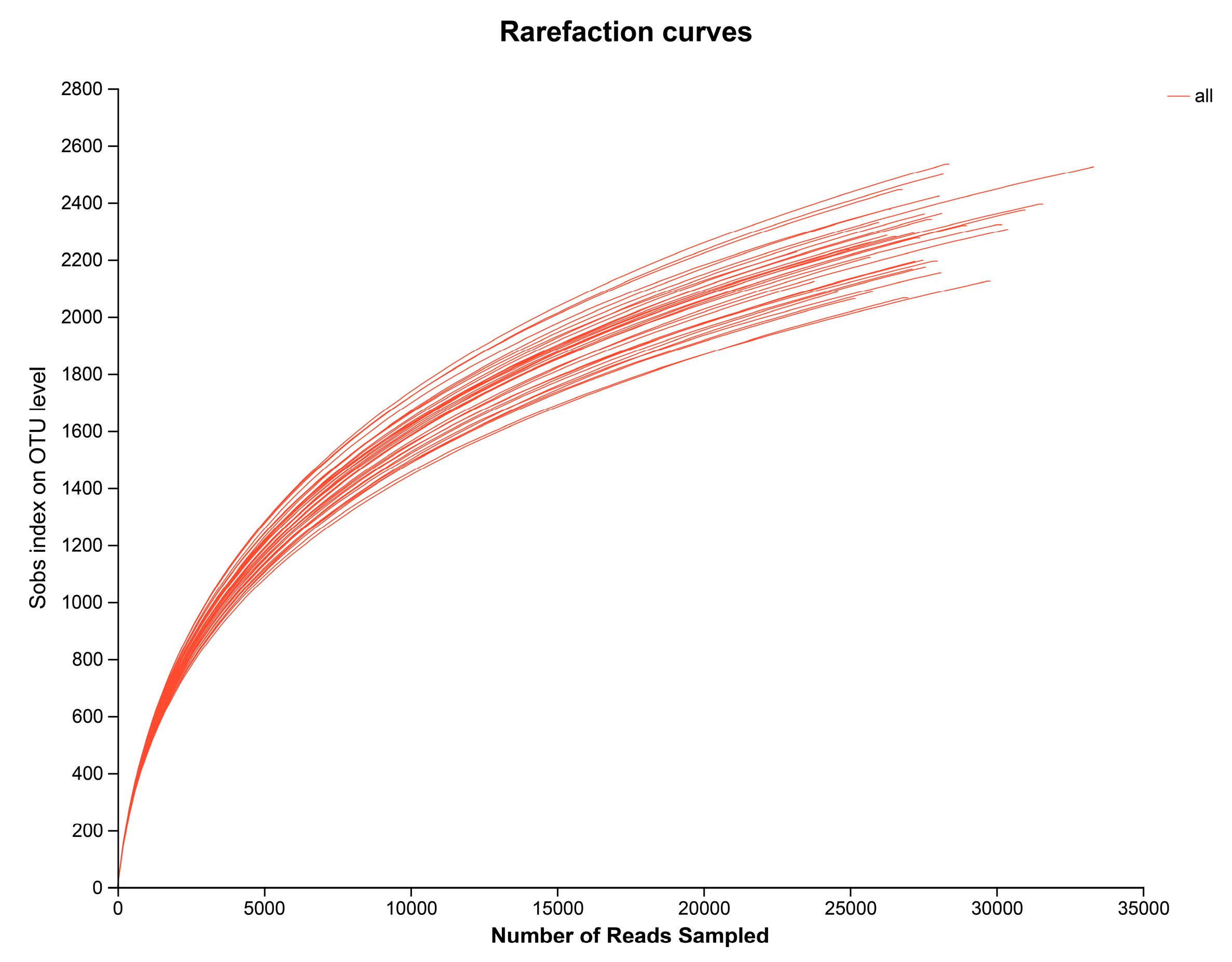
3.6. Alpha Diversity of Bacterial Community Structure in Barley Rhizosphere
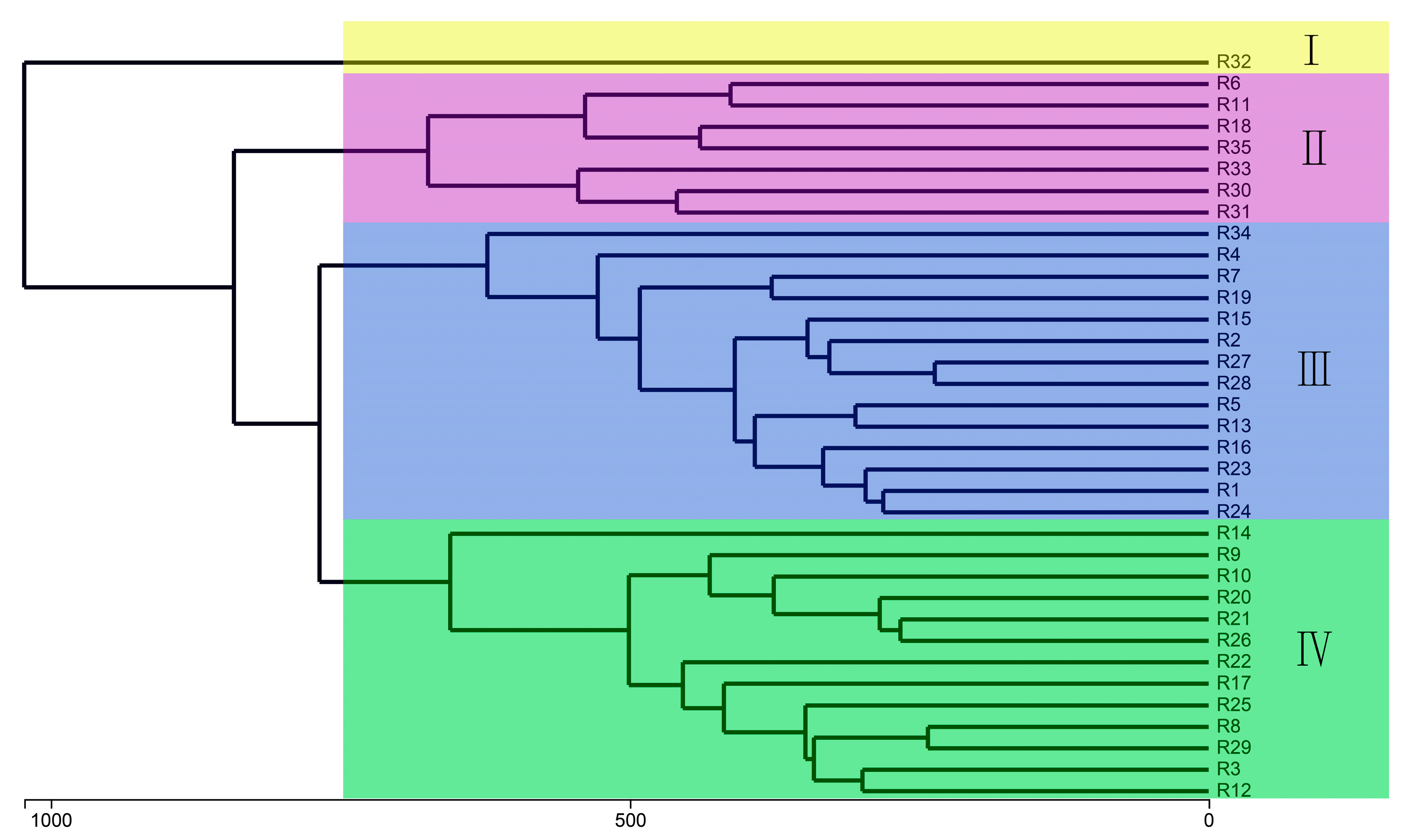
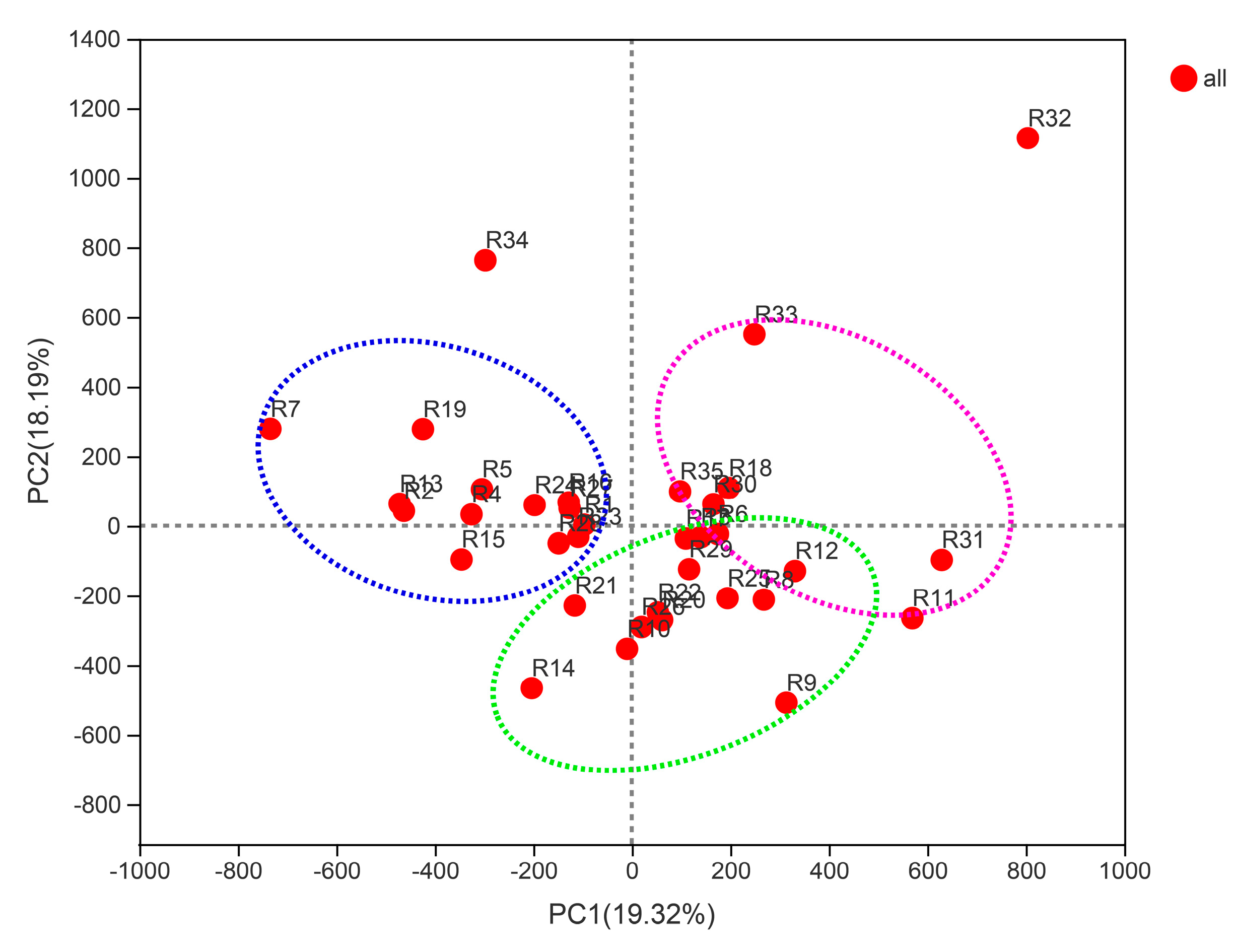
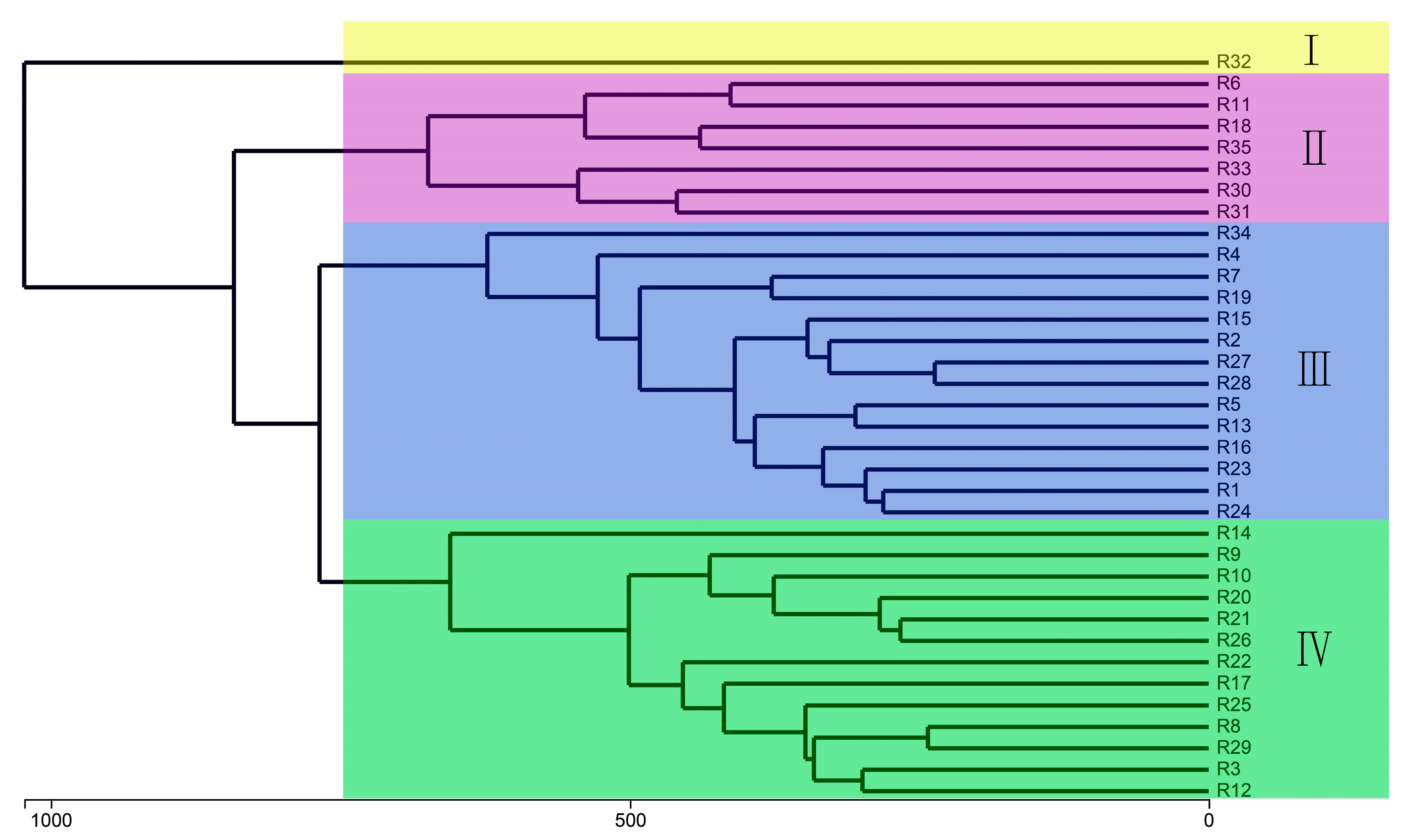
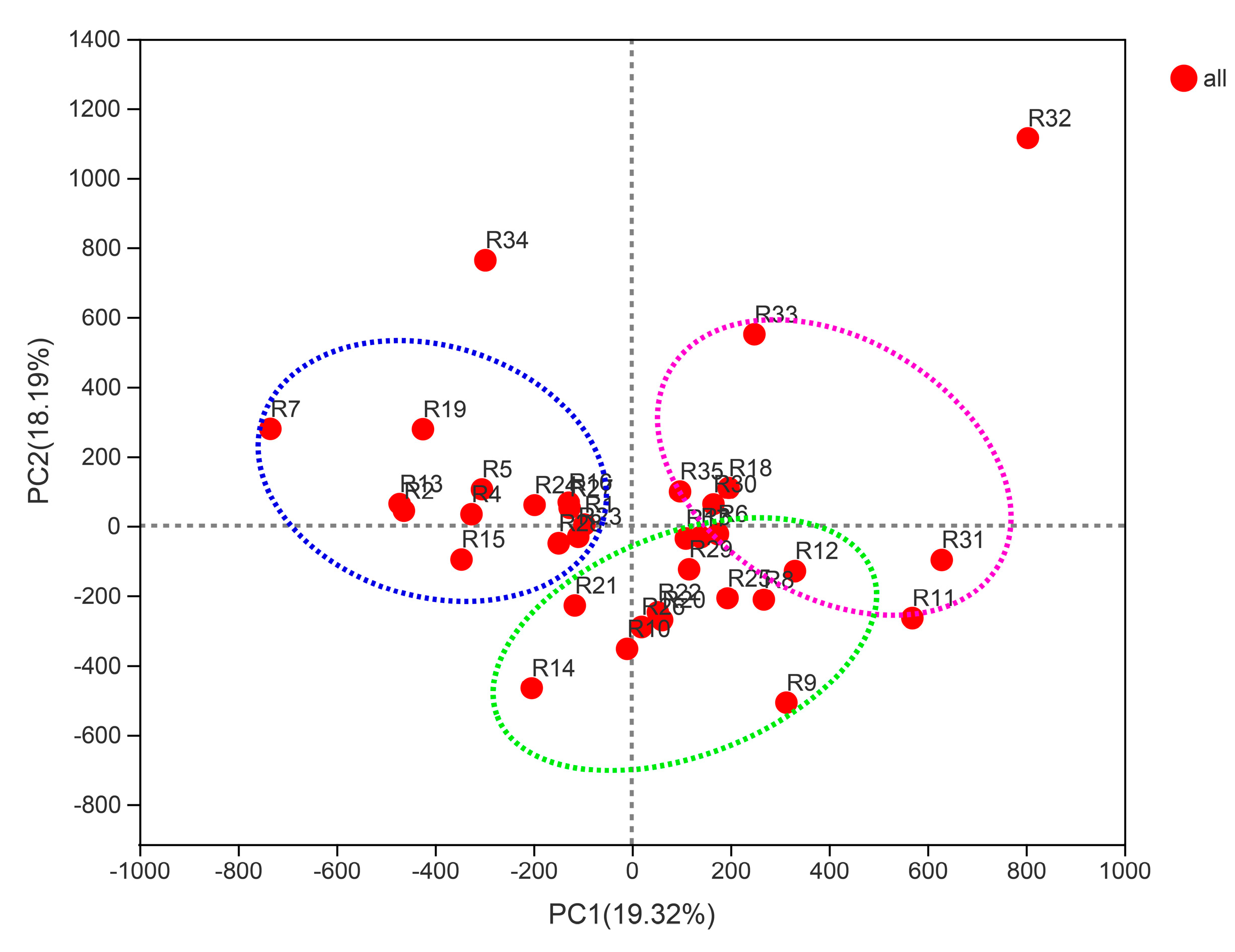
3.7. Beta Diversity of Bacterial Community Structure in Barley Rhizosphere
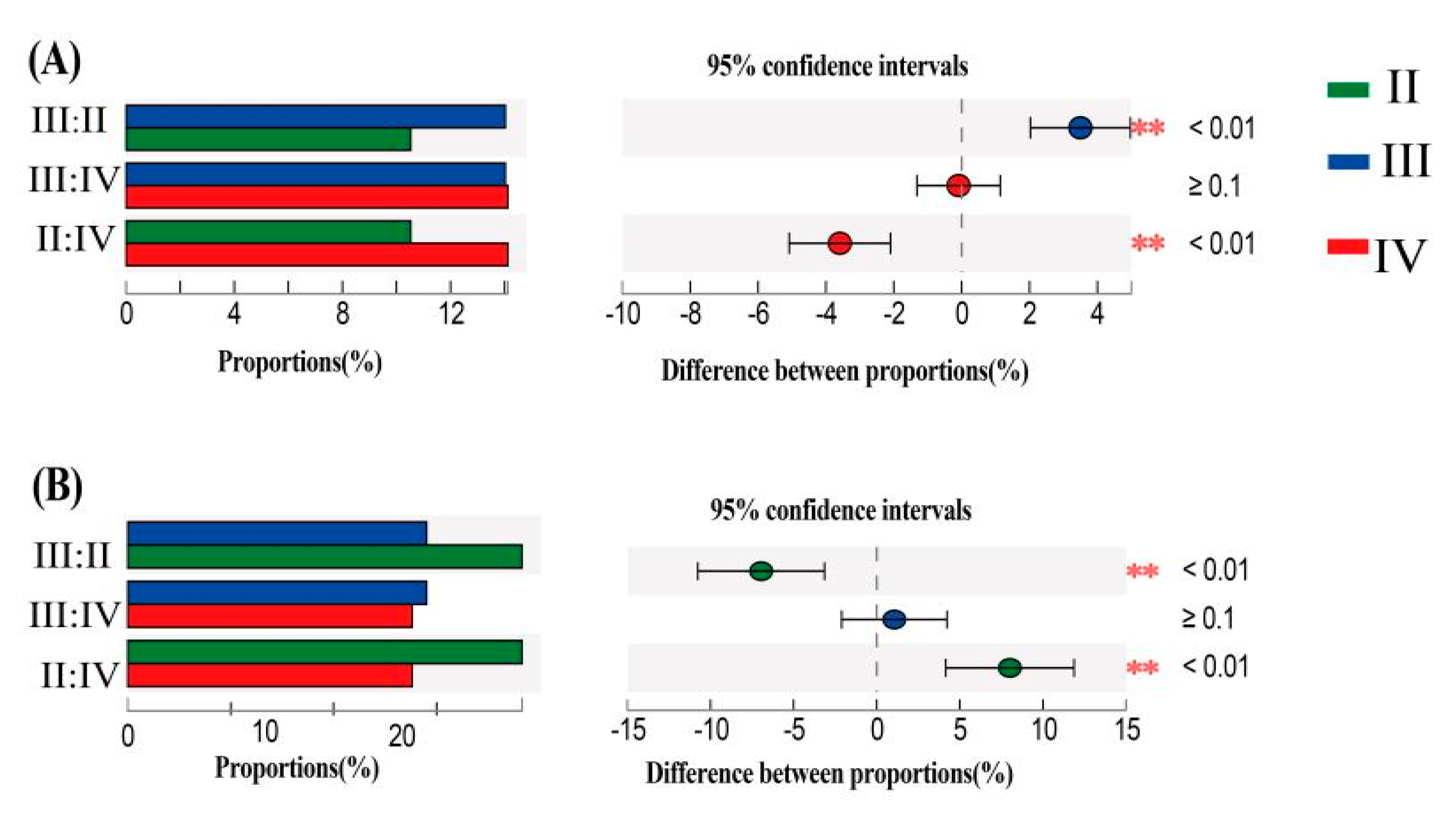
3.8. Differences in Rhizosphere Bacterial Communities of Different Barley Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bhaduri, D.; Pal, S.; Purakayastha, T.J.; Chakraborty, K.; Yadav, R.S.; Akhtar, M.S. Soil quality and plant-microbe interactions in the rhizosphere. Sustain. Agric. Rev. 2015, 17, 307–335. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Fan, K.K.; Cardona, C.; Li, Y.T.; Shi, Y.; Xiang, X.J.; Chen, C.C.; Wang, H.F.; Gilbert, J.A.; Chu, H.Y. Rhizosphere-associated bacterial network structure and spatial distribution differ significantly from bulk soil in wheat crop fields. Soil Biol. Biochem. 2017, 113, 275–284. [Google Scholar] [CrossRef]
- He, C.; Zhu, H.; Kong, X.; Li, C.; Yan, X. Environmental effects of returning rice and wheat straw to fields. Agric. Sci. Technol. 2017, 18, 1710–1715. [Google Scholar] [CrossRef]
- Butler, J.L.; Williams, M.A.; Bottomley, P.J.; Myrold, D.D. Microbial community dynamics associated with rhizosphere carbon flow. Appl. Environ. Microbiol. 2003, 69, 6793–6800. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, H.; Wan, S.; Hua, K.; Wang, D.; Guo, X.; He, C. Fertilisation practice changes rhizosphere microbial community structure in the agroecosystem. Ann. Appl. Biol. 2019, 174, 123–132. [Google Scholar] [CrossRef]
- Zhou, Y.; Lu, G.; Sun, G.L.; Sun, D.K.; Ren, X.F. Transcriptome and metabolite insights into domestication process of cultivated barley in China. Plants 2022, 11, 209. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, Y.; Lu, G.; Zhang, A.; Zhao, T.; Sun, G. Assessment of soil quality for guided fertilization in 7 barley agro-ecological areas of China. PLoS ONE 2021, 17, e0261638. [Google Scholar] [CrossRef]
- Tarjoc, F.C.; Tabără, V. Issues concerning the impact of foliar fertilization on the production of barley in Timisoara. Res. J. Agric. Sci. 2011, 43, 184–189. [Google Scholar]
- Chala, G.; Terbush, A.; Assegid, D. Performance evaluation and selection of improved food barley (Hordeum vulgare L.) varieties for their adaptability in West Hararghe Zone, Eastern Oromia, Ethiopia. Asian J. Plant Sci. Res. 2021, 11, 49–56. [Google Scholar]
- Ansari, M.; Rajabi, R.; Azimzadeh, S.; Hesami, A.; Soleymani, K.; Abediasl, G.R. Study on adaptation and stability of grain yield of barley genotypes under cold rainfed condition. Seed Plant 2017, 23, 387–402. [Google Scholar]
- Ievina, S.; Arta, K.; Vija, S.; Aina, K.; Mauritz, A.; Kari, B.O.A. Adaptability of hull-less barley varieties to different cropping systems and climatic conditions. Acta Agric. Scand. 2018, 69, 1–11. [Google Scholar] [CrossRef]
- Azarbad, H.; Tremblay, J.; Bainard, L.D.; Yergeau, E. Relative and quantitative rhizosphere microbiome profiling result in distinct abundance patterns. Front. Microbiol. 2021, 12, 798023. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; van Themaat, E.V.L.; Ahmadinejad, N.; Assenza, F.; Rauf, P.; Huettel, B.; Reinhardt, R.; Schmelzer, E.; et al. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellín, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, 911–920. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Schirmer, M.; Ijaz, U.Z.; D’Amore, R.; Hall, N.; Sloan, W.T.; Quince, C. Insight into biases and sequencing errors for amplicon sequencing with the Illumina MiSeq platform. Nucleic Acids Res. 2015, 43, e37. [Google Scholar] [CrossRef]
- Li, H.; Luo, L.Y.; Tang, B.; Guo, H.L.; Cao, Z.Y.; Zeng, Q.; Chen, S.L.; Chen, Z.H. Dynamic changes of rhizosphere soil bacterial community and nutrients in cadmium polluted soils with soybean-corn intercropping. BMC Microbiol. 2022, 22, 57. [Google Scholar] [CrossRef]
- Emmett, B.D.; Buckley, D.H.; Drinkwater, L.E. Plant growth rate and nitrogen uptake shape rhizosphere bacterial community composition and activity in an agricultural field. New Phytol. 2020, 225, 960–973. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Lan, Y.M.; Wang, Q.; Cole, J.R.; Rosen, G.L. Using the RDP classifier to predict taxonomic novelty and reduce the search space for finding novel organisms. PLoS ONE 2012, 7, e0032491. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. 2017. Available online: https://cran.r-project.org/web/packages/corrplot/index.html (accessed on 21 July 2023).
- Zhou, W.; Lv, D.; Qin, S. Research progress in interaction between plant and rhizosphere microorganism. J. Jilin Agric. Univ. 2016, 38, 253–260. (In Chinese) [Google Scholar]
- Alegria, T.R.; Balbirnie-Cumming, K.; Morris, J.; Hedley, P.E.; Russell, J.; Paterson, E.; Baggs, E.M.; Fridman, E.; Bulgarelli, D. A footprint of plant eco-geographic adaptation on the composition of the barley rhizosphere bacterial microbiota. Sci. Rep. 2020, 10, 12916. [Google Scholar] [CrossRef]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Levy, A.; Gonzalez, I.S.; Mittelviefhaus, M.; Clingenpeel, S.; Paredes, S.H.; Miao, J.M.; Wang, K.R.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef]
- Walters, W.A.; Jin, Z.; Youngblut, N.; Wallace, J.G.; Sutter, J.; Zhang, W.; Gonzalez-Pena, A.; Peiffer, J.; Koren, O.; Shi, Q.J.; et al. Large-scale replicated field study of maize rhizosphere identifies heritable microbes. Proc. Natl. Acad. Sci. USA 2018, 115, 7368–7373. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Kantor, R.S.; Wrighton, K.C.; Handley, K.M.; Sharon, I.; Hug, L.A.; Castelle, C.J.; Thomas, B.C.; Banfield, J.F. Small genomes and sparse metabolisms of sediment-associated bacterial from four candicate phyla. mBio 2013, 4, e00708-13. [Google Scholar] [CrossRef]
- Guo, H.J.; He, X.L.; Li, Y.P. Spatial distribution of arbuscular mycorrhiza and glomalin in the rhizosphere of Caragana korshinskii Kom. in the Otindag sandy land, China. Afr. J. Microbiol. Res. 2012, 6, 5745–5753. [Google Scholar] [CrossRef]
- Tian, L.X.; Wang, Y.H.; Yang, J.X.; Zhang, L.Y.; Feng, B.L. Rhizosphere bacterial community structure of three minor grain crops: A case--study from paired field sites in northern China. Land. Degrad. Dev. 2021, 33, 104–116. [Google Scholar] [CrossRef]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Xue, Y.; Deng, Y.; Li, H.; Zhou, J.; Li, D. Soil bacterial diversity patterns and drivers along an elevational gradient on Shennongjia Mountain, China. Microb. Biotechnol. 2014, 7, 601–609. [Google Scholar] [CrossRef]
- Donn, S.; Kirkegaard, J.A.; Perera, G.; Richardson, A.E.; Watt, M. Evolution of bacterial communities in the wheat crop rhizosphere. Environ. Microbiol. 2015, 17, 610–621. [Google Scholar] [CrossRef]
- Latif, S.; Bibi, S.; Kouser, R.; Fatimah, H.; Farooq, S.; Naseer, S.; Kousar, R. Characterization of bacterial community structure in the rhizosphere of Triticum aestivum L. Genomics 2020, 112, 4760–4768. [Google Scholar] [CrossRef]
- Shetty, S.A.; van Beek, J.; Bijvank, E.; Groot, J.; Kuiling, S.; Bosch, T.; van Baarle, D.; Fuentes, S. Associations and recovery dynamics of the nasopharyngeal microbiota during influenza-like illness in the aging population. Sci. Rep. 2022, 12, 1915. [Google Scholar] [CrossRef]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Variety Name | Code | Variety Name | Code | Variety Name |
|---|---|---|---|---|---|
| R1 | Ganpi No.2 | R13 | Yunpi No.2 | R25 | Edamai 820352 |
| R2 | Ganpi No.3 | R14 | Yunnan S-500 | R26 | Huadamai 1707 |
| R3 | Ganpi No.5 | R15 | Huadamai No.2 | R27 | Huadamai 16316 |
| R4 | Kenpimai No.1 | R16 | Huadamai No.7 | R28 | Huadamai No.13 |
| R5 | Kenpimai No.2 | R17 | Huadamai No.9 | R29 | Huadamai No.15 |
| R6 | Kenpimai No.4 | R18 | Suhua No.2 | R30 | Huadamai No.18 |
| R7 | Kenpimai No.6 | R19 | Kanghanluodamai | R31 | Huadamai No.20 |
| R8 | Kenpimai No.8 | R20 | Hailaerdamai | R32 | Edamai 523898 |
| R9 | Kenpimai No.9 | R21 | Huadamai1539 | R33 | Edamai 720135 |
| R10 | Kenpimai No.10 | R22 | Changfupi No.1 | R34 | Edamai 720033 |
| R11 | Kenjianpi No.3 | R23 | Edamai 522600 | R35 | Huadamai 16312 |
| R12 | Edamai 029 | R24 | Edamai 730135 |
| PH | Ammonium Nitrogen (mg/kg) | Nitrate Nitrogen (mg/kg) | Organic Carbon (‰) | Available Phosphorus (mg/kg) | Available Potassium (mg/kg) | Effective Boron (mg/kg) |
|---|---|---|---|---|---|---|
| 6.3 | 26.92 | 99.78 | 19.92 | 48.57 | 149.4 | 0.12 |
| Phylum Name | Range | Minimum | Maximum | Median | Average | Standard Deviation | Coefficient of Variation |
|---|---|---|---|---|---|---|---|
| Actinobacteriota | 0.1292 | 0.1901 | 0.3193 | 0.2635 | 0.2639 | 0.0327 | 0.1238 |
| Proteobacteria | 0.2028 | 0.1556 | 0.3584 | 0.2212 | 0.2302 | 0.0458 | 0.1988 |
| Acidobacteriota | 0.2138 | 0.0547 | 0.2685 | 0.1531 | 0.1489 | 0.0450 | 0.3025 |
| Chloroflexi | 0.0862 | 0.0921 | 0.1783 | 0.1365 | 0.1328 | 0.0193 | 0.1455 |
| Gemmatimonadota | 0.0502 | 0.0301 | 0.0803 | 0.0510 | 0.0517 | 0.0101 | 0.1959 |
| Firmicutes | 0.0938 | 0.0221 | 0.1159 | 0.0382 | 0.0423 | 0.0179 | 0.4219 |
| Bacteroidota | 0.0747 | 0.0152 | 0.0899 | 0.0386 | 0.0403 | 0.0148 | 0.3678 |
| Myxococcota | 0.0306 | 0.0224 | 0.0530 | 0.0309 | 0.0324 | 0.0064 | 0.1967 |
| Verrucomicrobiota | 0.0183 | 0.0016 | 0.0199 | 0.0098 | 0.0103 | 0.0048 | 0.4700 |
| Planctomycetota | 0.0213 | 0.0024 | 0.0237 | 0.0085 | 0.0098 | 0.0051 | 0.5252 |
| Methylomirabilota | 0.0185 | 0.0023 | 0.0208 | 0.0057 | 0.0071 | 0.0036 | 0.5026 |
| Nitrospirota | 0.0075 | 0.0032 | 0.0107 | 0.0062 | 0.0063 | 0.0019 | 0.3038 |
| Patescibacteria | 0.0088 | 0.0020 | 0.0108 | 0.0049 | 0.0053 | 0.0019 | 0.3652 |
| Class Name | Range | Minimum | Maximum | Median | Average | Standard Deviation | Coefficient of Variation |
|---|---|---|---|---|---|---|---|
| Actinobacteria | 0.0887 | 0.1049 | 0.1936 | 0.1707 | 0.1416 | 0.0226 | 0.1596 |
| Alphaproteobacteria | 0.1314 | 0.0861 | 0.2175 | 0.1305 | 0.1397 | 0.0270 | 0.1932 |
| Gammaproteobacteria | 0.0756 | 0.0653 | 0.1409 | 0.0718 | 0.0904 | 0.0206 | 0.2277 |
| Thermoleophilia | 0.0957 | 0.0449 | 0.1406 | 0.0860 | 0.0877 | 0.0222 | 0.2532 |
| Vicinamibacteria | 0.1589 | 0.0152 | 0.1741 | 0.0893 | 0.0690 | 0.0309 | 0.4478 |
| Chloroflexia | 0.0518 | 0.0376 | 0.0894 | 0.0591 | 0.0664 | 0.0123 | 0.1859 |
| Gemmatimonadetes | 0.0498 | 0.0297 | 0.0795 | 0.0421 | 0.0504 | 0.0099 | 0.1967 |
| Bacteroidia | 0.0745 | 0.0150 | 0.0895 | 0.0326 | 0.0396 | 0.0148 | 0.3744 |
| Bacilli | 0.0915 | 0.0189 | 0.1104 | 0.0474 | 0.0386 | 0.0177 | 0.4586 |
| Blastocatellia | 0.0563 | 0.0097 | 0.0660 | 0.0397 | 0.0386 | 0.0141 | 0.3651 |
| Acidobacteriae | 0.0375 | 0.0190 | 0.0565 | 0.0233 | 0.0306 | 0.0073 | 0.2395 |
| Acidimicrobiia | 0.0188 | 0.0168 | 0.0356 | 0.0207 | 0.0233 | 0.0038 | 0.1613 |
| Polyangia | 0.0149 | 0.0148 | 0.0297 | 0.0258 | 0.0220 | 0.0038 | 0.1748 |
| Anaerolineae | 0.0298 | 0.0086 | 0.0384 | 0.0174 | 0.0212 | 0.0072 | 0.3396 |
| KD4-96 | 0.0269 | 0.0041 | 0.0310 | 0.0181 | 0.0149 | 0.0064 | 0.4293 |
| TK10 | 0.0221 | 0.0064 | 0.0285 | 0.0111 | 0.0130 | 0.0042 | 0.3257 |
| Ktedonobacteria | 0.0156 | 0.0054 | 0.0210 | 0.0106 | 0.0102 | 0.0034 | 0.3344 |
| Verrucomicrobiae | 0.0179 | 0.0012 | 0.0191 | 0.0097 | 0.0095 | 0.0046 | 0.4809 |
| Planctomycetes | 0.0202 | 0.0016 | 0.0218 | 0.0117 | 0.0087 | 0.0048 | 0.5488 |
| MB-A2-108 | 0.0167 | 0.0019 | 0.0186 | 0.0066 | 0.0075 | 0.0040 | 0.5345 |
| Methylomirabilia | 0.0185 | 0.0023 | 0.0208 | 0.0029 | 0.0071 | 0.0036 | 0.5040 |
| Holophagae | 0.0093 | 0.0028 | 0.0121 | 0.0054 | 0.0065 | 0.0019 | 0.2993 |
| Nitrospiria | 0.0075 | 0.0032 | 0.0107 | 0.0032 | 0.0062 | 0.0019 | 0.3027 |
| Myxococcia | 0.0206 | 0.0023 | 0.0229 | 0.0094 | 0.0061 | 0.0036 | 0.5866 |
| Saccharimonadia | 0.0087 | 0.0016 | 0.0103 | 0.0067 | 0.0048 | 0.0019 | 0.3924 |
| Minimum | Maximum | Mean Value | Standard Deviation | Coefficient of Variation | |
|---|---|---|---|---|---|
| sobs | 2062.00 | 2534.00 | 2273.9143 | 128.6018 | 0.0566 |
| Shannon | 6.31 | 6.62 | 6.4766 | 0.0877 | 0.0135 |
| Simpson | 0.00 | 0.01 | 0.0040 | 0.0008 | 0.1925 |
| ace | 2766.30 | 3394.67 | 3033.6982 | 154.3249 | 0.0509 |
| chao | 2731.20 | 3427.66 | 3051.9101 | 154.8281 | 0.0507 |
| coverage | 0.97 | 0.98 | 0.9735 | 0.0022 | 0.0022 |
| Total Variance | Average Variance | F Model | R2 | p | Padjust |
|---|---|---|---|---|---|
| 0.08966 | 0.04483 | 6.57782 | 0.29794 | 0.00813 | 0.0092 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; An, Y.; Zhou, Y.; Wang, X.; Tang, Y.; Zhang, D.; Sun, G.; Wang, Q.; Ren, X. Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy 2023, 13, 2825. https://doi.org/10.3390/agronomy13112825
Zhang S, An Y, Zhou Y, Wang X, Tang Y, Zhang D, Sun G, Wang Q, Ren X. Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy. 2023; 13(11):2825. https://doi.org/10.3390/agronomy13112825
Chicago/Turabian StyleZhang, Siyu, Yue An, Yu Zhou, Xiaofang Wang, Yiqing Tang, Daorong Zhang, Genlou Sun, Qifei Wang, and Xifeng Ren. 2023. "Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage" Agronomy 13, no. 11: 2825. https://doi.org/10.3390/agronomy13112825
APA StyleZhang, S., An, Y., Zhou, Y., Wang, X., Tang, Y., Zhang, D., Sun, G., Wang, Q., & Ren, X. (2023). Bacterial Community Structure in Rhizosphere of Barley at Maturity Stage. Agronomy, 13(11), 2825. https://doi.org/10.3390/agronomy13112825