

Evolving Soil Water Limitation Changes Maize Production Potential and Biomass Accumulation but Not Its Relationship with Grain Yield

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Experimental Design and Field Management
2.3. Measurements of Environmental Variables and Maize Productivity
2.3.1. Meteorological Conditions
2.3.2. Soil Water Availability
2.3.3. Crop Biomass Production and Grain Yield
2.4. Maize Biomass Production Potential
2.4.1. Photosynthetic Production Potential of Biomass
2.4.2. Light–Temperature Production Potential of Biomass
2.4.3. Climatic Production Potential of Biomass
2.5. Data and Statistical Analysis
3. Results
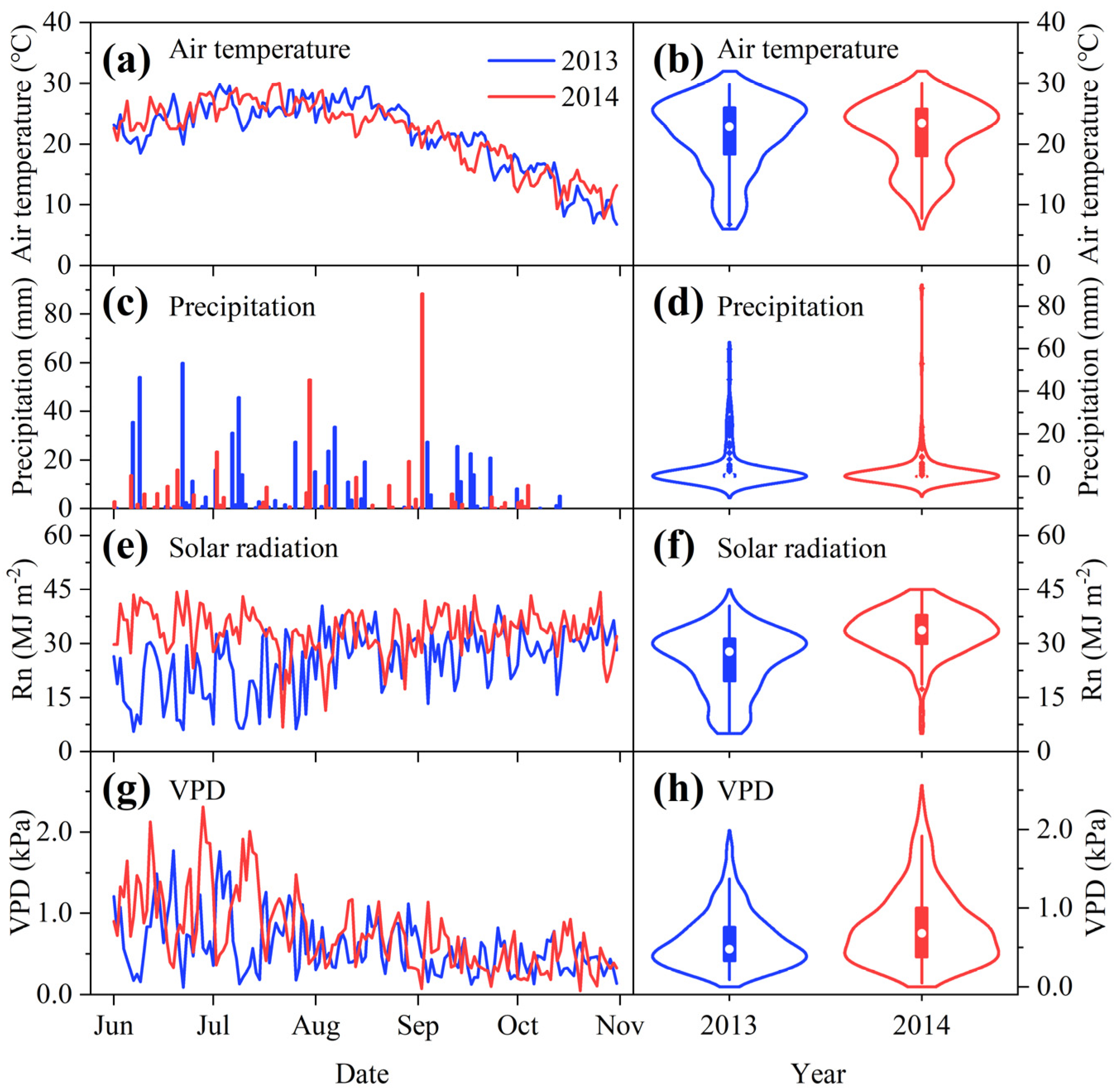
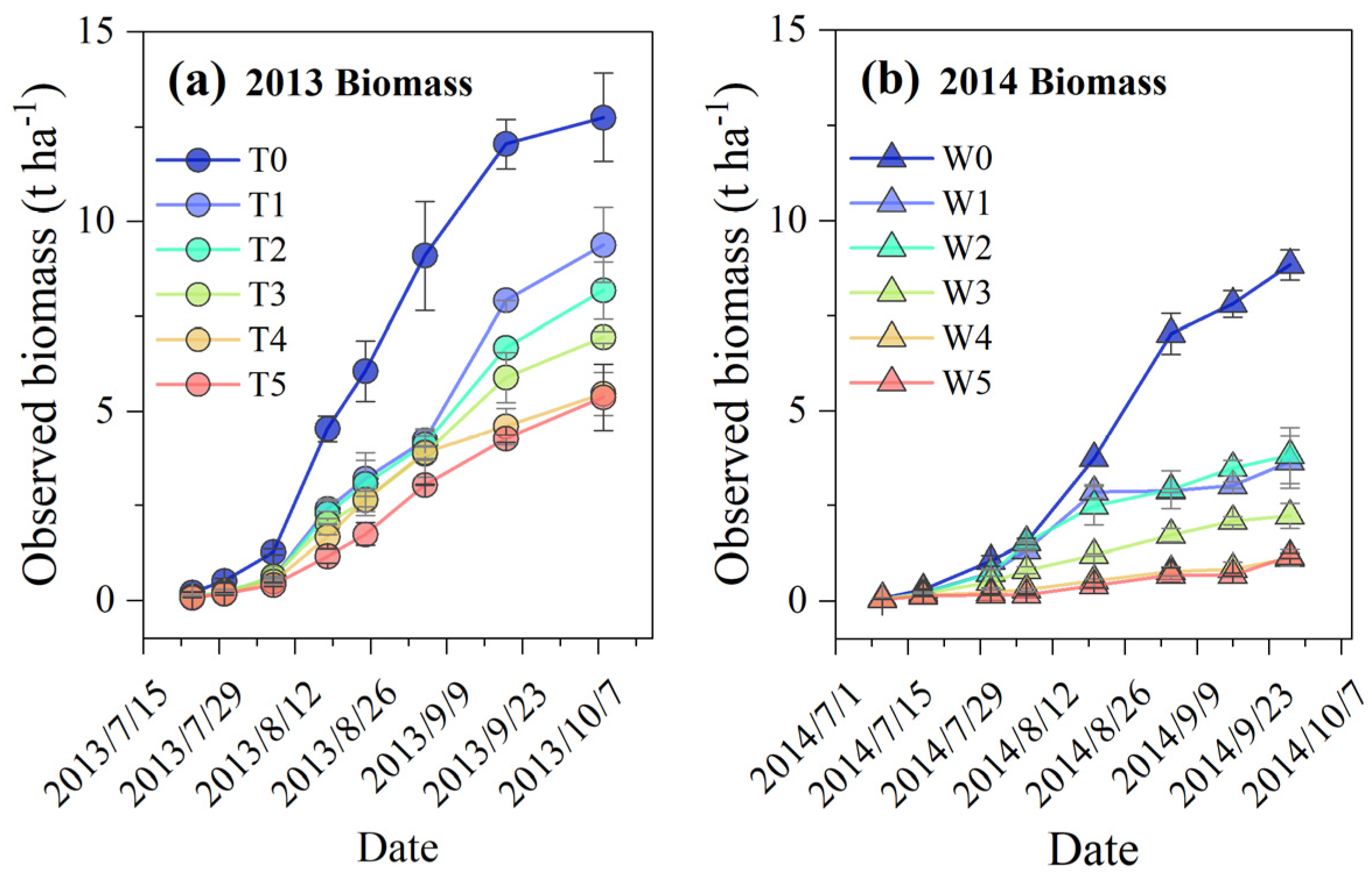
3.1. Meteorological Conditions during the Experimental Growing Seasons
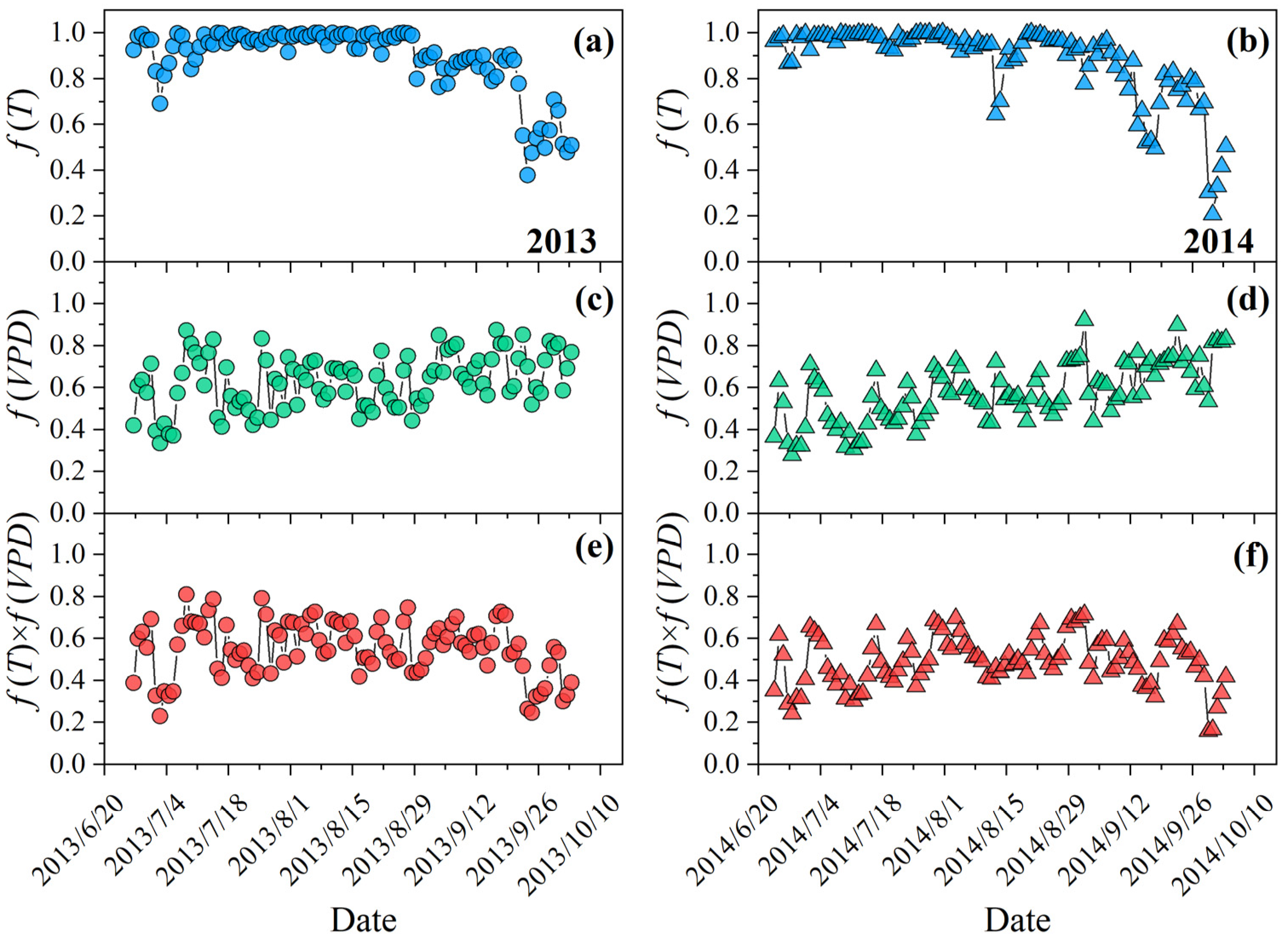
3.2. Dynamics of Downregulation Scalars for the Effects of Temperature and VPD on Climatic Production Potential
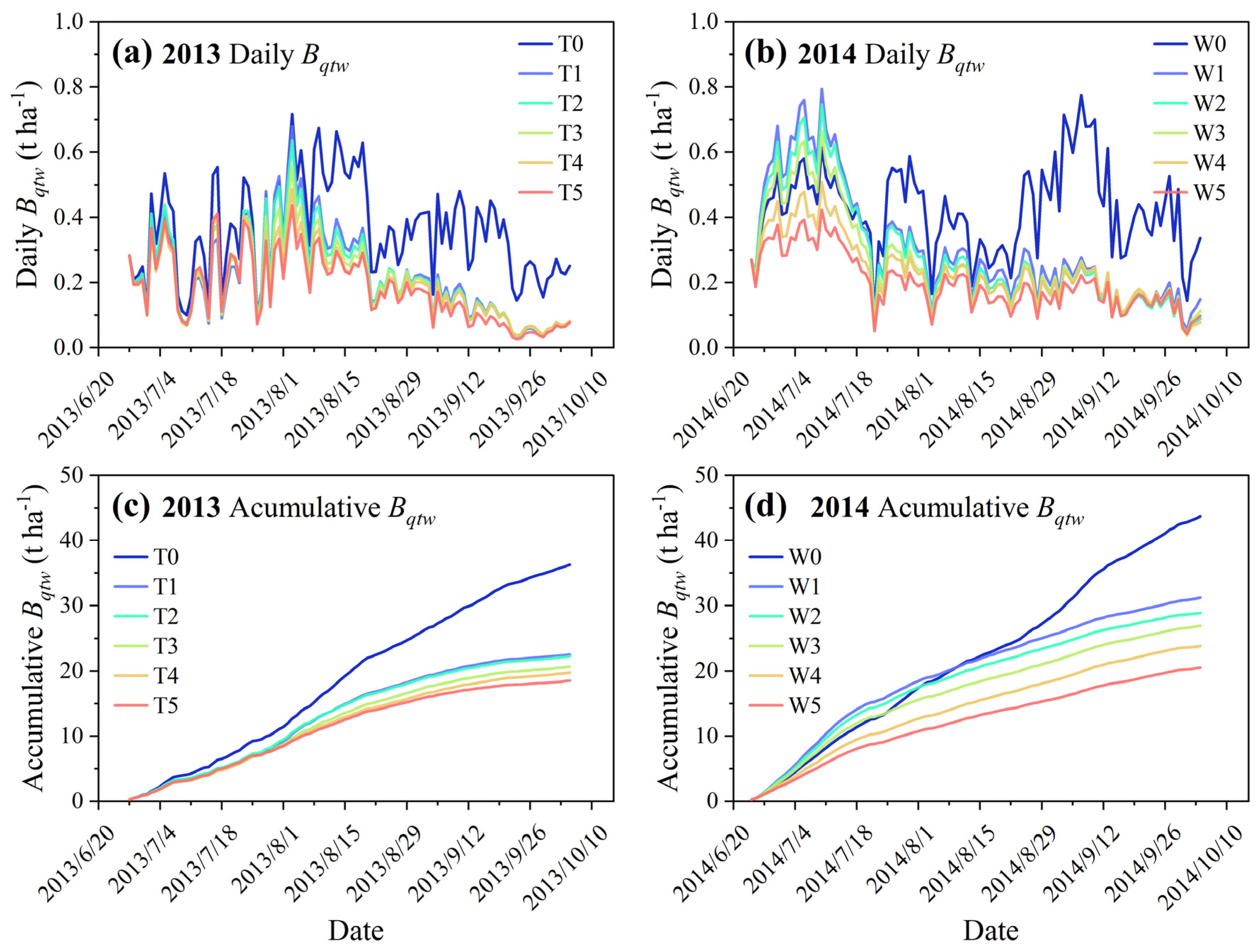
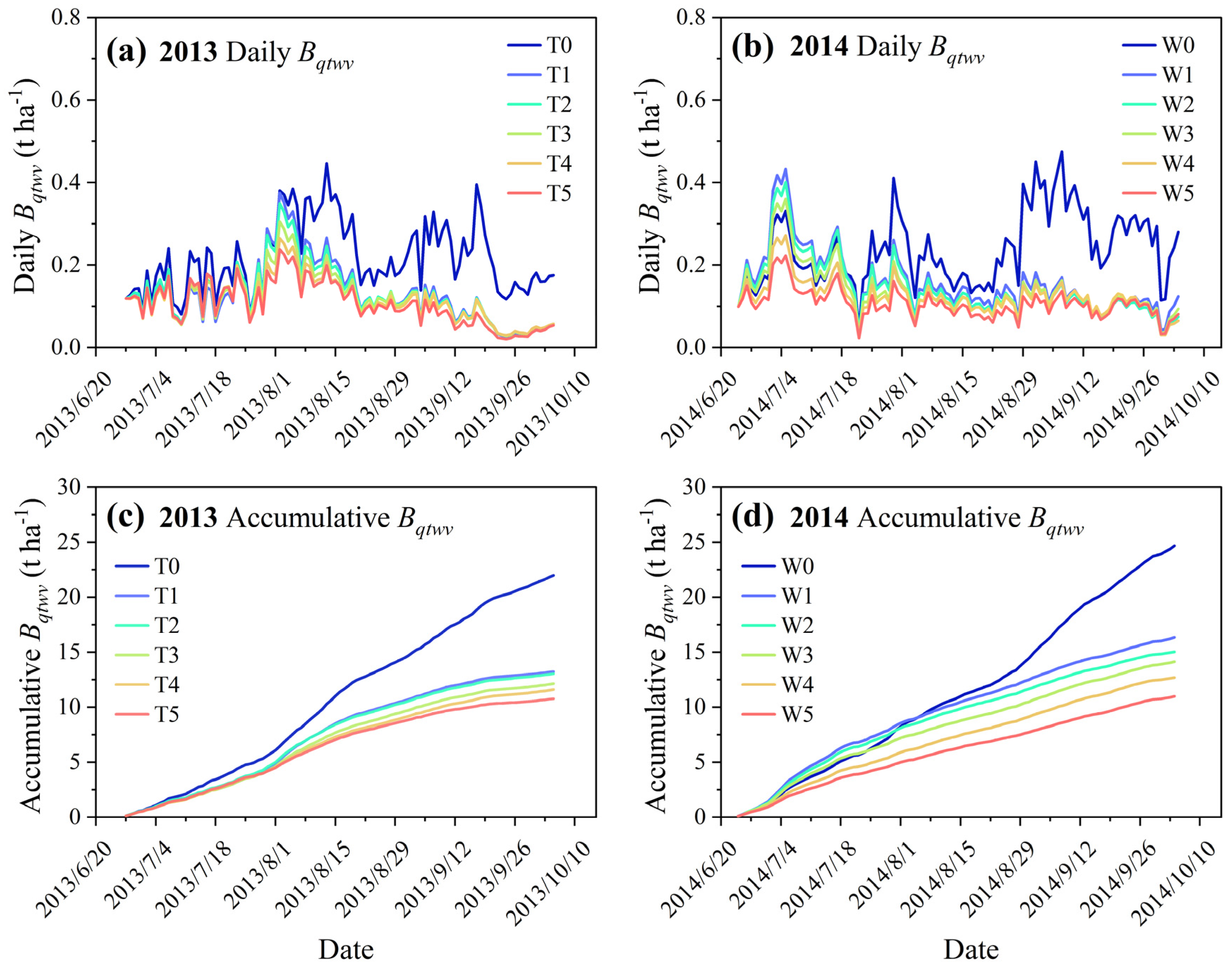
3.3. Climatic Production Potential across Rainfed and Soil Water Deficit Treatments
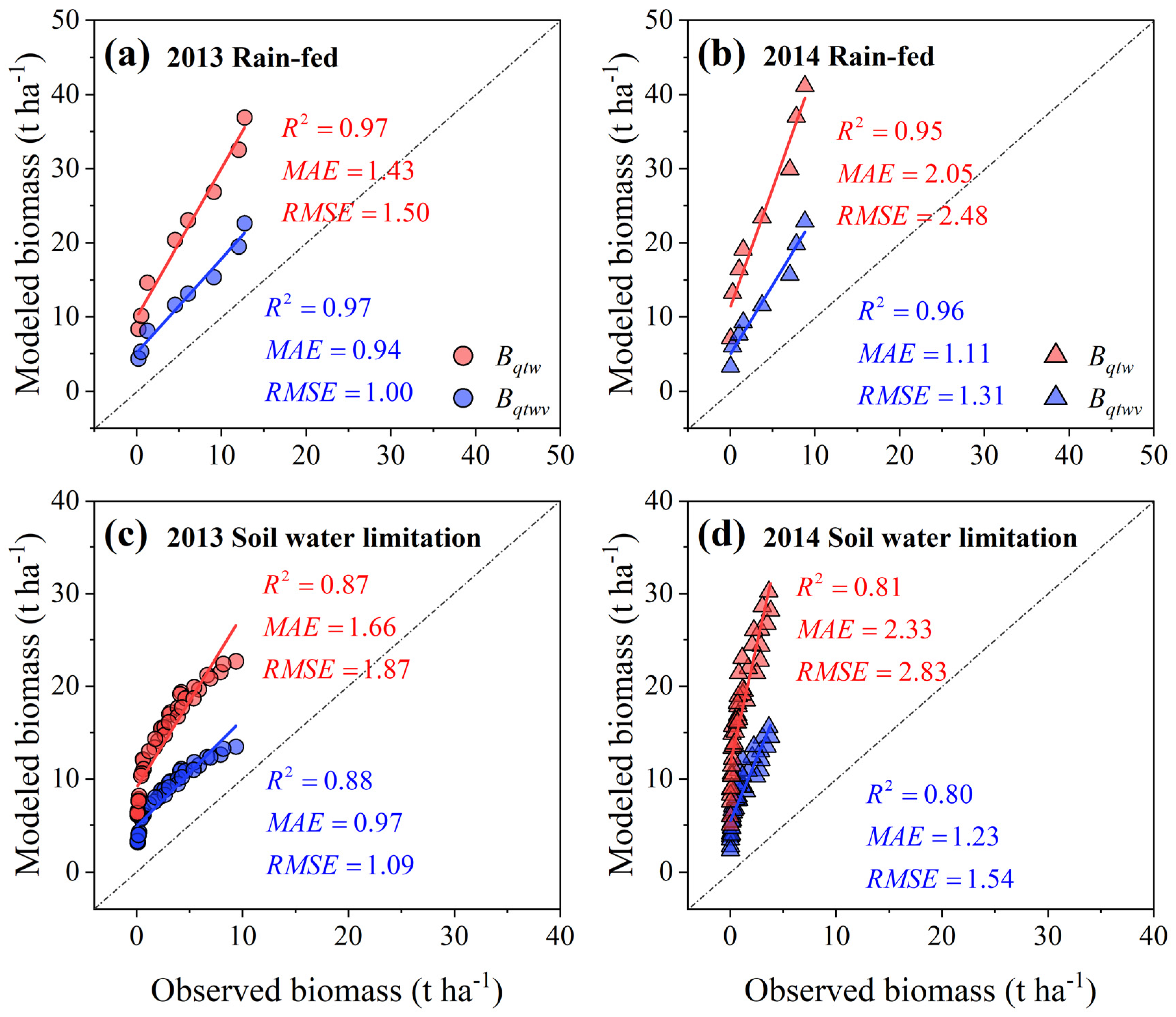
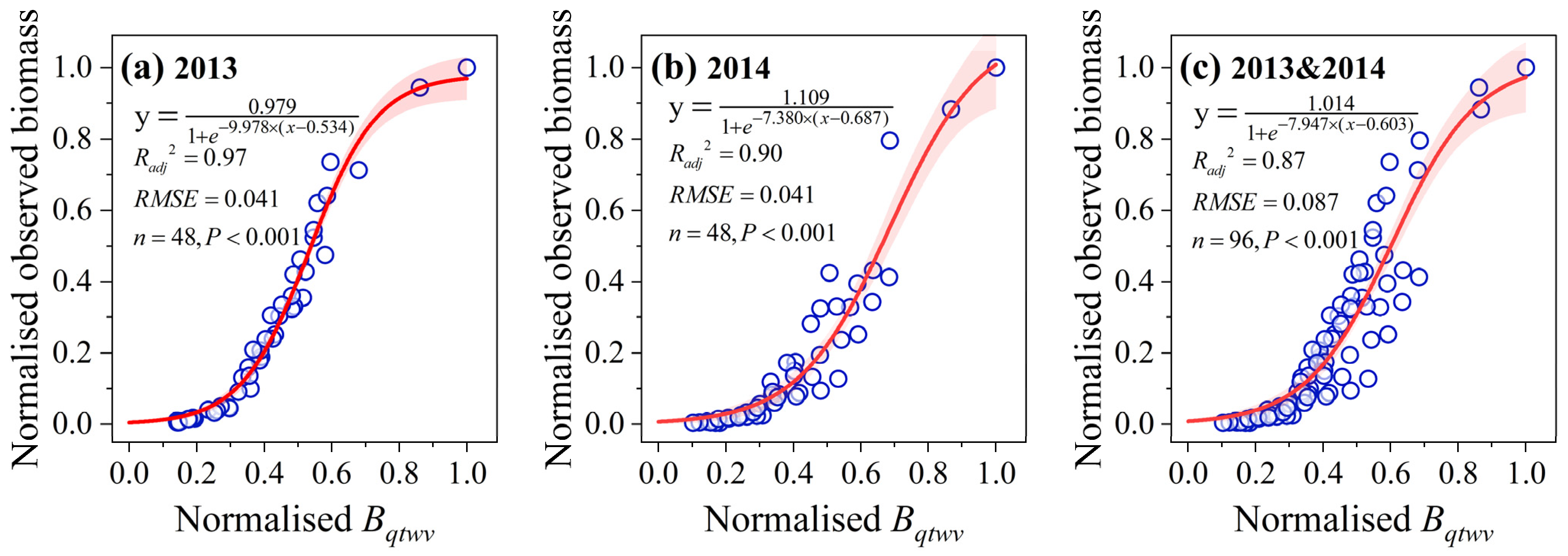
3.4. Effect of Evolving Soil Water Limitation on the Response Pattern of Biomass Accumulation to Climatic Production Potential
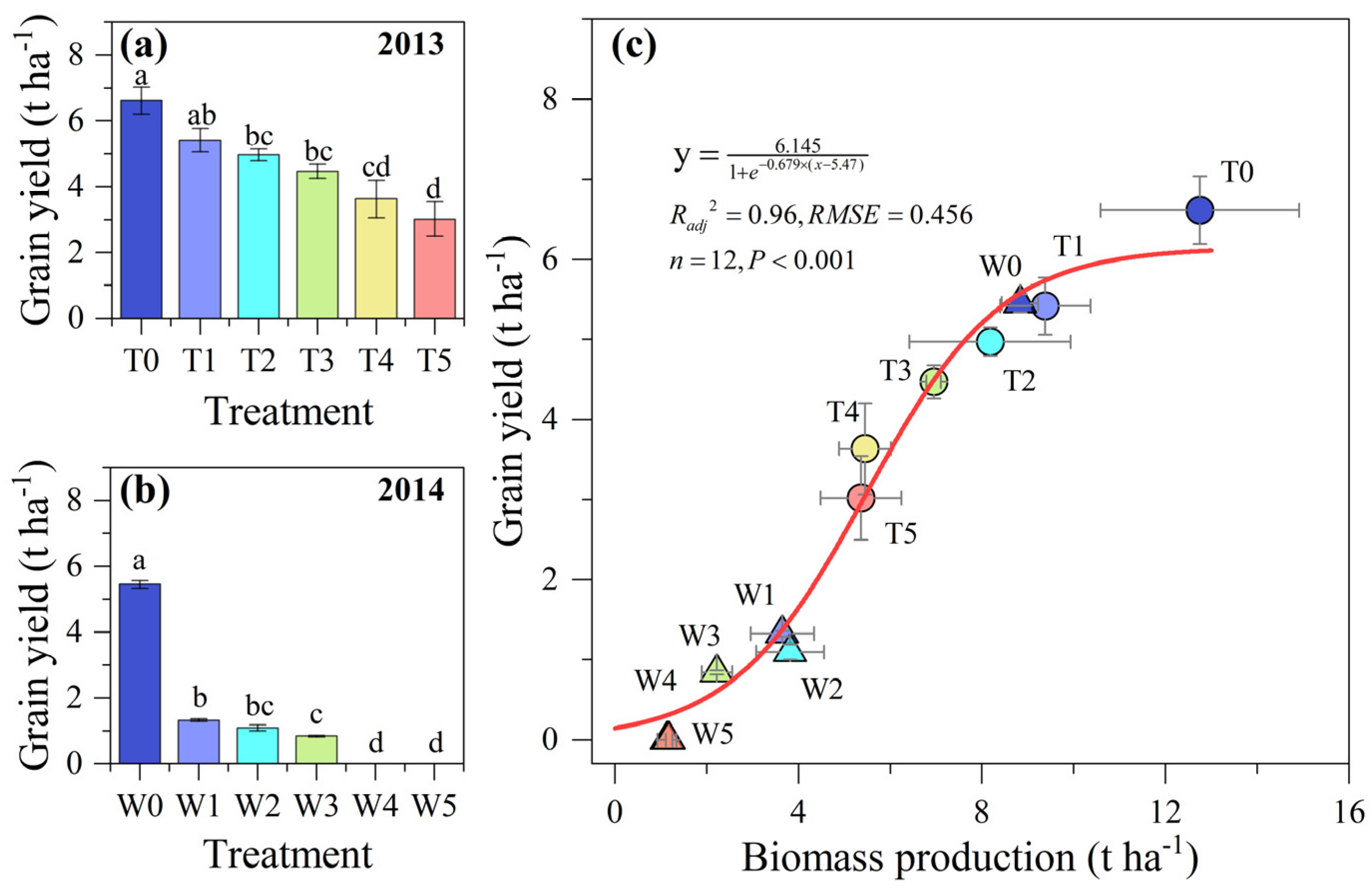
3.5. The Relationship between Biomass and Grain Yield across Rainfed and Evolving Soil Water Limitiation Conditions
4. Discussion
4.1. Climate Change Influences Crop Growth and Production Potential
4.2. Effect of Evolving Soil Water Limitation on Biomass Accumulation and Its Relation to Climatic Production Potential
4.3. The Relationship between Biomass and Grain Yield Affected by Soil Water Condition
4.4. Limitations and Future Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar]
- Saini, A.; Sahu, N.; Nayak, S. Determination of Grid-Wise Monsoon Onset and Its Spatial Analysis for India (1901–2019). Atmosphere 2023, 14, 1424. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2014, 5, 143–147. [Google Scholar] [CrossRef]
- Hsiao, J.; Swann, A.L.S.; Kim, S.-H. Maize yield under a changing climate: The hidden role of vapor pressure deficit. Agric. For. Meteorol. 2019, 279, 107692. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- van Ittersum, M.K.; Rabbinge, R. Concepts in production ecology for analysis and quantification of agricultural input-output combinations. Field Crops Res. 1997, 52, 197–208. [Google Scholar] [CrossRef]
- Evans, L.T. Crop Evolution, Adaptation and Yield; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Yang, X.; Cheng, C.; Li, Y. Effect of cropland occupation and supplement on light-temperature potential productivity in China from 2000 to 2008. Chin. Geogr. Sci. 2010, 20, 536–544. [Google Scholar] [CrossRef]
- He, D.; Wang, J.; Dai, T.; Feng, L.; Zhang, J.; Pan, X.; Pan, Z. Impact of climate change on maize potential productivity and the potential productivity gap in southwest China. J. Meteorolog. Res. 2015, 28, 1155–1167. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Elliott, J.; Deryng, D.; Ruane, A.C.; Müller, C.; Arneth, A.; Boote, K.J.; Folberth, C.; Glotter, M.; Khabarov, N.; et al. Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc. Natl. Acad. Sci. USA 2014, 111, 3268–3273. [Google Scholar] [CrossRef]
- van Wart, J.; Kersebaum, K.C.; Peng, S.; Milner, M.; Cassman, K.G. Estimating crop yield potential at regional to national scales. Field Crop. Res. 2013, 143, 34–43. [Google Scholar] [CrossRef]
- Wang, X.; Li, T.; Yang, X.; Zhang, T.; Liu, Z.; Guo, E.; Liu, Z.; Qu, H.; Chen, X.; Wang, L.; et al. Rice yield potential, gaps and constraints during the past three decades in a climate-changing Northeast China. Agric. For. Meteorol. 2018, 259, 173–183. [Google Scholar] [CrossRef]
- Saini, A.; Sahu, N.; Mishra, S.K.; Jain, S.; Behera, S.; Dash, S.K. The Spatio-Temporal Onset Characteristics of Indian Summer Monsoon Rainfall and Their Relationship with Climate Indices. Atmosphere 2022, 13, 1581. [Google Scholar] [CrossRef]
- de Wit, A.; Boogaard, H.; Fumagalli, D.; Janssen, S.; Knapen, R.; van Kraalingen, D.; Supit, I.; van der Wijngaart, R.; van Diepen, K. 25 years of the WOFOST cropping systems model. Agric. Syst. 2019, 168, 154–167. [Google Scholar] [CrossRef]
- van Diepen, C.A.; Wolf, J.; van Keulen, H.; Rappoldt, C. WOFOST: A simulation model of crop production. Soil Use Manag. 1989, 5, 16–24. [Google Scholar] [CrossRef]
- Jones, J.W.; Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Batchelor, W.D.; Hunt, L.A.; Wilkens, P.W.; Singh, U.; Gijsman, A.J.; Ritchie, J.T. The DSSAT cropping system model. Eur. J. Agron. 2003, 18, 235–265. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S.; Hammer, G.L.; Probert, M.E.; Robertson, M.J.; Holzworth, D.; Huth, N.I.; Hargreaves, J.N.G.; Meinke, H.; Hochman, Z.; et al. An overview of APSIM, a model designed for farming systems simulation. Eur. J. Agron. 2003, 18, 267–288. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; deVoil, P.G.; Zurcher, E.J.; Herrmann, N.I.; McLean, G.; Chenu, K.; van Oosterom, E.J.; Snow, V.; Murphy, C.; et al. APSIM—Evolution towards a new generation of agricultural systems simulation. Environ. Modell. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Brisson, N.; Gary, C.; Justes, E.; Roche, R.; Mary, B.; Ripoche, D.; Zimmer, D.; Sierra, J.; Bertuzzi, P.; Burger, P.; et al. An overview of the crop model stics. Eur. J. Agron. 2003, 18, 309–332. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Raes, D.; Fereres, E. AquaCrop—The FAO crop model to simulate yield response to water: I. Concepts and underlying principles. Agron. J. 2009, 101, 426–437. [Google Scholar] [CrossRef]
- Li, T.; Angeles, O.; Marcaida, M.; Manalo, E.; Manalili, M.P.; Radanielson, A.; Mohanty, S. From ORYZA2000 to ORYZA (v3): An improved simulation model for rice in drought and nitrogen-deficient environments. Agric. For. Meteorol. 2017, 237–238, 246–256. [Google Scholar] [CrossRef]
- Lieth, H. Primary production: Terrestrial ecosystems. Hum. Ecol. 1973, 1, 303–332. [Google Scholar] [CrossRef]
- Lieth, H. Modeling the primary productivity of the world. In Primary Productivity of the Biosphere; Lieth, H., Whittaker, R.H., Eds.; Springer: Berlin/Heidelberg, Germany, 1975; pp. 237–263. [Google Scholar] [CrossRef]
- Fischer, G.; Sun, L. Model based analysis of future land-use development in China. Agric. Ecosyst. Environ. 2001, 85, 163–176. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, X.; Sun, S. Constraints on maize yield and yield stability in the main cropping regions in China. Eur. J. Agron. 2018, 99, 106–115. [Google Scholar] [CrossRef]
- Liang, S.; Li, Y.; Zhang, X.; Sun, Z.; Sun, N.; Duan, Y.; Xu, M.; Wu, L. Response of crop yield and nitrogen use efficiency for wheat-maize cropping system to future climate change in northern China. Agric. For. Meteorol. 2018, 262, 310–321. [Google Scholar] [CrossRef]
- Gao, Z.; Feng, H.-Y.; Liang, X.-G.; Zhang, L.; Lin, S.; Zhao, X.; Shen, S.; Zhou, L.-L.; Zhou, S.-L. Limits to maize productivity in the North China Plain: A comparison analysis for spring and summer maize. Field Crop. Res. 2018, 228, 39–47. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Qiu, X.; Tang, L.; Zhu, Y.; Cao, W.; Liu, L. Quantifying the spatial variation in the potential productivity and yield gap of winter wheat in China. J. Integr. Agric. 2017, 16, 845–857. [Google Scholar] [CrossRef]
- Liu, B.; Chen, X.; Meng, Q.; Yang, H.; van Wart, J. Estimating maize yield potential and yield gap with agro-climatic zones in China—Distinguish irrigated and rainfed conditions. Agric. For. Meteorol. 2017, 239, 108–117. [Google Scholar] [CrossRef]
- Yang, X.; Chen, F.; Lin, X.; Liu, Z.; Zhang, H.; Zhao, J.; Li, K.; Ye, Q.; Li, Y.; Lv, S.; et al. Potential benefits of climate change for crop productivity in China. Agric. For. Meteorol. 2015, 208, 76–84. [Google Scholar] [CrossRef]
- Wang, J.; Wang, E.; Yin, H.; Feng, L.; Zhang, J. Declining yield potential and shrinking yield gaps of maize in the North China Plain. Agric. For. Meteorol. 2014, 195–196, 89–101. [Google Scholar] [CrossRef]
- Hay, R.K.M. Harvest index: A review of its use in plant breeding and crop physiology. Ann. Appl. Biol. 1995, 126, 197–216. [Google Scholar] [CrossRef]
- Bassu, S.; Fumagalli, D.; Toreti, A.; Ceglar, A.; Giunta, F.; Motzo, R.; Zajac, Z.; Niemeyer, S. Modelling potential maize yield with climate and crop conditions around flowering. Field Crop. Res. 2021, 271, 108226. [Google Scholar] [CrossRef]
- Unkovich, M.; Baldock, J.; Forbes, M. Variability in harvest index of grain crops and potential significance for carbon accounting: Examples from australian agriculture. Adv. Agron. 2010, 105, 173–219. [Google Scholar]
- Prihar, S.S.; Stewart, B.A. Using upper-bound slope through origin to estimate genetic harvest index. Agron. J. 1990, 82, 1160–1165. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Schubert, S. Harvest Index of Maize (Zea mays L.): Are There Possibilities for Improvement? Adv. Agron. 2017, 146, 37–82. [Google Scholar]
- Qiao, S.; Wang, H.; Prentice, I.C.; Harrison, S.P. Extending a first-principles primary production model to predict wheat yields. Agric. For. Meteorol. 2020, 287, 107932. [Google Scholar] [CrossRef]
- Fischer, R.A. Definitions and determination of crop yield, yield gaps, and of rates of change. Field Crop. Res. 2015, 182, 9–18. [Google Scholar] [CrossRef]
- van Ittersum, M.K.; Cassman, K.G.; Grassini, P.; Wolf, J.; Tittonell, P.; Hochman, Z. Yield gap analysis with local to global relevance—A review. Field Crop. Res. 2013, 143, 4–17. [Google Scholar] [CrossRef]
- Meng, Q.; Hou, P.; Wu, L.; Chen, X.; Cui, Z.; Zhang, F. Understanding production potentials and yield gaps in intensive maize production in China. Field Crop. Res. 2013, 143, 91–97. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Y.; Huang, S.; Xu, C.; Wang, X.; Gao, J.; Meng, Q.; Wang, P. The impact of drought and heat stress at flowering on maize kernel filling: Insights from the field and laboratory. Agric. For. Meteorol. 2022, 312, 108733. [Google Scholar] [CrossRef]
- Gabaldón-Leal, C.; Webber, H.; Otegui, M.E.; Slafer, G.A.; Ordóñez, R.A.; Gaiser, T.; Lorite, I.J.; Ruiz-Ramos, M.; Ewert, F. Modelling the impact of heat stress on maize yield formation. Field Crop. Res. 2016, 198, 226–237. [Google Scholar] [CrossRef]
- Tian, B.; Zhu, J.; Nie, Y.; Xu, C.; Meng, Q.; Wang, P. Mitigating heat and chilling stress by adjusting the sowing date of maize in the North China Plain. J. Agron. Crop Sci. 2019, 205, 77–87. [Google Scholar] [CrossRef]
- Ling, M.; Han, H.; Wei, X.; Lv, C. Temporal and spatial distributions of precipitation on the Huang-Huai-Hai Plain during 1960–2019, China. J. Water Clim. Chang. 2021, 12, 2232–2244. [Google Scholar] [CrossRef]
- Xue, W.; Guo, J.; Zhang, Y.; Zhou, S.; Wang, Y.; Miao, Y.; Liu, L.; Xu, H.; Li, J.; Chen, D.; et al. Declining diurnal temperature range in the North China Plain related to environmental changes. Clim. Dyn. 2019, 52, 6109–6119. [Google Scholar] [CrossRef]
- Meng, Q.; Liu, B.; Yang, H.; Chen, X. Solar dimming decreased maize yield potential on the North China Plain. Food Energy Secur. 2020, 9, e235. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.K.; Lei, P.; Shi, D.; Hashimi, M.H.; Wang, S.; Xie, D.; Ni, J.; Ni, C. Response of maize (Zea mays L.) towards vapor pressure deficit. Environ. Exp. Bot. 2021, 181, 104293. [Google Scholar] [CrossRef]
- Fu, Z.; Ciais, P.; Prentice, I.C.; Gentine, P.; Makowski, D.; Bastos, A.; Luo, X.; Green, J.K.; Stoy, P.C.; Yang, H.; et al. Atmospheric dryness reduces photosynthesis along a large range of soil water deficits. Nat. Commun. 2022, 13, 989. [Google Scholar] [CrossRef]
- Li, Q.; Wei, M.; Li, Y.; Feng, G.; Wang, Y.; Li, S.; Zhang, D. Effects of soil moisture on water transport, photosynthetic carbon gain and water use efficiency in tomato are influenced by evaporative demand. Agric. Water Manag. 2019, 226, 105818. [Google Scholar] [CrossRef]
- Craufurd, P.Q.; Wheeler, T.R. Climate change and the flowering time of annual crops. J. Exp. Bot. 2009, 60, 2529–2539. [Google Scholar] [CrossRef]
- Tao, F.; Zhang, Z. Adaptation of maize production to climate change in North China Plain: Quantify the relative contributions of adaptation options. Eur. J. Agron. 2010, 33, 103–116. [Google Scholar] [CrossRef]
- Ma, X.; He, Q.; Zhou, G. Sequence of Changes in Maize Responding to Soil Water Deficit and Related Critical Thresholds. Front. Plant Sci. 2018, 9, 511. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, G.; Song, X.; He, Q. Dynamic characteristics of canopy and vegetation water content during an entire maize growing season in relation to spectral-based indices. Remote Sens. 2022, 14, 584. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, G.; He, Q.; Zhou, L.; Ji, Y.; Zhou, M. Environmental explanation of maize specific leaf area under varying water stress regimes. Environ. Exp. Bot. 2020, 171, 103932. [Google Scholar] [CrossRef]
- Campbell, G.S.; Norman, J.M. Water Vapor and Other Gases. In An Introduction to Environmental Biophysics; Campbell, G.S., Norman, J.M., Eds.; Springer: New York, NY, USA, 1998; pp. 37–51. [Google Scholar] [CrossRef]
- Wang, S.; Huang, Y.; Sun, W.; Yu, L. Mapping the vertical distribution of maize roots in China in relation to climate and soil texture. J. Plant Ecol. 2018, 11, 899–908. [Google Scholar] [CrossRef]
- Anyia, A.O.; Herzog, H. Water-use efficiency, leaf area and leaf gas exchange of cowpeas under mid-season drought. Eur. J. Agron. 2004, 20, 327–339. [Google Scholar] [CrossRef]
- Yuan, B.; Guo, J.; Ye, M.; Zhao, J. Variety distribution pattern and climatic potential productivity of spring maize in Northeast China under climate change. Chin. Sci. Bull. 2012, 57, 3497–3508. [Google Scholar] [CrossRef][Green Version]
- Loomis, R.S.; Williams, W.A. Maximum crop productivity: An estimate. Crop Sci. 1963, 3, 67–72. [Google Scholar] [CrossRef]
- Jiang, X.; Tang, L.; Liu, X.; Cao, W.; Zhu, Y. Spatial and temporal characteristics of rice potential productivity and potential yield increment in main production regions of China. J. Integr. Agric. 2013, 12, 45–56. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, J.; Xu, Y.; Mu, J. Effects of climate change on cultivation patterns of spring maize and its climatic suitability in Northeast China. Agric. Ecosyst. Environ. 2015, 202, 178–187. [Google Scholar] [CrossRef]
- Qing, L.; Jianping, G.; Yali, M.; Xiangnan, M.; Limin, Z.; Mengwei, L. Evaluation of climate resource utilization rate of main grain crops in Shanxi Province. Chin. J. Ecol. 2021, 40, 1386–1396. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Patanè, C.; Sanzone, E.; Testa, G.; Scordia, D. Leaf gas exchange, water status and radiation use efficiency of giant reed (Arundo donax L.) in a changing soil nitrogen fertilization and soil water availability in a semi-arid Mediterranean area. Eur. J. Agron. 2016, 72, 56–69. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Cao, G.; Ma, Z.; Li, Y.; Zhang, F.; Zhao, X.; Zhao, X.; Jiang, L.; Sanders, N.J.; et al. Alpine grassland plants grow earlier and faster but biomass remains unchanged over 35 years of climate change. Ecol. Lett. 2020, 23, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Paine, C.E.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Huang, L.; Koubek, T.; Weiser, M.; Herben, T.; Cornelissen, H. Environmental drivers and phylogenetic constraints of growth phenologies across a large set of herbaceous species. J. Ecol. 2018, 106, 1621–1633. [Google Scholar] [CrossRef]
- Lizaso, J.I.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.A.; Lorite, I.J.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crop. Res. 2018, 216, 129–140. [Google Scholar] [CrossRef]
- Wild, M.; Gilgen, H.; Roesch, A.; Ohmura, A.; Long, C.N.; Dutton, E.G.; Forgan, B.; Kallis, A.; Russak, V.; Tsvetkov, A. From dimming to brightening: Decadal changes in solar radiation at Earth’s surface. Science 2005, 308, 847–850. [Google Scholar] [CrossRef]
- Gupta, R.; Somanathan, E.; Dey, S. Global warming and local air pollution have reduced wheat yields in India. Clim. Chang. 2017, 140, 593–604. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, S.; Zhang, F.; Guo, S.; Guo, E.; Liu, Z.; Zhao, J.; Zhao, C.; Li, T.; Yang, X. Using estimated radiation in crop models amplified the negative impacts of climate variability on maize and winter wheat yields in China. Agric. For. Meteorol. 2022, 318, 108914. [Google Scholar] [CrossRef]
- Tollenaar, M.; Fridgen, J.; Tyagi, P.; Stackhouse, P.W., Jr.; Kumudini, S. The Contribution of Solar Brightening to the US Maize Yield Trend. Nat. Clim. Chang. 2017, 7, 275–278. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef]
- Schauberger, B.; Archontoulis, S.; Arneth, A.; Balkovic, J.; Ciais, P.; Deryng, D.; Elliott, J.; Folberth, C.; Khabarov, N.; Müller, C.; et al. Consistent negative response of US crops to high temperatures in observations and crop models. Nat. Commun. 2017, 8, 13931. [Google Scholar] [CrossRef]
- L Hoffman, A.; R Kemanian, A.; E Forest, C. The response of maize, sorghum, and soybean yield to growing-phase climate revealed with machine learning. Environ. Res. Lett. 2020, 15, 094013. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Monteith, J.L. A reinterpretation of stomatal responses to humidity. Plant Cell Environ. 1995, 18, 357–364. [Google Scholar] [CrossRef]
- Verbraeken, L.; Wuyts, N.; Mertens, S.; Cannoot, B.; Maleux, K.; Demuynck, K.; De Block, J.; Merchie, J.; Dhondt, S.; Bonaventure, G.; et al. Drought affects the rate and duration of organ growth but not inter-organ growth coordination. Plant Physiol. 2021, 186, 1336–1353. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 00272. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Inze, D.; Nelissen, H.; Saibo, N.J.M. Source-Sink Regulation in Crops under Water Deficit. Trends Plant Sci. 2019, 24, 652–663. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 01147. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought Stress in Plants: An Overview. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar] [CrossRef]
- Araus, J.L.; Serret, M.D.; Edmeades, G.O. Phenotyping maize for adaptation to drought. Front. Physiol. 2012, 3, 00305. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Sun, H.; Shao, L.; Wang, Y. Changes in evapotranspiration over irrigated winter wheat and maize in North China Plain over three decades. Agric. Water Manag. 2011, 98, 1097–1104. [Google Scholar] [CrossRef]
- Borras, L.; Vitantonio-Mazzini, L.N. Maize reproductive development and kernel set under limited plant growth environments. J. Exp. Bot. 2018, 69, 3235–3243. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Borrás, L.; De Bruin, J.; Pedersen, P. Physiological strategies for seed number determination in soybean: Biomass accumulation, partitioning and seed set efficiency. Field Crop. Res. 2012, 135, 58–66. [Google Scholar] [CrossRef]
- van Oosterom, E.J.; Hammer, G.L. Determination of grain number in sorghum. Field Crop. Res. 2008, 108, 259–268. [Google Scholar] [CrossRef]
- Miralles, D.J.; Katz, S.D.; Colloca, A.; Slafer, G.A. Floret development in near isogenic wheat lines differing in plant height. Field Crop. Res. 1998, 59, 21–30. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | T0 | T1 | T2 | T3 | T4 | T5 | |
|---|---|---|---|---|---|---|---|
| 2013 | Irrigation amount (mm) | No, with 421 mm rainfall | 80 | 60 | 40 | 25 | 15 |
| Proportion of precipitation in July (%) | 280.7 | 53.3 | 40 | 26.7 | 16.7 | 10 | |
| Treatment | W0 | W1 | W2 | W3 | W4 | W5 | |
| 2014 | Irrigation amount (mm) | No, with 292 mm rainfall | 150 | 120 | 90 | 60 | 30 |
| Proportion of precipitation in July (%) | 194.7 | 100 | 80 | 60 | 40 | 20 |
| Parameter | Value | Meaning | Unit |
|---|---|---|---|
| 49 | Fraction of photosynthetically active radiation | % | |
| 22.4 | Light quantum efficiency | % | |
| 8 | Plant population reflectance | % | |
| 6 | Plant population transmittance | % | |
| 10 | Fraction of radiation intercepted by crop nonphotosynthetic organs | % | |
| 1 | Ratio of light beyond the light saturation point | % | |
| ω | 30 | Fraction of photosynthetic products consumed by respiration | % |
| 17.2 | Heat content per unit dry matter | MJ kg−1 | |
| 0.58 | Revised factor for dynamic change in crop leaf area | - | |
| 8 | Crop ash content | % |
| Growth Stages | (°C) | (°C) | (°C) |
|---|---|---|---|
| Sowing–emergence | 14 | 25 | 32 |
| Emergence–jointing | 14 | 27 | 35 |
| Jointing–tasseling | 17 | 27 | 35 |
| Tasseling–maturity | 10 | 26 | 32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Zhou, G.; Song, X.; Geng, J.; He, Q. Evolving Soil Water Limitation Changes Maize Production Potential and Biomass Accumulation but Not Its Relationship with Grain Yield. Agronomy 2023, 13, 2637. https://doi.org/10.3390/agronomy13102637
Zhou H, Zhou G, Song X, Geng J, He Q. Evolving Soil Water Limitation Changes Maize Production Potential and Biomass Accumulation but Not Its Relationship with Grain Yield. Agronomy. 2023; 13(10):2637. https://doi.org/10.3390/agronomy13102637
Chicago/Turabian StyleZhou, Huailin, Guangsheng Zhou, Xingyang Song, Jinjian Geng, and Qijin He. 2023. "Evolving Soil Water Limitation Changes Maize Production Potential and Biomass Accumulation but Not Its Relationship with Grain Yield" Agronomy 13, no. 10: 2637. https://doi.org/10.3390/agronomy13102637
APA StyleZhou, H., Zhou, G., Song, X., Geng, J., & He, Q. (2023). Evolving Soil Water Limitation Changes Maize Production Potential and Biomass Accumulation but Not Its Relationship with Grain Yield. Agronomy, 13(10), 2637. https://doi.org/10.3390/agronomy13102637