
Updates and Prospects: Morphological, Physiological, and Molecular Regulation in Crop Response to Waterlogging Stress


Abstract
:1. Introduction
2. Morphological Adaptation Mechanism of Plants in Response to Waterlogging Stress
3. Physiological Changes and Metabolic Regulation of Plants in Response to Waterlogging Stress
3.1. Hypoxic Response of Plants to Waterlogging Stress
3.2. Osmotic Regulation of Plants in Response to Waterlogging Stress
3.3. Active Oxygen Scavenging Mechanism of Plants in Response to Waterlogging Stress
3.4. Photosynthetic Signal Transduction in Response to Waterlogging Stress in Plants
3.5. Changes of Plant Hormone and Signal Molecules-Mediated Pathways under Waterlogging Stress
3.6. Energy Metabolism of Plants in Response to Waterlogging Stress
3.7. Nitrogen Uptake and Utilization under Waterlogging Stress
4. Transcription Factors in Plant Response to Waterlogging Stress
5. Stress Memory and Cross-Stress in Plants
5.1. Waterlogging Stress Memory in Plants
5.2. Effect of Temperature on Waterlogging
6. Challenges and Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AR | Adventitial roots |
| ROS | Reactive oxygen species |
| ROL | Radial oxygen loss |
| BR | Brassinolide |
| TCA | Tricarboxylic acid |
| ADH | Ethanol dehydrogenase |
| PCD | Programmed cell death |
| PDC | Pyruvate decarboxylase |
| ERF-VII | Ethylene response factor VII |
| QTL | Quantitative trait loci |
| XET | Xyloglucan endotransglucosylase |
| LDH | Lactate dehydrogenase |
| ADH | Ethanol dehydrogenase |
| QTL | Quantitative trait loci |
| MDA | Malondialdehyde |
| SOD | Superoxide dismutase |
| POD | Peroxidase |
| CAT | Catalase |
| GSH | Glutathione |
| GB | Glycine betaine |
| GABA | Gamma-aminobutyric acid |
| LACS2 | Long-chain acyl-CoA synthetase |
| PUB44 | PLANT U-BOX44 |
| CPK4 | Calc-dependent protein kinase |
| HvPgb1 | Phytoglobin 1 in Hordeum vulgare |
| mETC | Mitochondrial electron transport chain |
| SIRK | Senescence-induced receptor-like protein kinase |
| OsPIL1 | Photopigment-interacting factor-like protein |
| Chl | Chlorophyll |
| PEP | Phosphoenolpyruvate |
| LGF1 | Leaf gas film 1 |
| PCO | Cysteine oxidase |
| PGB1 | PHYTOGLOBIN1 |
| IAA | Indole acetic acid |
| ABA | Abscisic acid |
| GA | Gibberellin |
| SLR1 | SLENDER rice-1 |
| SLRL1 | SLR1-like-1 |
| SD1 | SEMIDWARF1 |
| H2S | Hydrogen sulfide |
| SUS | Sucrose synthetase |
| PA | Phosphatidyl acid |
| NR | Nitrate reductase |
| NO | Nitric oxide |
| ASA | Ascorbic acid |
| COP1 | Constitutive photomorphogenesis 1 |
| VTC1 | vitamin C deficiency 1 |
| GSH1 | Glutathione synthase 1 |
| SHYG | SPEEDY HYPONASTIC GROWTH |
| ACO5 | ACC oxidase5 |
| bHLH | Basic helix-loop-helix |
| HBI1 | HTH IBH 1 |
| GR | Glutathione reductase |
| GWA | Genome-wide association |
References
- Sharkey, T.D. The Physiology of Plants under Stress: Abiotic Factors; Nilsen, E.T., Orcutt, D.M., Eds.; The Quarterly Review of Biology; The University of Chicago Press: Chicago, IL, USA, 1996; Volume 72, pp. 476–477. ISBN 0-471-03512-6. [Google Scholar]
- Choudhary, P.; Muthamilarasan, M. Modulating physiological and transcriptional regulatory mechanisms for enhanced climate resilience in cereal crops. J. Plant Physiol. 2022, 278, 153815. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wang, S.; Yang, P.; Tao, Y. Experimental and modeling evaluation of siphon-type subsurface drainage performance in flooding and waterlogging removal. Agric. Water Manag. 2023, 275, 108031. [Google Scholar] [CrossRef]
- Xu, X.; Ji, J.; Xu, Q.; Qi, X.; Weng, Y.; Chen, X. The major-effect quantitative trait locus CsARN6.1 encodes an AAA ATPase domain-containing protein that is associated with waterlogging stress tolerance by promoting adventitious root formation. Plant J. 2018, 93, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Pais, I.P.; Moreira, R.; Semedo, J.N.; Ramalho, J.C.; Lidon, F.C.; Coutinho, J.; Macas, B.; Scotti-Campos, P. Wheat crop under waterlogging: Potential soil and plant effects. Plants 2022, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Anuradha, M.; Sivaraju, K.; Krishnamurthy, V. Effect of waterlogging on physiological characteristics, yield and quality of flue-cured tobacco. Indian J. Plant Physiol. 2013, 18, 67–70. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Gibbs, D.J.; Holdsworth, M.J.; Lee, S.C.; Licausi, F.; Perata, P.; Voesenek, L.A.; Van Dongen, J.T. Making sense of low oxygen sensing. Trends Plant Sci. 2012, 17, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Najeeb, U.; Bange, M.P.; Tan, D.K.Y.; Atwell, B.J. Consequences of waterlogging in cotton and opportunities for mitigation of yield losses. AoB Plants 2015, 7, plv080. [Google Scholar] [CrossRef] [PubMed]
- De San Celedonio, R.P.; Abeledo, L.G.; Miralles, D.J. Physiological traits associated with reductions in grain number in wheat and barley under waterlogging. Plant Soil 2018, 429, 469–481. [Google Scholar] [CrossRef]
- Zhen, B.; Li, H.; Niu, Q.; Qiu, H.; Tian, G.; Lu, H.; Zhou, X. Effects of combined high temperature and waterlogging stress at booting stage on root anatomy of rice (Oryza sativa L.). Water 2020, 12, 2524. [Google Scholar] [CrossRef]
- Shao, J.; Liu, P.; Zhao, B.; Zhang, J.; Zhao, X.; Ren, B. Combined effects of high temperature and waterlogging on yield and stem development of summer maize. Crop J. 2022, 11, 651–660. [Google Scholar] [CrossRef]
- Ren, B.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J. Responses of photosynthetic characteristics and leaf senescence in summer maize to simultaneous stresses of waterlogging and shading. Crop J. 2023, 11, 269–277. [Google Scholar] [CrossRef]
- Song, M.; Li, X.; Saikkonen, K.; Li, C.; Nan, Z. An asexual epichloë endophyte enhances waterlogging tolerance of Hordeum brevisubulatum. Fungal Ecol. 2015, 13, 44–52. [Google Scholar] [CrossRef]
- Ashraf, M.A. Waterlogging stress in plants: A review. Afr. J. Agric. Res. 2012, 7, 1976–1981. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Oladosu, Y.; Rafii, M.Y.; Arolu, F.; Chukwu, S.C.; Muhammad, I.; Kareem, I.; Salisu, M.A.; Arolu, I.W. Submergence tolerance in rice: Review of mechanism, breeding, and future prospects. Sustainability 2020, 12, 1632. [Google Scholar] [CrossRef]
- Vidoz, M.L.; Mignolli, F.; Aispuru, H.T.; Mroginski, L.A. Rapid formation of adventitious roots and partial ethylene sensitivity result in faster adaptation to flooding in the aerial roots (aer) mutant of tomato. Sci. Hortic. 2016, 201, 130–139. [Google Scholar] [CrossRef]
- Qian, L.; Chen, X.; Wang, X.; Huang, S.; Luo, Y. The effects of flood, drought, and flood followed by drought on yield in cotton. Agronomy 2020, 10, 555. [Google Scholar] [CrossRef]
- Negi, S.; Ivanchenko, M.G.; Muday, G.K. Ethylene regulates lateral root formation and auxin transport in Arabidopsis thaliana. Plant J. 2008, 55, 175–187. [Google Scholar] [CrossRef]
- Sasidharan, R.; Voesenek, L.A. Ethylene-mediated acclimations to flooding stress. Plant Physiol. 2015, 169, 3–12. [Google Scholar] [CrossRef]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef]
- Mergemann, H.; Sauter, M. Ethylene induces epidermal cell death at the site of adventitious root emergence in rice. Plant Physiol. 2000, 124, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, Q.; Ma, X.; Qian, C.; Wang, H.; Ren, N.; Shen, C.; Huang, S.; Xu, X.; Xu, Q.; et al. Waterlogging-induced adventitious root formation in cucumber is regulated by ethylene and auxin through reactive oxygen species signalling. Plant Cell Environ. 2019, 42, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Che, Q.; Su, S.; Liu, Y.; Wang, Y.; Xu, X. Genome-wide identification and characterization of Respiratory Burst Oxidase Homolog genes in six Rosaceae species and an analysis of their effects on adventitious rooting in apple. PLoS ONE 2020, 15, e0239705. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Liang, K.; Fang, T.; Zhao, H.; Han, X.; Cai, M.; Qiu, F. A group VII ethylene response factor gene, ZmEREB180, coordinates waterlogging tolerance in maize seedlings. Plant Biotechnol. J. 2019, 17, 2286–2298. [Google Scholar] [CrossRef] [PubMed]
- Eysholdt-Derzso, E.; Sauter, M. Hypoxia and the group VII ethylene response transcription factor HRE2 promote adventitious root elongation in Arabidopsis. Plant Biol. 2019, 21 (Suppl. S1), 103–108. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, S.; Yoshioka, T.; Yamamoto, R.; Hiraga, S.; Nakamura, T.; Shimada, S.; Komatsu, S. Role of abscisic acid in flood-induced secondary aerenchyma formation in soybean (Glycine max) hypocotyls. Plant Prod. Sci. 2014, 17, 131–137. [Google Scholar] [CrossRef]
- Kim, Y.H.; Hwang, S.J.; Waqas, M.; Khan, A.L.; Lee, J.H.; Lee, J.D.; Nguyen, H.T.; Lee, I.J. Comparative analysis of endogenous hormones level in two soybean (Glycine max L.) lines differing in waterlogging tolerance. Front. Plant Sci. 2015, 6, 714. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Guo, S.R. 24-epibrassinolide improves cucumber photosynthesis under hypoxia by increasing CO2 assimilation and photosystem II efficiency. Photosynthetica 2014, 52, 96–104. [Google Scholar] [CrossRef]
- Trewavas, A.J.; Malho, R. Ca2+ signalling in plant cells: The big network. Curr. Opin. Plant Biol. 1998, 1, 428–433. [Google Scholar] [CrossRef]
- Lai, W.-L.; Zhang, Y.; Chen, Z.-H. Radial oxygen loss, photosynthesis, and nutrient removal of 35 wetland plants. Ecol. Eng. 2012, 39, 24–30. [Google Scholar] [CrossRef]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Shen, Q.F.; Zhang, G.P. The mechanisms for the difference in waterlogging tolerance among sea barley, wheat and barley. Plant Growth Regul. 2022, 96, 431–441. [Google Scholar] [CrossRef]
- Liang, K.; Tang, K.; Fang, T.; Qiu, F. Waterlogging tolerance in maize: Genetic and molecular basis. Mol. Breed. 2020, 40, 111. [Google Scholar] [CrossRef]
- Matsukura, C.; Kawai, M.; Toyofuku, K.; Barrero, R.A.; Yamaguchi, J. Transverse vein differentiation associated with gas space formation—Fate of the middle cell layer in leaf sheath development of rice. Ann. Bot. 2000, 85, 19–27. [Google Scholar] [CrossRef]
- Shimamura, S.; Mochizuki, T.; Nada, Y.; Fukuyama, M. Formation and function of secondary aerenchyma in hypocotyl, roots and nodules of soybean (Glycine max) under flooded conditions. Plant Soil 2003, 251, 351–359. [Google Scholar] [CrossRef]
- Sundgren, T.; Uhlen, A.; Waalen, W.; Lillemo, M. Field Screening of Waterlogging Tolerance in Spring Wheat and Spring Barley. Agronomy 2018, 8, 38. [Google Scholar] [CrossRef]
- Seago, J.L., Jr.; Marsh, L.C.; Stevens, K.J.; Soukup, A.; Votrubova, O.; Enstone, D.E. A re-examination of the root cortex in wetland flowering plants with respect to aerenchyma. Ann. Bot. 2005, 96, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, D.; Lin, Z.; Guan, B.; Liu, D.; Yang, L.; Deng, X.; Mei, F.; Zhou, Z. Histone acetylation modification affects cell wall degradation and aerenchyma formation in wheat seminal roots under waterlogging. Plant Growth Regul. 2019, 87, 149–163. [Google Scholar] [CrossRef]
- Sakagami, J.; Joho, Y.; Ito, O. Contrasting physiological responses by cultivars of Oryza sativa and O. glaberrima to prolonged submergence. Ann. Bot. 2009, 103, 171–180. [Google Scholar] [CrossRef]
- Matthews, G. Rice in Deep Water. Crop Prot. 1994, 13, 316–317. [Google Scholar] [CrossRef]
- Colmer, T.D. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.). Ann. Bot. 2003, 91, 301–309. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Sun, S.; Zhao, J.; Huang, Z.; Peng, L.; Huang, C.; Tang, Z.; Huang, Q.; Wang, Z. UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination. Nat. Commun. 2023, 14, 2296. [Google Scholar] [CrossRef] [PubMed]
- Pantin, F.; Simonneau, T.; Muller, B. Coming of leaf age: Control of growth by hydraulics and metabolics during leaf ontogeny. New Phytol. 2012, 196, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Visser, E.J.; De Kroon, H.; Pierik, R.; Voesenek, L.A.; Huber, H. Fitness consequences of natural variation in flooding-induced shoot elongation in Rumex palustris. New Phytol. 2011, 190, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Van Aken, J.M.; Voesenek, L.A.C.J. Is elongation-induced leaf emergence beneficial for submerged Rumex species? Ann. Bot. 2009, 103, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Colmer, T.D. Plant tolerance of flooding stress—Recent advances. Plant Cell Environ. 2014, 37, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, H.; Akman, M.; Jamar, D.C.; Vreugdenhil, D.; Kooiker, M.; Van Tienderen, P.; Voesenek, L.A.; Schranz, M.E.; Sasidharan, R. Group VII ethylene response factor diversification and regulation in four species from flood-prone environments. Plant Cell Environ. 2014, 37, 2421–2432. [Google Scholar] [CrossRef]
- Tougou, M.; Hashiguchi, A.; Yukawa, K.; Nanjo, Y.; Hiraga, S.; Nakamura, T.; Nishizawa, K.; Komatsu, S. Responses to flooding stress in soybean seedlings with the alcohol dehydrogenase transgene. Plant Biotechnol. 2012, 29, 301–305. [Google Scholar] [CrossRef]
- Xuewen, X.; Huihui, W.; Xiaohua, Q.; Qiang, X.; Xuehao, C. Waterlogging-induced increase in fermentation and related gene expression in the root of cucumber (Cucumis sativus L.). Sci. Hortic. 2014, 179, 388–395. [Google Scholar] [CrossRef]
- Pan, D.L.; Wang, G.; Wang, T.; Jia, Z.H.; Guo, Z.R.; Zhang, J.Y. AdRAP2.3, a novel ethylene response factor VII from Actinidia deliciosa, enhances waterlogging resistance in transgenic tobacco through improving expression levels of PDC and ADH genes. Int. J. Mol. Sci. 2019, 20, 1189. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.J.; Harrison-Murray, R.S.; Taylor, J.M. Rapid flood-induced stomatal closure accompanies xylem sap transportation of root-derived acetaldehyde and ethanol in Forsythia. Environ. Exp. Bot. 2008, 64, 196–205. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Meng, Y.; Chandrasekaran, U.; Luo, X.; Yang, W.; Shu, K. Plant waterlogging/flooding stress responses: From seed germination to maturation. Plant Physiol. Biochem. 2020, 148, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.F.; Zheng, H.J.; Liu, Z.D. Effects of waterlogging at different stages on growth and ear quality of waxy maize. Agric. Water Manag. 2022, 266, 107603. [Google Scholar] [CrossRef]
- Chang, W.W.; Huang, L.; Shen, M.; Webster, C.; Burlingame, A.L.; Roberts, J.K. Patterns of protein synthesis and tolerance of anoxia in root tips of maize seedlings acclimated to a low-oxygen environment, and identification of proteins by mass spectrometry. Plant Physiol. 2000, 122, 295–317. [Google Scholar] [CrossRef]
- Zhou, Y.; Tan, W.J.; Xie, L.J.; Qi, H.; Yang, Y.C.; Huang, L.P.; Lai, Y.X.; Tan, Y.F.; Zhou, D.M.; Yu, L.J.; et al. Polyunsaturated linolenoyl-CoA modulates ERF-VII-mediated hypoxia signaling in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 330–348. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Liao, K.; Wang, L.N.; Shi, L.L.; Zhang, Y.; Xu, L.J.; Zhou, Y.; Li, J.F.; Chen, Y.Q.; Chen, Q.F.; et al. Calcium-dependent activation of CPK12 facilitates its cytoplasm-to-nucleus translocation to potentiate plant hypoxia sensing by phosphorylating ERF-VII transcription factors. Mol. Plant 2023, 16, 979–998. [Google Scholar] [CrossRef] [PubMed]
- Whan, A.; Dielen, A.-S.; Mieog, J.; Bowerman, A.F.; Robinson, H.M.; Byrne, K.; Colgrave, M.; Larkin, P.J.; Howitt, C.A.; Morell, M.K.; et al. Engineering alpha-amylase levels in wheat grain suggests a highly sophisticated level of carbohydrate regulation during development. J. Exp. Bot. 2014, 65, 5443–5457. [Google Scholar] [CrossRef]
- Park, M.; Yim, H.K.; Park, H.G.; Lim, J.; Kim, S.H.; Hwang, Y.S. Interference with oxidative phosphorylation enhances anoxic expression of rice alpha-amylase genes through abolishing sugar regulation. J. Exp. Bot. 2010, 61, 3235–3244. [Google Scholar] [CrossRef]
- Lee, K.W.; Chen, P.W.; Lu, C.A.; Chen, S.; Ho, T.H.D.; Yu, S.M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Plant Biol. 2009, 2, 61. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Pelayo, M.A.; Trijatmiko, K.R.; Gabunada, L.F.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, D.; Suh, M.C. Cuticle ultrastructure, cuticular lipid composition, and gene expression in hypoxia-stressed Arabidopsis stems and leaves. Plant Cell Rep. 2017, 36, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Tan, W.J.; Yang, Y.C.; Tan, Y.F.; Zhou, Y.; Zhou, D.M.; Xiao, S.; Chen, Q.F. Long-Chain acyl-CoA Synthetase LACS2 Contributes to Submergence Tolerance by Modulating Cuticle Permeability in Arabidopsis. Plants 2020, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Shabala, L.; Zhang, X.; Zhou, M.; Voesenek, L.A.C.J.; Hartman, S.; Yu, M.; Shabala, S. Cation transporters in cell fate determination and plant adaptive responses to a low-oxygen environment. J. Exp. Bot. 2022, 73, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Phukan, U.J.; Mishra, S.; Shukla, R.K. Waterlogging and submergence stress: Affects and acclimation. Crit. Rev. Biotechnol. 2016, 36, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Qiao, X.; Tian, Z.; Zhang, X.; Zou, X.; Cheng, Y.; Lu, G.; Zeng, L.; Fu, G.; Ding, X.; et al. Proteomic analysis of rapeseed root response to waterlogging stress. Plants 2018, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Barickman, T.C.; Simpson, C.R.; Sams, C.E. Waterlogging causes early modification in the physiological performance, carotenoids, chlorophylls, proline, and soluble sugars of cucumber plants. Plants 2019, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, A.R. Proline, a multifaceted signalling molecule in plant responses to abiotic stress: Understanding the physiological mechanisms. Plant Biol. 2021, 24, 227–239. [Google Scholar] [CrossRef]
- Yang, H.; Huang, T.; Ding, M.; Lu, D.; Lu, W. Effects of waterlogging around flowering stage on the grain yield and eating properties of fresh waxy maize. Cereal Chem. 2016, 93, 605–611. [Google Scholar] [CrossRef]
- Habibullah, M.; Sarkar, S.; Islam, M.M.; Ahmed, K.U.; Rahman, M.Z.; Awad, M.F.; Elsayed, A.I.; Mansour, E.; Hossain, M.S. Assessing the response of diverse sesame genotypes to waterlogging durations at different plant growth stages. Plants 2021, 10, 2294. [Google Scholar] [CrossRef]
- Fan, W.; Liao, X.L.; Tan, Y.Q.; Wang, X.R.; Schroeder, J.; Li, Z.X. Arabidopsis PLANT U-BOX44 down-regulates osmotic stress signaling by mediating Ca2+-DEPENDENT PROTEIN KINASE4 degradation. Plant Cell 2023, 35, 3870–3888. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Li, X.L.; Xie, X.; Liu, L.J.; Fu, J.Y.; Wang, Q. Maize transcription factor ZmNAC2 enhances osmotic stress tolerance in transgenic Arabidopsis. J. Plant Physiol. 2023, 282, 153948. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.A.D.; Oliveira, L.E.M.D.; Domiciano, D.; Carvalho, J.N.D.; Prudente, D.D.O.; Guimarães, R.J. Effect of nitrogen source and oxygen deficiency on carbon metabolism and antioxidant system of rubber tree plants (Hevea spp.). Aust. J. Crop Sci. 2018, 12, 116–125. [Google Scholar] [CrossRef]
- Basu, S.; Kumar, G.; Kumari, N.; Kumari, S.; Shekhar, S.; Kumar, S.; Rajwanshi, R. Reactive oxygen species and reactive nitrogen species induce lysigenous aerenchyma formation through programmed cell death in rice roots under submergence. Environ. Exp. Bot. 2020, 177, 104118. [Google Scholar] [CrossRef]
- Dantas, B.; Silva, R.; Ribeiro, R.; Aragão, C. Respiration and antioxidant enzymes activity in watermelon seeds and seedlings subjected to salt and temperature stresses. Am. J. Exp. Agric. 2015, 7, 70–77. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Ahmed, A.R.; Farris, F.F.; Ray, S.D. Lipid peroxidation. Biomed. Sci. 2023. [Google Scholar]
- Hodges, D.M.; Andrews, C.J.; Johnson, D.A.; Hamilton, R.I. Antioxidant enzyme responses to chilling stress in differentially sensitive inbred maize lines. J. Exp. Bot. 1997, 48, 1105–1113. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Tanyolaç, D.; Ayhan, B. Effects of cadmium on antioxidant enzyme and photosynthetic activities in leaves of two maize cultivars. J. Plant Physiol. 2008, 165, 600–611. [Google Scholar] [CrossRef]
- Abedi, T.; Pakniyat, H. Antioxidant enzymes changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Genet. Plant Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Li, M.-F.; Zhu, J.-Q.; Jiang, Z.-H. Plant growth regulators and nutrition applied to cotton after waterlogging. In Proceedings of the Third International Conference on Intelligent System Design and Engineering Applications, Hong Kong, China, 16–18 January 2013; pp. 1045–1048. [Google Scholar] [CrossRef]
- Das, K.K.; Panda, D.; Sarkar, R.K.; Reddy, J.N.; Ismail, A.M. Submergence tolerance in relation to variable floodwater conditions in rice. Environ. Exp. Bot. 2009, 66, 425–434. [Google Scholar] [CrossRef]
- Rich, S.M.; Ludwig, M.; Colmer, T.D. Photosynthesis in aquatic adventitious roots of the halophytic stem-succulent Tecticornia pergranulata (formerly Halosarcia pergranulata). Plant Cell Environ. 2008, 31, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Fu, S.; Chen, S.; Zhang, W.; Qi, C. Ethylene response factor BnERF2-like (ERF2.4) from Brassica napus L. enhances submergence tolerance and alleviates oxidative damage caused by submergence in Arabidopsis thaliana. Crop J. 2016, 4, 199–211. [Google Scholar] [CrossRef]
- Hill, R.D.; De Castro, J.; Mira, M.M.; Igamberdiev, A.U.; Hebelstrup, K.H.; Renault, S.; Xu, W.; Badea, A.; Stasolla, C. Over-expression of the barley Phytoglobin 1 (HvPgb1) evokes leaf-specific transcriptional responses during root waterlogging. J. Plant Physiol. 2023, 283, 153944. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gao, J.; Zhang, Z.; Liu, W.; Cheng, P.; Mu, W.; Su, T.; Chen, S.; Chen, F.; Jiang, J. Overexpression of CmSOS1 confers waterlogging tolerance in Chrysanthemum. J. Integr. Plant Biol. 2020, 62, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- An, Y.Y.; Lin, Q.; Wang, L.J. ALA pretreatment improves waterlogging tolerance of fig plants. PLoS ONE 2016, 11, e0147202. [Google Scholar] [CrossRef] [PubMed]
- Keya, S.S.; Mostofa, M.G.; Mezanur Rahman, M.; Das, A.K.; Abiar Rahman, M.; Anik, T.R.; Sultana, S.; Arifur Rahman Khan, M.; Robyul Islam, M.; Watanabe, Y.; et al. Effects of glutathione on waterlogging-induced damage in sesame crop. Ind. Crops Prod. 2022, 185, 115092. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotechnol. 2017, 93, 385–391. [Google Scholar] [CrossRef]
- Salah, A.; Zhan, M.; Cao, C.; Han, Y.; Ling, L.; Liu, Z.; Li, P.; Ye, M.; Jiang, Y. γ-Aminobutyric acid promotes chloroplast ultrastructure, antioxidant capacity, and growth of waterlogged maize seedlings. Sci. Rep. 2019, 9, 484. [Google Scholar] [CrossRef]
- Chang, R.; Jang, C.J.; Branco-Price, C.; Nghiem, P.; Bailey-Serres, J. Transient MPK6 activation in response to oxygen deprivation and reoxygenation is mediated by mitochondria and aids seedling survival in Arabidopsis. Plant Mol. Biol. 2012, 78, 109–122. [Google Scholar] [CrossRef]
- Wan, D. The Study of Calcium Signal Transduction Induced by Oxidative Stress (H2O2). Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2018. [Google Scholar]
- Bansal, R.; Srivastava, J.P. Effect of waterlogging on photosynthetic and biochemical parameters in pigeonpea. Russ. J. Plant Physiol. 2015, 62, 322–327. [Google Scholar] [CrossRef]
- Gravarr, D.A.; Kirby, C.J. Patterns of photosynthesis and starch allocation in seedlings of four bottomland hardwood tree species subjected to flooding. Tree Physiol. 1998, 18, 411–417. [Google Scholar] [CrossRef]
- Sairam, R.K.; Dharmar, K.; Chinnusamy, V.; Meena, R.C. Waterlogging-induced increase in sugar mobilization, fermentation, and related gene expression in the roots of mung bean (Vigna radiata). J. Plant Physiol. 2009, 166, 602–616. [Google Scholar] [CrossRef] [PubMed]
- Distefano, G.; Cirilli, M.; Las Casas, G.; La Malfa, S.; Continella, A.; Rugini, E.; Thomas, B.; Long, G.; Gentile, A.; Muleo, R. Ectopic expression of Arabidopsis phytochrome B in Troyer citrange affects photosynthesis and plant morphology. Sci. Hortic. 2013, 159, 1–7. [Google Scholar] [CrossRef]
- Lim, J.; Park, J.H.; Jung, S.; Hwang, D.; Nam, H.G.; Hong, S. Antagonistic roles of PhyA and PhyB in far-red light-dependent Leaf senescence in Arabidopsis thaliana. Plant Cell Physiol. 2018, 59, 1753–1764. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Wang, X.; Wang, J.; Fan, K.; Li, Z.; Lin, W. DELLA proteins negatively regulate dark-induced senescence and chlorophyll degradation in Arabidopsis through interaction with the transcription factor WRKY6. Plant Cell Rep. 2018, 37, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Panda, D.; Dash, G.K.; Mohanty, S.; Sekhar, S.; Roy, A.; Tudu, C.; Behera, L.; Tripathy, B.C.; Baig, M.J. Phytochrome A mediated modulation of photosynthesis, development and yield in rice (Oryza sativa L.) in fluctuating light environment. Environ. Exp. Bot. 2023, 206, 105183. [Google Scholar] [CrossRef]
- Jackson, M.B. Long-distance signalling from roots to shoots assessed the flooding story. J. Exp. Bot. 2002, 53, 175–181. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on leaf mesophyll cell ultrastructure and photosynthetic characteristics of summer maize. PLoS ONE 2016, 11, e0161424. [Google Scholar] [CrossRef]
- Liao, C.T.; Ho, L.C. Effect of flooding stress on photosynthetic activities of momordica charantia. Plant Physiol. Biochem. 1994, 32, 479–485. [Google Scholar]
- Hu, J.; Ren, B.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Poor development of spike differentiation triggered by lower photosynthesis and carbon partitioning reduces summer maize yield after waterlogging. Crop J. 2022, 10, 478–489. [Google Scholar] [CrossRef]
- Huang, C.; Gao, Y.; Qin, A.; Liu, Z.; Zhao, B.; Ning, D.; Ma, S.; Duan, A.; Liu, Z. Effects of waterlogging at different stages and durations on maize growth and grain yields. Agric. Water Manag. 2022, 261, 107334. [Google Scholar] [CrossRef]
- Tian, L.; Li, J.; Bi, W.; Zuo, S.; Li, L.; Li, W.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) Under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Wakchaure, G.C.; Minhas, P.S.; Kumar, S.; Khapte, P.S.; Rane, J.; Reddy, K.S. Bulb productivity and quality of monsoon onion (Allium cepa L.) as affected by transient waterlogging at different growth stages and its alleviation with plant growth regulators. Agric. Water Manag. 2023, 278, 108136. [Google Scholar] [CrossRef]
- Kurokawa, Y.; Nagai, K.; Huan, P.D.; Shimazaki, K.; Qu, H.; Mori, Y.; Toda, Y.; Kuroha, T.; Hayashi, N.; Aiga, S.; et al. Rice leaf hydrophobicity and gas films are conferred by a wax synthesis gene (LGF1) and contribute to flood tolerance. New Phytol. 2018, 218, 1558–1569. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.N.; Mostofa, M.G.; Rahman, M.M.; Tahjib-Ul-Arif, M.; Das, A.K.; Mohi-Ud-Din, M.; Rohman, M.M.; Hafiz, H.R.; Ansary, M.M.U.; Tran, L.P. Glutathione improves rice tolerance to submergence: Insights into its physiological and biochemical mechanisms. J. Biotechnol. 2021, 325, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Perata, P. Ethylene signaling controls fast oxygen sensing in plants. Trends Plant Sci. 2020, 25, 3–6. [Google Scholar] [CrossRef]
- Negi, S.; Sukumar, P.; Liu, X.; Cohen, J.D.; Muday, G.K. Genetic dissection of the role of ethylene in regulating auxin-dependent lateral and adventitious root formation in tomato. Plant J. 2010, 61, 3–15. [Google Scholar] [CrossRef]
- Liu, Z.; Hartman, S.; Van Veen, H.; Zhang, H.; Leeggangers, H.; Martopawiro, S.; Bosman, F.; De Deugd, F.; Su, P.; Hummel, M.; et al. Ethylene augments root hypoxia tolerance via growth cessation and reactive oxygen species amelioration. Plant Physiol. 2022, 190, 1365–1383. [Google Scholar] [CrossRef]
- Shin, S.Y.; Choi, Y.; Kim, S.G.; Park, S.J.; Park, J.S.; Moon, K.B.; Kim, H.S.; Jeon, J.H.; Cho, H.S.; Lee, H.J. Submergence promotes auxin-induced callus formation through ethylene-mediated post-transcriptional control of auxin receptors. Mol. Plant 2022, 15, 1947–1961. [Google Scholar] [CrossRef]
- White, M.D.; Kamps, J.; East, S.; Taylor Kearney, L.J.; Flashman, E. The plant cysteine oxidases from Arabidopsis thaliana are kinetically tailored to act as oxygen sensors. J. Biol. Chem. 2018, 293, 11786–11795. [Google Scholar] [CrossRef] [PubMed]
- Weits, D.A.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Hubberten, H.M.; Riegler, H.; Hoefgen, R.; Perata, P.; Van Dongen, J.T.; Licausi, F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nat. Commun. 2014, 5, 3425. [Google Scholar] [CrossRef] [PubMed]
- Hartman, S.; Liu, Z.; Van Veen, H.; Vicente, J.; Reinen, E.; Martopawiro, S.; Zhang, H.; Van Dongen, N.; Bosman, F.; Bassel, G.W.; et al. Ethylene-mediated nitric oxide depletion pre-adapts plants to hypoxia stress. Nat. Commun. 2019, 10, 4020. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Tedds, H.M.; Labandera, A.M.; Bailey, M.; White, M.D.; Hartman, S.; Sprigg, C.; Mogg, S.L.; Osborne, R.; Dambire, C.; et al. Oxygen-dependent proteolysis regulates the stability of angiosperm polycomb repressive complex 2 subunit VERNALIZATION 2. Nat. Commun. 2018, 9, 5438. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marin-De La Rosa, N.; Vicente Conde, J.; Sousa Correia, C.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: An ethylene response transcription factor that is important for hypoxia survival. Plant Physiol. 2010, 153, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.P.; Lin, T.Y.; Wang, N.N.; Shih, M.C. Differential expression of genes encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis during hypoxia. Plant Mol. Biol. 2005, 58, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef]
- Yin, D.; Sun, D.; Han, Z.; Ni, D.; Jiang, C.Z. PhERF2, an ethylene-responsive element binding factor, plays an essential role in waterlogging tolerance of petunia. Hortic. Res. 2019, 6, 83. [Google Scholar] [CrossRef]
- Luan, H.; Guo, B.; Shen, H.; Pan, Y.; Hong, Y.; Lv, C.; Xu, R. Overexpression of barley transcription factor HvERF2.11 in Arabidopsis enhances plant waterlogging tolerance. Int. J. Mol. Sci. 2020, 21, 1982. [Google Scholar] [CrossRef]
- Sun, D.; Nandety, R.S.; Zhang, Y.; Reid, M.S.; Niu, L.; Jiang, C.Z. A petunia ethylene-responsive element binding factor, PhERF2, plays an important role in antiviral RNA silencing. J. Exp. Bot. 2016, 67, 3353–3365. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell. 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Fukao, T.; Bailey-Serres, J. Ethylene—A key regulator of submergence responses in rice. Plant Sci. 2008, 175, 43–51. [Google Scholar] [CrossRef]
- Dawood, T.; Yang, X.; Visser, E.J.; Te Beek, T.A.; Kensche, P.R.; Cristescu, S.M.; Lee, S.; Flokova, K.; Nguyen, D.; Mariani, C.; et al. A Co-Opted hormonal cascade activates dormant adventitious root primordia upon flooding in solanum dulcamara. Plant Physiol. 2016, 170, 2351–2364. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Nagai, K.; Gamuyao, R.; Wang, D.R.; Furuta, T.; Nakamori, M.; Kitaoka, T.; Adachi, K.; Minami, A.; Mori, Y.; et al. Ethylene-gibberellin signaling underlies adaptation of rice to periodic flooding. Science 2018, 361, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Huang, S.N.; Wang, G.; Xuan, J.P.; Guo, Z.R. Overexpression of Actinidia deliciosa pyruvate decarboxylase 1 gene enhances waterlogging stress in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2016, 106, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Vidoz, M.L.; Loreti, E.; Mensuali, A.; Alpi, A.; Perata, P. Hormonal interplay during adventitious root formation in flooded tomato plants. Plant J. 2010, 63, 551–562. [Google Scholar] [CrossRef]
- Wang, X.; Huang, M.; Zhou, Q.; Cai, J.; Dai, T.; Cao, W.; Jiang, D. Physiological and proteomic mechanisms of waterlogging priming improves tolerance to waterlogging stress in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2016, 132, 175–182. [Google Scholar] [CrossRef]
- Jiang, Z.-H.; Zhua, J.-Q.; Yang, W.; Lia, M.-F.; Yua, Y. Effects of remedial measures implemented after waterlogging on cotton. In Proceedings of the Third International Conference on Intelligent System Design and Engineering Applications, Hong Kong, China, 16–18 January 2013; pp. 692–695. [Google Scholar] [CrossRef]
- Li, Y.; Sun, D.; Xu, K.; Jin, L.; Peng, R. Hydrogen sulfide enhances plant tolerance to waterlogging stress. Plants 2021, 10, 1928. [Google Scholar] [CrossRef]
- Zhong, Y.-H.; Guo, Z.-J.; Wei, M.-Y.; Wang, J.-C.; Song, S.-W.; Chi, B.-J.; Zhang, Y.-C.; Liu, J.-W.; Li, J.; Zhu, X.-Y.; et al. Hydrogen sulfide upregulates the alternative respiratory pathway in mangrove plant Avicennia marina to attenuate waterlogging-induced oxidative stress and mitochondrial damage in a calcium-dependent manner. Plant Cell Environ. 2023, 46, 1521–1539. [Google Scholar] [CrossRef]
- Hasnain, M.; Munir, N.; Abideen, Z.; Zulfiqar, F.; Koyro, H.W.; El-Naggar, A.; Caçador, I.; Duarte, B.; Rinklebe, J.; Yong, J.W.H. Biochar-plant interaction and detoxification strategies under abiotic stresses for achieving agricultural resilience: A critical review. Ecotoxicol. Environ. Saf. 2023, 249, 114408. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Yang, Y.; Zheng, C.; Luo, X.; Chandrasekaran, U.; Yin, H.; Chen, F.; Meng, Y.; Chen, L.; Shu, K. Flooding represses soybean seed germination by mediating anaerobic respiration, glycometabolism and phytohormones biosynthesis. Environ. Exp. Bot. 2021, 188, 104491. [Google Scholar] [CrossRef]
- Kaur, G.; Vikal, Y.; Kaur, L.; Kalia, A.; Mittal, A.; Kaur, D.; Yadav, I. Elucidating the morpho-physiological adaptations and molecular responses under long-term waterlogging stress in maize through gene expression analysis. Plant Sci. 2021, 304, 110823. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Meng, X.; Wang, M.; Zang, S.; Feng, L. Improvement in submergence tolerance of cherry through regulation of carbohydrate metabolism and pant growth by PsERF and PsCIPK. Appl. Biochem. Biotechnol. 2018, 184, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Phukan, U.J.; Jeena, G.S.; Tripathi, V.; Shukla, R.K. MaRAP2-4, a waterlogging-responsive ERF from Mentha, regulates bidirectional sugar transporter AtSWEET10 to modulate stress response in Arabidopsis. Plant Biotechnol. J. 2018, 16, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Yu, L.J.; Chen, Q.F.; Wang, F.Z.; Huang, L.; Xia, F.N.; Zhu, T.R.; Wu, J.X.; Yin, J.; Liao, B.; et al. Arabidopsis acyl-CoA-binding protein ACBP3 participates in plant response to hypoxia by modulating very-long-chain fatty acid metabolism. Plant J. 2015, 81, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Zhou, Y.; Chen, Q.F.; Xiao, S. New insights into the role of lipids in plant hypoxia responses. Prog. Lipid Res. 2021, 81, 101072. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, D.M.; Yu, W.W.; Shi, L.L.; Zhang, Y.; Lai, Y.X.; Huang, L.P.; Qi, H.; Chen, Q.F.; Yao, N.; et al. Phosphatidic acid modulates MPK3- and MPK6-mediated hypoxia signaling in Arabidopsis. Plant Cell 2022, 34, 889–909. [Google Scholar] [CrossRef]
- Bloom, A.J.; Sukrapanna, S.S.; Warner, R.L. Root respiration associated with ammonium and nitrate absorption and assimilation by barley. Plant Physiol. 1992, 99, 1294–1301. [Google Scholar] [CrossRef]
- Loque, D.; Ludewig, U.; Yuan, L.; Von Wiren, N. Tonoplast intrinsic proteins AtTIP2;1 and AtTIP2;3 facilitate NH3 transport into the vacuole. Plant Physiol. 2005, 137, 671–680. [Google Scholar] [CrossRef]
- Gupta, K.J.; Mur, L.A.J.; Wany, A.; Kumari, A.; Fernie, A.R.; Ratcliffe, R.G. The role of nitrite and nitric oxide under low oxygen conditions in plants. New Phytol. 2020, 225, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Borella, J.; Becker, R.; Lima, M.C.; Oliveira, D.D.S.C.D.; Braga, E.J.B.; Oliveira, A.C.B.D.; Amarante, L.D. Nitrogen source influences the antioxidative system of soybean plants under hypoxia and re-oxygenation. Sci. Agric. 2019, 76, 51–62. [Google Scholar] [CrossRef]
- Da-Silva, C.J.; Do Amarante, L. Short-term nitrate supply decreases fermentation and oxidative stress caused by waterlogging in soybean plants. Environ. Exp. Bot. 2020, 176, 104078. [Google Scholar] [CrossRef]
- Pucciariello, C.; Perata, P. New insights into reactive oxygen species and nitric oxide signalling under low oxygen in plants. Plant Cell Environ. 2017, 40, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Prakash, V.; Vishwakarma, K.; Singh, V.P.; Rai, P.; Ramawat, N.; Tripathi, D.K.; Sharma, S. NO and ROS implications in the organization of root system architecture. Physiol. Plant 2020, 168, 473–489. [Google Scholar] [CrossRef] [PubMed]
- Wany, A.; Kumari, A.; Gupta, K.J. Nitric oxide is essential for the development of aerenchyma in wheat roots under hypoxic stress. Plant Cell Environ. 2017, 40, 3002–3017. [Google Scholar] [CrossRef] [PubMed]
- Zottini, M.; Formentin, E.; Scattolin, M.; Carimi, F.; Lo Schiavo, F.; Terzi, M. Nitric oxide affects plant mitochondrial functionality in vivo. FEBS Lett. 2002, 515, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Liu, G.; Xu, S.; Dai, J.; Li, W.; Li, Z.; Zhang, D.; Li, C.; Dong, H. Nitric oxide increases the biomass and lint yield of field-grown cotton under temporary waterlogging through physiological and molecular regulation. Field Crops Res. 2021, 261, 107989. [Google Scholar] [CrossRef]
- Liu, C.; Lan, C.; Li, C.; Li, C.; Huang, J. Exogenous spermidine and calcium alleviate waterlogging stress in cherry tomato at the seedling stage. Sci. Hortic. 2023, 307, 111504. [Google Scholar] [CrossRef]
- Zhu, J.; Li, A.; Zhang, J.; Sun, C.; Tang, G.; Chen, L.; Hu, J.; Zhou, N.; Wang, S.; Zhou, Y.; et al. Effects of nitrogen application after abrupt drought-flood alternation on rice root nitrogen uptake and rhizosphere soil microbial diversity. Environ. Exp. Bot. 2022, 201, 105007. [Google Scholar] [CrossRef]
- Martínez-Arias, C.; Witzell, J.; Solla, A.; Martin, J.A.; Rodríguez-Calcerrada, J. Beneficial and pathogenic plant-microbe interactions during flooding stress. Plant Cell Environ. 2022, 45, 2875–2897. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jiang, Z.; Shi, M.; Zhou, Y.; Huo, L.; Li, X.; Xu, K. Comparative transcriptome provides insight into responding mechanism of waterlogging stress in Actinidia valvata Dunn. Gene 2022, 845, 146843. [Google Scholar] [CrossRef] [PubMed]
- Hess, N.; Klode, M.; Anders, M.; Sauter, M. The hypoxia responsive transcription factor genes ERF71/HRE2 and ERF73/HRE1 of Arabidopsis are differentially regulated by ethylene. Physiol. Plant 2011, 143, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.-F.; Li, Z.; Hu, C.-G.; Zhang, Y.-J.; Muhammad, A.; Zhong, Y.-P.; Fang, J.-B. Transcriptome-wide identification and expression analysis of ERF family genes in Actinidia valvata during waterlogging stress. Sci. Hortic. 2021, 281, 109994. [Google Scholar] [CrossRef]
- Mao, J.L.; Miao, Z.Q.; Wang, Z.; Yu, L.H.; Cai, X.T.; Xiang, C.B. Arabidopsis ERF1 mediates cross-talk between ethylene and auxin biosynthesis during primary root elongation by regulating ASA1 expression. PLoS Genet. 2016, 12, e1005760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhang, J.; Chen, S.; Zhang, Z.; Wan, G.; Mao, J.; Wang, Z.; Tan, S.; Xiang, C. ERF1 inhibits lateral root emergence by promoting local auxin accumulation and repressing ARF7 expression. Cell Rep. 2023, 42, 112565. [Google Scholar] [CrossRef]
- Wei, X.; Xu, H.; Rong, W.; Ye, X.; Zhang, Z. Constitutive expression of a stabilized transcription factor group VII ethylene response factor enhances waterlogging tolerance in wheat without penalizing grain yield. Plant Cell Environ. 2019, 42, 1471–1485. [Google Scholar] [CrossRef]
- Liu, B.; Jiang, Y.; Tang, H.; Tong, S.; Lou, S.; Shao, C.; Zhang, J.; Song, Y.; Chen, N.; Bi, H.; et al. The ubiquitin E3 ligase SR1 modulates the submergence response by degrading phosphorylated WRKY33 in Arabidopsis. Plant Cell 2021, 33, 1771–1789. [Google Scholar] [CrossRef]
- Meng, D.; Li, Y.; Bai, Y.; Li, M.; Cheng, L. Genome-wide identification and characterization of WRKY transcriptional factor family in apple and analysis of their responses to waterlogging and drought stress. Plant Physiol. Biochem. 2016, 103, 71–83. [Google Scholar] [CrossRef]
- Xie, L.-J.; Wang, J.-H.; Liu, H.-S.; Yuan, L.-B.; Tan, Y.-F.; Tan, W.-J.; Zhou, Y.; Chen, Q.-F.; Qi, H.; Li, J.-F.; et al. MYB30 integrates light signals with antioxidant biosynthesis to regulate plant responses during postsubmergence recovery. New Phytol. 2023, 237, 2238–2254. [Google Scholar] [CrossRef] [PubMed]
- Rauf, M.; Arif, M.; Fisahn, J.; Xue, G.P.; Balazadeh, S.; Mueller-Roeber, B. NAC transcription factor speedy hyponastic growth regulates flooding-induced leaf movement in Arabidopsis. Plant Cell 2013, 25, 4941–4955. [Google Scholar] [CrossRef]
- Berry, H.M.; Argueso, C.T. More than growth: Phytohormone-regulated transcription factors controlling plant immunity, plant development and plant architecture. Curr. Opin. Plant Biol. 2022, 70, 102309. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Dong, X. In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid-mediated gene activation in Arabidopsis. Plant Cell 2002, 14, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Di, D.W.; Li, G.; Li, Y.; Kronzucker, H.J.; Shi, W. Transcriptome analysis of rice (Oryza sativa L.) in response to ammonium resupply reveals the involvement of phytohormone signaling and the transcription factor OsJAZ9 in reprogramming of nitrogen uptake and metabolism. J. Plant Physiol. 2020, 246–247, 153137. [Google Scholar] [CrossRef]
- Yuan, L.B.; Dai, Y.S.; Xie, L.J.; Yu, L.J.; Zhou, Y.; Lai, Y.X.; Yang, Y.C.; Xu, L.; Chen, Q.F.; Xiao, S. Jasmonate regulates plant responses to postsubmergence reoxygenation through transcriptional activation of antioxidant synthesis. Plant Physiol. 2017, 173, 1864–1880. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, F.-L.; Jiang, D. Priming: A promising strategy for crop production in response to future climate. J. Integr. Agric. 2017, 16, 2709–2716. [Google Scholar] [CrossRef]
- Kartashov, A.V.; Zlobin, I.E.; Pashkovskiy, P.P.; Pojidaeva, E.S.; Ivanov, Y.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Kuznetsov, V.V. Effects of drought stress memory on the accumulation of stress-protective compounds in naturally grown pine and spruce. Plant Physiol. Biochem. 2023, 200, 107761. [Google Scholar] [CrossRef]
- Luis De La Fuente, J.; Zunzunegui, M.; Barradas, M.C.D. Physiological responses to water stress and stress memory in Argania spinosa. Plant Stress 2023, 7, 100133. [Google Scholar] [CrossRef]
- Sharma, M.; Kumar, P.; Verma, V.; Sharma, R.; Bhargava, B.; Irfan, M. Understanding plant stress memory response for abiotic stress resilience: Molecular insights and prospects. Plant Physiol. Biochem. 2022, 179, 10–24. [Google Scholar] [CrossRef]
- Kinoshita, T.; Seki, M. Epigenetic memory for stress response and adaptation in plant. Plant Cell Physiol. 2014, 55, 1859–1863. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2022, 27, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Gallusci, P.; Agius, D.R.; Moschou, P.N.; Dobranszki, J.; Kaiserli, E.; Martinelli, F. Deep inside the epigenetic memories of stressed plants. Trends Plant Sci. 2023, 28, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiang, D.; Wollenweber, B.; Li, Y.; Dai, T.; Cao, W. Waterlogging pretreatment during vegetative growth improves tolerance to waterlogging after anthesis in wheat. Plant Sci. 2011, 180, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Sharma, K.D.; Siddique, K.H.M.; Nayyar, H. Cold priming the chickpea seeds imparts reproductive cold tolerance by reprogramming the turnover of carbohydrates, osmo-protectants and redox components in leaves. Sci. Hortic. 2020, 261, 108929. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; An, J.; Yin, M.; Jia, X.; Guan, Y.; He, F.; Hu, J. Cold plasma treatment and exogenous salicylic acid priming enhances salinity tolerance of Oryza sativa seedlings. Protoplasma 2019, 256, 79–99. [Google Scholar] [CrossRef] [PubMed]
- Lukić, N.; Schurr, F.M.; Trifković, T.; Kukavica, B.; Walter, J. Transgenerational stress memory in plants is mediated by upregulation of the antioxidative system. Environ. Exp. Bot. 2023, 205, 105129. [Google Scholar] [CrossRef]
- Mohammadi, M.; Afshari, R.T.; Nabati, J.; Oskoueian, E. Stress memory in seedlings of primed seed chickpea (Cicer arietinum L.) using sodium nitroprusside under cold stress. Plant Stress 2023, 8, 100163. [Google Scholar] [CrossRef]
- Geng, S.; Lin, Z.; Xie, S.; Xiao, J.; Wang, H.; Zhao, X.; Zhou, Y.; Duan, L. Ethylene enhanced waterlogging tolerance by changing root architecture and inducing aerenchyma formation in maize seedlings. J. Plant Physiol. 2023, 287, 154042. [Google Scholar] [CrossRef]
- Rossatto, T.; Souza, G.M.; Do Amaral, M.N.; Auler, P.A.; Pérez-Alonso, M.-M.; Pollmann, S.; Braga, E.J.B. Cross-stress memory: Salt priming at vegetative growth stages improves tolerance to drought stress during grain-filling in rice plants. Environ. Exp. Bot. 2023, 206, 105187. [Google Scholar] [CrossRef]
- Harshavardhan, V.T.; Govind, G.; Kalladan, R.; Sreenivasulu, N.; Hong, C.-Y. Cross-Protection by Oxidative Stress: Improving Tolerance to Abiotic Stresses Including Salinity. In Salinity Responses and Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1, pp. 283–305. [Google Scholar] [CrossRef]
- Johnson, R.; Puthur, J.T. Seed priming as a cost effective technique for developing plants with cross tolerance to salinity stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gai, Z.; Qiu, X.; Liu, T.; Li, S.; Ye, F.; Jian, S.; Shen, Y.; Li, X. Salt stress improves the low-temperature tolerance in sugar beet in which carbohydrate metabolism and signal transduction are involved. Environ. Exp. Bot. 2023, 208, 105239. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.; Hu, W.; Wang, S.; Snider, J.L.; Zhou, Z. Co-occurring elevated temperature and waterlogging stresses disrupt cellulose synthesis by altering the expression and activity of carbohydrate balance-associated enzymes during fiber development in cotton. Environ. Exp. Bot. 2017, 135, 106–117. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Hu, W.; Wang, S.; Snider, J.L.; Zhou, Z. Carbohydrate metabolism in the subtending leaf cross-acclimates to waterlogging and elevated temperature stress and influences boll biomass in cotton (Gossypium hirsutum). Physiol. Plant 2017, 161, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, Y.; Hu, W.; Snider, J.L.; Zhou, Z. Short-term soil-waterlogging contributes to cotton cross tolerance to chronic elevated temperature by regulating ROS metabolism in the subtending leaf. Plant Physiol. Biochem. 2019, 139, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, H.; Hu, W.; Wang, S.; Wang, Y.; Snider, J.L.; Zhou, Z. Combined elevated temperature and soil waterlogging stresses inhibit cell elongation by altering osmolyte composition of the developing cotton (Gossypium hirsutum L.) fiber. Plant Sci. 2017, 256, 196–207. [Google Scholar] [CrossRef]
- Pociecha, E.; Rapacz, M.; Dziurka, M.; Kolasinska, I. Mechanisms involved in the regulation of photosynthetic efficiency and carbohydrate partitioning in response to low- and high-temperature flooding triggered in winter rye (Secale cereale) lines with distinct pink snow mold resistances. Plant Physiol. Biochem. 2016, 104, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Jurczyk, B.; Rapacz, M.; Krępski, T. Photosynthetic apparatus responses to short-term low-temperature flooding may contribute to freezing tolerance changes in forage grasses. J. Agron. Crop Sci. 2015, 201, 49–56. [Google Scholar] [CrossRef]
- Jurczyk, B.; Krępski, T.; Kosmala, A.; Rapacz, M. Different mechanisms trigger an increase in freezing tolerance in Festuca pratensis exposed to flooding stress. Environ. Exp. Bot. 2013, 93, 45–54. [Google Scholar] [CrossRef]
- Maruyama, K.; Sakuma, Y.; Kasuga, M.; Ito, Y.; Seki, M.; Goda, H.; Shimada, Y.; Yoshida, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Identification of cold-inducible downstream genes of the Arabidopsis DREB1A/CBF3 transcriptional factor using two microarray systems. Plant J. 2004, 38, 982–993. [Google Scholar] [CrossRef]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [PubMed]
- Olorunwa, O.J.; Adhikari, B.; Brazel, S.; Shi, A.; Popescu, S.C.; Popescu, G.V.; Barickman, T.C. Growth and photosynthetic responses of cowpea genotypes under waterlogging at the reproductive stage. Plants 2022, 11, 2315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-M.; Duan, S.-G.; Xia, Y.; Li, J.-T.; Liu, L.-X.; Tang, M.; Tang, J.; Sun, W.; Yi, Y. Transcriptomic, physiological, and metabolomic response of an alpine Plant, Rhododendron delavayi, to waterlogging stress and post-waterlogging recovery. Int. J. Mol. Sci. 2023, 24, 10509. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Veen, H.V.; Perata, P. Plant responses to flooding stress. Curr. Opin. Plant Biol. 2016, 33, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Guo, J.; Zhang, Z.; Kou, T.; Deng, A.; Zheng, C.; Ren, J.; Zhang, W. Impacts of planting systems on soil moisture, soil temperature and corn yield in rainfed area of Northeast China. Eur. J. Agron. 2013, 50, 66–74. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Liu, G.; Lian, T.; Li, Z.; Liang, T.; Zhang, D.; Cui, Z.; Zhan, L.; Sun, L.; et al. Ridge intertillage alters rhizosphere bacterial communities and plant physiology to reduce yield loss of waterlogged cotton. Field Crops Res. 2023, 293, 108849. [Google Scholar] [CrossRef]
- Devkota, K.P.; Futakuchi, K.; Mel, V.C.; Humphreys, E. Does wet seeding combined with Sub1 varieties increase yield in submergence prone lowlands of West Africa? Field Crops Res. 2022, 276, 108375. [Google Scholar] [CrossRef]
- Fischer, R.A.; Moreno Ramos, O.H.; Ortiz Monasterio, I.; Sayre, K.D. Yield response to plant density, row spacing and raised beds in low latitude spring wheat with ample soil resources: An update. Field Crops Res. 2019, 232, 95–105. [Google Scholar] [CrossRef]
- Du, X.; He, W.; Wang, Z.; Xi, M.; Xu, Y.; Wu, W.; Gao, S.; Liu, D.; Lei, W.; Kong, L. Raised bed planting reduces waterlogging and increases yield in wheat following rice. Field Crops Res. 2021, 265, 108119. [Google Scholar] [CrossRef]
- Den Besten, N.; Steele Dunne, S.; Mahmud, A.; Jackson, D.; Aouizerats, B.; De Jeu, R.; Burger, R.; Houborg, R.; Mcglinchey, M.; Van Der Zaag, P. Understanding Sentinel-1 backscatter response to sugarcane yield variability and waterlogging. Remote Sens. Environ. 2023, 290, 113555. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, X.; Shen, J.; Du, K. Poplar’s waterlogging resistance modeling and evaluating: Exploring and perfecting the feasibility of machine learning methods in plant science. Front. Plant Sci. 2022, 13, 821365. [Google Scholar] [CrossRef]

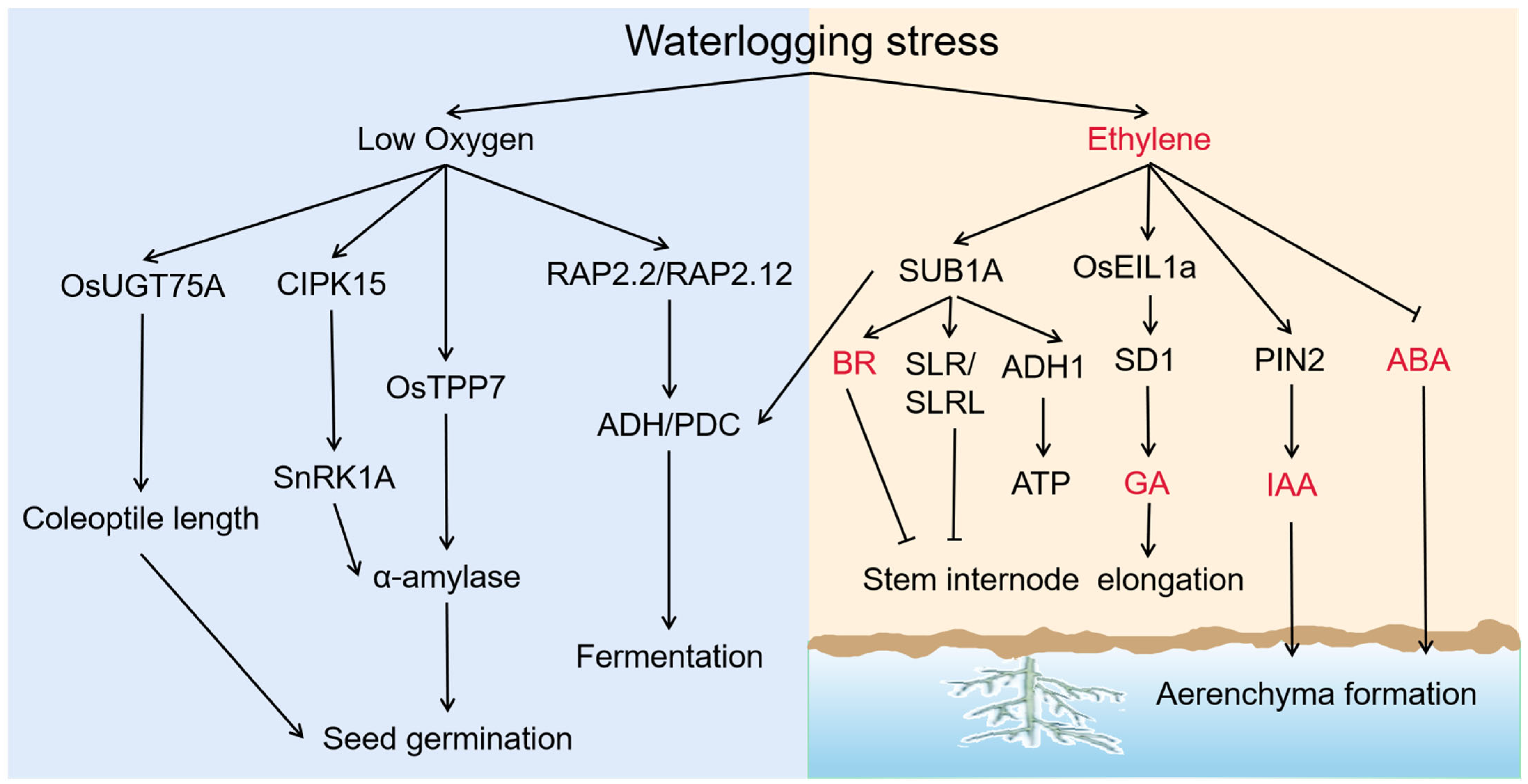
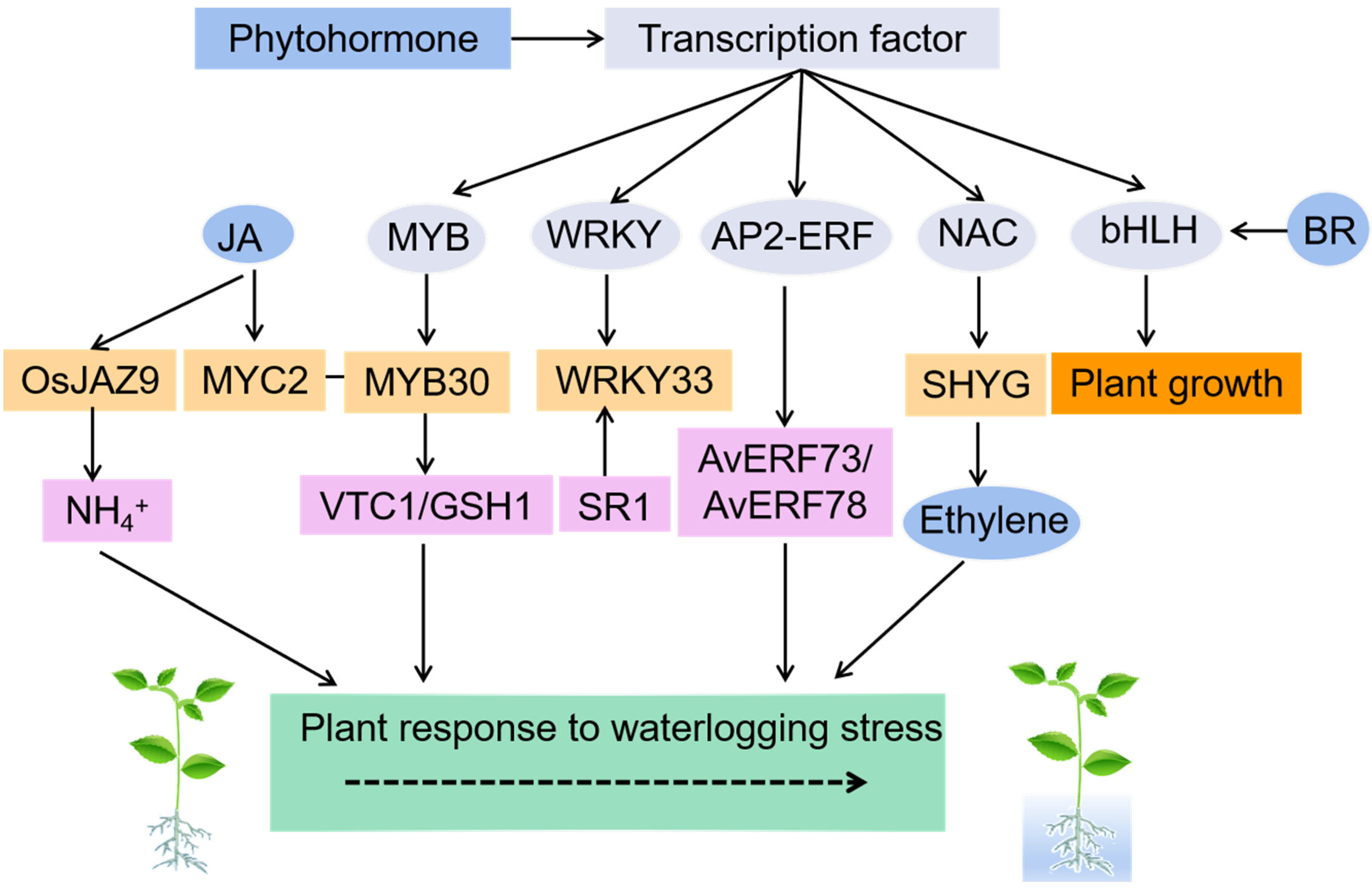

{kind=link}
{kind=link}
{kind=link}
| Plant Species | Genes | Ways | Genes or General Description | References |
|---|---|---|---|---|
| Oryza sativa | SUB1A | Overexpression | SUB1A mediates the expression of ethylene and GA and the overexpression of SUB1A enhances the transcription of ADH1 to improve the tolerance of rice under waterlogging stress. | [125] |
| Oryza sativa | OsCIPK15 | Overexpression | The CIPK15 gene is the main upstream positive regulator of SnRK1A, which can induce the expression of Amy3s and ultimately, mediate carbohydrate catabolism under hypoxic stress. | [60] |
| Oryza sativa | OsGA20ox2(SD1) | Overexpression | When submerged, the SD1 gene is activated by the ethylene-responsive transcription factor OsEIL1a and the overexpression of SD1 promotes the synthesis of GA, mainly GA4, and facilitates the rapid growth of rice leaf and stem internodes. | [128] |
| Oryza sativa | LG1 | Overexpression | LGF1 was identified as the gene that determines leaf air film. LGF1 regulates the synthesis of C30 primary alcohols, increases underwater photosynthesis by 8.2-fold, and contributes to underwater tolerance. | [107] |
| A. thaliana | AdPDC1 | Overexpression | The AdPDC1 gene in Arabidopsis inhibited seed germination and root length under ABA treatment, while overexpression of the kiwifruit PDC1 gene in Arabidopsis enhanced waterlogging tolerance suggesting that ABA negatively regulated the waterlogging tolerance. | [129] |
| A. thaliana | BnERF2.4 | Overexpression | The induced expression of BnERF2.4 in Arabidopsis showed the increased tolerance of plants to submergence and was involved in regulating soluble sugar content and the antioxidant system in defense against submergence stress. | [85] |
| A. thaliana | HvERF2.11 | Overexpression | Under waterlogging stress, overexpression of HvERF2.11 in Arabidopsis rapidly increases the expression of AtSOD1, AtPOD1, and AtACO1 and activates the antioxidant enzymes including ADH significantly. | [123] |
| A. thaliana | AtACO5 | Overexpression | Overexpression of AtACO5 (ACC OXIDASE 5) in Arabidopsis triggers ethylene production and cellular expansion activity enhances tolerance to waterlogging stress. | [166] |
| A. thaliana | HRE2 | Overexpression | HRE2 overexpression promotes ARs formation and elongation growth of ARs primordia. | [26] |
| A. thaliana | ERF73/ HRE1 | Overexpression | ERF73/HRE1 induced expression positively regulates ADH activity which was analyzed as a marker enzyme for metabolic adaptation to hypoxic stress and ethylene regulates metabolic adaptation to low oxygen stress in the Arabidopsis root through ERF73/HRE1. | [158] |
| Zea mays | ZmEREB180 | Overexpression | Overexpression of ZmEREB180 also enhanced survival rate after long-term flooding stress, apparently by enhancing ARs formation and modulating antioxidant levels. | [25] |
| Cherry | PsERF/PsCIPK | Overexpression | The activities of sucrose synthase (SUS), alpha-amylase (Ramy), ADH, and PDC in the leaves of PsERF/PsCIPK gene could resist waterlogging stress by regulating sugar metabolism. | [138] |
| Glycine max | GmADH2 | Overexpression | Enhanced germination ability of transgenic soybean seeds was detected through the induction of the GmADH2 gene during the process of glycolysis and ethanol fermentation under flooding stress. | [50] |
| Petunia × hybrida | PhERF2 | Overexpression | Overexpressing of PhERF2 promotes growth vigor and contributes to petunia waterlogging tolerance through modulation of PCD and alcoholic fermentation system. | [122] |
| Chrysanthemum | CmSOS1 | Overexpression | CmSOS1 coordinately mediates waterlogging tolerance by maintaining membrane integrity and minimizing the level of reactive oxygen species. | [87] |
| Cucumis sativus/A.thaliana | CsARN6.1 | Overexpression | CsARN6.1 played an important role in increasing cucumber and Arabidopsis ARs formation via its ATPase activity to promote waterlogging tolerance. | [4] |
| Nicotiana tabacum | AdRAP2.3 | Overexpression | Inducible expression levels of NtPDC, NtADH, NtHB1, NtHB2, NtPCO1, and NtPCO2 genes through the overexpression of AdRAP2.3 in tobacco plants are associated with enhanced waterlogging tolerance. | [52] |
| Triticum aestivum | TaERFVII.1 | Overexpression and VIGS | Constitutive expression of TaERFVII.1 with an MYC-peptide tag at its N terminus in wheat enhanced tolerance to waterlogging as evidenced by increasing grain weight per plant, survival rate, and chlorophyll content of leaves and improving the expression of waterlogging-responsive genes while silencing of TaERFVII.1 compromised the expression of waterlogging-responsive genes. | [162] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Li, N.; Liu, Y.; Miao, P.; Liu, J.; Wang, Z. Updates and Prospects: Morphological, Physiological, and Molecular Regulation in Crop Response to Waterlogging Stress. Agronomy 2023, 13, 2599. https://doi.org/10.3390/agronomy13102599
Yang L, Li N, Liu Y, Miao P, Liu J, Wang Z. Updates and Prospects: Morphological, Physiological, and Molecular Regulation in Crop Response to Waterlogging Stress. Agronomy. 2023; 13(10):2599. https://doi.org/10.3390/agronomy13102599
Chicago/Turabian StyleYang, Lu, Na Li, Yang Liu, Pengfei Miao, Ji Liu, and Zhi Wang. 2023. "Updates and Prospects: Morphological, Physiological, and Molecular Regulation in Crop Response to Waterlogging Stress" Agronomy 13, no. 10: 2599. https://doi.org/10.3390/agronomy13102599
APA StyleYang, L., Li, N., Liu, Y., Miao, P., Liu, J., & Wang, Z. (2023). Updates and Prospects: Morphological, Physiological, and Molecular Regulation in Crop Response to Waterlogging Stress. Agronomy, 13(10), 2599. https://doi.org/10.3390/agronomy13102599