
Soil Microbial Community Composition and Diversity Are Insusceptible to Nitrogen Addition in a Semi-Arid Grassland in Northwestern China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
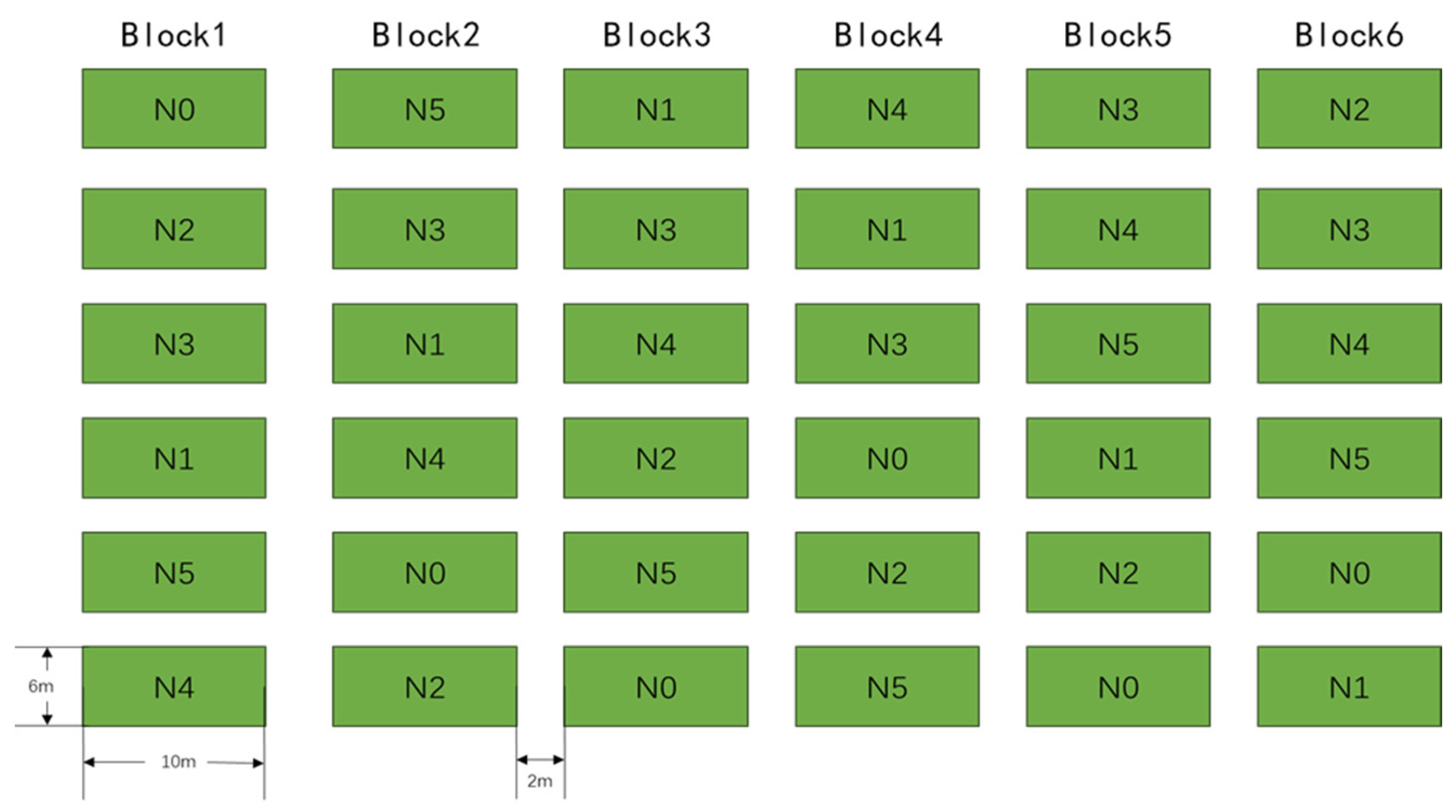
2.2. Experimental Design and Sampling
2.3. Leaf functional Trait Sample and Measurements
2.4. Determination Index and Method
2.5. DNA Extractions and High-Throughput Sequencing
2.6. Data Analysis
3. Results
3.1. The Vegetation Characteristics and Soil Properties across N Addition Gradients
3.2. The Microbial Communities across N Addition Gradients
3.2.1. Diversity of Microbial Communities
3.2.2. Composition of Microbial Communities
3.2.3. Assembly and Species Turnover of Microbial Communities
3.2.4. Drivers of Microbial Communities
4. Discussion
4.1. Changes in Vegetation Characteristics and Soil Properties under N Addition
4.2. Changes in Microbial Community Diversity and Assembly under N Addition
4.3. Microbial Community Composition and Its Driving Factors under N Addition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.X.; Li, W.B.; Adams, H.D.; Wang, A.Z.; Wu, J.B.; Jin, C.J.; Guan, D.; Yuan, F.H. Responses of woody plant functional traits to nitrogen addition: A meta-analysis of leaf economics, gas exchange, and hydraulic traits. Front. Pant Sci. 2018, 9, 683. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.J.; Tang, A.; Zhang, Y.; Luo, X.S.; Shen, J.L.; Wang, W.; Li, K.H.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Luo, X.S.; Pan, Y.P.; Zhang, L.; Tang, A.H.; Shen, J.L.; Zhang, Y.; Li, K.H.; Wu, Q.H.; Yang, D.W.; et al. Quantifying atmospheric nitrogen deposition through a nationwide monitoring network across China. Atmos. Chem. Phys. Discuss. 2015, 15, 18365–18405. [Google Scholar] [CrossRef]
- Jones, L.; Provins, A.; Holland, M.; Mills, G.; Hayes, F.; Emmett, B.; Hall, J.; Sheppard, L.; Smith, R.; Sutton, M.; et al. A review and application of the evidence for nitrogen impacts on ecosystem services. Ecosyst. Serv. 2014, 7, 76–88. [Google Scholar] [CrossRef]
- Zhu, J.X.; Chen, Z.; Wang, Q.F.; Xu, L.; He, N.P.; Jia, Y.L.; Zhang, Q.Y.; Yu, G.R. Potential transition in the effects of atmospheric nitrogen deposition in China. Environ. Pollut. 2020, 258, 113739. [Google Scholar] [CrossRef]
- Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Wilcox, K.R.; Wilson, G.W.T.; Smith, M.D.; Collins, S.L. Changes in plant community composition, not diversity, during a decade of nitrogen and phosphorus additions drive above-ground productivity in a tallgrass prairie. J. Ecol. 2014, 102, 1649–1660. [Google Scholar] [CrossRef]
- Lu, X.K.; Vitousek, P.M.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Turner, B.L.; Zhou, G.Y.; Mo, J.M. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. Proc. Natl. Acad. Sci. USA 2021, 118, e2020790118. [Google Scholar] [CrossRef]
- Sun, R.B.; Chen, Y.; Han, W.X.; Dong, W.X.; Zhang, Y.M.; Hu, C.S.; Liu, B.B.; Wang, F.H. Different contribution of species sorting and exogenous species immigration from manure to soil fungal diversity and community assemblage under long-term fertilization. Soil Biol. Biochem. 2020, 151, 108049. [Google Scholar] [CrossRef]
- Yang, H.J.; Li, Y.; Wu, M.Y.; Zhang, Z.; Li, L.H.; Wan, S.Q. Plant community responses to nitrogen addition and increased precipitation: The importance of water availability and species traits. Glob. Chang. Biol. 2011, 17, 2936–2944. [Google Scholar] [CrossRef]
- Liu, X.J.; Duan, L.; Mo, J.M.; Du, E.Z.; Shen, J.L.; Lu, X.K.; Zhang, Y.; Zhou, X.B.; He, C.; Zhang, F.S. Nitrogen deposition and its ecological impact in China: An overview. Environ. Pollut. 2011, 159, 2251–2264. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lu, X.K.; Mori, T.; Mao, Q.G.; Zhou, K.J.; Zhou, G.Y.; Nie, Y.X.; Mo, J.M. Responses of soil microbial community to continuous experimental nitrogen additions for 13 years in a nitrogen-rich tropical forest. Soil Biol. Biochem. 2018, 121, 103–112. [Google Scholar] [CrossRef]
- Lv, F.L.; Xue, S.; Wang, G.L.; Zhang, C. Nitrogen addition shifts the microbial community in the rhizosphere of Pinus tabuliformis in northwestern China. PLoS ONE 2017, 12, e0172382. [Google Scholar] [CrossRef]
- Liu, C.; Yao, M.J.; Stegen, J.C.; Rui, J.P.; Li, J.B.; Li, X.Z. Long-term nitrogen addition affects the phylogenetic turnover of soil microbial community responding to moisture pulse. Sci. Rep. 2017, 7, 17492. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sang, C.P.; Yang, J.Y.; Qu, L.R.; Xia, Z.W.; Sun, H.; Jiang, P.; Wang, X.G.; He, H.B.; Wang, C. Stoichiometric imbalance and microbial community regulate microbial elements use efficiencies under nitrogen addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Zhang, T.A.; Chen, H.Y.H.; Ruan, H.H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.Y.; Yuan, X.C.; Zhang, Q.F.; Zhou, J.C.; Lin, K.M.; Xu, J.G.; Zeng, Y.Z.; Wu, Y.; Cheng, L.; Zeng, Q.X.; et al. Nutrient availability is a dominant predictor of soil bacterial and fungal community composition after nitrogen addition in subtropical acidic forests. PLoS ONE 2021, 16, e0246263. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef]
- Widdig, M.; Heintz-Buschart, A.; Schleuss, P.M.; Guhr, A.; Borer, E.T.; Seabloom, E.W.; Spohn, M. Effects of nitrogen and phosphorus addition on microbial community composition and element cycling in a grassland soil. Soil Biol. Biochem. 2020, 151, 108041. [Google Scholar] [CrossRef]
- Birrer, S.C.; Dafforn, K.A.; Sun, M.Y.; Williams, R.B.H.; Potts, J.; Scanes, P.; Kelaher, B.P.; Simpson, S.L.; Kjelleberg, S.; Swarup, S.; et al. Using meta-omics of contaminated sediments to monitor changes in pathways relevant to climate regulation. Environ. Microbiol. 2019, 21, 389–401. [Google Scholar] [CrossRef]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B.H. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Trivedi, C.; Hu, H.W.; Anderson, I.C.; Jeffries, T.C.; Zhou, J.Z.; Singh, B.K. Microbial regulation of the soil carbon cycle: Evidence from gene-enzyme relationships. ISME J. 2016, 10, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Yevdokimov, I.; Larionova, A.; Blagodatskaya, E. Microbial immobilisation of phosphorus in soils exposed to drying-rewetting and freeze-thawing cycles. Biol. Fertil. Soils 2016, 52, 685–696. [Google Scholar] [CrossRef]
- Chen, Q.L.; Ding, J.; Li, C.Y.; Yan, Z.Z.; He, J.Z.; Hu, H.W. Microbial functional attributes, rather than taxonomic attributes, drive top soil respiration, nitrification and denitrification processes. Sci. Total Environ. 2020, 734, 139479. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.T.; Xiao, X.; Nuccio, E.E.; Yuan, M.T.; Zhang, N.; Xue, K.; Cohan, F.M.; Zhou, J.Z.; Sun, B. Differentiation strategies of soil rare and abundant microbial taxa in response to changing climatic regimes. Environ. Microbiol. 2020, 22, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Toju, H.; Peay, K.G.; Yamamichi, M.; Narisawa, K.; Hiruma, K.; Naito, K.; Fukuda, S.; Ushio, M.; Nakaoka, S.; Onoda, Y.; et al. Core microbiomes for sustainable agroecosystems. Nat. Plants 2018, 4, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.D.; Ren, K.X.; Isabwe, A.; Chen, H.H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2019, 7, 138, Erratum in Microbiome 2019, 7, 148. [Google Scholar] [CrossRef]
- Perronne, R.; Munoz, F.; Borgy, B.; Reboud, X.; Gaba, S. How to design trait-based analyses of community assembly mechanisms: Insights and guidelines from a literature review. Perspect. Plant Ecol. Evol. Syst. 2017, 25, 29–44. [Google Scholar] [CrossRef]
- Osburn, E.D.; Aylward, F.O.; Barrett, J.E. Historical land use has long-term effects on microbial community assembly processes in forest soils. ISME Commun. 2021, 1, 48. [Google Scholar] [CrossRef]
- Mo, Y.Y.; Peng, F.; Gao, X.F.; Xiao, P.; Logares, R.; Jeppesen, E.; Ren, K.X.; Xue, Y.Y.; Yang, J. Low shifts in salinity determined assembly processes and network stability of microeukaryotic plankton communities in a subtropical urban reservoir. Microbiome 2021, 9, 128. [Google Scholar] [CrossRef]
- Luan, L.; Liang, C.; Chen, L.J.; Wang, H.T.; Xu, Q.S.; Jiang, Y.J.; Sun, B. Coupling Bacterial Community Assembly to Microbial Metabolism across Soil Profiles. mSystems 2020, 5, e00298-20. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Romdhane, S.; Spor, A.; Aubert, J.; Bru, D.; Breuil, M.-C.; Hallin, S.; Mounier, A.; Ouadah, S.; Tsiknia, M.; Philippot, L. Unraveling negative biotic interactions determining soil microbial community assembly and functioning. ISME J. 2022, 16, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Chevallereau, A.; Pons, B.J.; van Houte, S.; Westra, E.R. Interactions between bacterial and phage communities in natural environments. Nat. Rev. Microbiol. 2022, 20, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, S.R.; Zhang, X.; Mao, Q.G.; Li, X.Z.; You, Y.M.; Wang, J.X.; Zheng, M.H.; Zhang, W.; Lu, X.K.; et al. Nitrogen addition reduces soil bacterial richness, while phosphorus addition alters community composition in an old-growth N-rich tropical forest in southern China. Soil Biol. Biochem. 2018, 127, 22–30. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhu, K.; Chiariello, N.R.; Field, C.B.; Peay, K.G. Fire history and plant community composition outweigh decadal multi-factor global change as drivers of microbial composition in an annual grassland. J. Ecol. 2020, 108, 611–625. [Google Scholar] [CrossRef]
- Xiao, Y.M.; Li, C.B.; Yang, Y.; Peng, Y.F.; Yang, Y.H.; Zhou, G.Y. Soil fungal community composition, not assembly process, was altered by nitrogen addition and precipitation changes at an alpine steppe. Front. Microbiol. 2020, 11, 579072. [Google Scholar] [CrossRef]
- Craig, H.; Antwis, R.E.; Cordero, I.; Ashworth, D.; Robinson, C.H.; Osborne, T.Z.; Bardgett, R.D.; Rowntree, J.K.; Simpson, L.T. Nitrogen addition alters composition, diversity, and functioning of microbial communities in mangrove soils: An incubation experiment. Soil Biol. Biochem. 2021, 153, 108076. [Google Scholar] [CrossRef]
- Guo, Q.; Wen, Z.M.; Ghanizadeh, H.; Fan, Y.M.; Zheng, C.; Yang, X.; Yan, X.H.; Li, W. Stochastic processes dominate assembly of soil fungal community in grazing excluded grasslands in northwestern China. J. Soils Sediments 2023, 23, 156–171. [Google Scholar] [CrossRef]
- Friedman, J.; Alm, E.J. Inferring correlation networks from genomic survey data. PLoS Comput. Biol. 2012, 8, e1002687. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Urbach, N.; Ahlemann, F. Structural equation modeling in information systems research using partial least squares. J. Inf. Technol. Theory Appl. JITTA 2010, 11, 5–40. [Google Scholar]
- Hair, J.F.; Ringle, C.M.; Sarstedt, M. PLS-SEM : Indeed a silver bullet. J. Mark. Theory Pract. 2011, 19, 139–152. [Google Scholar] [CrossRef]
- Tatarko, A.R.; Knops, J.M.H. Nitrogen addition and ecosystem functioning: Both species abundances and traits alter community structure and function. Ecosphere 2018, 9, e02087. [Google Scholar] [CrossRef]
- Xu, L.C.; Xing, A.J.; Du, E.Z.; Shen, H.H.; Yan, Z.B.; Jiang, L.; Tian, D.; Hu, H.F.; Fang, J.Y. Effects of nitrogen addition on leaf nutrient stoichiometry in an old-growth boreal forest. Ecosphere 2021, 12, e03335. [Google Scholar] [CrossRef]
- Zheng, L.L.; Zhao, Q.; Yu, Z.Y.; Zhao, S.Y.; Zeng, D.H. Altered leaf functional traits by nitrogen addition in a nutrient-poor pine plantation: A consequence of decreased phosphorus availability. Sci. Rep. 2017, 7, 7415. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Alonso, R.; Janssens, I.A.; Carnicer, J.; Vereseglou, S.; Rillig, M.C.; Fernández-Martínez, M.; Sanders, T.G.M.; Peñuelas, J. Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: Relationships with climate, N deposition and tree growth. Funct. Ecol. 2016, 30, 676–689. [Google Scholar] [CrossRef]
- You, C.M.; Wu, F.Z.; Yang, W.Q.; Xu, Z.F.; Tan, B.; Yue, K.; Ni, X.Y. Nutrient-limited conditions determine the responses of foliar nitrogen and phosphorus stoichiometry to nitrogen addition: A global meta-analysis. Environ. Pollut. 2018, 241, 740–749. [Google Scholar] [CrossRef]
- Deng, Q.; Hui, D.F.; Dennis, S.; Reddy, K.C. Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta-analysis. Glob. Ecol. Biogeogr. 2017, 26, 713–728. [Google Scholar] [CrossRef]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yang, Y.H.; Luo, Y.Q.; Fang, C.M.; Zhou, X.H.; Chen, J.K.; Yang, X.; Li, B. Responses of ecosystem nitrogen cycle to nitrogen addition: A meta-analysis. New Phytol. 2011, 189, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheng, H.; Gao, H.; An, S.S. Response and driving factors of soil microbial diversity related to global nitrogen addition. Land. Degrad. Dev. 2020, 31, 190–204. [Google Scholar] [CrossRef]
- Dai, Z.M.; Su, W.Q.; Chen, H.H.; Barberán, A.; Zhao, H.C.; Yu, M.J.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.W.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Wu, J.P.; Liu, W.F.; Zhang, W.X.; Shao, Y.H.; Duan, H.L.; Chen, B.D.; Wei, X.H.; Fan, H.B. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, X.J.; Song, L.; Lin, X.G.; Zhang, H.Z.; Shen, C.C.; Chu, H.Y. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K.; Luo, Y.Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.W.; Yang, S.; Li, X.B.; Top, E.M.; Wang, R.Z.; Zhang, Y.G.; Cai, J.P.; Yao, F.; Han, X.G.; et al. Responses of soil bacterial communities to nitrogen deposition and precipitation increment are closely linked with aboveground community variation. Microb. Ecol. 2016, 71, 974–989. [Google Scholar] [CrossRef]
- Liu, J.W.; Meng, Z.; Liu, X.Y.; Zhang, X.H. Microbial assembly, interaction, functioning, activity and diversification: A review derived from community compositional data. Mar. Life Sci. Technol. 2019, 1, 112–128. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; Van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.F.; Zhang, J.J.; Tu, Q.C.; Deng, Y.; Qiu, Q.F.; Xiong, J.B. Bacterioplankton assembly and interspecies interaction indicating increasing coastal eutrophication. Chemosphere 2017, 177, 317–325. [Google Scholar] [CrossRef]
- Freedman, Z.B.; Romanowicz, K.J.; Upchurch, R.A.; Zak, D.R. Differential responses of total and active soil microbial communities to long-term experimental N deposition. Soil Biol. Biochem. 2015, 90, 275–282. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.A.; Wang, F.M.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.F.; Li, Y.W.; Li, X.B.; Xia, H.P. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Markovskaja, S.; Kačergius, A. Morphological and molecular characterisation of Periconia pseudobyssoides sp. nov. and closely related P. byssoides. Mycol. Prog. 2014, 13, 291–302. [Google Scholar] [CrossRef]
- Neuhauser, S.; Huber, L.; Kirchmair, M. Is Roesleria subterranea a primary pathogen or a minor parasite of grapevines? Risk assessment and a diagnostic decision scheme. Eur. J. Plant Pathol. 2011, 130, 503–510. [Google Scholar] [CrossRef]
- Carey, C.J.; Michael Beman, J.; Eviner, V.T.; Malmstrom, C.M.; Hart, S.C. Soil microbial community structure is unaltered by plant invasion, vegetation clipping, and nitrogen fertilization in experimental semi-arid grasslands. Front. Microbiol. 2015, 6, 466. [Google Scholar] [CrossRef]
- McHugh, T.A.; Morrissey, E.M.; Mueller, R.C.; Gallegos-Graves, L.V.; Kuske, C.R.; Reed, S.C. Bacterial, fungal, and plant communities exhibit no biomass or compositional response to two years of simulated nitrogen deposition in a semiarid grassland. Environ. Microbiol. 2017, 19, 1600–1611. [Google Scholar] [CrossRef]
- She, W.W.; Bai, Y.X.; Zhang, Y.Q.; Qin, S.G.; Feng, W.; Sun, Y.F.; Zheng, J.; Wu, B. Resource availability drives responses of soil microbial communities to short-term precipitation and nitrogen addition in a desert shrubland. Front. Microbiol. 2018, 9, 186. [Google Scholar] [CrossRef]
- Huang, J.Y.; Xu, Y.X.; Yu, H.L.; Zhu, W.W.; Wang, P.; Wang, B.; Na, X.F. Soil prokaryotic community shows no response to 2 years of simulated nitrogen deposition in an arid ecosystem in northwestern China. Environ. Microbiol. 2021, 23, 1222–1237. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.X.; Zhang, Y.L.; Duan, C.J.; Wang, X.; Zhang, X.C.; Ju, W.L.; Chen, H.S.; Yue, S.C.; Wang, Y.Q.; Li, S.Q.; et al. Ecoenzymatic stoichiometry reveals microbial phosphorus limitation decreases the nitrogen cycling potential of soils in semi-arid agricultural ecosystems. Soil Tillage Res. 2020, 197, 104463. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef] [PubMed]
- Sagova-Mareckova, M.; Omelka, M.; Cermak, L.; Kamenik, Z.; Olsovska, J.; Hackl, E.; Kopecky, J.; Hadacek, F. Microbial communities show parallels at sites with distinct litter and soil characteristics. Appl. Environ. Microbiol. 2011, 77, 7560–7567. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Wippel, K.; Tao, K.; Niu, Y.L.; Zgadzaj, R.; Kiel, N.; Guan, R.; Dahms, E.; Zhang, P.F.; Jensen, D.B.; Logemann, E.; et al. Host preference and invasiveness of commensal bacteria in the Lotus and Arabidopsis root microbiota. Nat. Microbiol. 2021, 6, 1150–1162. [Google Scholar] [CrossRef]
- Nie, Y.X.; Wang, M.C.; Zhang, W.; Ni, Z.; Hashidoko, Y.; Shen, W.J. Ammonium nitrogen content is a dominant predictor of bacterial community composition in an acidic forest soil with exogenous nitrogen enrichment. Sci. Total Environ. 2018, 624, 407–415. [Google Scholar] [CrossRef]
- Niu, G.X.; Hasi, M.; Wang, R.Z.; Wang, Y.L.; Geng, Q.Q.; Hu, S.Y.; Xu, X.H.; Yang, J.J.; Wang, C.H.; Han, X.G.; et al. Soil microbial community responses to long-term nitrogen addition at different soil depths in a typical steppe. Appl. Soil Ecol. 2021, 167, 104054. [Google Scholar] [CrossRef]
- Yao, M.J.; Rui, J.P.; Li, J.B.; Dai, Y.M.; Bai, Y.F.; Heděnec, P.; Wang, J.M.; Zhang, S.H.; Pei, K.Q.; Liu, C.; et al. Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. Soil Biol. Biochem. 2014, 79, 81–90. [Google Scholar] [CrossRef]
- Zhou, J.; Guan, D.W.; Zhou, B.K.; Zhao, B.S.; Ma, M.C.; Qin, J.; Jiang, X.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | N0 | N1 | N2 | N3 | N4 | N5 | F | p |
|---|---|---|---|---|---|---|---|---|
| CWM.LC (g kg−1) | 490.28 ± 3.30 a | 480.29 ± 5.05 abc | 474.46 ± 3.78 bc | 470.01 ± 3.85 c | 486.73 ± 5.54 ab | 482.96 ± 7.23 abc | 2.318 | 0.068 |
| CWM.LN (g kg−1) | 14.96 ± 0.30 d | 17.93 ± 0.83 c | 18.30 ± 0.51 c | 20.27 ± 0.41 b | 21.81 ± 0.57 ab | 23.11 ± 1.09 a | 19.240 | <0.001 |
| CWM.LP (g kg−1) | 1.32 ± 0.04 a | 1.22 ± 0.11 ab | 1.16 ± 0.03 ab | 1.09 ± 0.05 b | 1.08 ± 0.06 b | 1.08 ± 0.10 b | 2.013 | 0.105 |
| CWM.SLA (cm−2 g−1) | 143.17 ± 11.02 a | 126.75 ± 8.52 ab | 131.21 ± 5.18 ab | 116.24 ± 5.98 b | 109.62 ± 2.78 b | 113.69 ± 9.76 b | 2.263 | 0.074 |
| Foliar N/P ratio | 11.29 ± 0.49 d | 14.97 ± 1.75 c | 15.79 ± 1.67 c | 18.84 ± 2.21 b | 20.33 ± 1.56 ab | 21.79 ± 2.62 a | 26.845 | <0.001 |
| FDis | 1.43 ± 0.11 | 1.34 ± 0.07 | 1.28 ± 0.11 | 1.55 ± 0.10 | 1.29 ± 0.16 | 1.19 ± 0.22 | 0.869 | 0.513 |
| S_plant | 13.33 ± 1.02 a | 11.50 ± 1.12 ab | 13.33 ± 0.95 a | 9.50 ± 1.15 b | 9.67 ± 0.88 b | 8.67 ± 1.17 b | 3.704 | 0.010 |
| H_plant | 2.27 ± 0.10 a | 2.01 ± 0.10 ab | 2.21 ± 0.09 a | 1.88 ± 0.13 b | 1.81 ± 0.10 b | 1.73 ± 0.12 b | 4.023 | 0.007 |
| Aboveground biomass (g m−2) | 202.02 ± 17.73 b | 279.95 ± 36.95 b | 304.64 ± 41.77 b | 296.11 ± 71.13 b | 484.54 ± 108.55 a | 491.15 ± 23.48 a | 2.492 | 0.041 |
| Parameters | N0 | N1 | N2 | N3 | N4 | N5 | F | p |
|---|---|---|---|---|---|---|---|---|
| OC (g kg−1) | 24.90 ± 1.55 | 24.88 ± 1.91 | 27.80 ± 1.43 | 26.37 ± 2.58 | 27.13 ± 1.14 | 26.38 ± 0.49 | 0.508 | 0.765 |
| TN (g kg−1) | 2.63 ± 0.32 | 2.60 ± 0.33 | 2.93 ± 0.30 | 2.77 ± 0.36 | 2.61 ± 0.58 | 2.71 ± 0.15 | 0.367 | 0.862 |
| AN (mg kg−1) | 67.90 ± 4.28 | 70.00 ± 4.50 | 78.40 ± 5.30 | 82.37 ± 5.75 | 77.70 ± 2.52 | 72.37 ± 6.96 | 1.177 | 0.376 |
| TP (g kg−1) | 0.62 ± 0.04 | 0.61 ± 0.03 | 0.62 ± 0.01 | 0.60 ± 0.04 | 0.62 ± 0.01 | 0.61 ± 0.02 | 0.260 | 0.926 |
| AP (mg kg−1) | 4.15 ± 0.06 a | 3.18 ± 0.40 b | 3.12 ± 0.30 b | 2.17 ± 0.08 c | 2.21 ± 0.35 c | 2.82 ± 0.32 bc | 6.638 | 0.003 |
| pH | 8.25 ± 0.05 | 8.26 ± 0.04 | 8.22 ± 0.04 | 8.30 ± 0.03 | 8.27 ± 0.04 | 8.26 ± 0.05 | 0.393 | 0.845 |
| SWC (%) | 21.55 ± 0.90 | 21.53 ± 0.75 | 23.95 ± 0.51 | 23.34 ± 1.43 | 23.41 ± 1.21 | 22.52 ± 1.88 | 0.677 | 0.649 |
| Parameters | N0 | N1 | N2 | N3 | N4 | N5 | F | p |
|---|---|---|---|---|---|---|---|---|
| B_ACE | 4139.95 ± 61.12 | 4115.67 ± 166.22 | 4270.66 ± 54.50 | 4312.69 ± 26.44 | 4266.26 ± 83.52 | 4125.14 ± 162.72 | 0.670 | 0.654 |
| B_Chao1 | 4034.67 ± 59.34 | 4000.78 ± 160.24 | 4158.49 ± 46.59 | 4193.21 ± 42.89 | 4184.37 ± 71.90 | 4041.05 ± 159.53 | 0.692 | 0.639 |
| B-Shannon | 9.70 ± 0.02 | 9.53 ± 0.16 | 9.71 ± 0.05 | 9.72 ± 0.06 | 9.69 ± 0.07 | 9.66 ± 0.10 | 0.650 | 0.667 |
| F_ACE | 2460.94 ± 207.22 | 2051.16 ± 272.71 | 2109.25 ± 158.80 | 2738.53 ± 445.16 | 2508.74 ± 253.72 | 2263.51 ± 298.78 | 0.833 | 0.551 |
| F_Chao1 | 2382.11 ± 195.04 | 2005.34 ± 306.95 | 2024.51 ± 142.49 | 2699.50 ± 469.47 | 2453.67 ± 264.52 | 2206.98 ± 267.11 | 0.843 | 0.545 |
| F-Shannon | 7.30 ± 0.43 ab | 6.74 ± 0.30 ab | 6.52 ± 0.13 b | 7.81 ± 0.30 a | 6.93 ± 0.55 ab | 6.84 ± 0.55 ab | 1.304 | 0.325 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuo, H.; Li, M.; Ghanizadeh, H.; Huang, J.; Yang, M.; Wang, Z.; Wang, Y.; Tian, H.; Ye, F.; Li, W. Soil Microbial Community Composition and Diversity Are Insusceptible to Nitrogen Addition in a Semi-Arid Grassland in Northwestern China. Agronomy 2023, 13, 2593. https://doi.org/10.3390/agronomy13102593
Tuo H, Li M, Ghanizadeh H, Huang J, Yang M, Wang Z, Wang Y, Tian H, Ye F, Li W. Soil Microbial Community Composition and Diversity Are Insusceptible to Nitrogen Addition in a Semi-Arid Grassland in Northwestern China. Agronomy. 2023; 13(10):2593. https://doi.org/10.3390/agronomy13102593
Chicago/Turabian StyleTuo, Hanghang, Meihui Li, Hossein Ghanizadeh, Jiandi Huang, Mengru Yang, Zilin Wang, Yibo Wang, Huihui Tian, Faming Ye, and Wei Li. 2023. "Soil Microbial Community Composition and Diversity Are Insusceptible to Nitrogen Addition in a Semi-Arid Grassland in Northwestern China" Agronomy 13, no. 10: 2593. https://doi.org/10.3390/agronomy13102593
APA StyleTuo, H., Li, M., Ghanizadeh, H., Huang, J., Yang, M., Wang, Z., Wang, Y., Tian, H., Ye, F., & Li, W. (2023). Soil Microbial Community Composition and Diversity Are Insusceptible to Nitrogen Addition in a Semi-Arid Grassland in Northwestern China. Agronomy, 13(10), 2593. https://doi.org/10.3390/agronomy13102593