
Genome-Wide Association Study for Spike Traits and Distribution of Two QTLs for Grain Number in Chinese Wheat Cultivars


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Field Trials
2.2. Phenotyping and Data Analysis
2.3. Genome-Wide Association Study and Linkage Disequilibrium Analysis
2.4. Haplotype Analysis
3. Results
3.1. Variation and Correlation Analysis of Spike-Related Traits
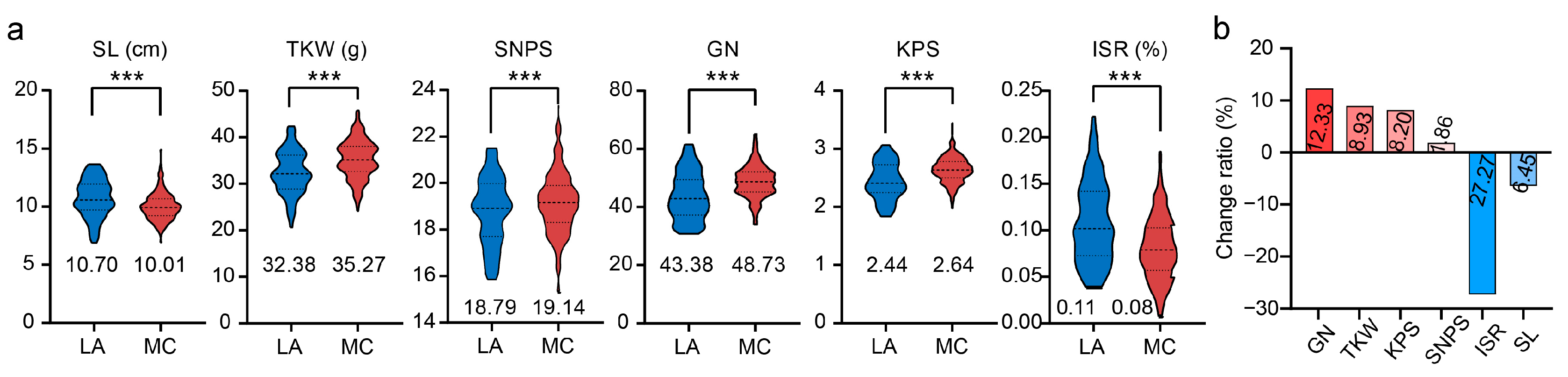
3.2. Variation of Spike-Related Traits during Modern Breeding
3.3. GWAS of Spike-Related Traits in Seven Environments
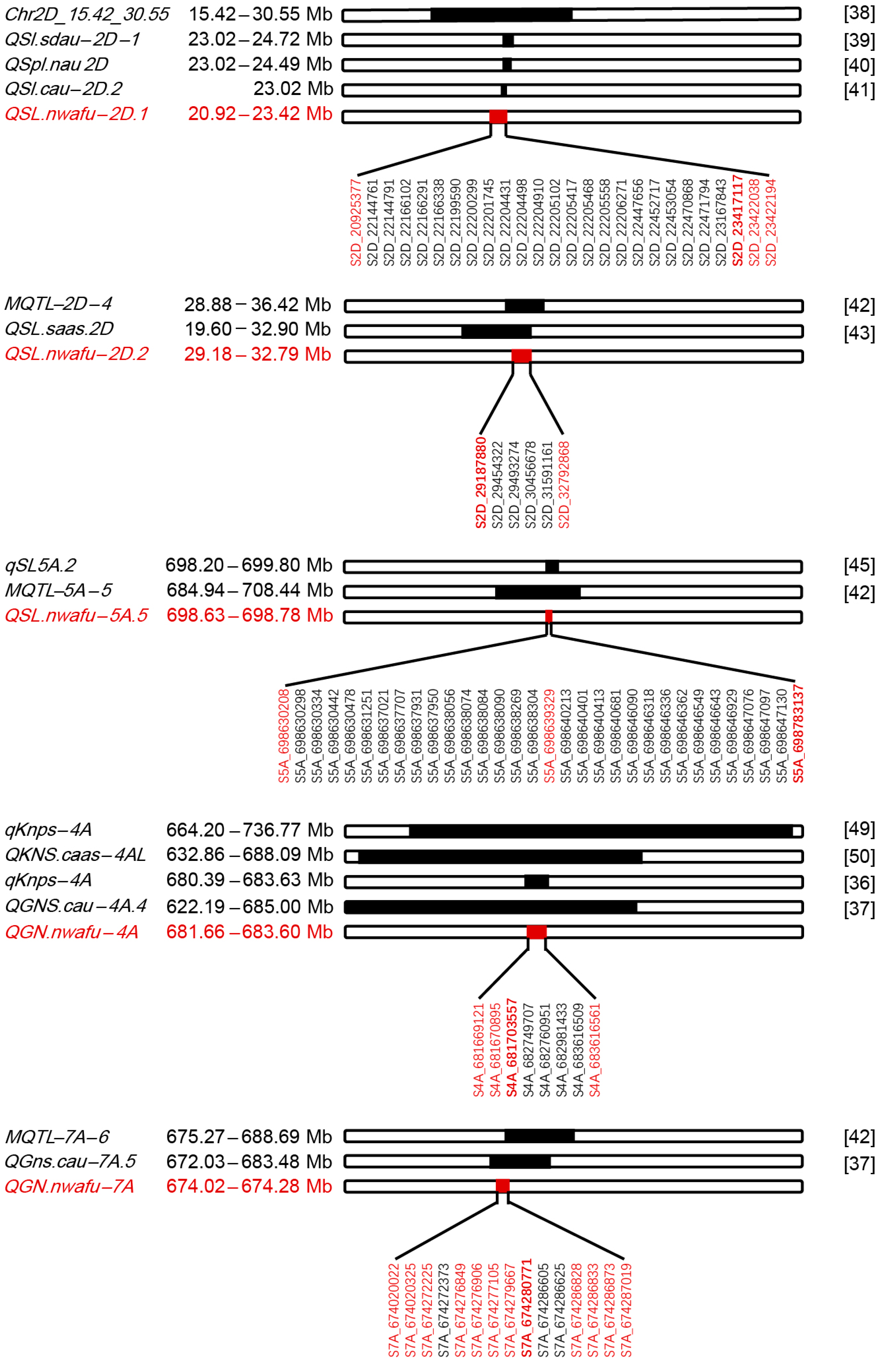
3.4. Haplotype Analysis of Reported QTLs of GN in Modern Wheat Breeding
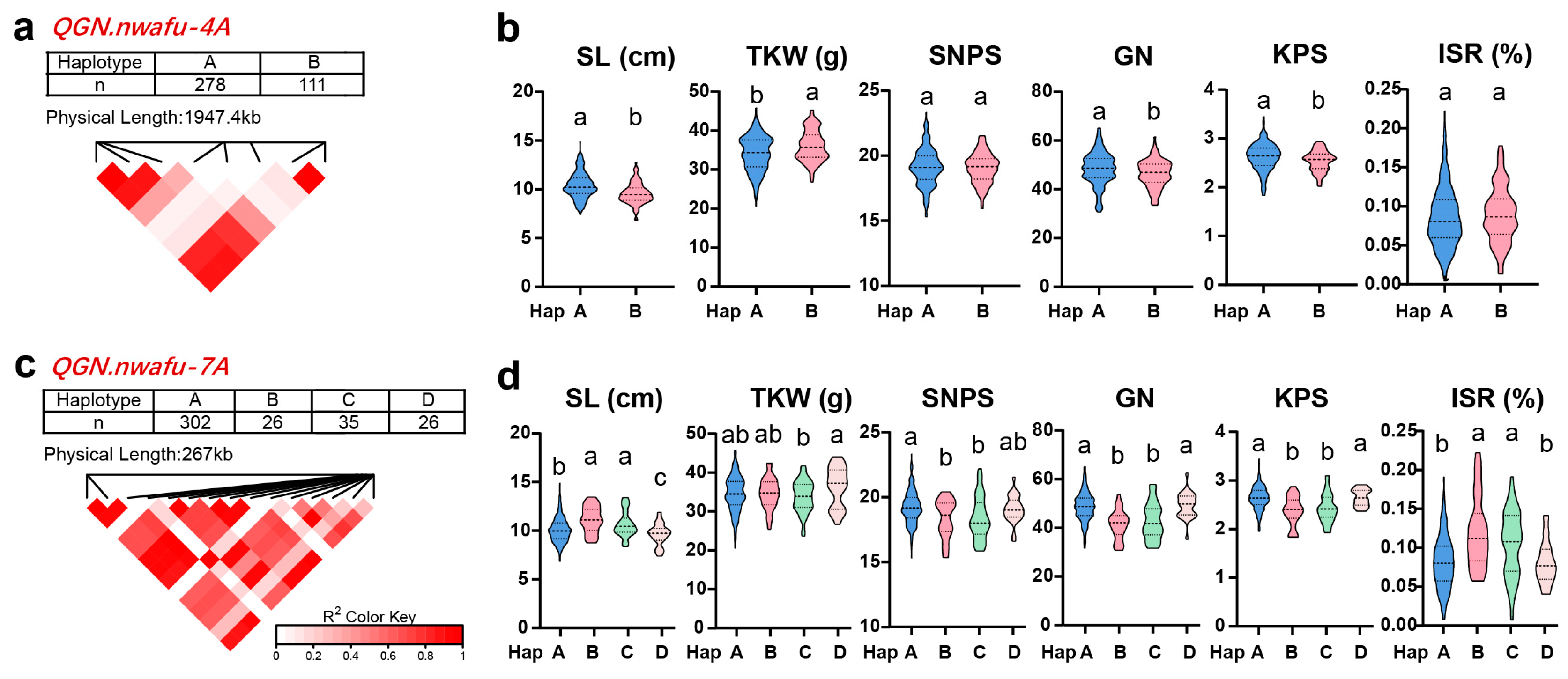
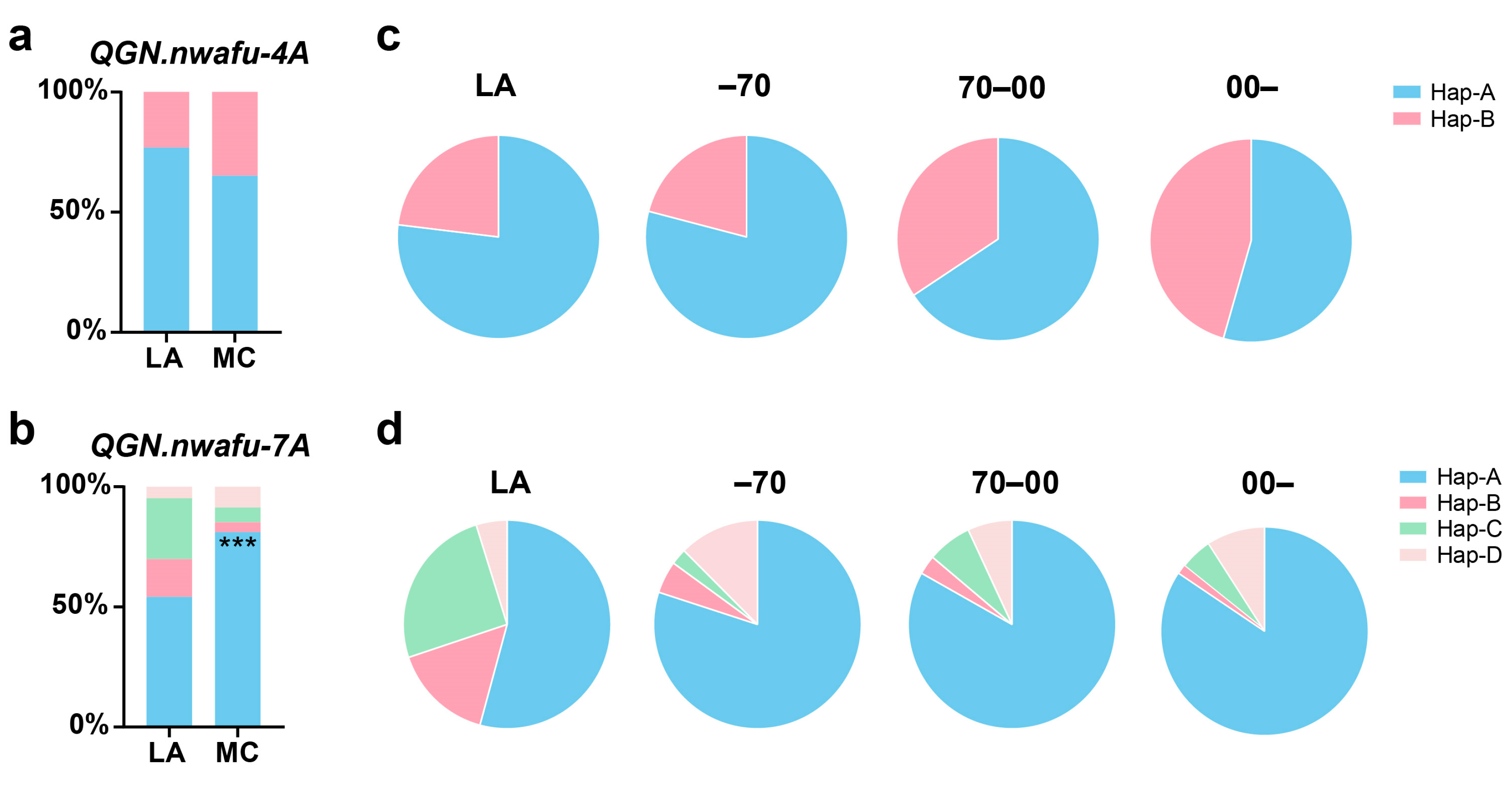
3.5. Selection and Application for QGN.nwafu-4A and QGN.nwafu-7A during Modern Breeding
3.6. Selection and Application for QGN.nwafu-4A and QGN.nwafu-7A in Different Chinese Wheat Regions
4. Discussion
4.1. The Contribution of Grain Number to Wheat Yield
4.2. Enrichment of Spike-Related QTLs Based on GWAS Assay
4.3. Repetitive and Novel QTLs of Spike-Related Traits Compared with Previous Research
4.4. Selection and Application of GN-Related QTLs in Wheat Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tadesse, W.; Sanchez-Garcia, M.; Assefa, S.G.; Amri, A.; Bishaw, Z.; Ogbonnaya, F.C.; Baum, M. Genetic Gains in Wheat Breeding and Its Role in Feeding the World. Crop Breed. Genet. Genom 2019, 1, e190005. [Google Scholar]
- Cao, S.; Xu, D.; Hanif, M.; Xia, X.; He, Z. Genetic architecture underpinning yield component traits in wheat. Theor. Appl. Genet. 2020, 133, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Doyle, D.; McAuley, S.; McHardy, F.; Danneels, Q.; Black, L.C.; White, E.M.; Spink, J. The impact of variation in grain number and individual grain weight on winter wheat yield in the high yield potential environment of Ireland. Eur. J. Agron. 2017, 87, 40–49. [Google Scholar] [CrossRef]
- Feng, F.; Han, Y.; Wang, S.; Yin, S.; Peng, Z.; Zhou, M.; Gao, W.; Wen, X.; Qin, X.; Siddique, K.H.M. The Effect of Grain Position on Genetic Improvement of Grain Number and Thousand Grain Weight in Winter Wheat in North China. Front. Plant Sci. 2018, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Schnurbusch, T. A genetic playground for enhancing grain number in cereals. Trends Plant Sci. 2012, 17, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Hanif, M.; Langer, R.H.M. The Vascular System of the Spikelet in Wheat (Triticum aestivum L.). Ann. Bot. 1972, 36, 721–727. [Google Scholar] [CrossRef]
- Wang, Y.; Du, F.; Wang, J.; Wang, K.; Tian, C.; Qi, X.; Lu, F.; Liu, X.; Ye, X.; Jiao, Y. Improving Bread Wheat Yield through Modulating an Unselected AP2/ERF Gene. Nat. Plants. 2022, 8, 930–939. [Google Scholar] [CrossRef]
- Sakuma, S.; Schnurbusch, T. Of floral fortune: Tinkering with the grain yield potential of cereal crops. New Phytol. 2020, 225, 1873–1882. [Google Scholar] [CrossRef]
- Kole, C.; Muthamilarasan, M.; Henry, R.; Edwards, D.; Sharma, R.; Abberton, M.; Batley, J.; Bentley, A.; Blakeney, M.; Bryant, J.; et al. Application of Genomics-Assisted Breeding for Generation of Climate Resilient Crops: Progress and Pro-spects. Front. Plant Sci. 2015, 6, 563. [Google Scholar] [CrossRef]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef]
- Henry, R.J.; Furtado, A.; Rangan, P. Wheat seed transcriptome reveals genes controlling key traits for human preference and crop adaptation. Curr. Opin. Plant Biol. 2018, 45, 231–236. [Google Scholar] [CrossRef]
- Segura, V.; Vilhjálmsson, B.J.; Platt, A.; Korte, A.; Seren, Ü.; Long, Q.; Nordborg, M. An Efficient Multi-Locus Mixed-Model Approach for Genome-Wide Association Studies in Structured Populations. Nat. Genet. 2012, 44, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, D.; Guo, X.; Yang, W.; Sun, J.; Wang, D.; Zhang, A. Genomic Distribution of Quantitative Trait Loci for Yield and Yield-Related Traits in Common Wheat. J. Integr. Plant Biol. 2010, 52, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, S.; Golan, G.; Guo, Z.; Ogawa, T.; Tagiri, A.; Sugimoto, K.; Bernhardt, N.; Brassac, J.; Mascher, M.; Hensel, G.; et al. Unleashing floret fertility in wheat through the mutation of a homeobox gene. Proc. Natl. Acad. Sci. USA 2019, 116, 5182–5187. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.E.; Greenwood, J.R.; Bencivenga, S.; Zhang, P.; Cockram, J.; Mellers, G.; Ramm, K.; Cavanagh, C.; Swain, S.M.; Boden, S.A. TEOSINTE BRANCHED1 Regulates Inflorescence Architecture and Development in Bread Wheat (Triticum aestivum L.). Plant Cell 2018, 30, 563–581. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, H.; Li, T.; Wu, J.; Nagarajan, R.; Lei, L.; Powers, C.; Kan, C.C.; Hua, W.; Liu, Z.; et al. TaCol-B5 Modifies Spike Architecture and Enhances Grain Yield in Wheat. Science 2022, 376, 180–183. [Google Scholar] [CrossRef]
- Dobrovolskaya, O.; Pont, C.; Sibout, R.; Martinek, P.; Badaeva, E.; Murat, F.; Chosson, A.; Watanabe, N.; Prat, E.; Gautier, N.; et al. FRIZZY PANICLE Drives Supernumerary Spikelets in Bread Wheat. Plant Physiol. 2015, 167, 189–199. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Zhao, M.; Guo, L.; Guo, X.; Zhao, D.; Batool, A.; Dong, B.; Xu, H.; Cui, S.; et al. Wheat FRIZZY PANICLE Activates VERNALIZATION1-A and HOMEOBOX4-A to Regulate Spike Development in Wheat. Plant Biotechnol. J. 2021, 19, 1141–1154. [Google Scholar] [CrossRef]
- Zhang, J.; Xiong, H.; Guo, H.; Li, Y.; Xie, X.; Xie, Y.; Zhao, L.; Gu, J.; Zhao, S.; Ding, Y.; et al. Identification of the Q Gene Playing a Role in Spike Morphology Variation in Wheat Mutants and Its Regulatory Network. Front. Plant Sci. 2021, 12, 807731. [Google Scholar] [CrossRef]
- Okada, T.; Jayasinghe, J.E.A.R.M.; Eckermann, P.; Watson-Haigh, N.S.; Warner, P.; Hendrikse, Y.; Baes, M.; Tucker, E.J.; Laga, H.; Kato, K.; et al. Effects of Rht-B1 and Ppd-D1 loci on pollinator traits in wheat. Theor. Appl. Genet. 2019, 132, 1965–1979. [Google Scholar] [CrossRef]
- Achilli, A.L.; Roncallo, P.F.; Larsen, A.O.; Dreisigacker, S.; Echenique, V. Population structure, allelic variation at Rht-B1 and Ppd-A1 loci and its effects on agronomic traits in Argentinian durum wheat. Sci. Rep. 2022, 12, 9629. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.M.; Zhou, R.H.; Gao, L.F.; Zhao, G.Y.; Jia, J.Z. The Characterization and Geographical Distribution of the Genes Responsible for Vernalization Requirement in Chinese Bread Wheat. J. Integr. Plant Biol. 2009, 51, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhou, Y.; Hu, W.; Zhang, Y.; Zhou, Y.; Chen, Y.; Wang, X.; Zhao, H.; Cao, T.; Liu, Z. Unlocking the Rela-tionships among Population Structure, Plant Architecture, Growing Season, and Environmental Adaptation in Henan Wheat Cultivars. BMC Plant Biol. 2020, 20, 469. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Smith, S.E.; Kuehl, R.O.; Ray, I.M.; Hui, R.; Soleri, D. Evaluation of Simple Methods for Estimating Broad-Sense Heritability in Stands of Randomly Planted Genotypes. Crop Sci. 1998, 38, 1125–1129. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, P.; Lai, X.; Wang, X.; Ji, W.; Xu, S. The Selection and Application of Peduncle Length QTL Qpl_6d.1 in Modern Wheat (Triticum aestivum L.) Breeding. Theor. Appl. Genet. 2023, 136, 32. [Google Scholar] [CrossRef]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef]
- Tian, F.; Bradbury, P.; Brown, P.; Sun, Q.; Flint-Garcia, S.; Rocheford, T.; McMullen, M.; Holland, J.; Buckler, E. Genome-Wide Association Study of Leaf Architecture in the Maize Nested Association Mapping Population. Nat. Genet. 2011, 43, 159–162. [Google Scholar] [CrossRef]
- Hirsch, C.N.; A Flint-Garcia, S.; Beissinger, T.M.; Eichten, S.R.; Deshpande, S.; Barry, K.; McMullen, M.D.; Holland, J.B.; Buckler, E.S.; Springer, N.; et al. Insights into the Effects of Long-Term Artificial Selection on Seed Size in Maize. Genetics 2014, 198, 409–421. [Google Scholar]
- Wang, X.; Zhao, P.; Guo, X.; Liu, Z.; Ma, X.; Zhao, Y.; Lai, X.; Huang, L.; Wang, W.; Han, D. Population Sequencing Reveals Rht-D1b Contributing the Bigger Seedling Root to Modern Wheat Cultivars. bioRxiv 2022.
- Liu, Z.; Lai, X.; Chen, Y.; Zhao, P.; Wang, X.; Ji, W.; Xu, S. Selection and application of four QTLs of grain protein content in modern wheat cultivars. J. Integr. Agric. 2023. [CrossRef]
- Zhang, C.; Dong, S.-S.; Xu, J.-Y.; He, W.-M.; Yang, T.-L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Valdar, W.; Holmes, C.C.; Mott, R.; Flint, J. Mapping in Structured Populations by Resample Model Averaging. Genetics 2009, 182, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Zhang, N.; Fan, X.; Zhang, W.; Zhao, C.; Yang, L.; Pan, R.; Chen, M.; Han, J.; Zhao, X. Utilization of a Wheat660k Snp Array-Derived High-Density Genetic Map for High-Resolution Mapping of a Major QTL for Kernel Number. Sci. Rep. 2017, 7, 3788. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Lu, L.; Jia, L.; Kabir, M.R.; Zhang, J.; Lan, T.; Zhao, Y.; Xin, M.; Hu, Z.; Yao, Y.; et al. Global QTL Analysis Identifies Genomic Regions on Chromosomes 4A and 4B Harboring Stable Loci for Yield-Related Traits Across Different Environments in Wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Deng, Z.; Kong, X.; Wang, F.; Guan, J.; Cui, D.; Sun, G.; Liao, R.; Fu, M.; Che, Y.; et al. Indels Identification and Association Analysis with Spike and Awn Length in Chinese Wheat Mini-Core Collection. Int. J. Mol. Sci. 2022, 23, 5587. [Google Scholar] [CrossRef]
- Deng, Z.; Cui, Y.; Han, Q.; Fang, W.; Li, J.; Tian, J. Discovery of Consistent QTLs of Wheat Spike-Related Traits under Nitrogen Treatment at Different Development Stages. Front. Plant Sci. 2017, 8, 2120. [Google Scholar] [CrossRef]
- Wu, X.; Cheng, R.; Xue, S.; Kong, Z.; Ma, Z. Precise Mapping of a Quantitative Trait Locus Interval for Spike Length and Grain Weight in Bread Wheat (Triticum Aestivum L.). Mol. Breed. 2014, 33, 129–138. [Google Scholar] [CrossRef]
- Zhai, H.; Feng, Z.; Li, J.; Liu, X.; Xiao, S.; Ni, Z.; Sun, Q. QTL Analysis of Spike Morphological Traits and Plant Height in Winter Wheat (Triticum Aestivum L.) Using a High-Density Snp and Ssr-Based Linkage Map. Front. Plant Sci. 2016, 7, 1617. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Amo, A.; Wei, D.; Chai, Y.; Zheng, J.; Qiao, P.; Cui, C.; Lu, S.; Chen, L.; Hu, Y. Large-scale integration of meta-QTL and genome-wide association study discovers the genomic regions and candidate genes for yield and yield-related traits in bread wheat. Theor. Appl. Genet. 2021, 134, 3083–3109. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, J.; Zhang, R.; Song, G.; Zhang, S.; Li, W.; Li, G. Identification of New QTL for Yield-Related Traits in Chinese Landrace and Elite Wheat Varieties through a Genome-Wide Linkage Mapping. Euphytica 2020, 216, 1–14. [Google Scholar] [CrossRef]
- Chen, D.; Wu, X.; Wu, K.; Zhang, J.; Liu, W.; Yang, X.; Li, X.; Lu, Y.; Li, L. Novel and Favorable Genomic Regions for Spike Related Traits in a Wheat Germplasm Pubing 3504 with High Grain Number Per Spike under Varying Environments. J. Integr. Agric. 2017, 11, 34–49. [Google Scholar] [CrossRef]
- Pang, Y.; Liu, C.; Wang, D.; Amand, P.S.; Bernardo, A.; Li, W.; He, F.; Li, L.; Wang, L.; Yuan, X.; et al. High-Resolution Ge-nome-Wide Association Study Identifies Genomic Regions and Candidate Genes for Important Agronomic Traits in Wheat. Mol. Plant 2020, 13, 1311–1327. [Google Scholar] [CrossRef]
- Li, F.; Wen, W.; He, Z.; Liu, J.; Jin, H.; Cao, S.; Geng, H.; Yan, J.; Zhang, P.; Wan, Y.; et al. Genome-wide linkage mapping of yield-related traits in three Chinese bread wheat populations using high-density SNP markers. Theor. Appl. Genet. 2018, 131, 1903–1924. [Google Scholar] [CrossRef]
- Zhou, Y.; Conway, B.; Miller, D.; Marshall, D.; Cooper, A.; Murphy, P.; Chao, S.; Brown-Guedira, G.; Costa, J. Quantitative Trait Loci Mapping for Spike Characteristics in Hexaploid Wheat. Plant Genome 2017, 10, 1–15. [Google Scholar] [CrossRef]
- Jantasuriyarat, C.; Vales, M.I.; Watson, C.J.W.; Riera-Lizarazu, O. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2003, 108, 261–273. [Google Scholar] [CrossRef]
- Cui, F.; Zhao, C.; Ding, A.; Li, J.; Wang, L.; Li, X.; Bao, Y.; Li, J.; Wang, H. Construction of an integrative linkage map and QTL mapping of grain yield-related traits using three related wheat RIL populations. Theor. Appl. Genet. 2014, 127, 659–675. [Google Scholar] [CrossRef]
- Gao, F.; Wen, W.; Liu, J.; Rasheed, A.; Yin, G.; Xia, X.; Wu, X.; He, Z. Genome-Wide Linkage Mapping of QTL for Yield Components, Plant Height and Yield-Related Physiological Traits in the Chinese Wheat Cross Zhou 8425B/Chinese Spring. Front. Plant Sci. 2015, 6, 1099. [Google Scholar] [CrossRef]
- Luo, X.; Yang, Y.; Lin, X.; Xiao, J. Deciphering spike architecture formation towards yield improvement in wheat. J. Genet. Genom. 2023, in press. [CrossRef] [PubMed]
- Sheoran, S.; Jaiswal, S.; Raghav, N.; Sharma, R.; Sabhyata; Gaur, A.; Jaisri, J.; Tandon, G.; Singh, S.; Sharma, P.; et al. Genome-Wide Association Study and Post-genome-Wide Association Study Analysis for Spike Fertility and Yield Related Traits in Bread Wheat. Front. Plant Sci. 2021, 12, 820761. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wen, W.; Liu, J.; Zhang, Y.; Cao, S.; He, Z.; Rasheed, A.; Jin, H.; Zhang, C.; Yan, J.; et al. Genetic architecture of grain yield in bread wheat based on genome-wide association studies. BMC Plant Biol. 2019, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Pretini, N.; Vanzetti, L.S.; Terrile, I.I.; Donaire, G.; González, F.G. Mapping QTL for spike fertility and related traits in two doubled haploid wheat (Triticum aestivum L.) populations. BMC Plant Biol. 2021, 21, 353. [Google Scholar] [CrossRef]
- Breseghello, F.; Sorrells, M.E. QTL analysis of kernel size and shape in two hexaploid wheat mapping populations. Field Crop. Res. 2007, 101, 172–179. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Singh, S.; Balyan, H.S.; Gupta, P.K. Multi-Locus Genome Wide Association Mapping for Yield and Its Contributing Traits in Hexaploid Wheat under Different Water Regimes. Sci. Rep. 2019, 9, 19468. [Google Scholar] [CrossRef]
- Saini, D.K.; Chopra, Y.; Singh, J.; Sandhu, K.S.; Kumar, A.; Bazzer, S.; Srivastava, P. Comprehensive evaluation of mapping complex traits in wheat using genome-wide association studies. Mol. Breed. 2022, 42, 1. [Google Scholar] [CrossRef]
- Marigorta, U.M.; Rodríguez, J.A.; Gibson, G.; Navarro, A. Replicability and Prediction: Lessons and Challenges from GWAS. Trends Genet. 2018, 34, 504–517. [Google Scholar] [CrossRef]
- Quarrie, S.A.; Steed, A.; Calestani, C.; Semikhodskii, A.; Lebreton, C.; Chinoy, C.; Steele, N.; Pljevljakusić, D.; Waterman, E.; Weyen, J.; et al. A High-Density Genetic Map of Hexaploid Wheat (Triticum aestivum L.) from the Cross Chinese Spring X Sq1 and Its Use to Compare QTLs for Grain Yield across a Range of Environments. Theor. Appl. Genet. 2005, 110, 865–880. [Google Scholar] [CrossRef]
- Cuthbert, J.L.; Somers, D.J.; Brûlé-Babel, A.L.; Brown, P.D.; Crow, G.H. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 117, 595–608. [Google Scholar] [CrossRef]
- Jia, H.; Wan, H.; Yang, S.; Zhang, Z.; Kong, Z.; Xue, S.; Zhang, L.; Ma, Z. Genetic dissection of yield-related traits in a recombinant inbred line population created using a key breeding parent in China’s wheat breeding. Theor. Appl. Genet. 2013, 126, 2123–2139. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Jia, L.; Lu, L.; Qin, D.; Zhang, J.; Guan, P.; Ni, Z.; Yao, Y.; Sun, Q.; Peng, H. Mapping QTLs of yield-related traits using RIL population derived from common wheat and Tibetan semi-wild wheat. Theor. Appl. Genet. 2014, 127, 2415–2432. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Hao, C.; Zhang, Y.; Cheng, J.; Zhang, Z.; Liu, J.; Yi, X.; Cheng, X.; Sun, D.; Xu, Y.; et al. A Combined Association Mapping and Linkage Analysis of Kernel Number Per Spike in Common Wheat (Triticum aestivum L.). Front. Plant Sci. 2017, 8, 1412. [Google Scholar] [CrossRef] [PubMed]
- Kuzay, S.; Xu, Y.; Zhang, J.; Katz, A.; Pearce, S.; Su, Z.; Fraser, M.; Anderson, J.A.; Brown-Guedira, G.; DeWitt, N.; et al. Identification of a Candidate Gene for a QTL for Spikelet Number Per Spike on Wheat Chromosome Arm 7AL by High-Resolution Genetic Mapping. Theor. Appl. Genet. 2019, 132, 2689–2705. [Google Scholar] [CrossRef]
- Guan, P.; Shen, X.; Mu, Q.; Wang, Y.; Wang, X.; Chen, Y.; Zhao, Y.; Chen, X.; Zhao, A.; Mao, W.; et al. Dissection and Validation of a QTL Cluster Linked to Rht-B1 Locus Controlling Grain Weight in Common Wheat (Triticum aestivum L.) Using near-Isogenic Lines. Theor. Appl. Genet. 2020, 133, 2639–2653. [Google Scholar] [CrossRef]
- Hu, J.; Wang, X.; Zhang, G.; Jiang, P.; Chen, W.; Hao, Y.; Ma, X.; Xu, S.; Jia, J.; Kong, L.; et al. QTL mapping for yield-related traits in wheat based on four RIL populations. Theor. Appl. Genet. 2020, 133, 917–933. [Google Scholar] [CrossRef]
- Isham, K.; Wang, R.; Zhao, W.; Wheeler, J.; Klassen, N.; Akhunov, E.; Chen, J. QTL mapping for grain yield and three yield components in a population derived from two high-yielding spring wheat cultivars. Theor. Appl. Genet. 2021, 134, 2079–2095. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, F.; Yan, X.; Zhang, X.; Dong, Z.; Cui, D.; Chen, F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotechnol. J. 2017, 15, 953–969. [Google Scholar] [CrossRef]
- Jiang, Y.; Jiang, Q.; Hao, C.; Hou, J.; Wang, L.; Zhang, H.; Zhang, S.; Chen, X.; Zhang, X. A yield-associated gene TaCWI, in wheat: Its function, selection and evolution in global breeding revealed by haplotype analysis. Theor. Appl. Genet. 2014, 128, 131–143. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | Min | Max | CV (%) | h2 |
|---|---|---|---|---|---|
| SL (cm) | 10.06 | 6.00 | 16.56 | 16.95 | 0.92 |
| TKW (g) | 35.09 | 12.33 | 71.71 | 21.37 | 0.82 |
| SNPS | 19.39 | 12.33 | 25.80 | 12.03 | 0.81 |
| GN | 48.72 | 21.00 | 78.60 | 20.33 | 0.78 |
| KPS | 2.53 | 1.14 | 3.81 | 18.74 | 0.72 |
| ISR | 0.12 | 0.00 | 0.59 | 43.95 | 0.46 |
| Traits | QTLs | pSNP | Chr | Pos (Mb) | Env | PVE (%) | RMIP | QTLs Detected in Previous Studies | |
|---|---|---|---|---|---|---|---|---|---|
| Traits | Bibliography | ||||||||
| SL | QSL.nwafu-2D.1 | S2D_23417117 | 2D | 20.92–23.42 | 0/1/2/3/4/5/6/7 | 7.59–11.32 | 5–44 | SL | [38,39,40,41] |
| QSL.nwafu-2D.2 | S2D_29187880 | 2D | 29.18–32.79 | /0/4/5 6/7 | 9.66 | 17–38 | SL | [42,43] | |
| QSL.nwafu-4A.1 | S4A_681703557 | 4A | 681.70–683.61 | 0/3/5 | 4.69–5.99 | 5–46 | SL | [44] | |
| QSL.nwafu-5A.2 | S5A_586308393 | 5A | 586.3 | 0/1/3 /5 | 4.48–6.08 | 6–12 | SL | [42] | |
| QSL.nwafu-5A.5 * | S5A_698783137 | 5A | 698.63–698.78 | 6/7 | 4.21–5.08 | 5–39 | SL | [45] | |
| GN, TKW | [42] | ||||||||
| TKW | QTKW.nwafu-3A | S3A_33423910 | 3A | 33.42–35.39 | 0/4/5 | 5.07–6.41 | 5–13 | TKW | [46] |
| QTKW.nwafu-4B.1 | S4B_40752468 | 4B | 36.15–40.75 | 0/4/7 | 4.15–6.64 | 5–8 | GN | [45] | |
| QTKW.nwafu-4B.2 | S4B_44185162 | 4B | 44.18 | 0/3/7 | 4.28–5.36 | 6–18 | GN | [45] | |
| QTKW.nwafu-7D | S7D_55412560 | 7D | 55.41 | 0/4/5 | 3.11–4.57 | 5–12 | TKW | [2] | |
| SNPS | QSNPS.nwafu-2D | S2D_628937584 | 2D | 628.93 | 0/4/6 | 4.34–5.32 | 5–30 | FS | [47] |
| QSNPS.nwafu-4A | S4A_597024251 | 4A | 597.02 | 0/6 | 5.83–7.55 | 6 | SNPS | [48] | |
| GN | QGN.nwafu-2A | S2A_715307274 | 2A | 715.30–715.33 | 0/2/4 | 4.56–5.38 | 5–20 | TKW | [37] |
| QGN.nwafu-4A * | S4A_681703557 | 4A | 681.7 | 0/6 | 4.69–7.88 | 7–10 | GN | [36,37,49,50] | |
| QGN.nwafu-7A | S7A_674280771 | 7A | 674.02–674.28 | 0/2/4 | 5.28–8.68 | 5–31 | GN | [37,42] | |
| KPS | QKPS.nwafu-4A * | S4A_681703557 | 4A | 681.66–681.70 | 0/6 | 7.56–7.88 | 5–9 | GN | [36,49,50] |
| QKPS.nwafu-4B.1 | S4B_27522453 | 4B | 27.52–30.86 | 0/2/6 7 | 8.07–11.49 | 5–9 | GN, KPS | [21] | |
| QKPS.nwafu-4B.2 * | S4B_41013810 | 4B | 36.15–41.01 | 0/2 | 5.58–7.39 | 5–19 | GN | [45] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Chen, Y.; Lai, X.; Liu, Z.; Liu, M.; Xu, S. Genome-Wide Association Study for Spike Traits and Distribution of Two QTLs for Grain Number in Chinese Wheat Cultivars. Agronomy 2023, 13, 2538. https://doi.org/10.3390/agronomy13102538
Wang J, Chen Y, Lai X, Liu Z, Liu M, Xu S. Genome-Wide Association Study for Spike Traits and Distribution of Two QTLs for Grain Number in Chinese Wheat Cultivars. Agronomy. 2023; 13(10):2538. https://doi.org/10.3390/agronomy13102538
Chicago/Turabian StyleWang, Junzhe, Yijin Chen, Xiangjun Lai, Zihui Liu, Manshuang Liu, and Shengbao Xu. 2023. "Genome-Wide Association Study for Spike Traits and Distribution of Two QTLs for Grain Number in Chinese Wheat Cultivars" Agronomy 13, no. 10: 2538. https://doi.org/10.3390/agronomy13102538
APA StyleWang, J., Chen, Y., Lai, X., Liu, Z., Liu, M., & Xu, S. (2023). Genome-Wide Association Study for Spike Traits and Distribution of Two QTLs for Grain Number in Chinese Wheat Cultivars. Agronomy, 13(10), 2538. https://doi.org/10.3390/agronomy13102538