

Quantitative Trait Analysis Shows the Potential for Alleles from the Wild Species Arachis batizocoi and A. duranensis to Improve Groundnut Disease Resistance and Yield in East Africa

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Populations
2.2. Field Experiment
2.3. Phenotypic Screening for Late Leaf Spot (LLS) and Groundnut Rosette Disease (GRD)
2.4. Agronomic Evaluations
2.5. Data Analysis
2.6. SNP Genotyping, Analysis and Data Filtering
2.7. Genetic Mapping and QTL Discovery
3. Results
3.1. Agronomic Evaluation
3.1.1. Total Yield (g/m2)
3.1.2. 100-Pod Weight—Proxy for Seed Size
3.1.3. Earliness
3.2. Disease Evaluation
3.2.1. Late Leaf Spot Resistance
3.2.2. Groundnut Rosette Disease Resistance
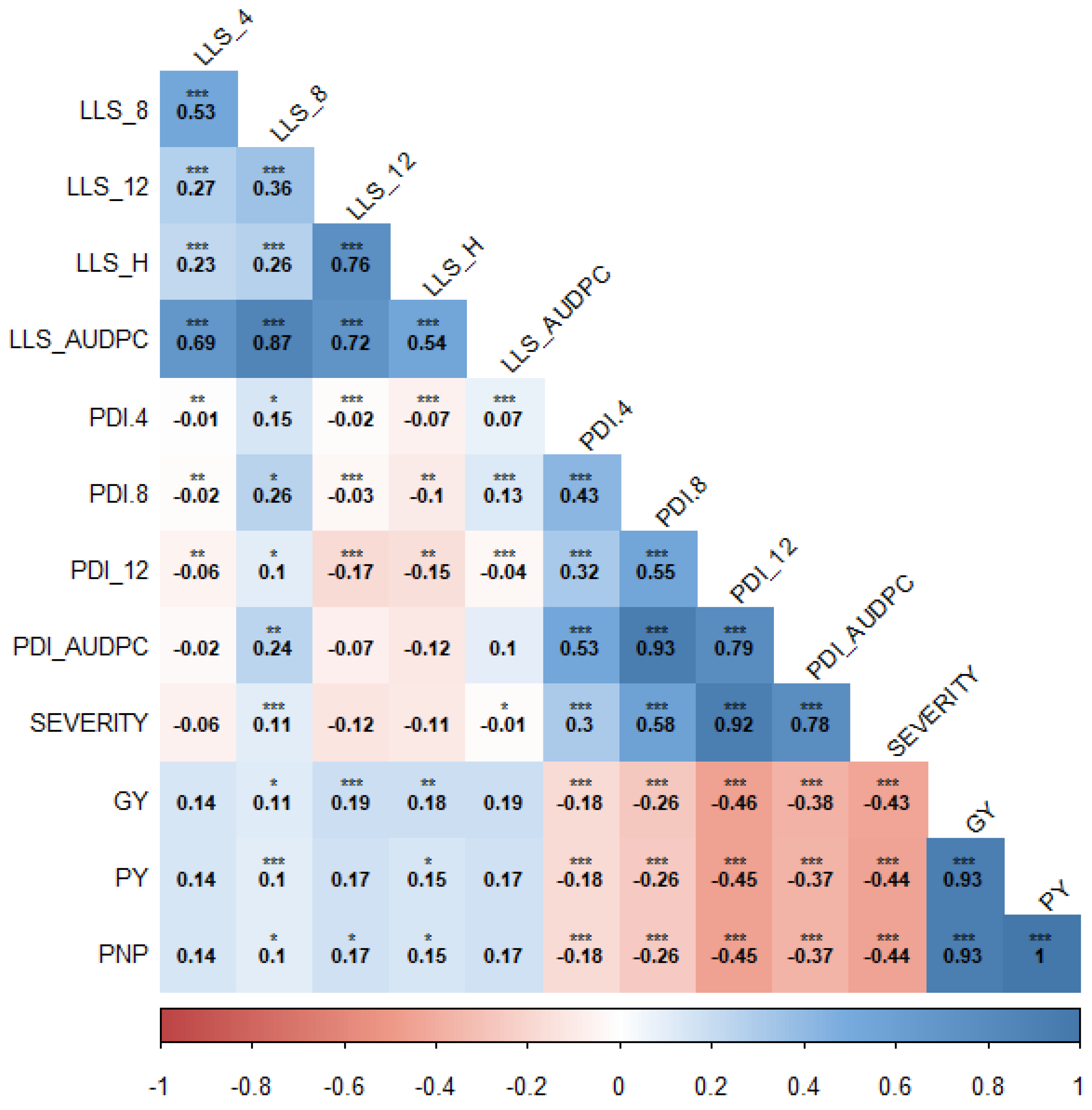
3.3. Analysis of Variance and Heritability for Groundnut Rosette and Late Leaf Spot Diseases across Two Locations
3.4. Identification of QTL for Disease Resistance
3.4.1. QTL for LLS Resistance
3.4.2. QTL for GRD Resistance
3.5. Identification of Markers Linked to Yield and 100-Seed Weight
4. Discussion
4.1. Variations per Location and Season
4.2. Phenotypic Evaluation and Selection of Superior Lines
4.3. QTL Identification
4.4. Usefulness of Lines to the Groundnut Breeding Program
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.C.M.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef]
- Kochert, G.; Stalker, H.T.; Gimenes, M.; Galgaro, L.; Lopes, C.R.; Moore, K. RFLP and cytogenetic evidence on the origin and evolution of allotetraploid domesticated peanut, Arachis hypogaea (Leguminosae). Am. J. Bot. 1996, 83, 1282–1291. [Google Scholar] [CrossRef]
- Chintu, J.M.M. Breeding Groundnut for Resistance to Rosette Disease and Its Aphid Vector, Aphis craccivora Koch in Malawi. Ph.D. Thesis, University of KwaZulu-Natal, Durban, South Africa, 2013. [Google Scholar]
- Stalker, H.T. Utilizing wild species for peanut improvement. Crop Sci. 2017, 57, 1102–1120. [Google Scholar] [CrossRef]
- Naidu, R.A.; Kimmins, F.M.; Deom, C.; Subrahmanyam, P.; Chiyembekeza, A.J.; van der Merwe, P.J.A. Groundnut rosette: A virus disease affecting groundnut production in sub-Saharan africa. Plant Dis. 1999, 83, 700–709. [Google Scholar] [CrossRef]
- Okello, D.K.; Biruma, M.; Deom, C.M. Overview of groundnuts research in Uganda: Past, present and future. Afr. J. Biotechnol. 2010, 9, 6448–6459. [Google Scholar]
- Laker-Ojok, R. Returns to Oilseed and Maize Research in Uganda. In The Case of Groundnut and Sesame. MSU International Development Working Paper No. 45; University of Michigan, Department of Agronomical Economic: East Lansing, MI, USA, 1996. [Google Scholar]
- Subrahmanyam, P.; Hildebrand, G.L.; Naidu, R.A.; Reddy, L.J.; Singh, A.K. Sources of resistance to groundnut rosette disease in global groundnut germplasm. Ann. Appl. Biol. 1998, 132, 473–485. [Google Scholar] [CrossRef]
- Subrahmanyam, P.; McDonald, D.; Waliyar, F.; Reddy, L.J.; Nigam, S.N.; Gibbons, R.W.; Rao, V.R.; Singh, A.K.; Pande, S.; Reddy, P.M.; et al. Screening Methods and Sources of Resistance to Rust and Late Leaf Spot of Groundnut. In Technical Report Information Bulletin No 47; Tropics ICRIftS-A, Ed.; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Andhra Pradesh, India, 1995. [Google Scholar]
- Mohammed, K.E.; Afutu, E.; Odong, T.L.; Okello, D.K.; Nuwamanya, E.; Grigon, O.; Rubaihayo, P.R.; Okori, P. Assessment of groundnut (Arachis hypogaea L.) genotypes for yield and resistance to late leaf spot and rosette diseases. J. Exp. Agric. Int. 2018, 21, 1–13. [Google Scholar] [CrossRef]
- Waliyar, F.; Kumar, P.; Ntare, B.; Monyo, E.; Nigam, S.; Reddy, A.; Osiru, M.; Diallo, A. A Century of Research on Groundnut Rosette Disease and Its Management. In Information Bulletin; Tropics TRICRIftS-A, Ed.; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, Andhra Pradesh, India, 2007; Volume 75. [Google Scholar]
- Wangai, A.; Pappu, S.; Pappu, H.; Deom, C.; Naidu, R. Distribution and characteristics of groundnut rosette disease in Kenya. Plant Dis. 2001, 85, 470–474. [Google Scholar] [CrossRef]
- Okello, D.K.; Akello, L.B.; Odong, T.L. Groundnut rosette disease symptoms types distribution and management of the disease in Uganda. Afr. J. Plant Sci. 2014, 8, 153–163. [Google Scholar]
- Nutman, F.J.; Roberts, F.M.; Williamson, J.G. Studies of varietal resistance in the groundnut, (Arachis hypogaea L.) to rosette disease. Rhod. J. Agric. Res. 1964, 2, 63–77. [Google Scholar]
- Bock, K.; Murant, A.; Rajeshwari, R. The nature of the resistance in groundnut to rosette disease. Ann. Appl. Biol. 1990, 117, 379–384. [Google Scholar] [CrossRef]
- Olorunju, P.E.; Kuhn, C.W.; Demski, J.W.; Misari, S.M.; Ansa, O.A. Disease reactions and yield performance of peanut genotypes grown under groundnut rosette and rosette-free field environments. Plant Dis. 1991, 75, 1269–1273. [Google Scholar] [CrossRef]
- Deom, C.M.; Naidu, R.A.; Chiyembekeza, A.J.; Ntare, B.R.; Subrahmanyam, P. Sequence diversity within the three agents of groundnut rosette disease. Phytopathology 2000, 90, 214–219. [Google Scholar] [CrossRef]
- Deom, C.M.; Okello, D.K. Developing Improved Varieties of Groundnut. In Achieving Sustainable Cultivation of Grain Legumes; Sivasankar, S., Bergvinson, D., Gaur, P., Kumar, S., Beebe, S., Tamo, M., Eds.; Burleigh Dodds Scientific Publishing: Cambridge, UK, 2018; Volume 2. [Google Scholar]
- Subrahmanyam, P.; Naidu, R.A.; Reddy, L.J.; Kumar, P.L.; Ferguson, M.E. Resistance to groundnut rosette disease in wild Arachis species. Ann. Appl. Biol. 2001, 139, 45–50. [Google Scholar] [CrossRef]
- Pande, S.; Rao, J.N. Resistance of wild Arachis species to late leaf spot and rust in greenhouse trials. Plant Dis. 2001, 85, 851–855. [Google Scholar] [CrossRef]
- Stalker, H.T.; Tallury, S.; Ozias-Akins, P.; Bertioli, D.J.; Leal-Bertioli, S. The value of diploid peanut relatives for breeding and genomics. Peanut Sci. 2013, 40, 70–88. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Seijo, G.; Freitas, F.O.; Valls, J.F.; Leal-Bertioli, S.C.; Moretzsohn, M.C. An overview of peanut and its wild relatives. Plant Genet. Resour. 2011, 9, 134–149. [Google Scholar] [CrossRef]
- Fonceka, D.; Tossim, H.; Rivallan, R.; Vignes, H.; Lacut, E.; de Bellis, F.; Faye, I.; Ndoye, O.; Leal-Bertioli, S.; Valls, J.; et al. Construction of chromosome segment substitution lines in peanut (Arachis hypogaea L.) using a wild synthetic and QTL mapping for plant morphology. PLoS ONE 2012, 7, e48642. [Google Scholar] [CrossRef]
- Fonceka, D.; Tossim, H.A.; Rivallan, R.; Vignes, H.; Faye, I.; Ndoye, O. Fostered and left behind alleles in peanut: Interspecific QTL mapping reveals footprints of domestication and useful natural variation for breeding. BMC Plant Biol. 2012, 12, 26. [Google Scholar] [CrossRef]
- Moss, J.P.; Singh, A.K.; Reddy, L.J.; Nigam, S.N.; Subrahmanyam, P.; McDonald, D.; Reddy, A.G.S. Registration of ICGV 87165 Peanut Germplasm Line with Multiple Resistance. Crop Sci. 1998, 37, 1028. [Google Scholar] [CrossRef]
- Okello, D.; Deom, C.; Puppala, N. Registration of ‘Naronut 2T’ groundnut. J. Plant Regist. 2021, 15, 62–67. [Google Scholar] [CrossRef]
- Van der Merwe, P.J.A.; Subrahmanyam, P.; Hildebrand, G.L.; Reddy, L.J.; Nigam, S.N.; Chiyembekeza, A.J.; Busolo-Bulafu, C.M.; Kapewa, T. Registration of groundnut cultivar ICGV-SM 90704 with resistance to groundnut rosette. Int. Arachis Newsl. 2001, 21, 19–20. [Google Scholar]
- Bertioli, D.J.; Clevenger, J.; Godoy, I.J.; Stalker, H.T.; Wood, S.; Santos, J.F.; Ballén-Taborda, C.; Abernathy, B.; Azevedo, V.; Campbell, J.; et al. Legacy genetics of Arachis cardenasii in the peanut crop shows the profound benefits of international seed exchange. Proc. Natl. Acad. Sci. USA 2021, 118, e2104899118. [Google Scholar] [CrossRef]
- Nguepjop, J.R.; Tossim, H.-A.; Bell, J.M.; Rami, J.-F.; Sharma, S.; Courtois, B.; Mallikarjuna, N.; Sane, D.; Fonceka, D. Evidence of genomic exchanges between homeologous chromosomes in a cross of peanut with newly synthetized allotetraploid hybrids. Front. Plant Sci. 2016, 7, 1635. [Google Scholar] [CrossRef]
- Tossim, H.-A.; Nguepjop, J.R.; Diatta, C.; Sambou, A.; Seye, M.; Sane, D.; Rami, J.-F.; Fonceka, D. Assessment of 16 peanut (Arachis hypogaea L.) CSSLs derived from an interspecific cross for yield and yield component traits: QTL validation. Agronomy 2020, 10, 583. [Google Scholar] [CrossRef]
- Faye, I.; Hodo-Abalo, T.; Ndoye, O.; Fonceka, D. Dossier Technique D’homologation de Nouvelles Variétés D′Arachide; ISRA, Ed.; Ministry of Agriculture: Dakar, Senegal, 2016; Volume 34. [Google Scholar]
- Mallikarjuna, N.; Senthilvel, S.; Hoisington, D. Development of new sources of tetraploid Arachis to broaden the genetic base of cultivated groundnut (Arachis hypogaea L.). Genet. Resour. Crop Evol. 2011, 58, 889–907. [Google Scholar] [CrossRef]
- Sambou, A. Genetic Studies and Mapping of Beneficial Alleles from Wild Species to Improve Agronomic Traits in Cultivated Groundnut (Arachis hypogaea L.); University of Legon: Accra, Ghana, 2017. [Google Scholar]
- Fávero, A.P.; Simpson, C.E.; Valls, F.M.J.; Velo, N.A. Study of evolution of cultivated peanut trough crossability studies among Arachis ipaënsis, A duranensis and A hypogaea. Crop Sci. 2006, 46, 1546–1552. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.M.; Godoy, I.J.; Santos, J.F.; Doyle, J.J.; Guimarães, P.M.; Abernathy, B.L.; Jackson, S.A.; Moretzsohn, M.C.; Bertioli, D.J. Segmental allopolyploidy in action: Increasing diversity through polyploid hybridization and homoeologous recombination. Am. J. Bot. 2018, 105, 1053–1066. [Google Scholar] [CrossRef]
- Fonceka, D.; Hodo-Abalo, T.; Rivallan, R.; Faye, I.; Sall, M.N.; Ndoye, O.; Favero, A.P.; Bertioli, D.J.; Glaszmann, J.C.; Courtois, B.; et al. Genetic mapping of wild introgressions into cultivated peanut: A way toward enlarging the genetic basis of a recent allotetraploid. BMC Plant Biol. 2009, 9, 103. [Google Scholar] [CrossRef]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.; Comstock, R. Estimates of genetic and environmental variability in soybeans 1. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Team, R.D.C. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Korani, W.; Clevenger, J.P.; Chu, Y.; Ozias-Akins, P. Machine learning as an effective method for identifying true single nucleotide polymorphisms in polyploid plants. Plant Genome 2019, 12, 180023. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Broman, K.W. Sen S; Springer: Berlin, Germany, 2009. [Google Scholar]
- Churchill, G.A.; Doerge, R.W. Empirical threshold values for quantitative trait mapping. Genetics 1994, 138, 963–971. [Google Scholar] [CrossRef]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.; Moretzsohn, M.C.; Santos, S.P.; Brasileiro, A.C.; Guimarães, P.M.; Bertioli, D.J.; Araujo, A.C. Phenotypic effects of allotetraploidization of wild Arachis and their implications for peanut domestication. Am. J. Bot. 2017, 104, 379–388. [Google Scholar] [CrossRef]
- Alyr, M.H.; Pallu, J.; Sambou, A.; Nguepjop, J.R.; Seye, M.; Tossim, H.A.; Djiboune, Y.R.; Sane, D.; Rami, J.F.; Fonceka, D. Fine-mapping of a wild genomic region involved in pod and seed size reduction on chromosome A07 in peanut (Arachis hypogaea L.). Genes 2020, 11, 1402. [Google Scholar] [CrossRef]
- Mugisa, I.O.; Karungi, J.; Akello, B.; Ochwo-Ssemakula, M.K.N.; Biruma, M.; Okello, D.K.; Otim, G. Assessing the effect of farmers’ practices on the severity of groundnut rosette virus disease in Uganda. Afr. J. Agric. Res. 2015, 10, 995–1003. [Google Scholar] [CrossRef]
- Gonzales, M.; Kemerait, R.; Bertioli, D.; Bertioli, S. Strong resistance to early and late leaf spot in peanut-compatible wild-derived induced allotetraploids. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Michelotto, M.D.; Barioni, W., Jr.; Resende, M.D.V.; Godoy, I.J.; Leonardecz, E.; Fávero, A.P. Identification of fungus resistant wild accessions and interspecific hybrids of the genus Arachis. PLoS ONE 2015, 10, e0128811. [Google Scholar] [CrossRef]
- Bua, B.; Opio, M. Variability in reactions of groundnuts varieties to groundnut rosette virus isolates from Uganda. Am. J. Exp. Agric. 2014, 4, 541–549. [Google Scholar] [CrossRef]
- Tshilenge-Lukanda, L.; Nkongolo, K.K.C.; Kalonji-Mbuyi, A.; Kizungu, R.V. Epidemiology of the groundnut (Arachis hypogaea L.) leaf spot disease: Genetic analysis and developmental cycles. Am. J. Plant Sci. 2012, 2012, 19476. [Google Scholar]
- Clevenger, J.; Chu, Y.; Chavarro, C.; Botton, S.; Culbreath, A.; Isleib, T.G.; Holbrook, C.C.; Ozias-Akins, P. Mapping late leaf spot resistance in peanut (Arachis hypogaea) using QTL-seq reveals markers for marker-assisted selection. Front. Plant Sci. 2018, 9, 83. [Google Scholar] [CrossRef]
- Khera, P.; Pandey, M.K.; Mallikarjuna, N.; Sriswathi, M.; Roorkiwal, M.; Janila, P.; Sharma, S.; Shilpa, K.; Sudini, H.; Guo, B. Genetic imprints of domestication for disease resistance, oil quality, and yield component traits in groundnut (Arachis hypogaea L.). Mol. Genet. Genom. 2019, 294, 365–378. [Google Scholar] [CrossRef]
- Kolekar, R.M.; Sujay, V.; Shirasawa, K.; Sukruth, M.; Khedikar, Y.P.; Gowda, M.V.C.; Pandey, M.K.; Varshney, R.K.; Bhat, R.S. QTL mapping for late leaf spot and rust resistance using an improved genetic map and extensive phenotypic data on a recombinant inbred line population in peanut (Arachis hypogaea L.). Euphytica 2016, 209, 147–156. [Google Scholar] [CrossRef]
- Pandey, M.K.; Khan, A.W.; Singh, V.K.; Vishwakarma, M.K.; Shasidhar, Y.; Kumar, V.; Garg, V.; Bhat, R.S.; Chitikineni, A.; Janila, P.; et al. QTL-seq approach identified genomic regions and diagnostic markers for rust and late leaf spot resistance in groundnut (Arachis hypogaea L.). Plant Biotechnol. J. 2017, 15, 927–941. [Google Scholar] [CrossRef]
- Han, S.; Yuan, M.; Clevenger, J.P.; Li, C.; Hagan, A.; Zhang, X.; Chen, C.; He, G. A SNP-based linkage map revealed QTLs for resistance to early and late leaf spot diseases in peanut (Arachis hypogaea L.). Front. Plant Sci. 2018, 9, 1012. [Google Scholar] [CrossRef]
- Liang, Y.; Baring, M.; Wang, S.; Septiningsih, E.M. Mapping QTLs for leafspot resistance in peanut using SNP-based next-generation sequencing markers. Plant Breed. Biotechnol. 2017, 5, 115–122. [Google Scholar] [CrossRef]
- Zhou, X.; Xia, Y.; Liao, J.; Liu, K.; Li, Q.; Dong, Y.; Ren, X.; Chen, Y.; Huang, L.; Liao, B.; et al. Quantitative trait locus analysis of late leaf spot resistance and plant-type-related traits in cultivated peanut (Arachis hypogaea L.) under multi-environments. PLoS ONE 2016, 11, e0166873. [Google Scholar] [CrossRef]
- Leal-Bertioli, S.C.; Jose, A.C.; Alves-Freitas, D.M.; Moretzsohn, M.C.; Guimaraes, P.M.; Nielen, S. Identification of candidate genome regions controlling disease resistance in Arachis. BMC Plant Biol. 2009, 9, 112. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean | Range | |||||||
|---|---|---|---|---|---|---|---|---|
| Trait | AB-QTL2 | AB-QTL1 | CSSL | Fleur11 | AB-QTL2 | AB-QTL1 | CSSL | p |
| PNP | 15.4 a | 15.9 a | 16.9 a | 21.3 a | 6.6–26.0 | 6.6–21.4 | 7.9–25.6 | 0.37 |
| HSW | 68.1 a | 65.41 b | 70.65 a | 67.1 ab | 38.2–95.7 | 38.7–88.5 | 51.9–92.0 | 0.00 |
| SY | 103.9 b | 85.3 ab | 117.3 a | 150.6 ab | 14.5–239.8 | 10.5–221.7 | 37.0–283.3 | 0.03 |
| DTF | 32.7 a | 32.8 a | 32.60 a | 31.5 a | 30.0–35 | 28.2–36.2 | 30.7–35.7 | 0.48 |
| Trait | AB-QTL1 | AB-QTL2 | CSSL | Dif | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | CV % | H2 | Mean ± SE | CV % | H2 | Mean ± SE | CV % | H2 | ||
| LLS@4WEEKS | 1.73 ± 0.02 | 14.61 | 0 | 1.72 ± 0.02 | 30.9 | 0.06 | 1.70 ± 0.03 | 33.5 | 0.32 | * |
| LLS@8WEEKS | 3.56 ± 0.02 | 11.05 | 0.14 | 3.48 ± 0.02 | 15.1 | 0.11 | 3.50 ± 0.03 | 14.3 | 0 | ** |
| LLS@12WEEKS | 5.94 ± 0.03 | 10.84 | 0.49 | 5.91 ± 0.02 | 11.2 | 0.56 | 5.98 ± 0.05 | 8.8 | 0.24 | ** |
| LLS@Harvest | 6.80 ± 0.03 | 10.41 | 0.39 | 6.82 ± 0.02 | 10.4 | 0.54 | 6.0.09 ± 0.05 | 8.7 | 0.32 | ** |
| LLS AUDPC | 221.80 ± 0.88 | 319.42 | 0.45 | 218.91 ± 0.72 | 10.9 | 0.43 | 220.18 ± 1.49 | 10.1 | 0.19 | ** |
| PDI@4WEEKS | 3.00 ± 0.29 | 82.56 | 1.14 × 10−13 | 2.94 ± 0.24 | 281.1 | 0 | 3.14 ± 0.50 | 200.0 | 0 | ns |
| PDI@8WEEKS | 25.21 ± 0.92 | 32.09 | 0.10 | 23.01 ± 0.70 | 60.1 | 0.39 | 26.45 ± 1.51 | 69.9 | 0.07 | ** |
| PDI@12WEEKS | 47.31 ± 1.22 | 26.28 | NC | 45.38 ± 0.99 | 36.8 | 0.40 | 46.17 ± 2.01 | 23.5 | 0.21 | ** |
| GRD Severity | 2.55 ± 0.06 | 47.12 | 0.04 | 2.44 ± 0.05 | 30.7 | 0.48 | 2.48 ± 0.10 | 18.9 | ** | |
| GRD AUDPC | 151.10 ± 44.43 | 47.66 | NC | 141.30 ± 34.70 | 49.2 | 0.43 | 153 ± 74.48 | 45.6 | 0.11 | ** |
| Trait Name | Trait Description | Nearest Marker | LG | Physical Position (Mb) | LOD | p-Val | LOD Threshold (5%) | 99% Bayes Interval (Mb) | R2 (%) |
|---|---|---|---|---|---|---|---|---|---|
| LLS_12_WEEKS_NAK_20 | LLS@12 wk Nakabango 2020 | B04_17,705,669 | B04 | 17.70 | 3.04 | 0.04 | 2.79 | 7.08–90.02 | 9.43 |
| LLS_AUDPC_NAK_20 | LLS AUDPC Nakabango 2020 | B04_9,708,846 | B04 | 9.71 | 3.00 | 0.03 | 2.84 | 7.08–90.02 | 9.60 |
| GRD_PDI_SEV_SER_20 | GRD Severity Serere 2020 | A08_22,758,270 | A08 | 22.80 | 3.20 | 0.02 | 2.78 | 22.76–23.42 | 7.90 |
| GRD_PDI_AUDPC_NAK_20 | GRD AUDPC Nakabango 2020 | B01_62,384,869 | B01 | 62.40 | 2.81 | 0.05 | 2.77 | 7.45–121.37 | 11.04 |
| GRD_PDI_8_WEEKS_NAK_20 | GRD PDI@8 wk Nakabango 2020 | B04_10,528,843 | B04 | 10.50 | 2.76 | 0.05 | 2.75 | 6.62–115.86 | 12.56 |
| GRD_PDI_4_WEEKS_NAK_20 | GRD PDI@4 wk Nakabango 2020 | B05_138,125,178 | B05 | 138.00 | 3.23 | 0.03 | 2.9 | 41.5–146.58 | 11.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essandoh, D.A.; Odong, T.; Okello, D.K.; Fonceka, D.; Nguepjop, J.; Sambou, A.; Ballén-Taborda, C.; Chavarro, C.; Bertioli, D.J.; Leal-Bertioli, S.C.M. Quantitative Trait Analysis Shows the Potential for Alleles from the Wild Species Arachis batizocoi and A. duranensis to Improve Groundnut Disease Resistance and Yield in East Africa. Agronomy 2022, 12, 2202. https://doi.org/10.3390/agronomy12092202
Essandoh DA, Odong T, Okello DK, Fonceka D, Nguepjop J, Sambou A, Ballén-Taborda C, Chavarro C, Bertioli DJ, Leal-Bertioli SCM. Quantitative Trait Analysis Shows the Potential for Alleles from the Wild Species Arachis batizocoi and A. duranensis to Improve Groundnut Disease Resistance and Yield in East Africa. Agronomy. 2022; 12(9):2202. https://doi.org/10.3390/agronomy12092202
Chicago/Turabian StyleEssandoh, Danielle A., Thomas Odong, David K. Okello, Daniel Fonceka, Joël Nguepjop, Aissatou Sambou, Carolina Ballén-Taborda, Carolina Chavarro, David J. Bertioli, and Soraya C. M. Leal-Bertioli. 2022. "Quantitative Trait Analysis Shows the Potential for Alleles from the Wild Species Arachis batizocoi and A. duranensis to Improve Groundnut Disease Resistance and Yield in East Africa" Agronomy 12, no. 9: 2202. https://doi.org/10.3390/agronomy12092202
APA StyleEssandoh, D. A., Odong, T., Okello, D. K., Fonceka, D., Nguepjop, J., Sambou, A., Ballén-Taborda, C., Chavarro, C., Bertioli, D. J., & Leal-Bertioli, S. C. M. (2022). Quantitative Trait Analysis Shows the Potential for Alleles from the Wild Species Arachis batizocoi and A. duranensis to Improve Groundnut Disease Resistance and Yield in East Africa. Agronomy, 12(9), 2202. https://doi.org/10.3390/agronomy12092202