
Integrating Ecological Principles for Addressing Plant Production Security and Move beyond the Dichotomy ‘Good or Bad’ for Nitrogen Inputs Choice



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
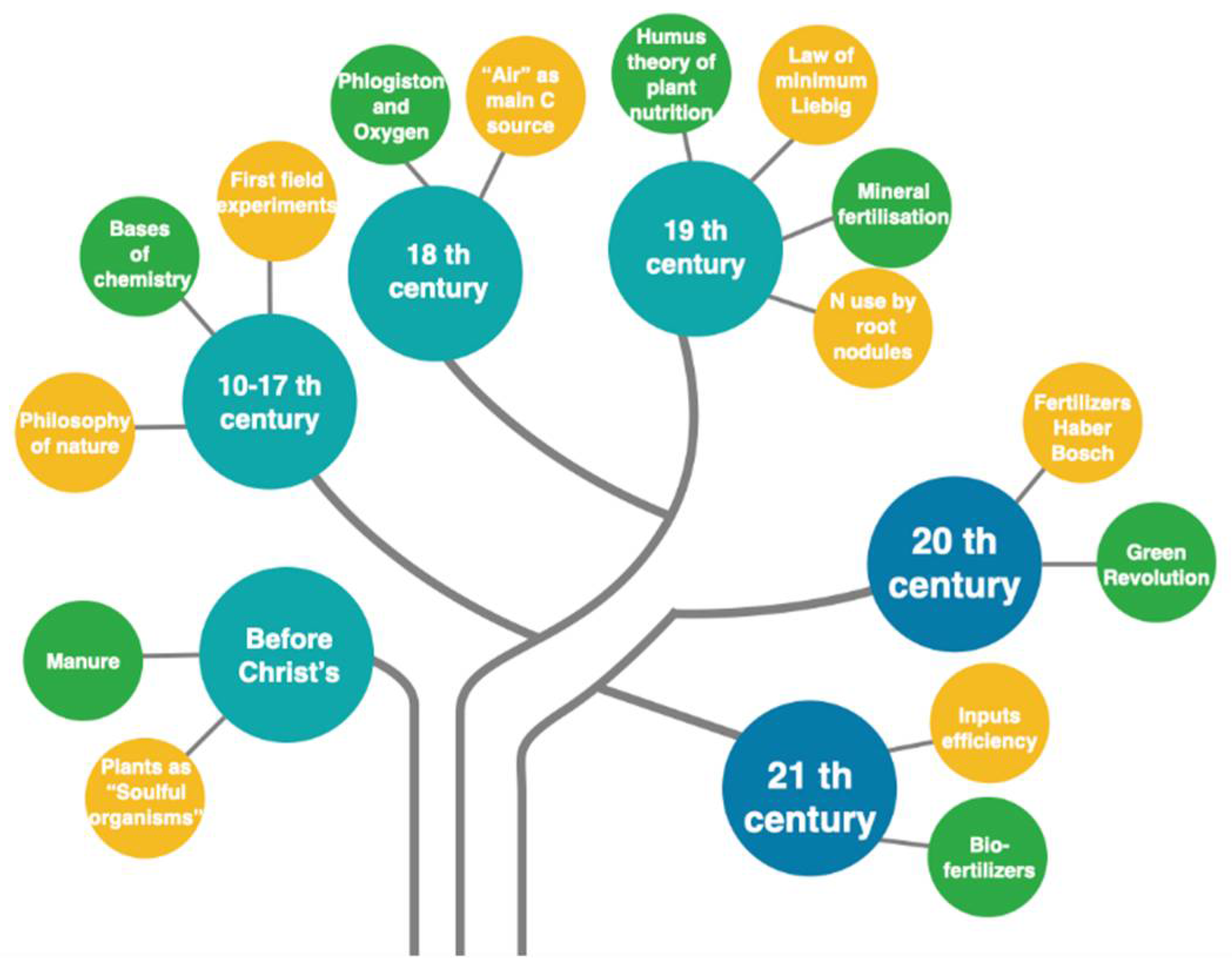
2. Plant Nutrition: The History of Humans on Earth
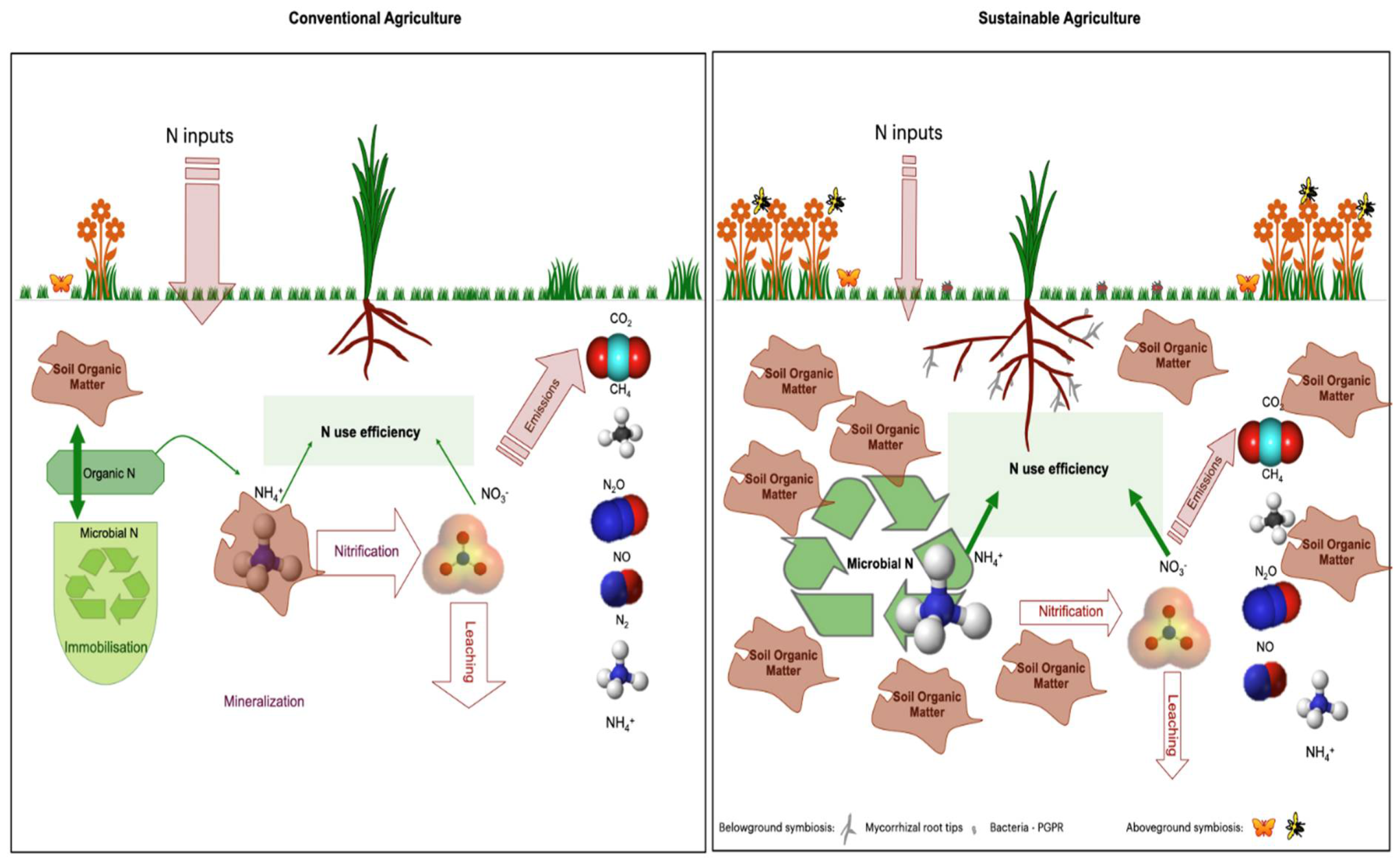
3. Plant Nitrogen Use Efficiency
4. Nature-Based Solutions for Better NUE
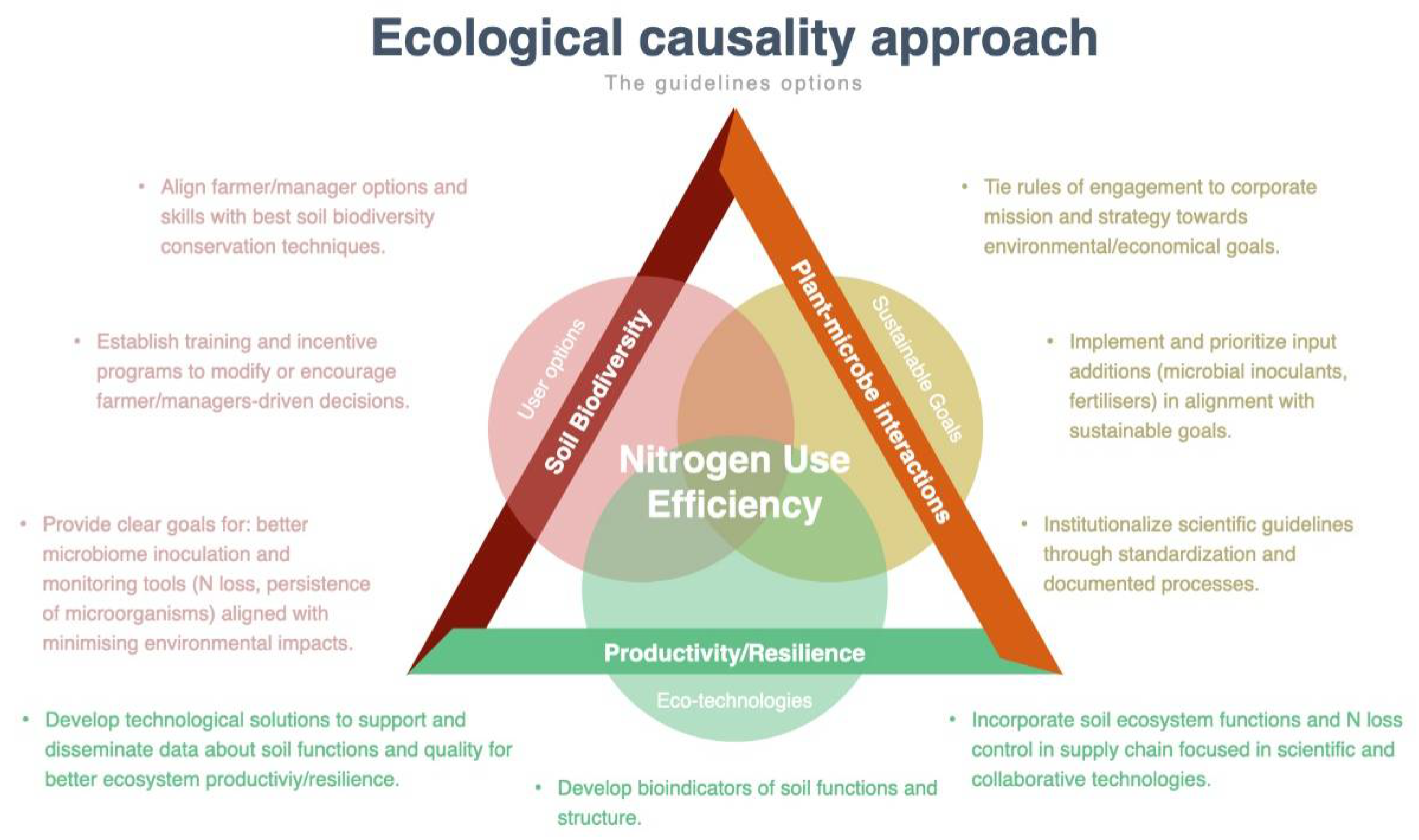
5. One Health Context within the Ecosystem
6. The Gospel of Choice
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richards, J.F. Land transformation. In The Earth as Transformed by Human Action: Global and Regional Changes in the Biosphere over the Past 300 Years; Turner, B.L., Clark, W.C., Kates, R.W., Richards, J.F., Mathews, J.T., Meyer, W.B., Eds.; CUP Archive: Cambridge, UK, 1990; pp. 163–178. [Google Scholar]
- Spengler, R.; Frachetti, M.; Doumani, P.; Rouse, L.; Cerasetti, B.; Bullion, E.; Mar’yashev, A. Early agriculture and crop transmission among Bronze Age mobile pastoralists of Central Eurasia. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133382. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Dukes, A.; Antunes, P.M. Accounting for soil biotic effects on soil health and crop productivity in the design of crop rotations. J. Sci. Food Agric. 2015, 95, 447–454. [Google Scholar] [CrossRef]
- Dias, T.; Pimentel, V.; Cogo, A.J.D.; Costa, R.; Bertolazi, A.A.; Miranda, C.; de Souza, S.B.; Melo, J.; Carolino, M.; Varma, A.; et al. The Free-Living Stage Growth Conditions of the Endophytic Fungus Serendipita indica May Regulate Its Potential as Plant Growth Promoting Microbe. Front. Microbiol. 2020, 11, 562238. [Google Scholar] [CrossRef]
- Galloway, J.; Aber, J.; Erisman, J. The Nitrogen Cascade. Bioscience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Sutton, M.A.; Howard, C.M.; Erisman, J.W.; Billen, G.; Bleeker, A.; Grennfelt, P.; van Grinsven, H.; Grizzetti, B. The European Nitrogen Assessment; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Smil, V. Nitrogen and Food. Ambio 2002, 31, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Malakoff, D. Death by suffocation in the Gulf of Mexico. Science 1998, 281, 190–192. [Google Scholar] [CrossRef]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The Genetics of Nitrogen Use Efficiency in Crop Plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef]
- Clarkson, D.T.; Hanson, J.B. The mineral nutrition of higher plants. Annu. Rev. Plant Physiol. 1980, 31, 239–298. [Google Scholar] [CrossRef]
- Fageria, N.K. The Use of Nutrients in Crop Plants; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Antonkiewicz, J.; Łabętowicz, J. Chemical innovation in plant nutrition in a historical continuum from ancient Greece and Rome until modern times. Chem.-Didact.-Ecol.-Metrol. 2016, 21, 29–43. [Google Scholar] [CrossRef]
- Finck, A. Fertilizers and Fertilization. In Introduction and Practical Guide to Crop Fertilization; Verlag Chemie: Weinheim, Germany, 1982. [Google Scholar]
- Starck, Z. Fizjologia roślin: Jak było wczoraj, jak jest dziś, a co przyniesie jutro? Kosmos 2014, 63, 569–589. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Burris, R.H. Biological nitrogen fixation: A scientific perspective. Plant Soil 1988, 108, 7–14. [Google Scholar] [CrossRef]
- Marciniec, B. Misja nauk chemicznych w zrównoważonym rozwoju. Nauka 2012, 1, 123–132. [Google Scholar]
- Gondek, K. Effect of fertilization with farmyard manure, municipal sewage sludge and compost from biodegradable waste on yield and mineral composition of spring wheat grain. J. Elem. 2012, 17, 231–245. [Google Scholar] [CrossRef]
- Voisin, A. Fertilizer Application. In Soil, Plant, Animal; Crosby Lockwood: London, UK, 1965. [Google Scholar]
- Kozhakhanov, T.E.; Lukashenko, S.N.; Larionova, N.V. Accumulation of artificial radionuclides in agricultural plants in the area used for surface nuclear tests. J. Environ. Radioact. 2014, 137, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.E.P.; Zocchi, S.S.; Baron, D. Reconciling the Mitscherlic’s law of diminishing returns with Liebig’s law of the minimum. Some results on crop modeling. Math. Biosci. 2017, 293, 29–37. [Google Scholar] [CrossRef]
- Huang, J.; Pray, C.; Rozelle, S. Enhancing the crops to feed the poor. Nature 2002, 418, 678–684. [Google Scholar] [CrossRef]
- Filipek-Mazur, B.; Lepiarczyk, A.; Tabak, M. Nitrogen and sulphur fertilization on yielding and zinc content in seeds of winter rape ‘Baldur’ cultivar. Ecol. Chem. Eng. A 2013, 20, 1351–1358. [Google Scholar]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. The effect of nitrogen source on photosynthesis of carob at high CO2concentrations. Physiol. Plant 1993, 89, 552–556. [Google Scholar] [CrossRef]
- Kelly, S. The Amount of Nitrogen Used for Photosynthesis Modulates Molecular Evolution in Plants. Mol. Biol. Evol. 2018, 35, 1616–1625. [Google Scholar] [CrossRef] [PubMed]
- Heffer, P.; Prud’homme, M. Global nitrogen fertilizer demand and supply: Trend, current level and outlook. In Proceedings of the Solutions to Improve Nitrogen Use Efficiency for the World, Melbourne, Australia, 4–8 December 2016. [Google Scholar]
- Galloway, J.N.; Bleeker, A.; Erisman, J.W. The Human Creation and Use of Reactive Nitrogen: A Global and Regional Perspective. Annu. Rev. Environ. Resour. 2021, 46, 255–288. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef]
- Pinho, P.; Dias, T.; Cruz, C.; Tang, Y.S.; Sutton, M.A.; Martins-Loucao, M.A.; Maguas, C.; Branquinho, C. Using lichen functional diversity to assess the effects of atmospheric ammonia in Mediterranean woodlands. J. Appl. Ecol. 2011, 48, 1107–1116. [Google Scholar] [CrossRef]
- Pinho, P.; Theobald, M.R.; Dias, T.; Tang, Y.S.; Cruz, C.; Martins-Loução, M.A.; Máguas, C.; Sutton, M.; Branquinho, C. Critical loads of nitrogen deposition and critical levels of atmospheric ammonia for semi-natural Mediterranean evergreen woodlands. Biogeosciences 2012, 9, 1205–1215. [Google Scholar] [CrossRef]
- Dias, T.; Oakley, S.; Alarcón-Gutiérrez, E.; Ziarelli, F.; Trindade, H.; Martins-Loução, M.A.; Sheppard, L.; Ostle, N.; Cruz, C. N-driven changes in a plant community affect leaf-litter traits and may delay organic matter decomposition in a Mediterranean maquis. Soil Biol. Biochem. 2013, 58, 163–171. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Searchinger, T.D. Opinion: A “more ammonium solution” to mitigate nitrogen pollution and boost crop yields. Proc. Natl. Acad. Sci. USA 2021, 118, e2107576118. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. Growth and nutrition of carob plants as affected by nitrogen sources. J. Plant Nutr. 1993, 16, 1–15. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. Interactions between nitrate and ammonium during uptake by carob seedlings and the effect of the form of earlier nitrogen nutrition. Physiol. Plant 1993, 89, 544–551. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. Nitrogen assimilation and transport in carob plants. Physiol. Plant 1993, 89, 524–531. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. The effect of root temperature on carob growth. Nitrate versus ammonium nutrition. J. Plant Nutr. 1993, 16, 1517–1530. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loução, M.A. Uptake of ammonium and nitrate by carob (Ceratonia siliqua) as affected by root temperature and inhibitors. Physiol. Plant 1993, 89, 532–543. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, H.; Martins-Loução, M.A. Nitrogen use efficiency by a slow-growing species as affected by CO2 levels, root temperature, N source and availability. J. Plant Physiol. 2003, 160, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.; Bio, A.F.M.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Martins-Loução, M.A. How does glutamine synthetase activity determine plant tolerance to ammonium? Planta 2006, 223, 1068–1080. [Google Scholar] [CrossRef]
- von Wirén, N.; Gazzarrini, S.; Gojon, A.; Frommer, W.B. The molecular physiology of ammonium uptake and retrieval. Curr. Opin. Plant Biol. 2000, 3, 254–261. [Google Scholar] [CrossRef]
- Ladha, J.K.; Kirk, G.J.D.; Bennett, J.; Peng, S.; Reddy, C.K.; Reddy, P.M.; Singh, U. Opportunities for increased nitrogen-use efficiency from improved lowland rice germplasm. Field Crop Res. 1998, 56, 41–71. [Google Scholar] [CrossRef]
- Iqbal, A.; Qiang, D.; Alamzeb, M.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Nianchang, P.; Xiling, Z.; Meizhen, S. Untangling the molecular mechanisms and functions of nitrate to improve nitrogen use efficiency. J. Sci. Food Agric. 2020, 100, 904–914. [Google Scholar] [CrossRef]
- Hawkesford, M.J. Genetic variation in traits for nitrogen use efficiency in wheat. J. Exp. Bot. 2017, 68, 2627–2632. [Google Scholar] [CrossRef]
- Pujarula, V.; Pusuluri, M.; Bollam, S.; Das, R.R.; Ratnala, R.; Adapala, G.; Thuraga, V.; Rathore, A.; Srivastava, R.K.; Gupta, R. Genetic Variation for Nitrogen Use Efficiency Traits in Global Diversity Panel and Parents of Mapping Populations in Pearl Millet. Front. Plant Sci. 2021, 12, 625915. [Google Scholar] [CrossRef]
- Poorter, H.; Lambers, H. Is interspecific variation in relative growth rate positively correlated with biomass allocation to the leaves? Am. Nat. 1991, 138, 1264–1268. [Google Scholar] [CrossRef]
- Atkin, O.K.; Botman, B.; Lambers, H. The causes of inherently slow growth in alpine plants: An analysis based on the underlying carbon economies of alpine and lowland Poa species. Funct. Ecol. 1996, 10, 698–707. [Google Scholar] [CrossRef]
- Mata, C.; van Vemde, N.; Clarkson, D.T.; Martins-Loucao, M.A.; Lambers, H. Influx, efflux and net uptake of nitrate in Quercus suber seedlings. Plant Soil 2000, 221, 25–32. [Google Scholar] [CrossRef]
- Cruz, C.; Bio, A.M.F.; Jullioti, A.; Tavares, A.; Dias, T.; Martins-Loução, M.A. Heterogeneity of soil surface ammonium concentration and other characteristics, related to plant specific variability in a Mediterranean-type ecosystem. Environ. Pollut. 2008, 154, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Martins-Loução, M.A.; Sheppard, L.; Cruz, C. The strength of the biotic compartment in retaining nitrogen additions prevents nitrogen losses from a Mediterranean maquis. Biogeosciences 2012, 9, 193–201. [Google Scholar] [CrossRef]
- Dias, T.; Malveiro, S.; Martins-Loução, M.; Shepard, L.; Cruz, C. Linking N-driven biodiversity changes with soil N availability in a Mediterranean ecosystem. Plant Soil 2011, 341, 125–136. [Google Scholar] [CrossRef]
- Dias, T.; Clemente, A.; Martins-Loução, M.; Sheppard, L.; Bobbink, R.; Cruz, C. Ammonium as a Driving Force of Plant Diversity and Ecosystem Functioning: Observations Based on 5 Years’ Manipulation of N Dose and Form in a Mediterranean Ecosystem. PLoS ONE 2014, 9, e92517. [Google Scholar] [CrossRef]
- Dias, T.; Crous, C.J.; Liberati, D.; Munzi, S.; Gouveia, C.; Ulm, F.; Afonso, A.C.; Ochoa-Hueso, R.; Manrique, E.; Sheppard, L.; et al. Alleviating Nitrogen limitation in Mediterranean maquis vegetation leads to ecological degradation. Land Degrad. Dev. 2017, 28, 2482–2492. [Google Scholar] [CrossRef]
- Ulm, F.; Gouveia, C.; Dias, T.; Cruz, C. N fertilization in a Mediterranean ecosystem alters N and P turnover in soil, roots and the ectomycorrhizal community. Soil Biol. Biochem. 2017, 113, 60–70. [Google Scholar] [CrossRef]
- Dias, T.; Crous, C.J.; Ochoa-Hueso, R.; Manrique, E.; Martins-Loução, M.A.; Cruz, C. Nitrogen inputs may improve soil biocrusts multifunctionality in dryland ecosystems. Soil Biol. Biochem. 2020, 149, 107947. [Google Scholar] [CrossRef]
- Cruz, C.; Lips, S.H.; Martins-Loucao, M.A. Uptake regions of inorganic nitrogen in roots of carob seedlings. Physiol. Plant 1995, 95, 167–175. [Google Scholar] [CrossRef]
- Cruz, C.; Green, J.J.; Watson, C.A.; Wilson, F.; Martins-Loução, M.A. Functional aspects of root architecture and mycorrhizal inoculation with respect to nutrient uptake capacity. Mycorrhiza 2004, 14, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C.; Lips, S.H.; Martins-Loucao, M.A. Changes in the morphology of roots and leaves of carob seedlings induced by nitrogen source and atmospheric carbon dioxide. Ann. Bot. 1997, 80, 817–823. [Google Scholar] [CrossRef]
- Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Cruz, C.; Martins-Loução, M.A.; Moran, J.F. Nitrogen nutrition and antioxidant metabolism in ammonium-tolerant and -sensitive plants. Physiol. Plant 2008, 132, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Neto, D.; Martins-Loucao, M.A.; Sheppard, L.; Cruz, C. Patterns of nitrate reductase activity vary according to the plant functional group in a Mediterranean maquis. Plant Soil 2011, 347, 363–376. [Google Scholar] [CrossRef][Green Version]
- Dias, T.; Martins-Loução, M.; Sheppard, L.; Cruz, C. Plant tolerance of ammonium varies between co-existing Mediterranean species. Plant Soil 2015, 395, 243–252. [Google Scholar] [CrossRef]
- Marschner, H.; Kirkby, E.A.; Cakmak, I. Effect of mineral nutritional status on shoot-root partitioning of photoassimilates and cycling of mineral nutrients. J. Exp. Bot. 1996, 47, 1255–1263. [Google Scholar] [CrossRef]
- Martins-Loução, M.A.; Cruz, C. The Role of Nitrogen Source in Carbon Balance. In Modes of Nitrogen Nutrition in Higher Plants; Srivastava, H.S., Singh., R.P., Eds.; Oxford & IBH Publishing Co. Pvt: New Delhi, India, 1999; pp. 231–282. [Google Scholar]
- Wang, L.; Ruan, Y.-L. Shoot–root carbon allocation, sugar signalling and their coupling with nitrogen uptake and assimilation. Funct. Plant Biol. 2016, 43, 105–113. [Google Scholar] [CrossRef]
- Buet, A.; Galatro, A.; Ramos-Artuso, F.; Simontacchi, M. Nitric oxide and plant mineral nutrition: Current knowledge. J. Exp. Bot. 2019, 70, 4461–4476. [Google Scholar] [CrossRef]
- Forde, B. The role of long-distance signalling in plant responses to nitrate and other nutrients. J. Exp. Bot. 2002, 53, 39–43. [Google Scholar]
- Kang, A.; Zhang, N.; Xun, W.; Dong, X.; Xiao, M.; Liu, Z.; Xu, Z.; Feng, H.; Zou, J.; Shen, Q.; et al. Nitrogen fertilization modulates beneficial rhizosphere interactions through signaling effect of nitric oxide. Plant Physiol. 2022, 188, 1129–1140. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Glass, A.D.M.; Siddiqi, M.Y. Nitrate induction in spruce: An approach using compartmental analysis. Planta 1995, 196, 683–690. [Google Scholar] [CrossRef]
- Lips, S.H.; Ben Zioni, A.; Vaadia, Y. K+ recirculation in plants and its importance for adequate nitrate reduction. In Recent Advances in Plant Nutrition; Samish, R.M., Gordon, Breach, Eds.; Gordon and Breach Science Publishers Inc.: New York, NY, USA, 1970; Volume 1, pp. 207–215. [Google Scholar]
- Ben-Zioni, A.; Vaadia, Y.; Lips, S.H. Nitrate uptake by roots as regulated by nitrate reduction products of the shoot. Physiol. Plant 1971, 24, 288–290. [Google Scholar] [CrossRef]
- Lips, S.H.; Soares, M.I.M.; Kaiser, J.J.; Lewis, O.A.M. K+modulation of nitrogen uptake and assimilation in plants. In Inorganic Nitrogen Metabolism; Syrett, P.J., Ulrich, W.R., Aparicio, P.J., Castillo, F., Eds.; Springer: Berlin, Germany, 1987; pp. 233–239. [Google Scholar]
- Martins-Loução, M.A.; Cruz, C.; Correia, P.M. New approaches to enhanced ammonium assimilation in plants. In Nitrogen in a Sustainable Ecosystem: From the Cell to the Plant; Martins-Loução, M., Lips, S.H., Eds.; Backhuys: Leiden, The Netherlands, 2000; pp. 349–360. [Google Scholar]
- Fonseca, M.B.; Dias, T.; Carolino, M.M.; França, M.G.C.; Cruz, C. Belowground microbes mitigate plant-plant competition. Plant Sci. 2017, 262, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef]
- Cruz, C.; Domínguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Bio, A.; Martins-Loução, M.A.; Moran, J.F. Intra-specific variation in pea responses to ammonium nutrition leads to different degrees of tolerance. Environ. Exp. Bot. 2011, 70, 233–243. [Google Scholar] [CrossRef]
- Ariz, I.; Cruz, C.; Moran, J.F.; González-Moro, M.B.; García-Olaverri, C.; González-Murua, C.; Martins-Loução, M.A.; Aparicio-Tejo, P.M. Depletion of the heaviest stable N isotope is associated with NH4+/NH3 toxicity in NH4+-fed plants. BMC Plant Biol. 2011, 11, 83. [Google Scholar] [CrossRef]
- Ariz, I.; Boeckstaens, M.; Gouveia, C.; Martins Ana, P.; Sanz-Luque, E.; Fernández, E.; Soveral, G.; von Wirén, N.; Marini Anna, M.; Aparicio-Tejo Pedro, M.; et al. Nitrogen isotope signature evidences ammonium deprotonation as a common transport mechanism for the AMT-Mep-Rh protein superfamily. Sci. Adv. 2018, 4, eaar3599. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Pohl, P.; Denisov, G.A. Permeation of ammonia across bilayer lipid membranes studied by ammonium ion selective microelectrodes. Biophys. J. 1997, 72, 2187–2195. [Google Scholar] [CrossRef]
- Ninnemann, O.; Jauniaux, J.; Frommer, W. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef]
- Martins-Loução, M.A.; Lips, S. Nitrogen from the Cell to the Plant: Recent Progress and Perspectives; Martins-Loução, M.A., Lips, S., Eds.; Backhuys: Leiden, The Netherlands, 2000; pp. 3–6. [Google Scholar]
- Ladha, J.K.; Pathak, H.; Krupnik, T.J.; Six, J.; van Kessel, C. Efficiency of Fertilizer Nitrogen in Cereal Production: Retrospects and Prospects. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2005; Volume 87, pp. 85–156. [Google Scholar]
- Baron, J.S.; Barber, M.; Adams, M.; Agboola, J.I.; Allen, E.B.; Bealey, W.J.; Bobbink, R.; Bobrovsky, M.V.; Bowman, W.D.; Branquinho, C. The effects of atmospheric nitrogen deposition on terrestrial and freshwater biodiversity. In Nitrogen Deposition, Critical Loads and Biodiversity; Springer: Berlin/Heidelberg, Germany, 2014; pp. 465–480. [Google Scholar]
- Bijlsma, R.; Lambers, H.; Kooijman, S. A dynamic whole-plant model of integrated metabolism of nitrogen and carbon. 1. Comparative ecological implications of ammonium-nitrate interactions. Plant Soil 2000, 220, 49–69. [Google Scholar] [CrossRef]
- Cruz, C.; Dias, T.; Matos, S.; Tavares, A.; Neto, D.; Martins-Loucao, M.A. Nitrogen availability and plant cover: The importance of nitrogen pools. In Ecosystems and Sustainable Development Iv, Vols 1 and 2; Tiezzi, E., Brebbia, C.A., Uso, J.L., Eds.; Advances in Ecological Sciences; WIT Press: Boston, MA, USA, 2003; Volume 18–19, pp. 123–135. [Google Scholar]
- Cruz, C.; Fegghi, Z.; Martins-Loução, M.; Varma, A. Plant Nitrogen Use Efficiency May Be Improved Through Symbiosis with Piriformospora indica. In Piriformospora Indica; Varma, A., Kost, G., Oelmüller, R., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 33, pp. 285–293. [Google Scholar]
- Dias, T.; Stürmer, S.; Chaves, S.; Fidalgo, C.; Tenreiro, R.; Correia, P.; Carvalho, L.; Martins-Loução, M.; Shepard, L.; Cruz, C. Species of Arbuscular Mycorrhizal Fungal Spores can Indicate Increased Nitrogen Availability in Mediterranean-type Ecosystems. In Nitrogen Deposition, Critical Loads and Biodiversity; Sutton, M.A., Mason, K.E., Sheppard, L.J., Sverdrup, H., Haeuber, R., Hicks, W.K., Eds.; Springer: Berlin, Germany; Dordrecht, The Netherlands, 2014; pp. 259–266. [Google Scholar]
- Cruz, C.; Dias, T.; Pinho, P.; Branquinho, C.; Máguas, C.; Pinto, M.J.; Martins-Loução, M.A. Policies for plant diversity conservation on a global scale: A SWOT analysis. Kew Bull. 2010, 65, 525–528. [Google Scholar] [CrossRef]
- FAO. Climate Change and Food Security: Risks and Responses; FAO: Rome, Italy, 2015; p. 86. [Google Scholar]
- Lawrencia, D.; Wong, S.K.; Low, D.Y.; Goh, B.H.; Goh, J.K.; Ruktanonchai, U.R.; Soottitantawat, A.; Lee, L.H.; Tang, S.Y. Controlled Release Fertilizers: A Review on Coating Materials and Mechanism of Release. Plants 2021, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Besharat, S.; Barão, L.; Cruz, C. New strategies to overcome water limitation in cultivated maize: Results from sub-surface irrigation and silicon fertilization. J. Environ. Manag. 2020, 263, 110398. [Google Scholar] [CrossRef] [PubMed]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J. The use of reed canary grass and giant miscanthus in the phytoremediation of municipal sewage sludge. Environ. Sci. Pollut. Res. 2016, 23, 9505–9517. [Google Scholar] [CrossRef]
- Ulm, F.; Avelar, D.; Hobson, P.; Penha-Lopes, G.; Dias, T.; Máguas, C.; Cruz, C. Sustainable urban agriculture using compost and an open-pollinated maize variety. J. Clean. Prod. 2019, 212, 622–629. [Google Scholar] [CrossRef]
- Gondek, K.; Mierzwa-Hersztek, M. Effect of low-temperature biochar derived from pig manure and poultry litter on mobile and organic matter-bound forms of Cu, Cd, Pb and Zn in sandy soil. Soil Use Manag. 2016, 32, 357–367. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Chapter Two-Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 162, pp. 31–87. [Google Scholar]
- Hawkes, C.V.; Bull, J.J.; Lau, J.A. Symbiosis and stress: How plant microbiomes affect host evolution. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190590. [Google Scholar] [CrossRef]
- Munzi, S.; Cruz, C.; Corrêa, A. When the exception becomes the rule: An integrative approach to symbiosis. Sci. Total Environ. 2019, 672, 855–861. [Google Scholar] [CrossRef]
- Ramos, A.C.; Melo, J.; de Souza, S.B.; Bertolazi, A.A.; Silva, R.A.; Rodrigues, W.P.; Campostrini, E.; Olivares, F.L.; Eutrópio, F.J.; Cruz, C.; et al. Inoculation with the endophytic bacterium Herbaspirillum seropedicae promotes growth, nutrient uptake and photosynthetic efficiency in rice. Planta 2020, 252, 87. [Google Scholar] [CrossRef] [PubMed]
- Antoun, H.; Prévost, D. Ecology of Plant Growth Promoting Rhizobacteria. In PGPR: Biocontrol and Biofertilization; Siddiqui, Z.A., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–38. [Google Scholar]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant-microbiome interactions under a changing world: Responses, consequences, and perspective. New Phytol. 2022, 34, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Correia, P.; Carvalho, L.; Melo, J.; de Varennes, A.; Cruz, C. Arbuscular mycorrhizal fungal species differ in their capacity to overrule the soil’s legacy from maize monocropping. Appl. Soil Ecol. 2018, 125, 177–183. [Google Scholar] [CrossRef]
- Basílio, F.; Dias, T.; Santana, M.; Melo, J.; Carvalho, L.; Correia, P.; Cruz, C. Multiple modes of action are needed to unlock soil phosphorus fractions unavailable for plants: The example of bacteria- and fungi-based biofertilizers. Appl. Soil Ecol. 2022; under review. [Google Scholar]
- Fravel, D.R. Commercialization and Implementation of Biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Ramakuwela, T.; Hatting, J.; Bock, C.; Vega, F.E.; Wells, L.; Mbata, G.N.; Shapiro-Ilan, D. Establishment of Beauveria bassiana as a fungal endophyte in pecan (Carya illinoinensis) seedlings and its virulence against pecan insect pests. Biol. Control 2020, 140, 104102. [Google Scholar] [CrossRef]
- Melo, J.; Carolino, M.; Carvalho, L.; Correia, P.; Tenreiro, R.; Chaves, S.; Meleiro, A.I.; de Souza, S.B.; Dias, T.; Cruz, C.; et al. Crop management as a driving force of plant growth promoting rhizobacteria physiology. SpringerPlus 2016, 5, 1574. [Google Scholar] [CrossRef]
- Santana, M.M.; Gonzalez, J.M.; Cruz, C. Nitric Oxide Accumulation: The Evolutionary Trigger for Phytopathogenesis. Front. Microbiol. 2017, 8, 1947. [Google Scholar] [CrossRef]
- Canton, G.C.; Bertolazi, A.A.; Cogo, A.J.; Eutrópio, F.J.; Melo, J.; de Souza, S.B.; AKrohling, C.; Campostrini, E.; da Silva, A.G.; Façanha, A.R.; et al. Biochemical and ecophysiological responses to manganese stress by ectomycorrhizal fungus Pisolithus tinctorius and in association with Eucalyptus grandis. Mycorrhiza 2016, 26, 475–487. [Google Scholar] [CrossRef]
- Lefort, M.-C.; McKinnon, A.C.; Nelson, T.L.; Glare, T.R. Natural occurrence of the entomopathogenic fungi Beauveria bassiana as a vertically transmitted endophyte of Pinus radiata and its effect on above- and below-ground insect pests. N. Z. Plant Prot. 2016, 69, 68–77. [Google Scholar] [CrossRef]
- Destoumieux-Garzón, D.; Mavingui, P.; Boetsch, G.; Boissier, J.; Darriet, F.; Duboz, P.; Fritsch, C.; Giraudoux, P.; Le Roux, F.; Morand, S.; et al. The One Health Concept: 10 Years Old and a Long Road Ahead. Front. Vet. Sci. 2018, 5, 14. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.A.-O.; Delgado-Baquerizo, M.; Duarte, E.; Marigliano, O.; Görgen, C.; Maestre, F.T.; Eisenhauer, N.A.-O. Global projections of the soil microbiome in the Anthropocene. Glob. Ecol. Biogeogr. 2021, 30, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Fox, A.; Widmer, F.; Barreiro, A.; Jongen, M.; Musyoki, M.; Vieira, Â.; Zimmermann, J.; Cruz, C.; Dimitrova-Mårtensson, L.M.; Rasche, F.; et al. Small-scale agricultural grassland management can affect soil fungal community structure as much as continental scale geographic patterns. FEMS Microbiol. Ecol. 2021, 97, fiab148. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, A.; Fox, A.; Jongen, M.; Melo, J.; Musyoki, M.; Vieira, A.; Zimmermann, J.; Carlsson, G.; Cruz, C.; Lüscher, A.; et al. Soil bacteria respond to regional edapho-climatic conditions while soil fungi respond to management intensity in grasslands along a European transect. Appl. Soil Ecol. 2022, 170, 104264. [Google Scholar] [CrossRef]
- Hirota, M.; Oliveira, R. Crossing thresholds on the way to ecosystem shifts. Science 2020, 367, 739–740. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Winiwarter, W.; Leip, A.; Leach, A.M.; Bleeker, A.; Erisman, J.W. Nitrogen footprints: Past, present and future. Environ. Res. Lett. 2014, 9, 115003. [Google Scholar] [CrossRef]
- Erisman, J.W.; Sutton, M.A.; Galloway, J.; Klimont, Z.; Winiwarter, W. How a century of ammonia synthesis changed the world. Nat. Geosci. 2008, 1, 636–639. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Munzi, S.; Alonso, R.; Arróniz-Crespo, M.; Avila, A.; Bermejo, V.; Bobbink, R.; Branquinho, C.; Concostrina-Zubiri, L.; Cruz, C.; et al. Ecological impacts of atmospheric pollution and interactions with climate change in terrestrial ecosystems of the Mediterranean Basin: Current research and future directions. Environ. Pollut. 2017, 227, 194–206. [Google Scholar] [CrossRef]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, Å.; Chapin, F.S.I.; Lambin, E.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. Planetary Boundaries: Exploring the Safe Operating Space for Humanity. Ecol. Soc. 2009, 14, 32–63. [Google Scholar] [CrossRef]
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef]
- Cruz, C.; Martins-Loução, M.A.; Varma, A. The influence of plant co-culture of tomato plants with Piriformospora indica on biomass accumulation and stress tolerance. Acta Hortic. 2010, 868, 123–127. [Google Scholar] [CrossRef]
- Hessini, K.; Kronzucker, H.J.; Abdelly, C.; Cruz, C. Drought stress obliterates the preference for ammonium as an N source in the C4 plant Spartina alterniflora. J. Plant Physiol. 2017, 213, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, N.; Caeiro, M.F.; Mahdhi, M.; Tenreiro, R.; Ulm, F.; Mars, M.; Cruz, C.; Dias, T. Arbuscular mycorrhizal traits are good indicators of soil multifunctionality in drylands. Geoderma 2021, 397, 115099. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-Loução, M.A.; Dias, T.; Cruz, C. Integrating Ecological Principles for Addressing Plant Production Security and Move beyond the Dichotomy ‘Good or Bad’ for Nitrogen Inputs Choice. Agronomy 2022, 12, 1632. https://doi.org/10.3390/agronomy12071632
Martins-Loução MA, Dias T, Cruz C. Integrating Ecological Principles for Addressing Plant Production Security and Move beyond the Dichotomy ‘Good or Bad’ for Nitrogen Inputs Choice. Agronomy. 2022; 12(7):1632. https://doi.org/10.3390/agronomy12071632
Chicago/Turabian StyleMartins-Loução, Maria Amélia, Teresa Dias, and Cristina Cruz. 2022. "Integrating Ecological Principles for Addressing Plant Production Security and Move beyond the Dichotomy ‘Good or Bad’ for Nitrogen Inputs Choice" Agronomy 12, no. 7: 1632. https://doi.org/10.3390/agronomy12071632
APA StyleMartins-Loução, M. A., Dias, T., & Cruz, C. (2022). Integrating Ecological Principles for Addressing Plant Production Security and Move beyond the Dichotomy ‘Good or Bad’ for Nitrogen Inputs Choice. Agronomy, 12(7), 1632. https://doi.org/10.3390/agronomy12071632