
Fertilizer Potential of Organic-Based Soil Amendments on cv. Sangiovese (V. vinifera L.) Vines: Preliminary Results

 ,
, 
 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vineyard Description and Treatments
- (1)
- Mineral fertilization (MIN): N supply was split into spring (half May) and summer (half June) and fertilization was performed with N, phosphorous (P), and potassium (K) fertilizer, and urea, respectively. Fertilizers were distributed manually on the soil at a total rate (spring + summer) of 40 and 60 kg N ha−1, in 2020 and 2021, respectively. The increased rate from 2020 to 2021 is a consequence of the increase of plants’ needs due to plant growth and is in line with the ICM Guidelines of the Emilia-Romagna region [26].
- (2)
- MOW compost (MOW): applied at a rate of 12.9 and 19.1 t DW ha−1 in 2020 and 2021, respectively.
- (3)
- SS compost (SS): applied at a rate of 18.6 and 11.1 t DW ha−1 in 2020 and 2021, respectively.
2.2. Soil Sampling and Analysis
2.3. Leaf Sampling and Analysis
2.4. Berry Sampling and Analysis
2.5. Vegetative Growth Determination
2.6. Statistical Analysis
3. Results
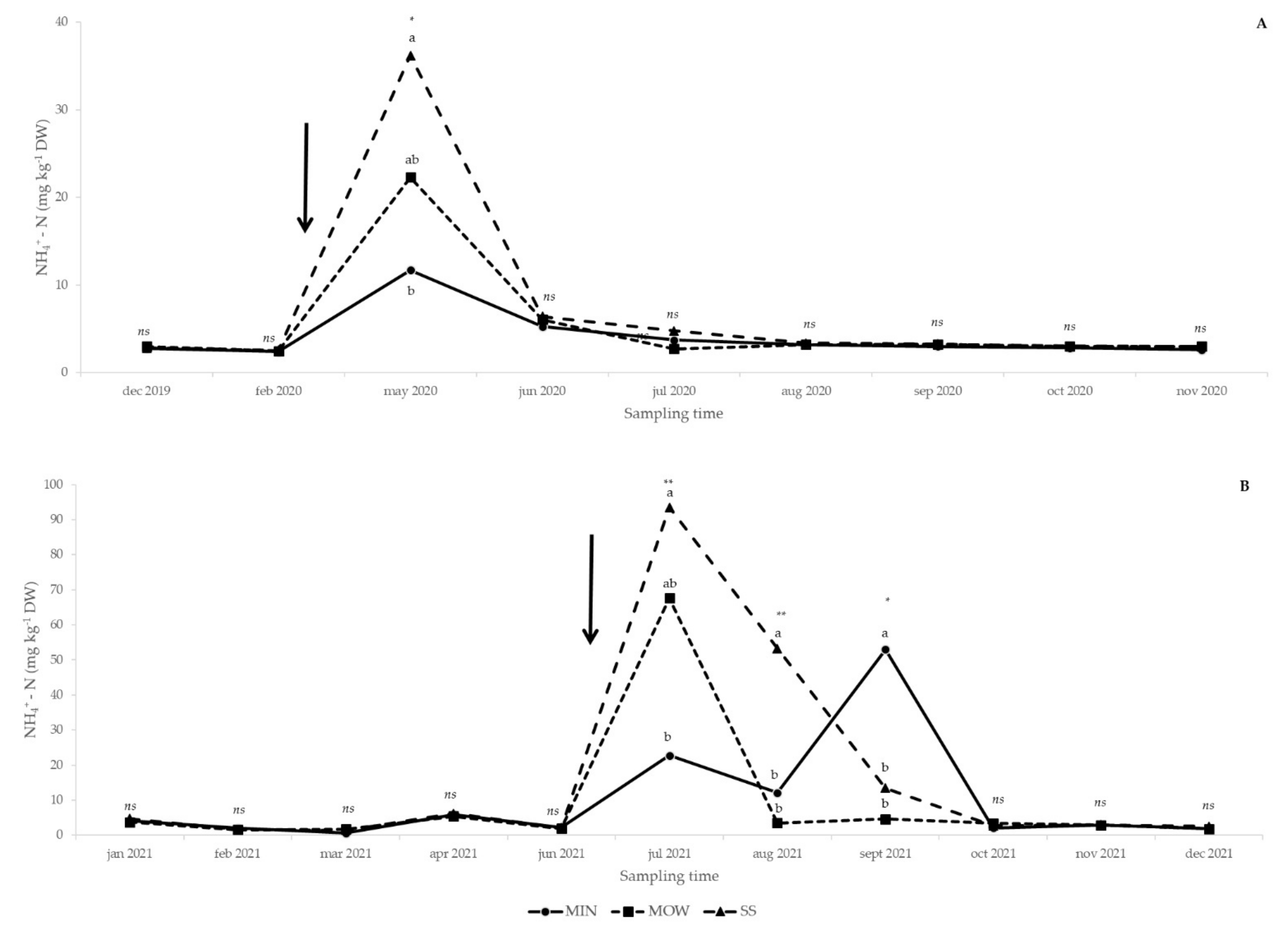
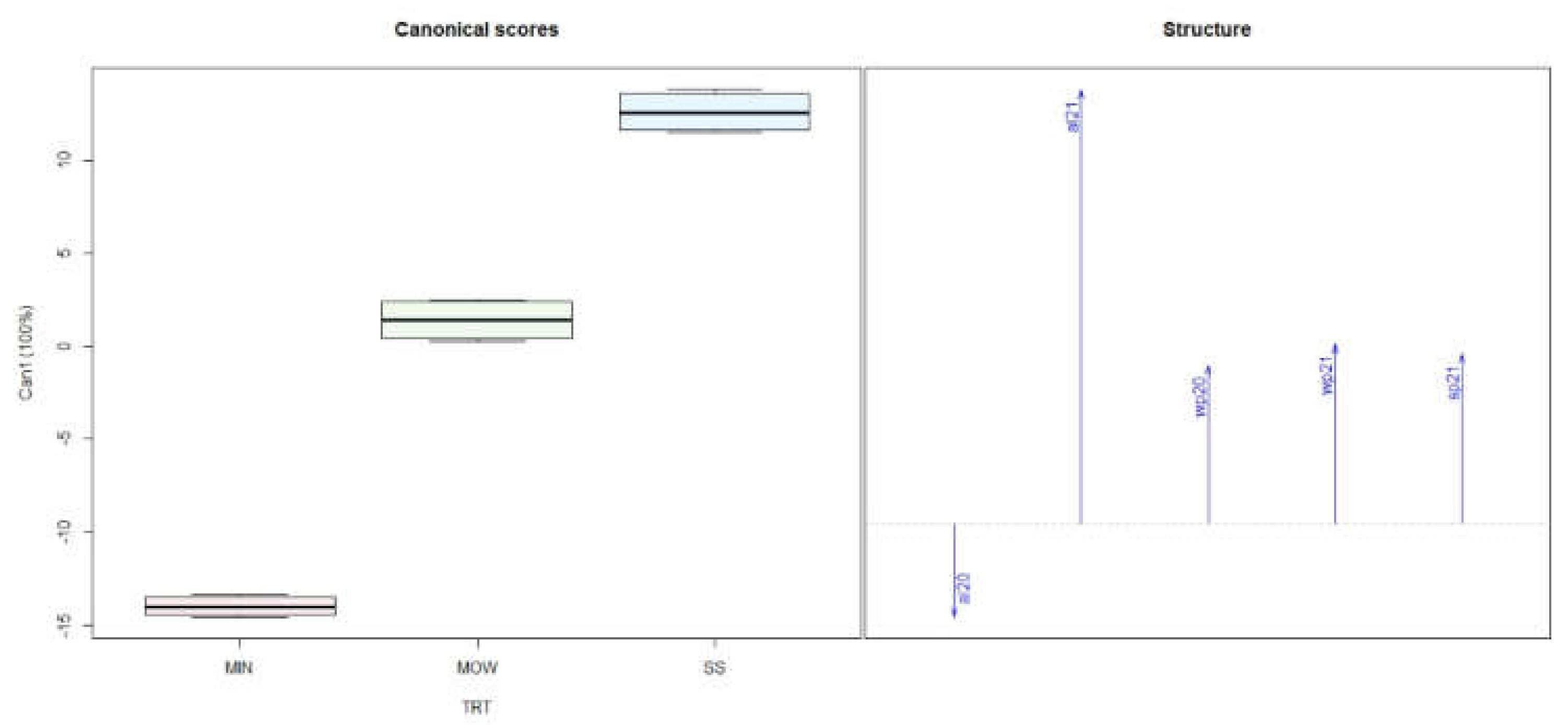
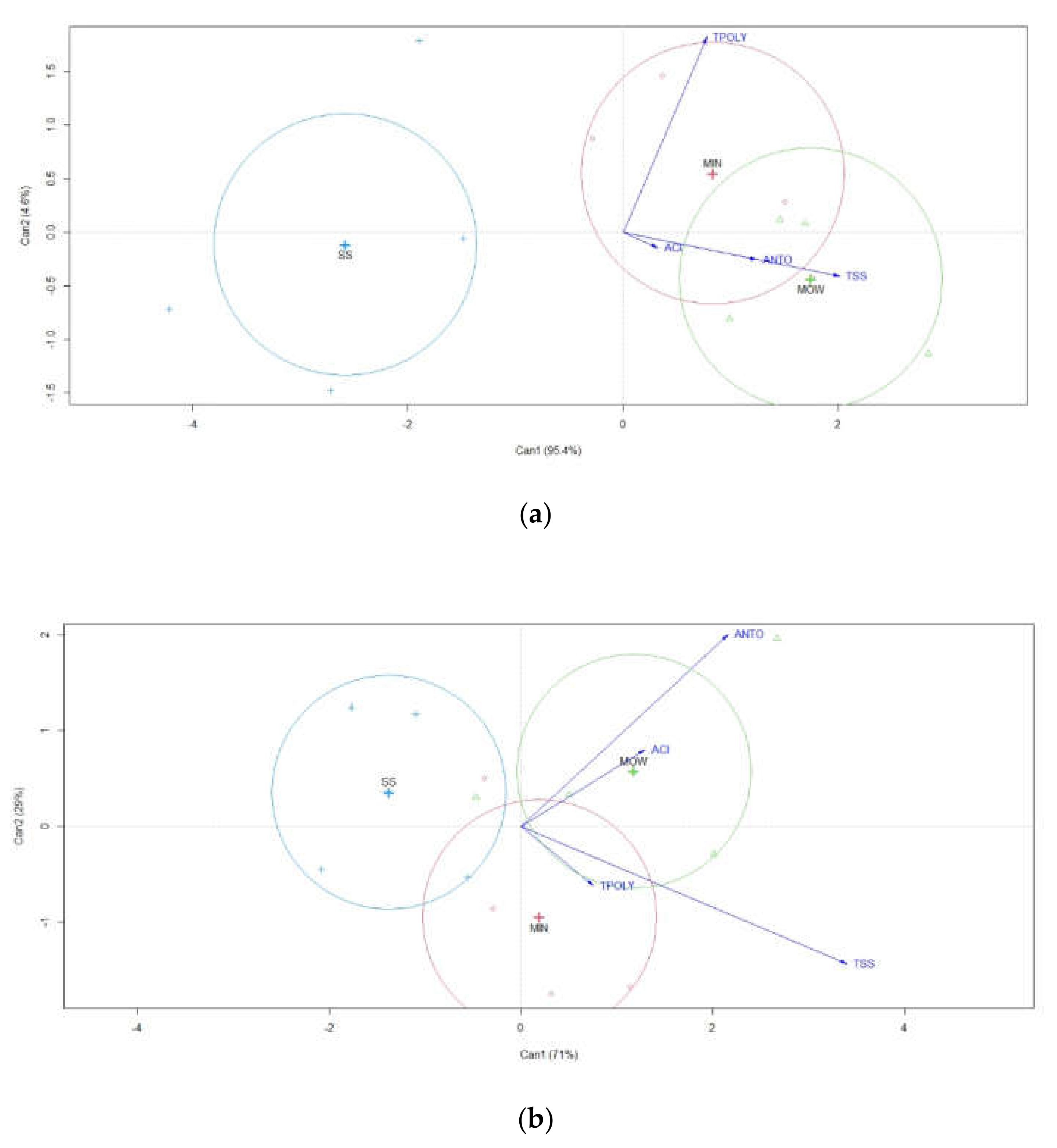
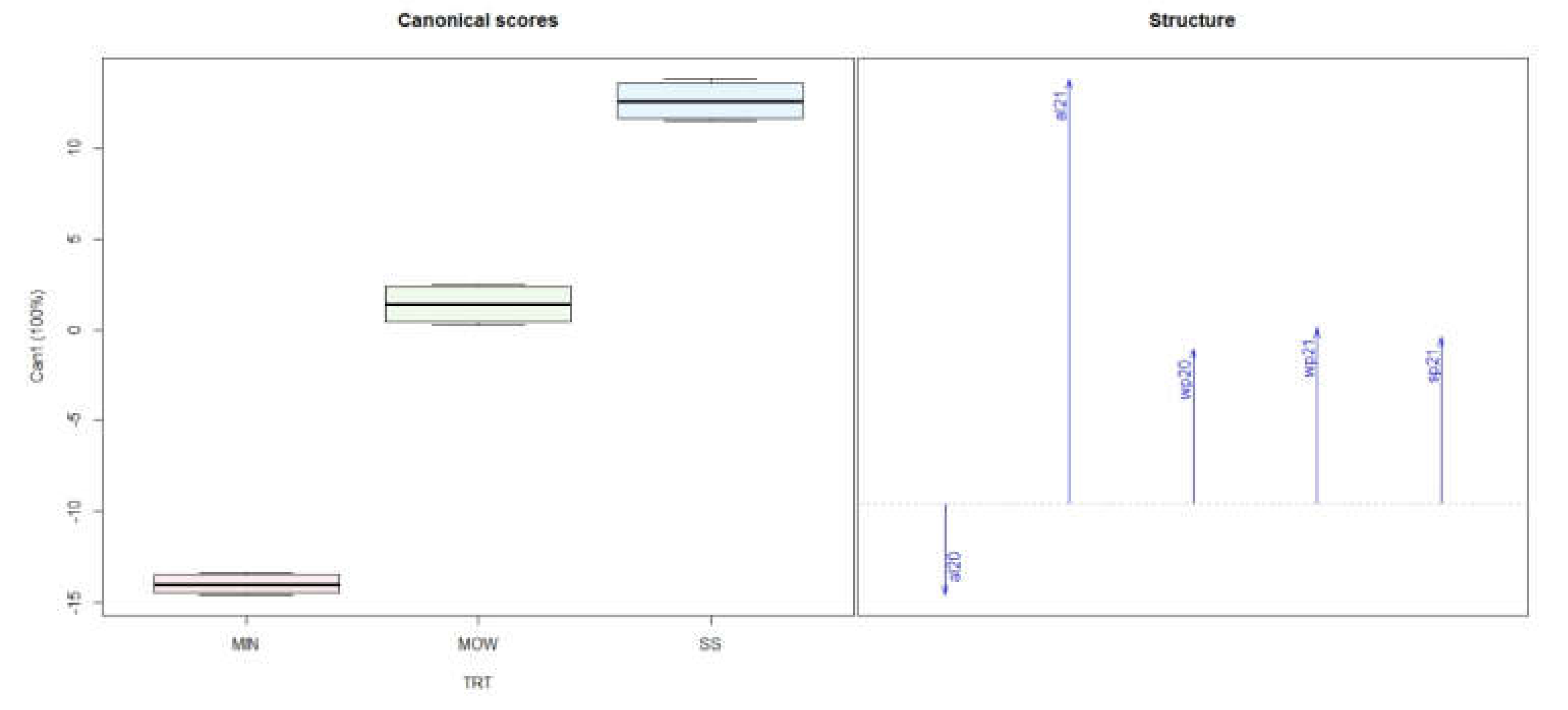
3.1. Soil Analysis
3.2. Leaf Analysis
3.3. Yield and Berry Composition
3.4. Vegetative Growth
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cerda, A.; Artola, A.; Font, X.; Barrena, R.; Gea, T.; Sánchez, A. Composting of food wastes: Status and challenges. Bioresour. Technol. 2018, 248, 57–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, J.C.; Adl, M.S.; Warman, P.R. A review of the use of composted municipal solid waste in agriculture. Agric. Ecosyst. Environ. 2008, 123, 1–14. [Google Scholar] [CrossRef]
- Ayilara, M.S.; Olanrewaju, O.S.; Babalola, O.O.; Odeyemi, O. Waste management through composting: Challenges and potentials. Sustainability 2020, 12, 4456. [Google Scholar] [CrossRef]
- Tondello, A.; Fasolo, A.; Marcato, S.; Treu, L.; Bonato, T.; Zanardi, W.; Concheri, G.; Squartini, S.; Baldan, B. Characterization of bacterial communities isolated from municipal waste compost and screening of their plant-interactive phenotypes. Sci. Total Environ. 2022, 806, 150592. [Google Scholar] [CrossRef] [PubMed]
- López-González, J.A.; Suárez-Estrella, F.; Vargas-García, M.C.; López, M.J.; Jurado, M.M.; Moreno, J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour. Technol. 2015, 175, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Costantini, E.A.; Dazzi, C. The Soils of Italy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- White, R.E.; Balachandra, L.; Edis, R.; Chen, D. The soil component of terroir. OENO One 2007, 41, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Carey, P.L.; Benge, J.R.; Haynes, R.J. Comparison of soil quality and nutrient budgets between organic and conventional kiwifruit orchards. Agric. Ecosyst. Environ. 2009, 132, 7–15. [Google Scholar] [CrossRef]
- Baldi, E.; Marcolini, G.; Quartieri, M.; Sorrenti, G.; Muzzi, E.; Toselli, M. Organic fertilization in nectarine (Prunus persica var. nucipersica) orchard combines nutrient management and pollution impact. Nutr. Cycl. Agroecosyst. 2016, 105, 39–50. [Google Scholar] [CrossRef]
- Morlat, R.; Chaussod, R. Long-term additions of organic amendments in a Loire Valley vineyard. I. Effects on properties of a calcareous sandy soil. Am. J. Enol. Vitic. 2008, 59, 353–363. [Google Scholar]
- Morlat, R.; Symoneaux, R. Long-term additions of organic amendments in a Loire Valley vineyard on a calcareous sandy soil. III. Effects on fruit composition and chemical and sensory characteristics of Cabernet franc wine. Am. J. Enol. Vitic. 2008, 59, 375–386. [Google Scholar]
- Nendel, C.; Reuter, S. Soil biology and nitrogen dynamics of vineyard soils as affected by a mature biowaste compost application. Compost Sci. Util. 2007, 15, 70–77. [Google Scholar] [CrossRef]
- Gaiotti, F.; Marcuzzo, P.; Belfiore, N.; Lovat, L.; Fornasier, F.; Tomasi, D. Influence of compost addition on soil properties, root growth and vine performances of Vitis vinifera cv Cabernet sauvignon. Sci. Hort. 2017, 225, 88–95. [Google Scholar] [CrossRef]
- Ambus, P.; Kure, L.K.; Jensen, E.S. Gross N transformation after application of household compost and domestic sewage sludge on agricultural soils. Agronomie 2002, 22, 723–730. [Google Scholar] [CrossRef]
- Mohanty, M.; Reddy, K.S.; Probert, M.E.; Dalal, R.C.; Rao, A.S.; Menzies, N.W. Modelling N mineralization from green manure and farmyard manure from a laboratory incubation study. Ecol. Model. 2011, 222, 719–726. [Google Scholar] [CrossRef]
- Masunga, R.H.; Uzokwe, V.N.; Mlay, P.D.; Odeh, I.; Singh, A.; Buchan, D.; De Neve, S. Nitrogen mineralization dynamics of different valuable organic amendments commonly used in agriculture. Appl. Soil Ecol. 2016, 101, 185–193. [Google Scholar] [CrossRef]
- Gioacchini, P.; Montecchio, D.; Gnudi, E.; Terzi, V.; Stanca, A.M.; Ciavatta, C.; Marzadori, C. Fate of N in soil amended with 15N-labelled residues of winter cereals combined with an organic N fertiliser. Soil Res. 2016, 54, 182–190. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Austr. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A. Nitrate transport, sensing, and responses in plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Verdenal, T.; Dienes-Nagy, Á.; Spangenberg, J.E.; Zufferey, V.; Spring, J.L.; Viret, O.; Carbonne, J.M.; van Leeuwen, C. Understanding and managing nitrogen nutrition in grapevine: A review. OENO One 2021, 55, 1–43. [Google Scholar] [CrossRef]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef]
- Cordovil, C.D.S.; Coutinho, J.; Goss, M.; Cabral, F. Potentially mineralizable nitrogen from organic materials applied to a sandy soil: Fitting the one-pool exponential model. Soil Use Manag. 2005, 21, 65–72. [Google Scholar] [CrossRef]
- Reeve, J.R.; Carpenter-Boggs, L.; Reganold, J.P.; York, A.L.; McGourty, G.; McCloskey, L.P. Soil and wine grape quality in biodynamically and organically managed vineyards. Am. J. Enol. Vitic. 2005, 56, 367–376. [Google Scholar]
- Downey, M.O.; Dokoozlian, N.K.; Krstic, M.P. Cultural practice and environmental impacts on the flavonoid composition of grapes and wine: A review of recent research. Am. J. Enol. Vitic. 2006, 57, 257–268. [Google Scholar]
- Möller, K. Soil fertility status and nutrient input–output flows of specialized organic cropping systems: A review. Nutr. Cycl. Agroecosyst. 2018, 112, 147–164. [Google Scholar] [CrossRef]
- Emilia-Romagna PDO and PGI Products: Created Here, Enjoyed Worldwide. Available online: https://www.regione.emilia-romagna.it/en/agriculture-and-food (accessed on 18 March 2022).
- WRB. World Reference Base for Soil Resources 2014. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports 106. FAO, Rome. Available online: http://www.fao.org/soils-portal/soil-survey/soil-classification/world-reference-base/en/ (accessed on 18 March 2022).
- Weather Forecast. Data and Observations. Available online: https://www.arpae.it/it/temi-ambientali/meteo/dati-e-osservazioni (accessed on 18 March 2022).
- Bran+Luebbe Auto Analyzer III Applications and Operation Manual, November 1998, Germany. Method No. G-109-94 Rev. 1 (multitest MT7/MT8) for NO2/NO3-N analysis and Method No. G-102-93 Rev. 1 (Multitest MT7/MT8) for NH4-N Analysis.
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Schumann, G.E.; Stanley, M.A.; Knudsen, D. Automated total nitrogen analysis of soil and plant samples. Soil Sci. Soc. Am. J. 1973, 37, 480–481. [Google Scholar] [CrossRef]
- Kingston, H.M. Microwave Assisted Acid Digestion of Siliceous and Organically-Based Matrices, Method 3052 1988 U.S. Environmental Protection Agency IAG DWI-393254-01-0, Quarterly Report, 1 January–31 March. Available online: https://www.scienceopen.com/document?vid=c835d7c2-a13d-4634-812e-58fa2a6853cd (accessed on 18 March 2022).
- Di Stefano, R.; Mattivi, F.; Carburazzi, M.; Giustini, E.; Bonifazi, L. Evoluzione della composizione fenolica dell’uva Sagrantino durante la maturazione. Riv. Vitic. Enol. 2008, 1, 39–61. [Google Scholar]
- Cruz-Castillo, J.G.; Ganeshanandam, S.; MacKay, B.R.; Lawes, G.S.; Lawoko, C.R.O.; Woolley, D.J. Applications of canonical discriminant analysis in horticultural research. HortScience 1994, 29, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Laudicina, V.A.; Badalucco, L.; Palazzolo, E. Effects of compost input and tillage intensity on soil microbial biomass and activity under Mediterranean conditions. Biol. Fert. Soil. 2011, 47, 63–70. [Google Scholar] [CrossRef]
- Nemet, F.; Perić, K.; Lončarić, Z. Microbiological activities in the composting process: A review. Columella J. Agric. Environ. Sci. 2021, 8, 41–53. [Google Scholar] [CrossRef]
- Sparling, G. The substrate induced respiration method. In Methods in Applied Soil Microbiology and Biochemistry; Academic Press: London, UK, 1995; pp. 397–404. [Google Scholar]
- Baldi, E.; Toselli, M.; Marcolini, G.; Quartieri, M.; Cirillo, E.; Innocenti, A.; Marangoni, B. Compost can successfully replace mineral fertilizers in the nutrient management of commercial peach orchard. Soil Use Manag. 2010, 26, 346–353. [Google Scholar] [CrossRef]
- Leifeld, J.; Siebert, S.; Kogel-Knabner, I. Changes in the chemical composition of soil organic matter after application of compost. Europ. J. Soil Sci. 2002, 53, 299–309. [Google Scholar] [CrossRef]
- Insam, H.; Domsch, K.H. Relationship between soil organic carbon and microbial biomass on chrono sequences of reclamation sites. Microb. Ecol. 1988, 15, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Murwira, H.K.; Kirchmann, H.; Swift, M.J. The effect of moisture on the decomposition rate of cattle manure. Plant Soil 1990, 122, 197–199. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C. Nitrate leaching in temperate agroecosystems: Sources, factors and mitigating strategies. Nutr. Cycl. Agroecosyst. 2002, 64, 237–256. [Google Scholar] [CrossRef]
- Forster, S.S.D.; Cripps, A.C.; Smith-Carington, A. Nitrate leaching to groundwater. Philosophical Transactions of the Royal Society of London. Biol. Sci. 1982, 296, 477–489. [Google Scholar]
- Holzapfel, B.P.; Treeby, M.T. Effects of timing and rate of N supply on leaf nitrogen status, grape yield and juice composition from Shiraz grapevines grafted to one of three different rootstocks. Austr. J. Grape Wine Res. 2007, 13, 14–22. [Google Scholar] [CrossRef]
- Gatti, M.; Squeri, C.; Garavani, A.; Vercesi, A.; Dosso, P.; Diti, I.; Poni, S. Effects of variable rate nitrogen application on cv. Barbera performance: Vegetative growth and leaf nutritional status. Am. J. Enol. Vitic. 2018, 69, 196–209. [Google Scholar] [CrossRef]
- Thomidis, T.; Zioziou, E.; Koundouras, S.; Karagiannidis, C.; Navrozidis, I.; Nikolaou, N. Effects of nitrogen and irrigation on the quality of grapes and the susceptibility to Botrytis bunch rot. Sci. Hort. 2016, 212, 60–68. [Google Scholar] [CrossRef]
- Winter, C.K.; Davis, S.F. Organic food. J. Food Sci. 2006, 71, 117–124. [Google Scholar] [CrossRef]
- Brandt, K.; Mølgaard, J.P. Organic agriculture: Does it enhance or reduce the nutritional value of plant foods? J. Sci. Food Agri. 2001, 81, 924–931. [Google Scholar] [CrossRef]
- Mitchell, A.E.; Chassy, A.W. Antioxidants and the nutritional quality of organic agriculture The Mitchell Lab–Phytochemicals & Health–Beyond Antioxidants. Ann. Meet. Am. Advancem. Sci. Chic. 2009, 12–16. [Google Scholar]
- Duchene, E.; Schneider, C.; Gaudillere, J.P. Effects of nitrogen nutrition timing on fruit set of grapevine, cv. Grenache. Vitis-Geilwelerhof 2011, 40, 45–46. [Google Scholar]
- Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Portu, J.; Moreno-Simunovic, Y.; Martínez-Gil, A.M. Foliar nitrogen application in Cabernet Sauvignon vines: Effects on wine flavonoid and amino acid content. Food Res. Intern. 2017, 96, 46–53. [Google Scholar] [CrossRef]
- Hilbert, G.; Soyer, J.-P.; Giraudon, J.; Milin, S.; Gaudillère, J.P. Effects of nitrogen supply on must quality and anthocyanin accumulation in berries of cv. Merlot. Vitis 2003, 42, 69–76. [Google Scholar]
- Schreiner, R.P.; Osborne, J.; Skinkis, P.A. Nitrogen requirements of Pinot noir based on growth parameters, must composition, and fermentation behavior. Am. J. Enol. Vitic. 2018, 69, 45–58. [Google Scholar] [CrossRef]
- Bravdo, B.; Hepner, Y.; Loinger, C.; Cohen, S.; Tabacman, H. Effect of crop level and crop load on growth, yield, must and wine composition, and quality of Cabernet Sauvignon. Am. J. Enol. Vitic. 1985, 36, 125–131. [Google Scholar]
- Dai, Z.W.; Ollat, N.; Gomès, E.; Decroocq, S.; Tandonnet, J.P.; Bordenave, L.; Pieri, P.; Hilbert, G.; Kappel, C.; van Leeuwen, C.; et al. Ecophysiological, genetic, and molecular causes of variation in grape berry weight and composition: A review. Am. J. Enol. Vitic. 2011, 62, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Soubeyrand, E.; Colombié, S.; Beauvoit, B.; Dai, Z.; Cluzet, S.; Hilbert, G.; Christel, R.; Christel, R.; Maneta-Peyret, L.; Dieuaide-Noubhani, M.; et al. Constraint-based modeling highlights cell energy, redox status and α-ketoglutarate availability as metabolic drivers for anthocyanin accumulation in grape cells under nitrogen limitation. Front. Plant Sci. 2018, 9, 421. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, J.I.; Rombolà, A.D. Nutrition of grapes in relation to cultivation techniques. Inf. Agrar. 2012, 68, 56–58. [Google Scholar]
- Sikora, J.L. Effect of compost-fertilizer blends on crop growth. In The Science of Composting; Bertoldi, M., Sequi, P., Lammers, B., Papi, T., Eds.; Blackie Academic & Professional Publications: Glasgow, UK, 1995; pp. 423–430. [Google Scholar]
- Pinamonti, F. Compost mulch effects on soil fertility, nutritional status and performance of grapevine. Nutr. Cycl. Agroecosyst. 1998, 51, 239–248. [Google Scholar] [CrossRef]
- Holzapfel, B.P.; Smith, J.P.; Field, S.K. Seasonal vine nutrient dynamics and distribution of Shiraz grapevines. OENO One 2019, 53, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Schreiner, R.P.; Scagel, C.F.; Baham, J. Nutrient uptake and distribution in a mature ‘Pinot Noir’ vineyard. HortScience 2006, 41, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Pradubsuk, S.; Davenport, J.R. Seasonal Uptake and Partitioning of Macronutrients in Mature ‘Concord’ Grape. J. Am. Soc. Hort. Sci. 2012, 135, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Briat, J.F.; Gojon, A.; Plassard, C.; Rouached, H.; Lemaire, G. Reappraisal of the central role of soil nutrient availability in nutrient management in light of recent advances in plant nutrition at crop and molecular levels. Europ. J. Agron. 2020, 116, 126069. [Google Scholar] [CrossRef]
- Forde, B.G. The role of long-distance signalling in plant responses to nitrate and other nutrients. J. Exp. Bot. 2002, 53, 39–43. [Google Scholar]
- Touraine, B.; Clarkson, D.T.; Muller, B. Regulation of nitrate uptake at the whole plant level. In A Whole Plant Perspective on Carbon-Nitrogen Interactions; SPB Academic Publishing: Amsterdam, The Netherlands, 1994. [Google Scholar]
- Imsande, J.; Touraine, B. N demand and the regulation of nitrate uptake. Plant Physiol. 1994, 105, 3. [Google Scholar] [CrossRef] [Green Version]
- Conradie, W.J. Seasonal uptake of nutrients by Chenin blanc in sand culture: I. Phosphorus, potassium, calcium and magnesium. S. Afr. J. Enol. Vitic. 1981, 2, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Bäßler, R.; Zou, C.; Römheld, V. Is iron phloem mobile during senescence in trees? A reinvestigation of Rissmüller’s finding of 1874. Plant Physiol. Biochem. 2011, 49, 489–493. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Characteristics | Unit | Value |
|---|---|---|
| Sand 1 | g kg−1 | 237 ± 24 |
| Silt 1 | g kg−1 | 610 ± 30 |
| Clay 1 | g kg−1 | 153 ± 5 |
| pH 1 | g kg−1 | 6.52 ± 0.110 |
| Total N 1 | g kg−1 | 0.80 ± 0.100 |
| Organic matter 1 | g kg−1 | 1.09 |
| C/N | - | 7.88 |
| Phosphorous Olsen 1 | mg kg−1 | 49.9 ± 2.3 |
| C.E.C. 1 | cmol+ kg−1 | 12.5 ± 1.10 |
| Mercury 2 (Hg) | mg kg−1 | 0.20 ± 0.02 |
| Cadmium 3 (Cd) | mg kg−1 | 0.26 ± 0.02 |
| Zinc 3 (Zn) | mg kg−1 | 76.1 ± 4.2 |
| Nickel 3 (Ni) | mg kg−1 | 43.2 ± 2.2 |
| Lead 3 (Pb) | mg kg−1 | 39.3 ± 1.3 |
| Copper 3 (Cu) | mg kg−1 | 31.3 ± 1.9 |
| Characteristics | Unit | MOW | SS | ||
|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | ||
| Dry weight 105 °C | % | 26 | 24 | 45.4 | 35.6 |
| Total organic C | % DW | 22 | 25 | 27.2 | 31.6 |
| Total N | % DW | 3.1 | 2.1 | 1.6 | 1.4 |
| C/N | 7 | 12 | 17 | 23 | |
| pH | 8.8 | 8.8 | 8.03 | 8.01 | |
| Cd | mg kg−1 DW | <0.200 | 0.63 | 0.190 | 0.24 |
| Cr6+ | mg kg−1 DW | <0.100 | 67 | <0.1 | <0.1 |
| Hg | mg kg−1 DW | 0.200 | 0.130 | 0.200 | 0.100 |
| Ni | mg kg−1 DW | 21 | 31 | 40.1 | 25.2 |
| Pb | mg kg−1 DW | 18 | 30 | 15.7 | 26.6 |
| Cu | mg kg−1 DW | 65 | 109 | 72.3 | 53.7 |
| Zn | mg kg−1 DW | 178 | 225 | 177 | 138 |
| Salmonella spp. | on 25 g wet basis | Absent | Absent | absent | absent |
| Escherichia coli | CFU | <100 | <100 | <10 | 970 |
| Treatment | chl | N | P | K | Ca | Mg | S |
|---|---|---|---|---|---|---|---|
| MIN 1 | 41.2 | 1.99 ab 3 | 0.143 | 0.474 c | 2.02 | 0.332 | 0.131 |
| MOW | 41.1 | 1.96 b | 0.153 | 0.695 a | 2.04 | 0.284 | 0.134 |
| SS | 42.1 | 2.09 a | 0.155 | 0.567 b | 1.88 | 0.306 | 0.141 |
| Significance | ns2 | *** | ns | *** | ns | ns | ns |
| Year | |||||||
| 2020 | 44.5 | 2.08 | 0.148 | 0.608 | 2.21 | 0.332 | 0.131 |
| 2021 | 38.4 | 1.95 | 0.153 | 0.549 | 1.75 | 0.282 | 0.140 |
| Significance | *** | ** | ns | ** | *** | ** | * |
| Year × treatment | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldi, E.; Polidori, G.; Germani, M.; Larocca, G.N.; Mazzon, M.; Allegro, G.; Pastore, C.; Quartieri, M.; Marzadori, C.; Filippetti, I.; et al. Fertilizer Potential of Organic-Based Soil Amendments on cv. Sangiovese (V. vinifera L.) Vines: Preliminary Results. Agronomy 2022, 12, 1604. https://doi.org/10.3390/agronomy12071604
Baldi E, Polidori G, Germani M, Larocca GN, Mazzon M, Allegro G, Pastore C, Quartieri M, Marzadori C, Filippetti I, et al. Fertilizer Potential of Organic-Based Soil Amendments on cv. Sangiovese (V. vinifera L.) Vines: Preliminary Results. Agronomy. 2022; 12(7):1604. https://doi.org/10.3390/agronomy12071604
Chicago/Turabian StyleBaldi, Elena, Greta Polidori, Margherita Germani, Greta Nicla Larocca, Martina Mazzon, Gianluca Allegro, Chiara Pastore, Maurizio Quartieri, Claudio Marzadori, Ilaria Filippetti, and et al. 2022. "Fertilizer Potential of Organic-Based Soil Amendments on cv. Sangiovese (V. vinifera L.) Vines: Preliminary Results" Agronomy 12, no. 7: 1604. https://doi.org/10.3390/agronomy12071604
APA StyleBaldi, E., Polidori, G., Germani, M., Larocca, G. N., Mazzon, M., Allegro, G., Pastore, C., Quartieri, M., Marzadori, C., Filippetti, I., Ciavatta, C., & Toselli, M. (2022). Fertilizer Potential of Organic-Based Soil Amendments on cv. Sangiovese (V. vinifera L.) Vines: Preliminary Results. Agronomy, 12(7), 1604. https://doi.org/10.3390/agronomy12071604