
Impact of Combined Drought and Heat Stress and Nitrogen on Winter Wheat Productivity and End-Use Quality

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wheat Material
2.2. Nitrogen Application
2.3. Determination of the HMW-GS Alleles at the Glu-A1 and Glu-D1 Loci
2.4. SE-HPLC Analysis
2.5. Statistical Analysis
3. Results
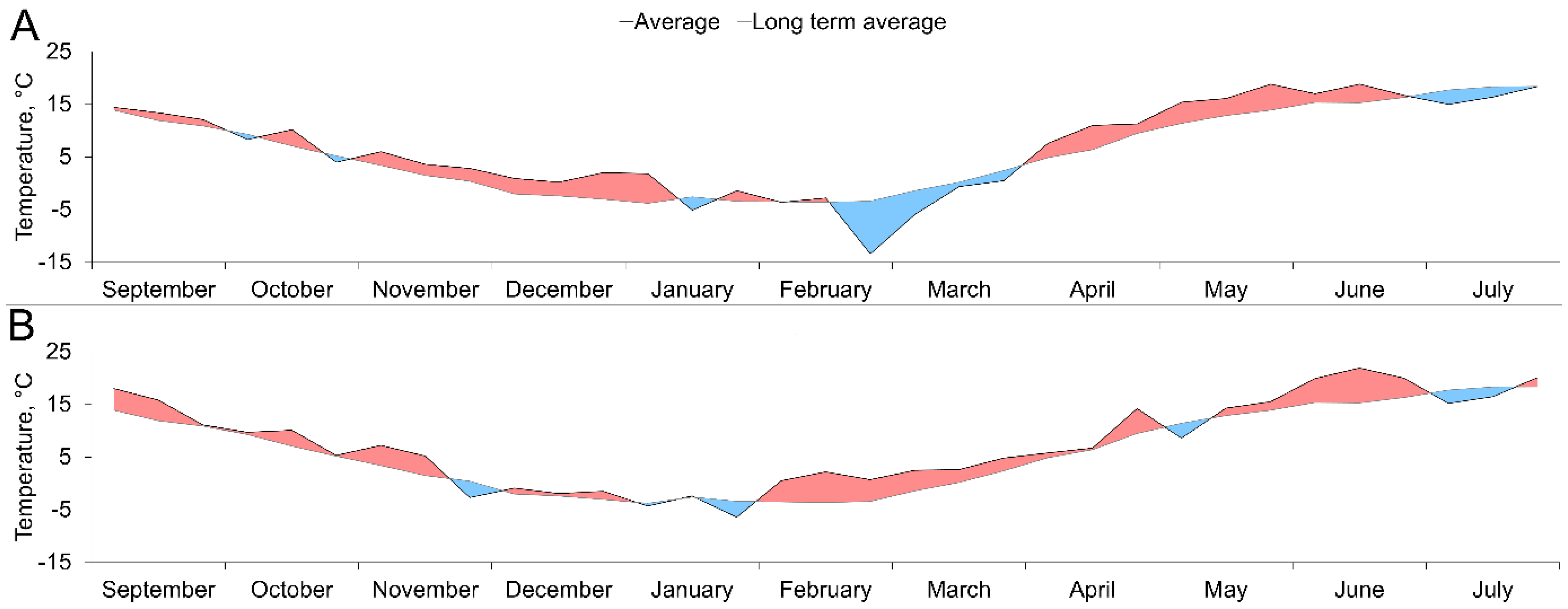
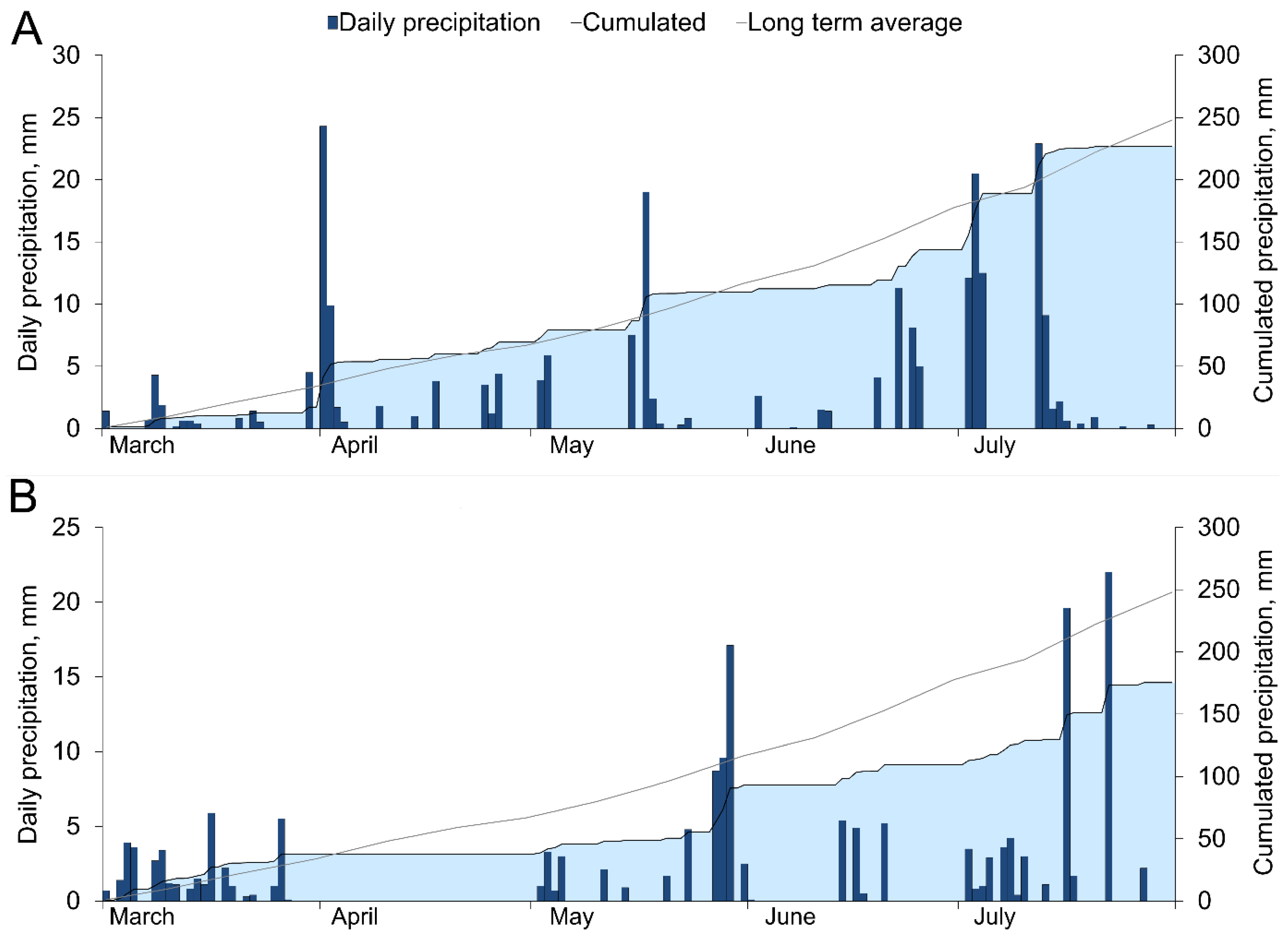
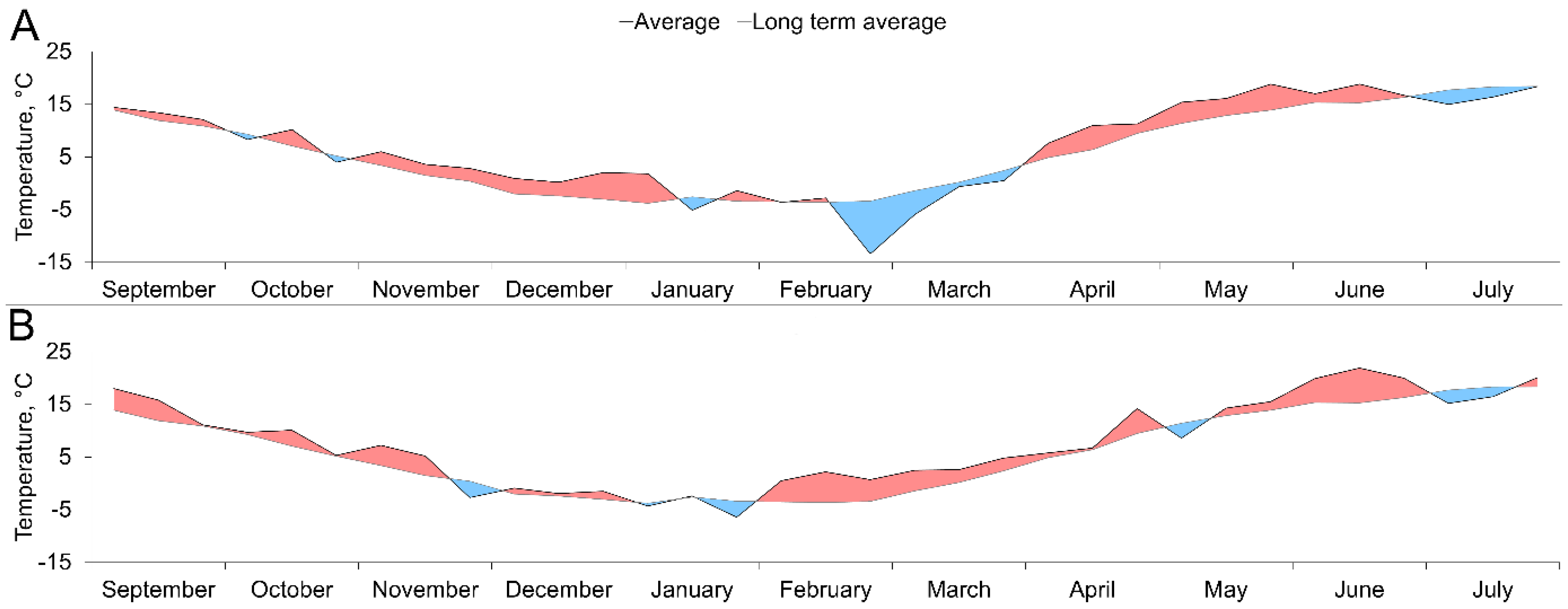
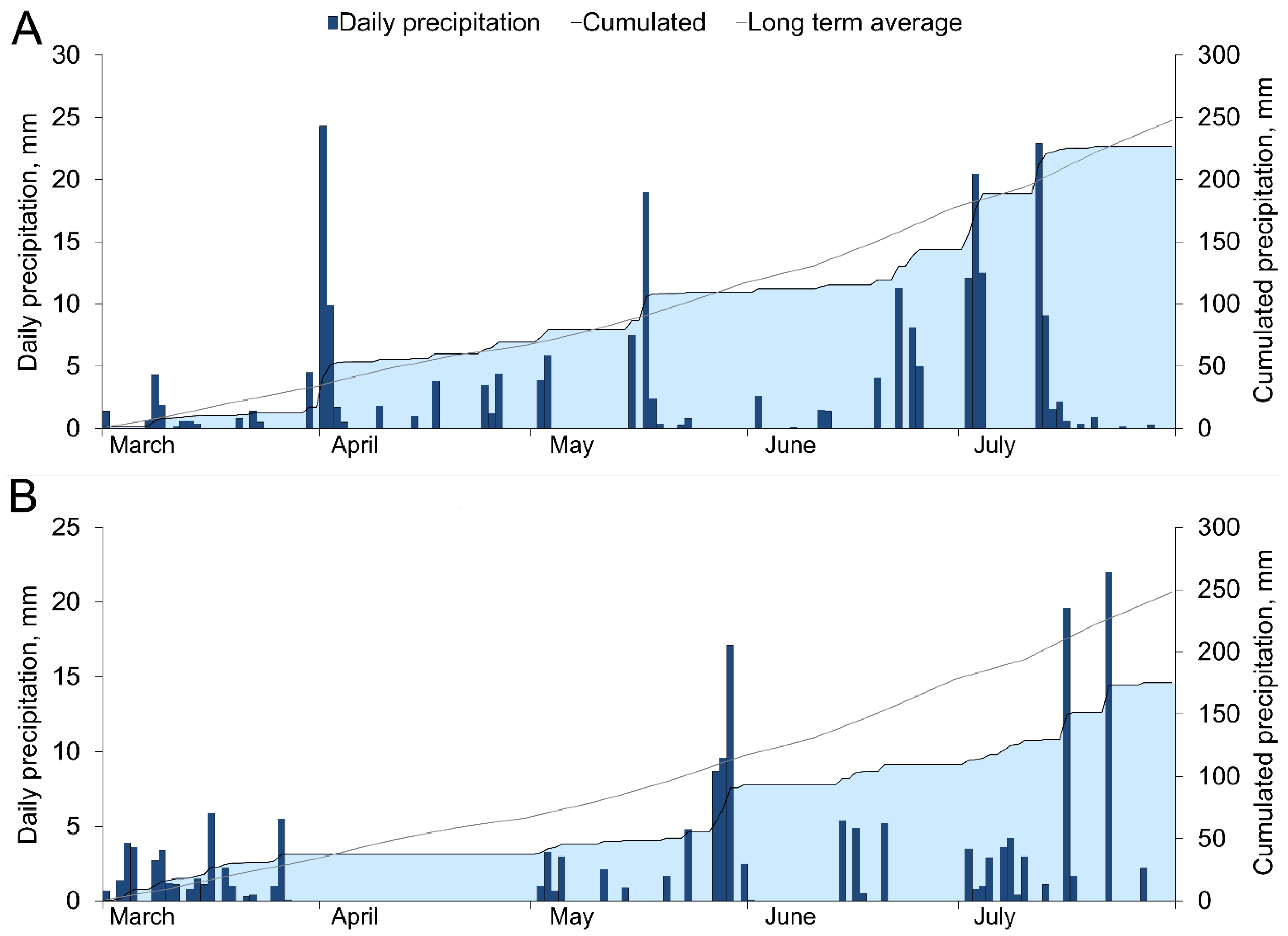
3.1. Temperature and Precipitation Variation
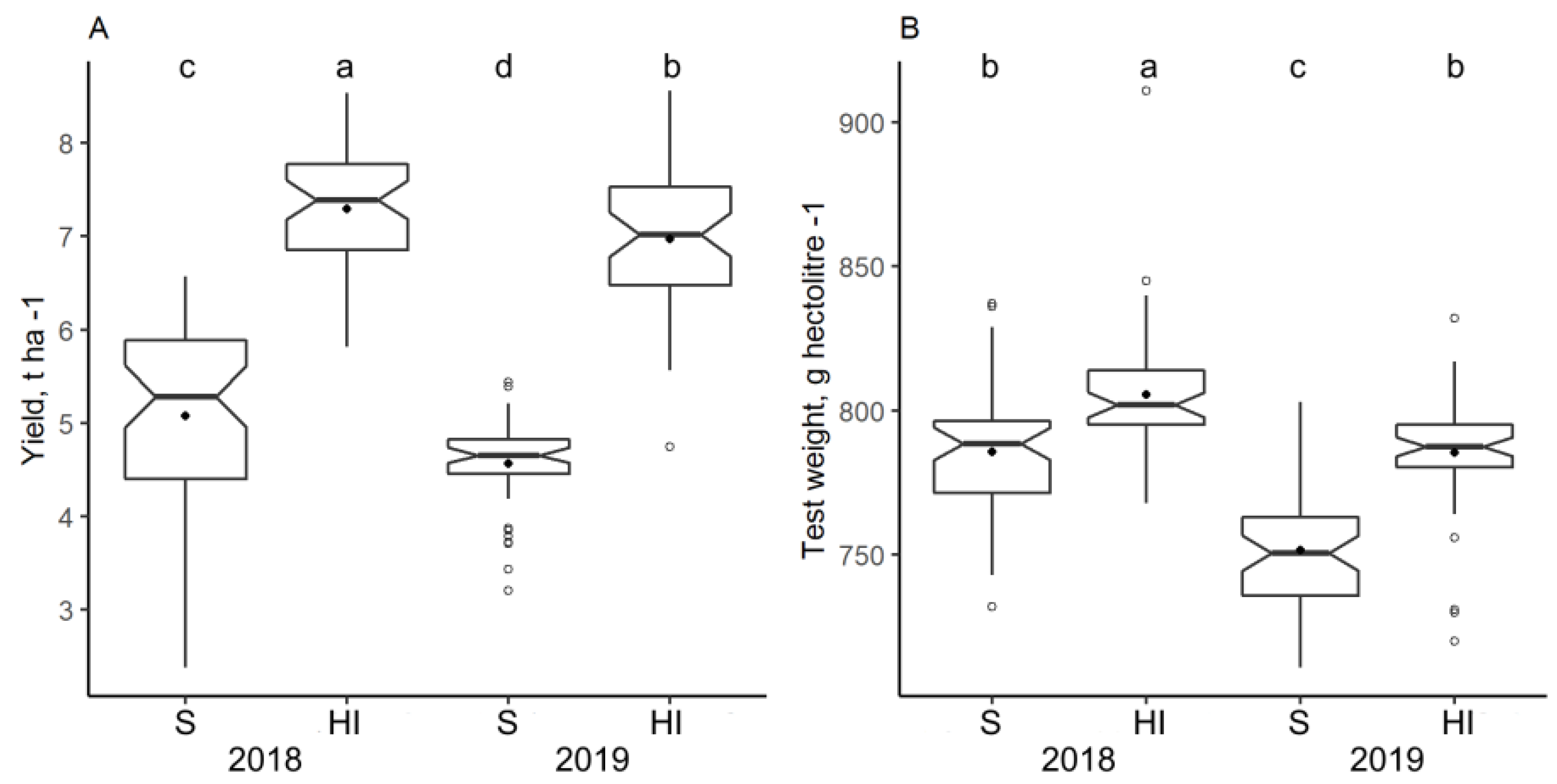
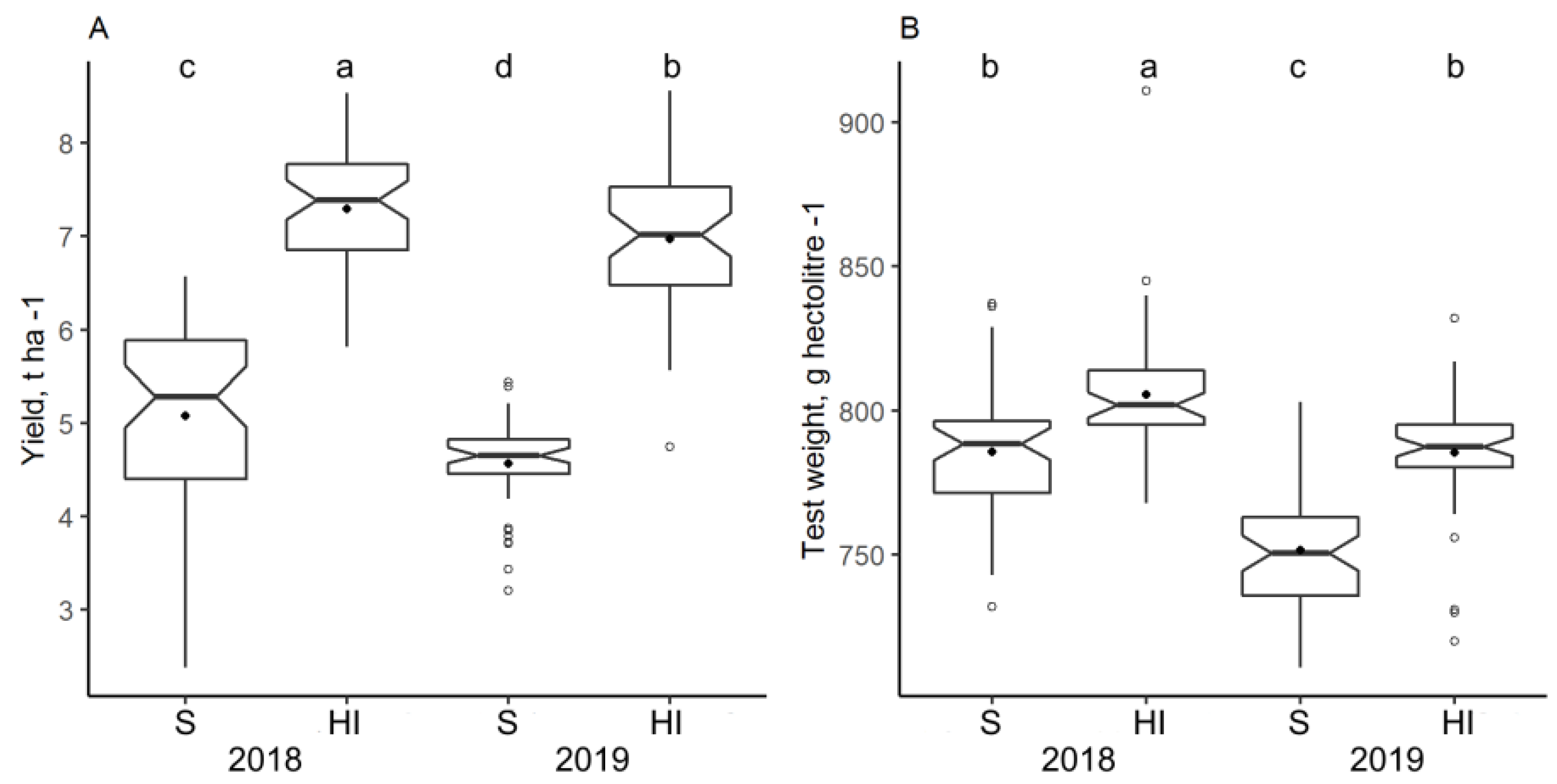
3.2. Impact of Climate and Nitrogen Application on Yield and Grain Test Weight
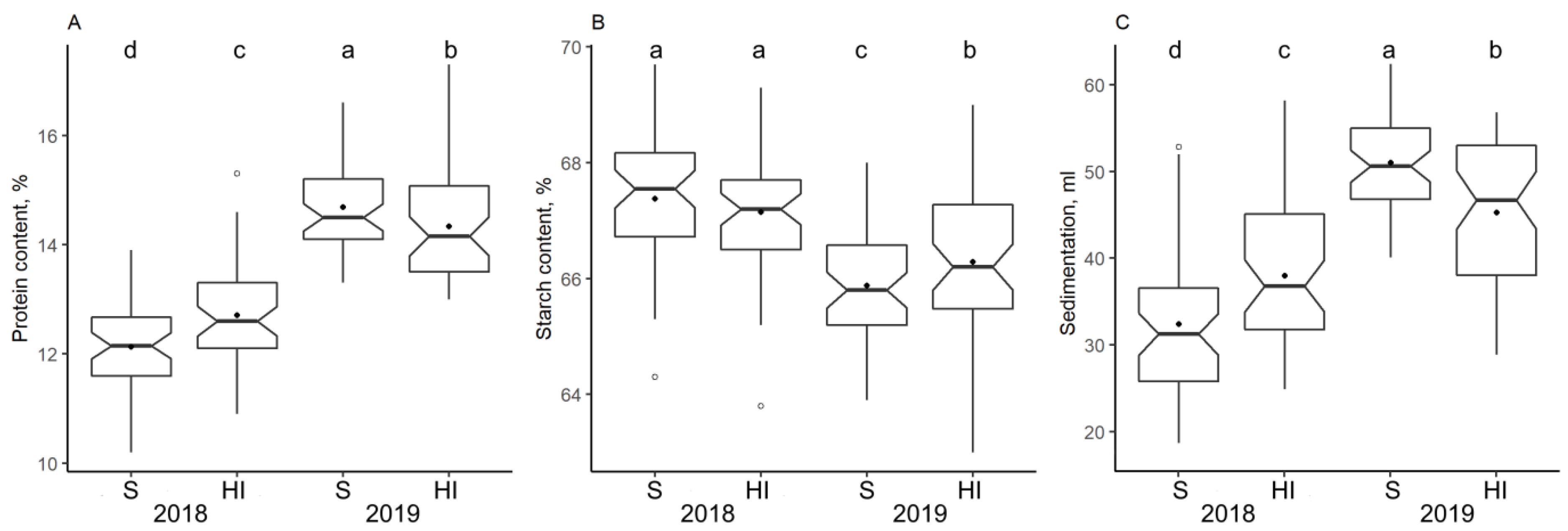
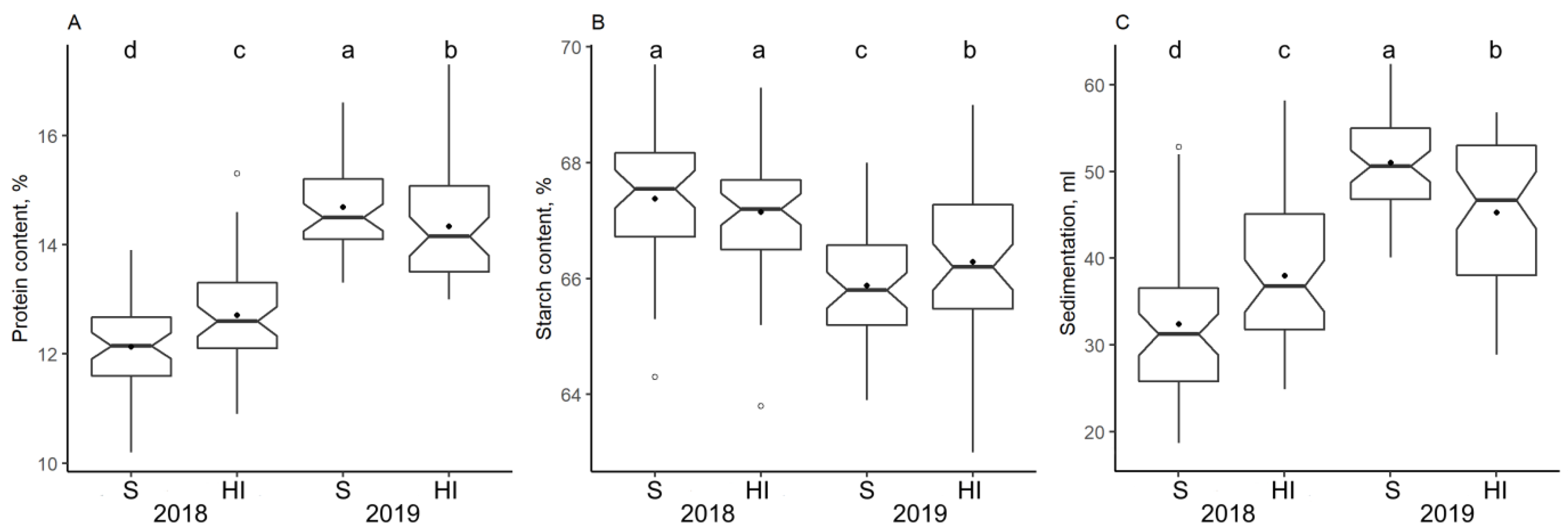
3.3. Impact of Climate and Nitrogen Application on Protein Concentration and Flour Quality Characteristics
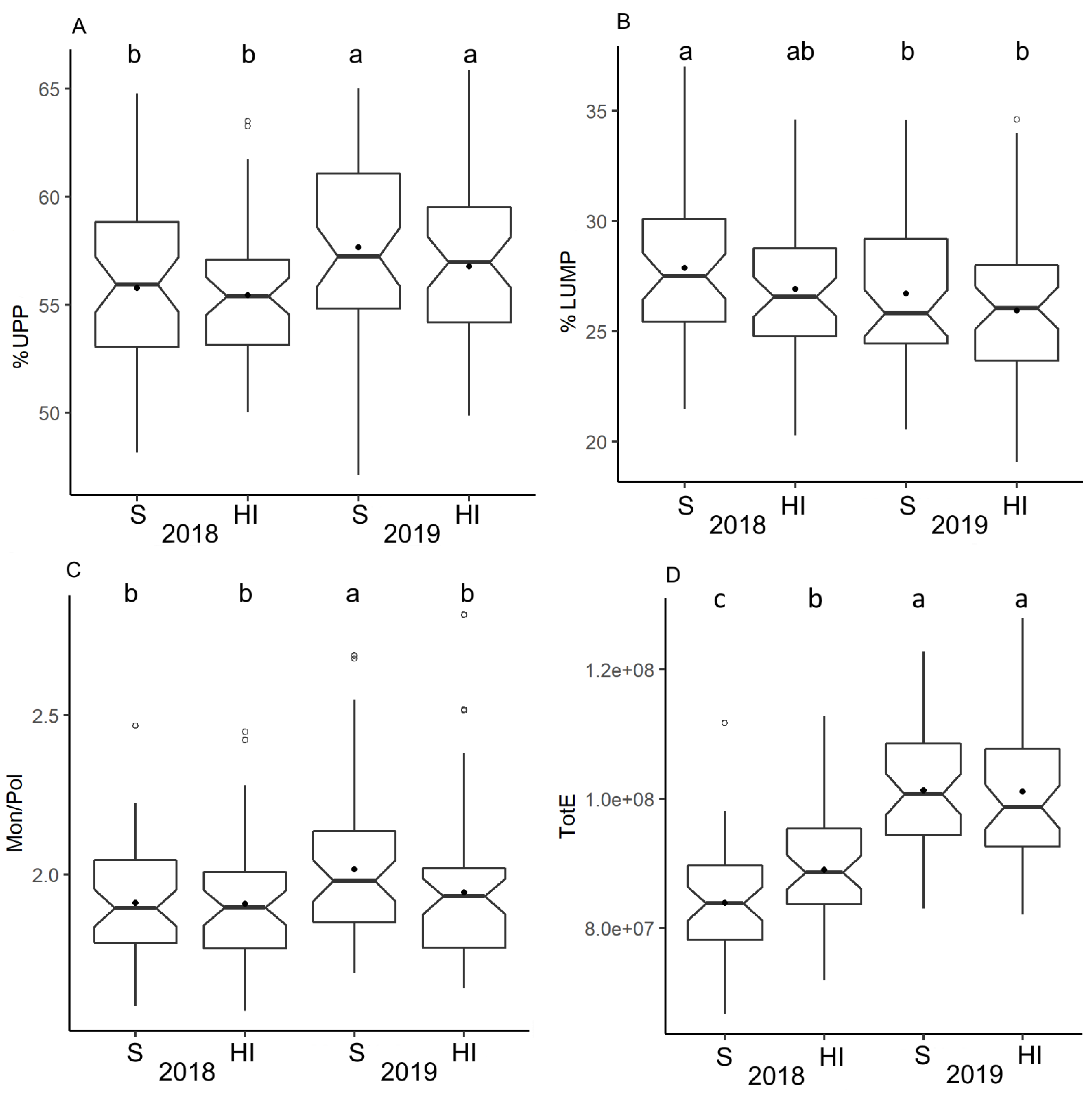
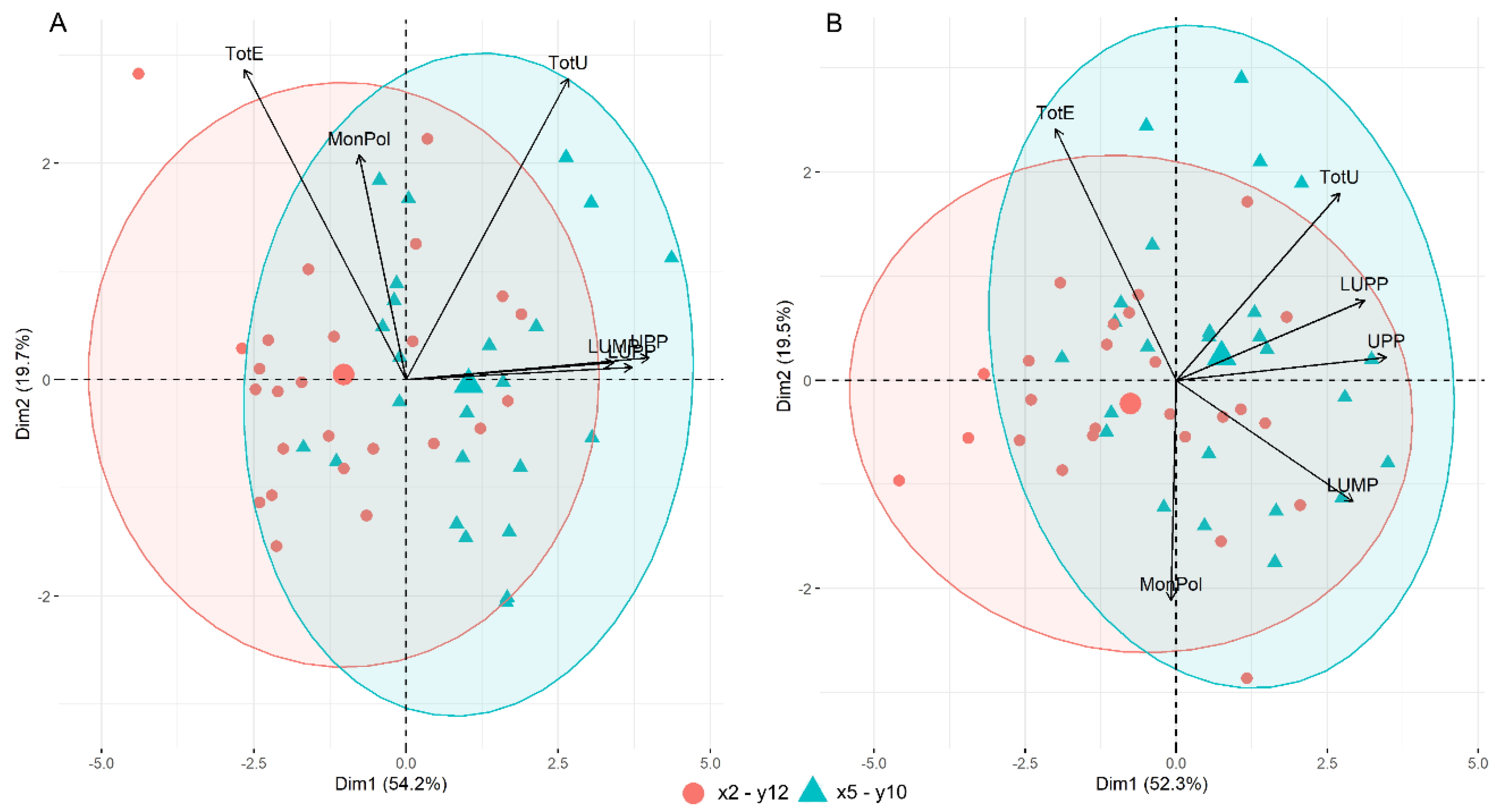
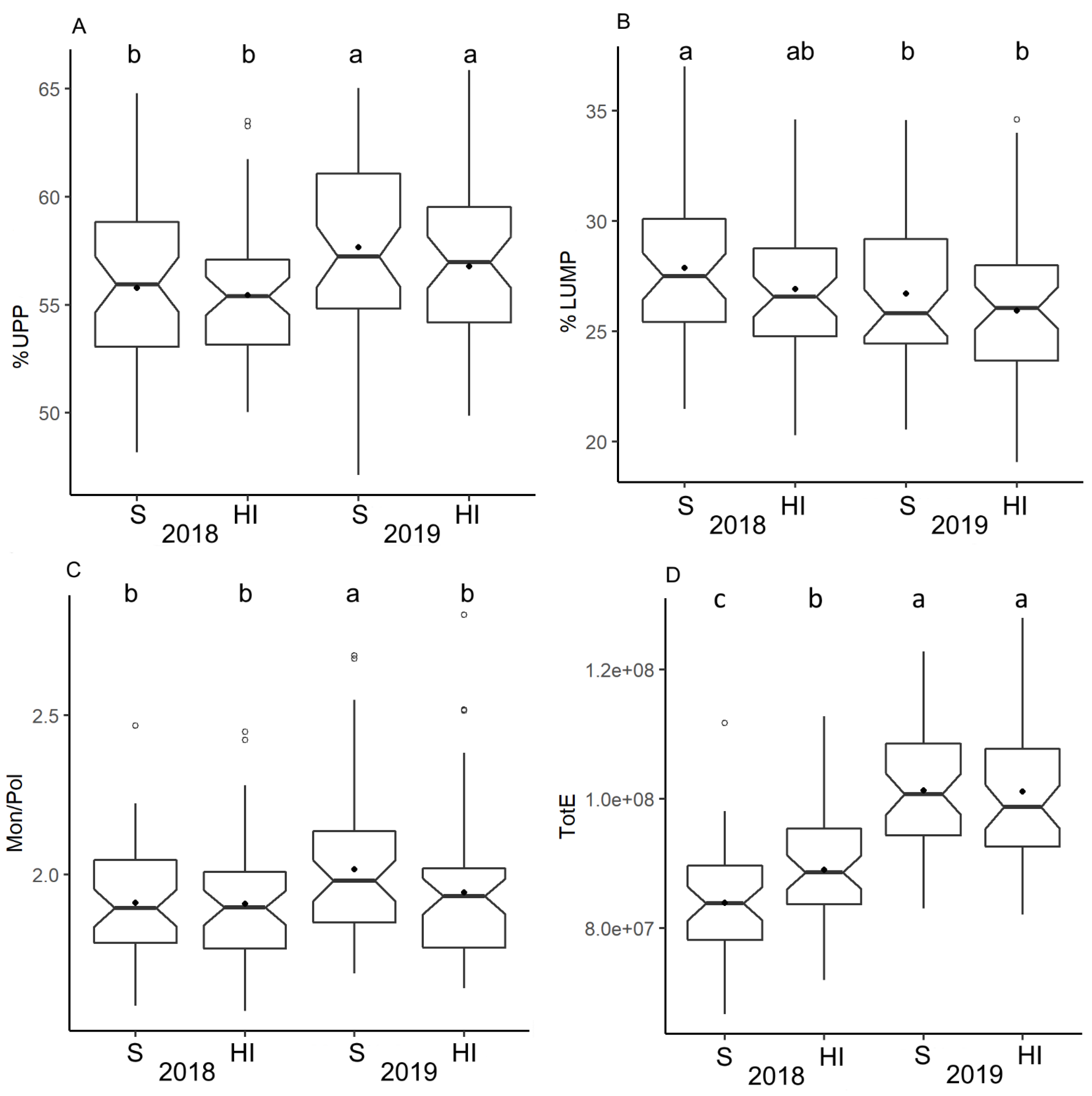
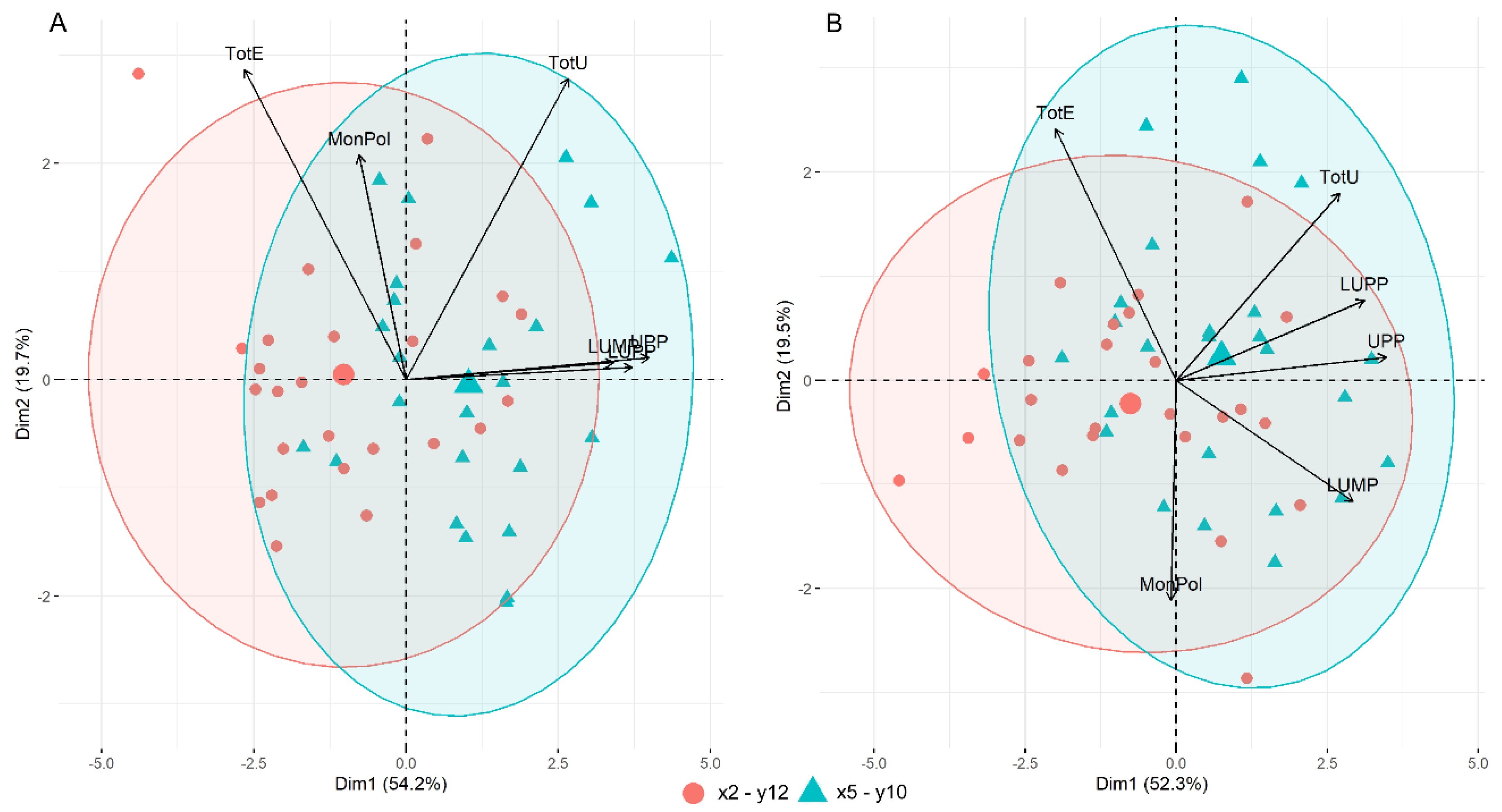
3.4. Gluten Protein Parameters by SE-HPLC
3.5. Gluten Protein Parameters and Genetic Composition of HMW-GS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lithuanian Agriculture Facts & Figures. Semiannual Statistical Report. 2021. Volume 2. Available online: https://www.vic.lt/download/faktai-ir-skaiciai-2021-m-nr-2-28/ (accessed on 30 March 2022).
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising Temperatures Reduce Global Wheat Production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Ortiz, R.; Sayre, K.D.; Govaerts, B.; Gupta, R.; Subbarao, G.V.; Ban, T.; Hodson, D.; Dixon, J.M.; Iván Ortiz-Monasterio, J.; Reynolds, M. Climate Change: Can Wheat Beat the Heat? Agric. Ecosyst. Environ. 2008, 126, 46–58. [Google Scholar] [CrossRef]
- Le Gouis, J.; Oury, F.X.; Charmet, G. How Changes in Climate and Agricultural Practices Influenced Wheat Production in Western Europe. J. Cereal Sci. 2020, 93, 102960. [Google Scholar] [CrossRef]
- Trnka, M.; Hlavinka, P.; Semenov, M.A. Adaptation Options for Wheat in Europe Will Be Limited by Increased Adverse Weather Events under Climate Change. J. R. Soc. Interface 2015, 12, 20150721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, A. Increasing Drought under Global Warming in Observations and Models. Nat. Clim. Chang. 2013, 3, 52–58. [Google Scholar] [CrossRef]
- Yadav, M.R.; Choudhary, M.; Singh, J.; Lal, M.K.; Jha, P.K.; Udawat, P.; Gupta, N.K.; Rajput, V.D.; Garg, N.K.; Maheshwari, C.; et al. Impacts, Tolerance, Adaptation, and Mitigation of Heat Stress on Wheat under Changing Climates. Int. J. Mol. Sci. 2022, 23, 2838. [Google Scholar] [CrossRef]
- Islam, M.; Begum, M.C.; Kabir, A.H.; Alam, M.F. Molecular and Biochemical Mechanisms Associated with Differential Responses to Drought Tolerance in Wheat (Triticum aestivum L.). J. Plant Interact. 2015, 10, 195–201. [Google Scholar] [CrossRef]
- Gregorová, Z.; Kováčik, J.; Klejdus, B.; Maglovski, M.; Kuna, R.; Hauptvogel, P.; Matušíková, I. Drought-Induced Responses of Physiology, Metabolites, and PR Proteins in Triticum aestivum. J. Agric. Food Chem. 2015, 63, 8125–8133. [Google Scholar] [CrossRef]
- Itam, M.; Mega, R.; Tadano, S.; Abdelrahman, M.; Matsunaga, S.; Yamasaki, Y.; Akashi, K.; Tsujimoto, H. Metabolic and Physiological Responses to Progressive Drought Stress in Bread Wheat. Sci. Rep. 2020, 10, 17189. [Google Scholar] [CrossRef] [PubMed]
- Lawas, L.M.F.; Zuther, E.; Jagadish, S.K.; Hincha, D.K. Molecular Mechanisms of Combined Heat and Drought Stress Resilience in Cereals. Curr. Opin. Plant Biol. 2018, 45, 212–217. [Google Scholar] [CrossRef]
- Senapati, N.; Stratonovitch, P.; Paul, M.J.; Semenov, M.A. Drought Tolerance during Reproductive Development Is Important for Increasing Wheat Yield Potential under Climate Change in Europe. J. Exp. Bot. 2019, 70, 2549–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous Oxide Emissions from Soils: How Well Do We Understand the Processes and Their Controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laidig, F.; Piepho, H.P.; Rentel, D.; Drobek, T.; Meyer, U.; Huesken, A. Breeding Progress, Environmental Variation and Correlation of Winter Wheat Yield and Quality Traits in German Official Variety Trials and on-Farm during 1983–2014. Theor. Appl. Genet. 2017, 130, 223–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, C.; Autrique, E.; Mondal, S.; Huerta-Espino, J.; Singh, R.P.; Vargas, M.; Crossa, J.; Amaya, A.; Peña, R.J. Genetic Improvement of Grain Quality Traits for CIMMYT Semi-Dwarf Spring Bread Wheat Varieties Developed during 1965–2015: 50 Years of Breeding. Field Crops Res. 2017, 210, 192–196. [Google Scholar] [CrossRef]
- Nehe, A.; Akin, B.; Sanal, T.; Evlice, A.K.; Ünsal, R.; Dinçer, N.; Demir, L.; Geren, H.; Sevim, I.; Orhan, Ş.; et al. Genotype x Environment Interaction and Genetic Gain for Grain Yield and Grain Quality Traits in Turkish Spring Wheat Released between 1964 and 2010. PLoS ONE 2019, 14, e0219432. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, A.; Xia, X.; Yan, Y.; Appels, R.; Mahmood, T.; He, Z. Wheat Seed Storage Proteins: Advances in Molecular Genetics, Diversity and Breeding Applications. J. Cereal Sci. 2014, 60, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, F.; Plivelic, T.S.; Kuktaite, R.; Hedenqvist, M.S.; Johansson, E. Unraveling the Structural Puzzle of the Giant Glutenin Polymer—An Interplay between Protein Polymerization, Nanomorphology, and Functional Properties in Bioplastic Films. ACS Omega 2018, 3, 5584–5592. [Google Scholar] [CrossRef]
- Branlard, G.; Dardevet, M.; Saccomano, R.; Lagoutte, F.; Gourdon, J. Genetic Diversity of Wheat Storage Proteins and Bread Wheat Quality. Euphytica 2001, 119, 59–67. [Google Scholar] [CrossRef]
- Dhaka, V.; Khatkar, B.S. Effects of Gliadin/Glutenin and HMW-GS/LMW-GS Ratio on Dough Rheological Properties and Bread-Making Potential of Wheat Varieties. J. Food Qual. 2015, 38, 71–82. [Google Scholar] [CrossRef]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Prieto-Linde, M.L.; Johansson, E. Towards the Understanding of Bread-Making Quality in Organically Grown Wheat: Dough Mixing Behaviour, Protein Polymerisation and Structural Properties. J. Cereal Sci. 2012, 56, 659–666. [Google Scholar] [CrossRef]
- Ceresino, E.B.; Kuktaite, R.; Hedenqvist, M.S.; Sato, H.H.; Johansson, E. Processing Conditions and Transglutaminase Sources to “Drive” the Wheat Gluten Dough Quality. Innov. Food Sci. Emerg. Technol. 2020, 65, 102439. [Google Scholar] [CrossRef]
- Lama, S.; Vallenback, P.; Hall, S.A.; Kuzmenkova, M.; Kuktaite, R. Prolonged Heat and Drought versus Cool Climate on the Swedish Spring Wheat Breeding Lines: Impact on the Gluten Protein Quality and Grain Microstructure. Food Energy Secur. 2022, 11, e376. [Google Scholar] [CrossRef]
- Lafiandra, D.; Tucci, G.F.; Pavoni, A.; Turchetta, T.; Margiotta, B. PCR Analysis of X- and y-Type Genes Present at the Complex Glu-A1 Locus in Durum and Bread Wheat. Theor. Appl. Genet. 1997, 94, 235–240. [Google Scholar] [CrossRef]
- Ma, W.; Zhang, W.; Gale, K.R. Multiplex-PCR Typing of High Molecular Weight Glutenin Alleles in Wheat. Euphytica 2003, 134, 51–60. [Google Scholar] [CrossRef]
- De Bustos, A.; Rubio, P.; Jouve, N. Molecular Characterisation of the Inactive Allele of the Gene Glu-A1 and the Development of a Set of AS-PCR Markers for HMW Glutenins of Wheat. Theor. Appl. Genet. 2000, 100, 1085–1094. [Google Scholar] [CrossRef]
- Ishikawa, G.; Nakamura, T. A New Co-Dominant PCR-Based Marker to Identify the High-Molecular-Weight Glutenin Subunit Combination. Wheat Inf. Servise 2007, 103, 1–4. [Google Scholar]
- Smith, R.L.; Schweder, M.E.; Barnett, R.D. Identification of Glutenin Alleles in Wheat and Triticale Using PCR-Generated DNA Markers. Crop Sci. 1994, 34, 1373–1378. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 22 June 2020).
- Olivoto, T.; Lúcio, A.D.C. Metan: An R Package for Multi-Environment Trial Analysis. Methods Ecol. Evol. 2020, 2020, 783–789. [Google Scholar] [CrossRef]
- Lüdecke, D. Sjstats: Statistical Functions for Regression Models (Version 0.18.0). 2020. Available online: https://zenodo.org/record/1489175#.YqvmLOxBzIU (accessed on 20 May 2020). [CrossRef]
- de Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research 2020. Available online: https://cran.r-project.org/package=agricolae (accessed on 20 May 2020).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A Meta-Analysis of Crop Yield under Climate Change and Adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef]
- Baruth, B.; Bassu, S.; Bussay, A.; Cerrani, I.; Chemin, Y.; De Palma, P.; Fumagalli, D.; Lecerf, R.; Manfron, G.; Nisini Scacchiafichi, L.; et al. JRC MARS Bulletin—Crop Monitoring in Europe; Van Den Berg, M., Baruth, B., Niemeyer, S., Chemin, Y., Eds.; Publications Office of the European Union: Luxembourg, 2019; Volume 27, p. 44. [Google Scholar]
- Bussay, A.; Bassu, S.; Ceglar, A.; Cerrani, I.; Garcia Condado, S.; Lecerf, R.; Lopez Lozano, R.; Manfron, G.; Nisini Scacchiafichi, L.; Panarello, L.; et al. JRC MARS Bulletin—Crop Monitoring in Europe; Van Den Berg, M., Baruth, B., Van Der Velde, M., Niemeyer, S., Eds.; Publications Office of the European Union: Luxembourg, 2018; Volume 26, p. 48. [Google Scholar]
- Farooq, M.; Hussain, M.; Siddique, K.H.M. Drought Stress in Wheat during Flowering and Grain-Filling Periods. CRC. Crit. Rev. Plant Sci. 2014, 33, 331–349. [Google Scholar] [CrossRef]
- Balla, K.; Rakszegi, M.; Li, Z.; Békés, F.; Bencze, S.; Veisz, O. Quality of Winter Wheat in Relation to Heat and Drought Shock after Anthesis. Czech J. Food Sci. 2011, 29, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.V.V.; Djanaguiraman, M. Response of Floret Fertility and Individual Grain Weight of Wheat to High Temperature Stress: Sensitive Stages and Thresholds for Temperature and Duration. Funct. Plant Biol. 2014, 41, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Onyemaobi, I.; Liu, H.; Siddique, K.H.M.; Yan, G. Both Male and Female Malfunction Contributes to Yield Reduction under Water Stress during Meiosis in Bread Wheat. Front. Plant Sci. 2017, 7, 2071. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Liu, L.; Asseng, S.; Zhang, D.; Ma, W.; Tang, L.; Cao, W.; Zhu, Y. Modelling the Effects of Post-Heading Heat Stress on Biomass Partitioning, and Grain Number and Weight of Wheat. J. Exp. Bot. 2020, 71, 6015–6031. [Google Scholar] [CrossRef]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; Sabagh, A.E.L. Evaluation of Fourteen Bread Wheat (Triticum aestivum L.) Genotypes by Observing Gas Exchange Parameters, Relative Water and Chlorophyll Content, and Yield Attributes under Drought Stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Urban, O.; Hlaváčová, M.; Klem, K.; Novotná, K.; Rapantová, B.; Smutná, P.; Horáková, V.; Hlavinka, P.; Škarpa, P.; Trnka, M. Combined Effects of Drought and High Temperature on Photosynthetic Characteristics in Four Winter Wheat Genotypes. Field Crops Res. 2018, 223, 137–149. [Google Scholar] [CrossRef]
- Hlaváčová, M.; Klem, K.; Rapantová, B.; Novotná, K.; Urban, O.; Hlavinka, P.; Smutná, P.; Horáková, V.; Škarpa, P.; Pohanková, E.; et al. Interactive Effects of High Temperature and Drought Stress during Stem Elongation, Anthesis and Early Grain Filling on the Yield Formation and Photosynthesis of Winter Wheat. Field Crops Res. 2018, 221, 182–195. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of Diverse Wheat (Triticum aestivum L.) Genotypes Varying in Sensitivity to Heat and Drought Stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozbicki, J.; Ceglińska, A.; Gozdowski, D.; Jakubczak, M.; Cacak-Pietrzak, G.; Madry, W.; Golba, J.; Piechociński, M.; Sobczyński, G.; Studnicki, M.; et al. Influence of the Cultivar, Environment and Management on the Grain Yield and Bread-Making Quality in Winter Wheat. J. Cereal Sci. 2015, 61, 126–132. [Google Scholar] [CrossRef]
- Šíp, V.; Vavera, R.; Chrpová, J.; Kusá, H.; Růžek, P. Winter Wheat Yield and Quality Related to Tillage Practice, Input Level and Environmental Conditions. Soil Tillage Res. 2013, 132, 77–85. [Google Scholar] [CrossRef]
- Gagliardi, A.; Carucci, F.; Masci, S.; Flagella, Z.; Gatta, G.; Giuliani, M.M. Effects of Genotype, Growing Season and Nitrogen Level on Gluten Protein Assembly of Durum Wheat Grown under Mediterranean Conditions. Agronomy 2020, 10, 755. [Google Scholar] [CrossRef]
- Ozturk, A.; Erdem, E.; Aydin, M.; Karaoglu, M.M. The Effects of Drought after Anthesis on the Grain Quality of Bread Wheat Depend on Drought Severity and Drought Resistance of the Variety. Cereal Res. Commun. 2022, 50, 105–116. [Google Scholar] [CrossRef]
- Malalgoda, M.; Ohm, J.B.; Meinhardt, S.; Simsek, S. Association between Gluten Protein Composition and Breadmaking Quality Characteristics in Historical and Modern Spring Wheat. Cereal Chem. 2018, 95, 226–238. [Google Scholar] [CrossRef]
- Anjum, F.M.; Khan, M.R.; Din, A.; Saeed, M.; Pasha, I.; Arshad, M.U. Wheat Gluten: High Molecular Weight Glutenin Subunits—Structure, Genetics, and Relation to Dough Elasticity. J. Food Sci. 2007, 72, R56–R63. [Google Scholar] [CrossRef]
- He, Z.; Joshi, A.K.; Zhang, W. Climate Vulnerabilities and Wheat Production. In Climate Vulnerability; Pielke, R.A.B.T.-C.V., Ed.; Academic Press: Oxford, UK, 2013; pp. 57–67. ISBN 978-0-12-384704-1. [Google Scholar]
- Thungo, Z.; Shimelis, H.; Odindo, A.; Mashilo, J. Genotype-by-Environment Effects on Grain Quality among Heat and Drought Tolerant Bread Wheat (Triticum aestivum L.) Genotypes. J. Plant Interact. 2020, 15, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Tomás, D.; Viegas, W.; Silva, M. Effects of Post-Anthesis Heat Waves on the Grain Quality of Seven European Wheat Varieties. Agronomy 2020, 10, 268. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, D.; Berger, A.; Prieto-Linde, M.L.; Johansson, E. Can Nitrogen Fertilization Be Used to Modulate Yield, Protein Content and Bread-Making Quality in Uruguayan Wheat? J. Cereal Sci. 2019, 85, 153–161. [Google Scholar] [CrossRef]
- Labuschagne, M.T.; Moloi, J.; van Biljon, A. Abiotic Stress Induced Changes in Protein Quality and Quantity of Two Bread Wheat Cultivars. J. Cereal Sci. 2016, 69, 259–263. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined Effect of Genetic and Environmental Factors on the Accumulation of Proteins in the Wheat Grain and Their Relationship to Bread-Making Quality. J. Cereal Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- He, Z.H.; Liu, L.; Xia, X.C.; Liu, J.J.; Peña, R.J. Composition of HMW and LMW Glutenin Subunits and Their Effects on Dough Properties, Pan Bread, and Noodle Quality of Chinese Bread Wheats. Cereal Chem. 2005, 82, 345–350. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Gene/Allele | Product Size (bp) | Reference |
|---|---|---|---|
| Glu-A1 | Ax null | 920 | Lafiandra et al. [25] |
| Ax1 or Ax2 * | 1500 or 1400 | Lafiandra et al. [25] | |
| Ax2 * | 1319 | Ma et al. [26] | |
| Ax2 * | 2650 | De Bustos et al. [27] | |
| Glu-D1 | Dx5 | 478 | Ma et al. [26] |
| Dx5-y10 | 343 + 320, other alleles 361 | Ishikawa and Nakamura [28] | |
| Dy10 or Dy12 | 576 or 612 | Smith et al. [29] |
| Variable | Genotype | Year | Nitrogen | Year × Nitrogen |
|---|---|---|---|---|
| Yield | 0.06 | 0.02 | 0.63 | n.s. |
| Test weight | 0.22 | 0.23 | 0.21 | n.s. |
| Protein content | 0.30 | 0.57 | n.s. | 0.03 |
| Starch content | 0.57 | 0.22 | n.s. | 0.02 |
| Sedimentation volume | 0.40 | 0.34 | n.s. | 0.07 |
| %UPP | 0.74 | 0.04 | 0.01 | n.s. |
| %LUPP | 0.71 | 0.04 | n.s | n.s. |
| %LUMP | 0.59 | 0.02 | 0.02 | n.s. |
| Mon/Pol ratio | 0.81 | 0.03 | 0.01 | 0.01 |
| TotU | 0.42 | 0.34 | n.s. | 0.01 |
| TotE | 0.47 | 0.37 | 0.01 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Statkevičiūtė, G.; Liatukas, Ž.; Cesevičienė, J.; Jaškūnė, K.; Armonienė, R.; Kuktaite, R.; Brazauskas, G. Impact of Combined Drought and Heat Stress and Nitrogen on Winter Wheat Productivity and End-Use Quality. Agronomy 2022, 12, 1452. https://doi.org/10.3390/agronomy12061452
Statkevičiūtė G, Liatukas Ž, Cesevičienė J, Jaškūnė K, Armonienė R, Kuktaite R, Brazauskas G. Impact of Combined Drought and Heat Stress and Nitrogen on Winter Wheat Productivity and End-Use Quality. Agronomy. 2022; 12(6):1452. https://doi.org/10.3390/agronomy12061452
Chicago/Turabian StyleStatkevičiūtė, Gražina, Žilvinas Liatukas, Jurgita Cesevičienė, Kristina Jaškūnė, Rita Armonienė, Ramune Kuktaite, and Gintaras Brazauskas. 2022. "Impact of Combined Drought and Heat Stress and Nitrogen on Winter Wheat Productivity and End-Use Quality" Agronomy 12, no. 6: 1452. https://doi.org/10.3390/agronomy12061452
APA StyleStatkevičiūtė, G., Liatukas, Ž., Cesevičienė, J., Jaškūnė, K., Armonienė, R., Kuktaite, R., & Brazauskas, G. (2022). Impact of Combined Drought and Heat Stress and Nitrogen on Winter Wheat Productivity and End-Use Quality. Agronomy, 12(6), 1452. https://doi.org/10.3390/agronomy12061452