Biological Response of Triticum aestivum L. to the Abiotic Stress Induced by Winemaking Waste


Abstract
1. Introduction
- (a)
- the phytotoxic effect of grape marc extracts on wheat; grains;
- (b)
- the genotoxic potential of grape marc extracts on wheat cells.
2. Materials and Methods
2.1. Extracts Preparation
2.2. Experimental Procedure
2.3. Evaluation of Germination Rate
2.4. Cytogenetic Investigations
- (1)
- MI (%) = number of dividing cells/total cells (dividing and nondividing) × 100;
- (2a)
- cells in prophase (%) = number of prophase cells/total cells (dividing and nondividing) × 100;
- (2b)
- cells in metaphase (%) = number of metaphase cells/total cells (dividing and nondividing) × 100;
- (2c)
- cells in anaphase (%) = number of anaphase cells /total cells (dividing and nondividing) × 100;
- (2d)
- cells in telophase (%) = number of telophase cells/total cells (dividing and nondividing) × 100;
- (3a)
- cells with chromosomal aberrations from ana-telophases (%) = number of cells in aberrant ana-telophases/total cells (dividing and nondividing) × 100;
- (3b)
- cells with aberrant interphases (%) = number of cells with aberrant interphases/total cells (dividing and nondividing) × 100;
- (4)
- cells with different types of genetic abnormalities from total cells (%) = number of cells with a certain type of genetic abnormality/total cells (dividing and nondividing) × 100.
2.5. Statistical Analysis
3. Results
3.1. Germination Rate of Triticum aestivum Grains
3.2. Cytogenetic Investigations
3.2.1. Mitotic Index
3.2.2. Frequency of Mitotic Phases
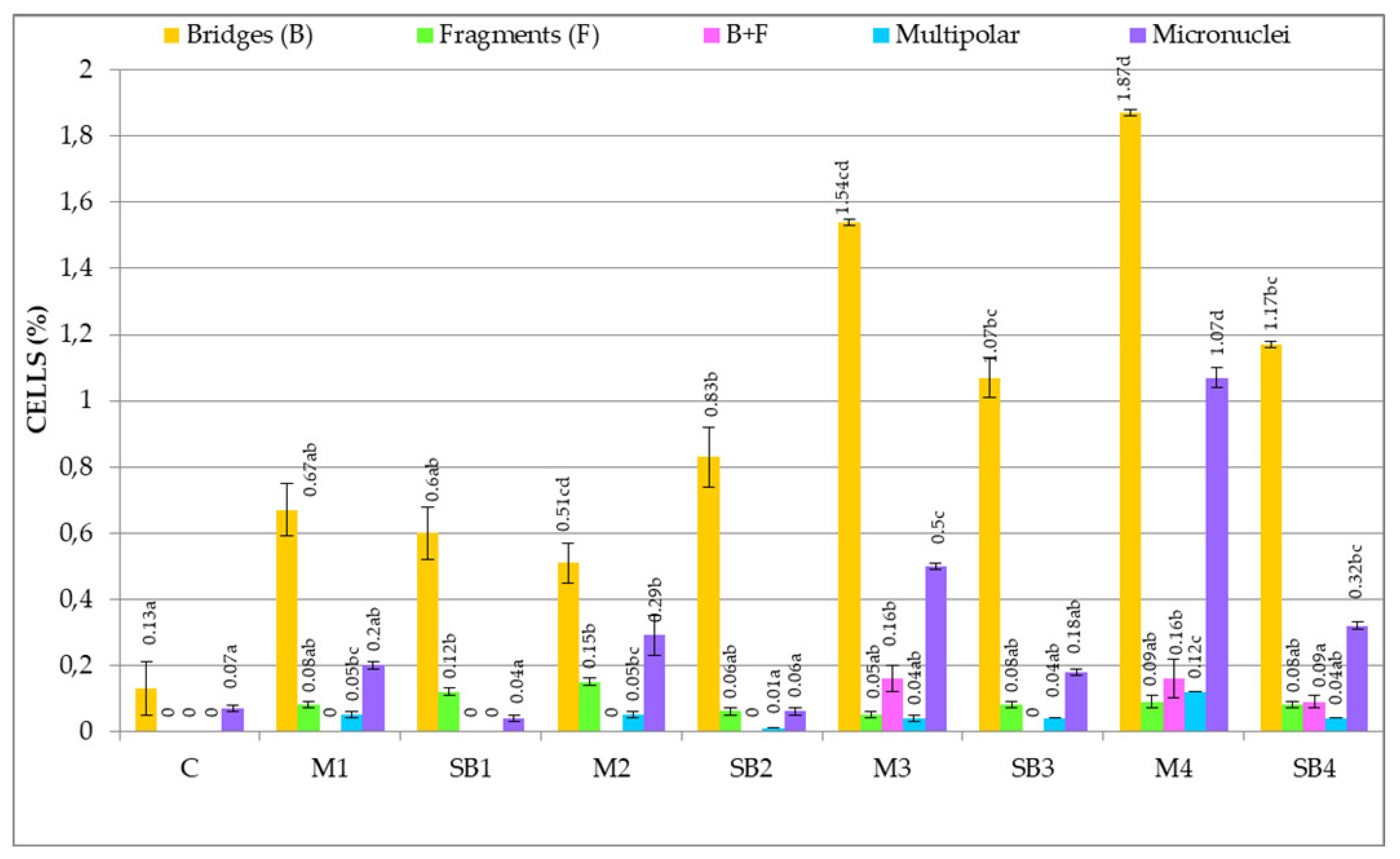
3.2.3. Genetic Abnormalities
4. Discussion
4.1. Germination Rate of Triticum aestivum Grains
4.2. Mitotic Index
4.3. Frequency of Mitotic Phases
4.4. Genetic Abnormalities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spigno, G.; Marinoni, L.; Garrido, G.D. State of the art in grape processing by-products. In Handbook of Grape Processing By-Products; Galanakis, C., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 1–27. [Google Scholar]
- Dávila, I.; Robles, E.; Egüés, I.; Labidi, J.; Gullón, P. The biorefinery concept for the industrial valorization of grape processing by-products. In Handbook of Grape Processing By-Products; Galanakis, C., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 29–53. [Google Scholar]
- Tylewicz, U.; Nowacka, M.; Martín-García, B.; Wiktor, A.; Caravaca, A.M.G. Target sources of polyphenols in different food products and their processing by-products. In Polyphenols: Properties, Recovery and Applications; Galanakis, C., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 135–175. [Google Scholar]
- Cristea, E.; Sturza, R.; Jauregi, P.; Niculaua, M.; Ghendov Moșanu, A.; Patras, A. Influence of pH and ionic strength on the color parameters and antioxidant properties of an ethanolic red grape marc extract. J. Food Biochem. 2019, 43, e12788. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic content s of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, V.; Brandelli, A.; Marczak, L.D.F.; Tessaro, I.C. Kinetic modeling of total polyphenol extraction from grape marc and characterization of the extracts. Sep. Purif. Technol. 2012, 100, 82–87. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Friant, P.; Chone, X.; Tregoat, O.; Koundouras, S.; Dubourdieu, D. Influence of climate, soil, and cultivar on terroir. Am. J. Enol. Vitic. 2004, 55, 207–217. [Google Scholar]
- Pereira, G.E.; Gaudillere, J.P.; van Leeuwen, C.; Hilbert, G.; Maucourt, M.; Deborde, C.; Moing, A.; Rolin, D. 1H NMR metabolite fingerprints of grape berry: Comparison of vintage and soil effects in Bordeaux grapevine growing areas. Anal. Chim. Acta 2006, 563, 346–352. [Google Scholar] [CrossRef]
- Alibante, A.; Lakka, A.; Bozinou, E.; Chatzilazarou, A.; Lalas, S.; Makris, D.P. Integrated Green Process for the Extraction of Red Grape Pomace Antioxidant Polyphenols Using Ultrasound-Assisted Pretreatment and beta-Cyclodextrin. Beverages 2021, 7, 59. [Google Scholar] [CrossRef]
- Cristea, E.; Sturza, R.; Patras, A. The influence of temperature and time on the stability of the antioxidant activity and colour parameters of grape marc ethanolic extract. Food Technol. 2015, 39, 96–104. [Google Scholar]
- Tanase, C.; Bujor, O.C.; Popa, V.I. Phenolic natural compounds and their influence on physiological processes in plants. In Polyphenols in Plants, 2nd ed.; Academic Press: Bucharest, Romania, 2019; pp. 45–58. [Google Scholar]
- Fuhrman, B.; Volkova, N.; Suraski, A.; Aviram, M. White wine with red wine-like properties: Increased extraction of grape skin polyphenols improves the antioxidant capacity of the derived white wine. J. Agric. Food Chem. 2001, 49, 3164–3168. [Google Scholar] [CrossRef]
- Ivanova, V.; Stefova, M.; Vojnoski, B.; Dörnyei, Á.; Márk, L.; Dimovska, V.; Stafilov, T.; Kilár, F. Identification of polyphenolic compounds in red and white grape varieties grown in R. Macedonia and changes of their content during ripening. Food Res. Int. 2011, 44, 2851–2860. [Google Scholar] [CrossRef]
- Rahbar, A.R.; Shakouri, M.M.; Shahidul Islam, M. Comparative effects of red and white grapes on oxidative markers and lipidemic parameters in adult hypercholesterolemic humans. Food Funct. 2015, 6, 1992–1998. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.M.; Toaldo, I.M.; Silva Haas, I.C.; Burin, V.M.; Caliari, V.; Luna, A.S.; Santos de Gois, J.; Bordignon-Luiz, M.T. Differential contribution of grape peel, pulp, and seed to bioaccessibility of micronutrients and major polyphenolic compounds of red and white grapes through simulated human digestion. J. Func. Foods 2019, 52, 699–708. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Hernanz, D.; Cifuentes-Gomez, T.; Escudero-Gilete, M.L.; Heredia, F.J.; Spencer, J.P.E. Assessment of white grape pomace from winemaking as source of bioactive compounds, and its antiproliferative activity. Food Chem. 2015, 183, 78–82. [Google Scholar] [CrossRef]
- Tian, Q.; Xu, Z.; Sun, X.; Deavila, J.; Du, M. Grape pomace inhibits colon carcinogenesis by suppressing cell proliferation and inducing epigenetic modifications. J. Nutr. Biochem. 2020, 84, 108443. [Google Scholar] [CrossRef]
- Yasui, Y.; Kim, M.; Oyama, T.; Tanaka, T. Colorectal carcinogenesis and suppression of tumor development by inhibition of enzymes and molecular targets. Curr. Enzym. Inhib. 2009, 5, 1–26. [Google Scholar] [CrossRef]
- Shan, B.E.; Wang, M.X.; Li, R.G. Quercetin inhibit human sw480 colon cancer growth in association with inhibition of cyclin d1 and surviving expression through Wnt/β-catenin signaling pathway. Cancer Invest. 2009, 27, 604–612. [Google Scholar] [CrossRef]
- Gómez-Alonso, S.; Collins, V.J.; Vauzour, D.; Rodríguez-Mateos, A.; Corona, G.; Spencer, J.P.E. Inhibition of colon adenocarcinoma cell proliferation by flavonols is linked to a G2/M cell cycle block and reduction in cyclin D1 expression. Food Chem. 2012, 130, 493–500. [Google Scholar] [CrossRef]
- Abdel-Moneim, E.; Shehata, A.M.; Alzahrani, S.O.; Shafi, M.E.; Mesalam, N.M.; Taha, A.E.; Swelum, A.A.; Arif, M.; Fayyaz, M.; Abd El-Hack, M.E. The role of polyphenols in poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1851–1866. [Google Scholar] [CrossRef]
- Kim, E.Y.; Pai, T.K.; Han, O. Effect of bioactive dietary polyphenols on zinc transport across the intestinal Caco-2 cell monolayers. J. Agr. Food Chem. 2011, 59, 3606–3612. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Kim, E.Y.; Lindsay, E.A.; Han, O. Bioactive dietary polyphenols inhibit heme iron absorption in a dose-dependent manner in human intestinal Caco-2 cells. J. Food Sci. 2011, 76, 143–150. [Google Scholar] [CrossRef]
- Acésio, N.A.; Carrijo, G.S.; Batista, T.H.; Damasceno, J.L.; Côrrea, M.B.; Tozatti, M.G.; Cunha, W.R.; Tavares, D.C. Assessment of the antioxidant, cytotoxic, and genotoxic potential of the Annona muricata leaves and their influence on genomic stability. J Toxicol. Environ. Health Part A 2017, 80, 1290–1300. [Google Scholar] [CrossRef] [PubMed]
- Rody, H.V.S.; Gontyo, D.C.; Coelho, M.V.P.; Ventrella, M.C.; Pádua, R.M.; Fietto, L.G.; Leite, V.J.P. Mutagenic activity and chemical composition of phenolic-rich extracts of leaves from two species of Ficus medicinal plants. Toxicol. Environ. Health Part A 2018, 81, 861–872. [Google Scholar] [CrossRef]
- Ramos, S.; Alia, M.; Bravo, L.; Goya, L. Comparative effects of food-derived polyphenols on the viability and apoptosis of a human hepatoma cell line (HepG2). J. Agric. Food Chem. 2005, 53, 1271–1280. [Google Scholar] [CrossRef]
- Ugartondo, V.; Mitjans, M.; Lozano, C.; Torres, J.L.; Vinardell, M.P. Comparative study of the cytotoxicity induced by antioxidant epicatechin conjugates obtained from grape. J. Agric. Food Chem. 2006, 54, 6945–6950. [Google Scholar] [CrossRef]
- Stagos, D.; Spanou, C.; Margariti, M.; Stathopoulos, C.; Mamuris, Z.; Kazantzoglou, G.; Magiatis, P.; Kouretas, D. Cytogenetic effects of grape extracts (Vitis vinifera) and polyphenols on mitomycin C-induced sister chromatid exchanges (SCEs) in human blood lymphocytes. J. Agric. Food Chem. 2007, 55, 5246–5252. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Lou, H.X. Effects of polyphenols from grape seeds on oxidative damage to cellular DNA. Mol. Cell. Biochem. 2008, 267, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.Q.; Deng, G.F.; Guo, Y.J.; Li, H.B. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Jitareanu, A.; Caba, I.C.; Trifan, A.; Padureanu, S.; Agoroaei, L. Triticum aestivum assay—A useful tool for environmental monitoring and toxicity assessment. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 1005–1018. [Google Scholar] [CrossRef]
- Fiskesjö, G. The Allium test as a standard in environmental monitoring. Hereditas 1985, 102, 99–112. [Google Scholar] [CrossRef]
- Fiskesjö, G. Allium test I: A 2–3 day plant test for toxicity assessment by measuring the mean root growth of onions (Allium cepa L. ) Environ. Toxicol. Water Qual. 1993, 8, 461–470. [Google Scholar] [CrossRef]
- Aksoy, O.; Deveci, A. The investigation of the cytotoxic effects of some pesticides on soybean (Glycine max L.). Cytologia 2012, 77, 475–483. [Google Scholar] [CrossRef][Green Version]
- Shehab, A.S. Cytological effects of medicinal plants in Qatar. I. Mitotic effect of water extract of Pulicaria crispa on Allium cepa. Cytologia 1979, 44, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Türkoğlu, S. Genotoxicity of five food preservatives tested on root tips of Allium cepa L. Mutat. Res. 2007, 626, 4–14. [Google Scholar] [CrossRef]
- Nefic, H.; Musanovic, J.; Metovic, A.; Kurteshi, K. Chromosomal and nuclear alterations in root tip cells of Allium cepa L. induced by alprazolam. Med. Arch. 2013, 67, 388–392. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.D.; Hoagland, R.E. The effects of naturally occurring phenolic compounds on seed germination. Weed Sci. 1982, 30, 206–212. [Google Scholar] [CrossRef]
- Ianni, A.; Martino, G. Dietary grape pomace supplementation in dairy cows: Effect on nutritional quality of milk and its derived dairy products. Foods 2020, 9, 168. [Google Scholar] [CrossRef]
- Özkan, G.; Sagdiç, O.; Baydar, G.N.; Kurumahmutoglu, Z. Antibacterial activities and total phenolic contents of grape pomace extracts. J. Sci. Food Agric. 2004, 84, 1807–1811. [Google Scholar] [CrossRef]
- Olejar, K.J.; Vandermeer, C.; Fedrizzi, B.; Kilmartin, P.A. A horticultural medium established from the rapid removal of phytotoxins from winery grape marc. Horticulturae 2019, 5, 69. [Google Scholar] [CrossRef]
- Medina, A.L.; Haas, L.I.R.; Chaves, F.C.; Salvador, M.; Zambiazi, R.C.; Silva, W.P.; Nora, L.; Rombaldi, C.V. Araçá (Psidium cattleianum Sabine) fruit extracts with antioxidant and antimicrobial activities and antiproliferative effect on human cancer cells. Food Chem. 2011, 128, 916–922. [Google Scholar] [CrossRef]
- Bobinaitė, R.C.; Grootaert, C.; Camp, J.V.; Šarkinas, A.P.R.; Liaudanskas, M.; Žvikas, V.; Viškelis, P.; Venskutonis, P.R. Chemical composition, antioxidant, antimicrobial and antiproliferative activities of the extracts isolated from the pomace of rowanberry (Sorbus aucuparia L.). Food Res. Int. 2020, 136, 109310. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Ramos, S.J.; Siqueira, J.O.; Faquin, V.; Castro, E.M.; Amaral, D.C.; Techio, V.H.; Coelho, L.C.; Silva, P.H.P.; Schnug, E.; et al. Bioaccumulation and effects of lanthanum on growth and mitotic index in soybean plants. Ecotox. Environ. Saf. 2015, 122, 136–144. [Google Scholar] [CrossRef]
- Shehab, A.S. Cytological Effects of Medicinal Plants in Qatar. II. Mitotic effect of water extract of Teucrium pilosum on Allium cepa. Cytologia 1980, 45, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Shehab, A.S.; Adam, Z.M. Cytological effects of medicinal plants in Qatar. III. Mitotic effect of water extract of Anastatica hierochuntica L. on Allium cepa. Cytologia 1983, 48, 343–348. [Google Scholar] [CrossRef]
- Pozo-Guisado, E.; Alvarez-Barrientos, A.; Mulero-Navarro, S.; Santiago-Josefat, B.; Fernandez-Salguero, P.M. The antiproliferative activity of resveratrol results in apoptosis in MCF-7 but not in MDA-MB-231 human breast cancer cells: Cell-specific alteration of the cell cycle. Biochem. Pharmacol. 2002, 64, 1375–1386. [Google Scholar] [CrossRef]
- Teerarak, M.; Laosinwattana, C.; Charoenying, P. Evaluation of allelopathic, decomposition and cytogenetic activities of Jasminum officinale L. f. var. grandiflorum (L.) Kob. on bioassay plants. Bioresour. Technol. 2010, 101, 5677–5684. [Google Scholar]
- Gavrilă, L. Genomica, 1st ed.; Enciclopedica: Bucharest, Romania, 2003; Volume 1, pp. 46–52. [Google Scholar]
- Easwaran, H.P.; Leonhardt, H.; Cardoso, M.C. Cell cycle markers for live cell analyses. Cell Cycle 2005, 4, 453–455. [Google Scholar] [CrossRef]
- Hahn, A.T.; Jones, J.T.; Meyer, T. Quantitative analysis of cell cycle phase durations and PC12 differentiation using fluorescent biosensors. Cell Cycle 2009, 8, 1044–1052. [Google Scholar] [CrossRef]
- Padfield, D.; Rittscher, J.; Thomas, N.; Roysam, B. Spatio-temporal cell cycle phase analysis using level sets and fast marching methods. Med. Image Anal. 2009, 13, 143–155. [Google Scholar] [CrossRef]
- Adam, Z.M.; Farah, O.R. Cytological effects of water extracts of medicinal plants in Egypt mitotic disturbances induced by water extract of Cymbopogon proximus (Halfa barr) on Vicia faba. Citologia 1989, 54, 489–492. [Google Scholar] [CrossRef]
- Glińska, S.; Bartczak, M.; Oleksiak, S.; Wolska, A.; Gabara, B.; Posmyk, M.; Janas, K. Effects of anthocyanin-rich extract from red cabbage leaves on meristematic cells of Allium cepa L. Roots treated with heavy metals. Ecotoxicol. Environ. Saf. 2007, 68, 343–350. [Google Scholar] [CrossRef]
- Osuji, J.O.; Owei, S.D. Mitotic index studies on ‘Treculia Africana’ Decne in Nigeria. Aust. J. Agric. Eng. 2010, 1, 25–28. [Google Scholar]
- Akaneme, F.I.; Amaefule, C.C. Evaluation of the cytotoxicity and genotoxicity of aqueous leaf extracts of Azardirachta indica A. Juss. using the Allium test. J. Med. Plants Res. 2012, 6, 3898–3907. [Google Scholar] [CrossRef]
- Khanna, N.; Sharma, S. Allium cepa root chromosomal aberration assay: A Review. Indian J. Pharm. Biol. Res. 2013, 1, 105–119. [Google Scholar] [CrossRef]
- Kharitonov, V.S.; Semenov, V.V.; Barabanshchikov, B.I. Purine receptor agonists protect the genome of plant and animal cells from clastogen damage. Bull. Exp. Biol. Med. 2001, 132, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Vanhauwaert, A.; Vanparys, P.; Kirsch-Volders, M. The in vivo gut micronucleus test detects clastogens and aneugens given by gavage. Mutagenesis 2001, 16, 39–50. [Google Scholar] [CrossRef]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef]
- Leme, D.M.; Franceschi de Angelis, D.; Marin-Morales, M.A. Action mechanisms of petroleum hydrocarbons present in waters impacted by an oil spill on the genetic material of Allium cepa root cells. Aquat. Toxicol. 2008, 88, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Trushin, M.V.; Ratushnyak, A.Y.; Arkharova, I.A.; Ratushnyak, A.A. Genetic alterations revealed in Allium cepa-test system under the action of some xenobiotics. World Appl. Sci. J. 2013, 22, 342–344. [Google Scholar]
- Bolzán, A.D. Using telomeric chromosomal aberrations to evaluate clastogen-induced genomic instability in mammalian cells. Chromosome Res. 2020, 28, 259–276. [Google Scholar] [CrossRef]
- Akaneme, F.I.; Iyioke, I.V. Mutagenic potentials of the sterilizing fluid—Purital on root tip mitosis of Allium cepa. Bio-Res. 2008, 6, 293–297. [Google Scholar]
- Rodrigues, M.A.; Probst, C.E.; Zayats, A.; Davidson, B.; Riedel, M.; Li, Y.; Venkatachalam, V. The in vitro micronucleus assay using imaging flow cytometry and deep learning. NPJ Syst. Biol. Appl. 2021, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Hintzsche, H.; Ulrike, H.U.; Poth, A.; Utesch, D.; Lott, J.; Stopper, H. Fate of micronuclei and micronucleated cells. Mutat. Res./Rev. Mutat. Res. 2017, 771, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, M.J.; Milne, L.; Nicotera, P.; Orrenius, S. 1,10-Phenanthroline stimulates internucleosomal DNA fragmentation in isolated rat-liver nuclei by promoting redox activity of endogenous copper ions. Biochem. J. 1996, 313, 163–169. [Google Scholar] [CrossRef]
- Azmi, A.S.; Bhat, S.H.; Hadi, S.M. Resveratrol–Cu(II) induced DNA breakage in human peripheral lymphocytes: Implications for anticancer properties. FEBS Lett. 2005, 579, 3131–3135. [Google Scholar] [CrossRef] [PubMed]
- Hadi, S.M.; Bhat, S.H.; Azmi, A.S.; Hanif, S.; Shamim, U.; Ullah, M. Oxidative breakage of cellular DNA by plant polyphenols: A putative mechanism for anticancer properties. Semin. Cancer Biol. 2007, 17, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Sta, C.; Goujon, E.; Ferjani, E.; Ledoigt, G. Toxicity of sulcotrione and grape marc on Vicia faba cells. J. Agric. Food Chem. 2014, 62, 11777–11785. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | Mean Number of Cells | Mitotic Index (%) | Cells in Prophase (%) | Cells in Metaphase (%) | Cells in Anaphase (%) | Cells in Telophase (%) |
|---|---|---|---|---|---|---|
| Control | 10.465 | 25.33 ± 0.65 h | 11.09 ± 0.89 cd | 4.34 ± 0.06 d | 2.43 ± 0.54 c | 7.47 ± 0.38 cde |
| M1 | 10.060 | 22.17 ± 0.09 ef | 9.74 ± 0.45 bc | 4.22 ± 0.33 d | 2.14 ± 0.15 bc | 6.07 ± 0.78 bc |
| SB1 | 11.156 | 24.00 ± 0.13 g | 10.97 ± 0.02 cd | 2.69 ± 0.24 b | 1.88 ± 0.24 bc | 8.46 ± 0.35 de |
| M2 | 10.585 | 21.63 ± 0.18 de | 11.41 ± 0.35 d | 3.44 ± 0.04 c | 1.39 ± 0.08 ab | 5.39 ± 0.49 bc |
| SB2 | 10.574 | 22.88 ± 0.06 f | 9.44 ± 0.04 b | 2.46 ± 0.03 b | 1.66 ± 0.08 bc | 9.32 ± 0.05 e |
| M3 | 10.507 | 18.25 ± 0.23 b | 10.71 ± 0.27 bcd | 2.76 ± 0.12 b | 1.48 ± 0.11 bc | 3.30 ± 0.27 a |
| SB3 | 10.923 | 21.01 ± 0.72 cd | 10.59 ± 0.71 bcd | 2.33 ± 0.03 b | 1.21 ± 0.08 ab | 6.88 ± 0.37 bcd |
| M4 | 10.712 | 11.34 ± 0.56 a | 4.74 ± 0.35 a | 1.27 ± 0.15 a | 0.51 ± 0.06 a | 4.82 ± 0.66 ab |
| SB4 | 11.573 | 20.40 ± 0.13 c | 10.84 ± 0.92 bcd | 2.38 ± 0.37 b | 1.84 ± 0.08 bc | 5.34 ± 0.23 ab |
| Variant | Cells in A-T (%) | Cells in Interphase (%) | ||
|---|---|---|---|---|
| Normal A-T (%) | Aberrant A-T(%) | Normal Interphase (%) | Aberrant Interphase (%) | |
| Control | 9.77 ± 0.73 d | 0.13 ± 0.01 a | 74.67 ± 0.62 a | 0.07 ± 0.01 a |
| M1 | 7.41 ± 0.57 bc | 0.80 ± 0.06 b | 77.63 ± 0.08 d | 0.20 ± 0.03 ab |
| SB1 | 9.62 ± 0.11 cd | 0.72 ± 0.05 b | 75.96 ± 0.13 b | 0.04 ± 0.01 a |
| M2 | 5.07 ± 0.13 ab | 1.71 ± 0.09 cd | 78.08 ± 0.02 e | 0.29 ± 0.05 b |
| SB2 | 10.08 ± 0.09 d | 0.90 ± 0.04 b | 77.06 ± 0.07 c | 0.06 ± 0.01 a |
| M3 | 2.98 ± 0.42 a | 1.80 ± 0.04 cd | 81.25 ± 0.25 h | 0.50 ± 0.01 c |
| SB3 | 6.89 ± 0.48 b | 1.2 ± 0.02 bc | 78.81 ± 0.66 f | 0.18 ± 0.06 ab |
| M4 | 3.09 ± 0.64 a | 2.24 ± 0.11 d | 87.59 ± 0.42 i | 1.07 ± 0.07 d |
| SB4 | 5.81 ± 0.15 b | 1.37 ± 0.01 bcm | 79.28± 0.11 g | 0.32 ± 0.03 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padureanu, S.; Patras, A. Biological Response of Triticum aestivum L. to the Abiotic Stress Induced by Winemaking Waste. Agronomy 2022, 12, 1371. https://doi.org/10.3390/agronomy12061371
Padureanu S, Patras A. Biological Response of Triticum aestivum L. to the Abiotic Stress Induced by Winemaking Waste. Agronomy. 2022; 12(6):1371. https://doi.org/10.3390/agronomy12061371
Chicago/Turabian StylePadureanu, Silvica, and Antoanela Patras. 2022. "Biological Response of Triticum aestivum L. to the Abiotic Stress Induced by Winemaking Waste" Agronomy 12, no. 6: 1371. https://doi.org/10.3390/agronomy12061371
APA StylePadureanu, S., & Patras, A. (2022). Biological Response of Triticum aestivum L. to the Abiotic Stress Induced by Winemaking Waste. Agronomy, 12(6), 1371. https://doi.org/10.3390/agronomy12061371