
Intra- and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat

 ,
, 
 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
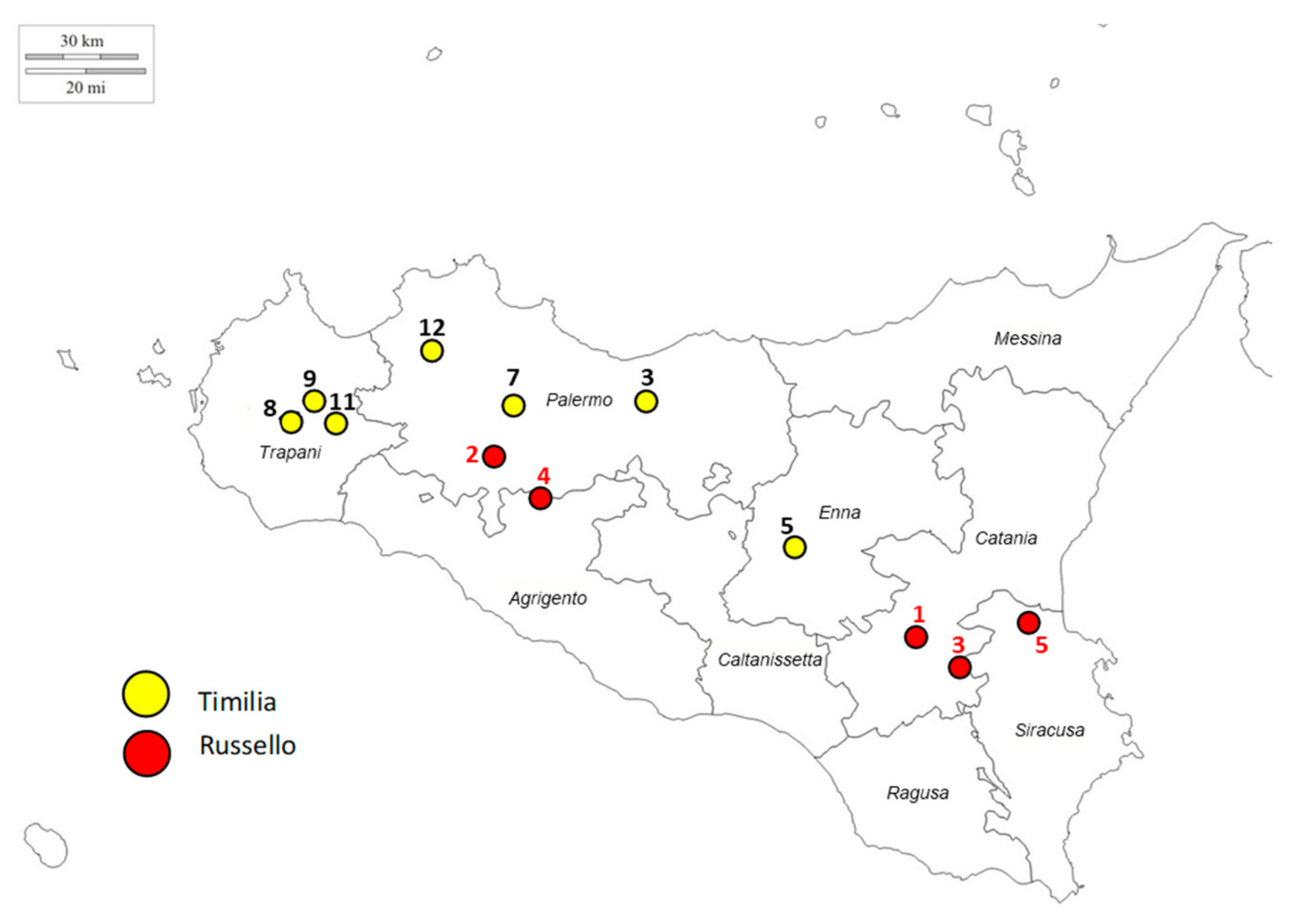
2.1. Plant Material, Genotyping, and SNP Filtering
2.2. Population Genetics and Diversity
2.3. Divergent Loci and Putative Genes under Selection
3. Results
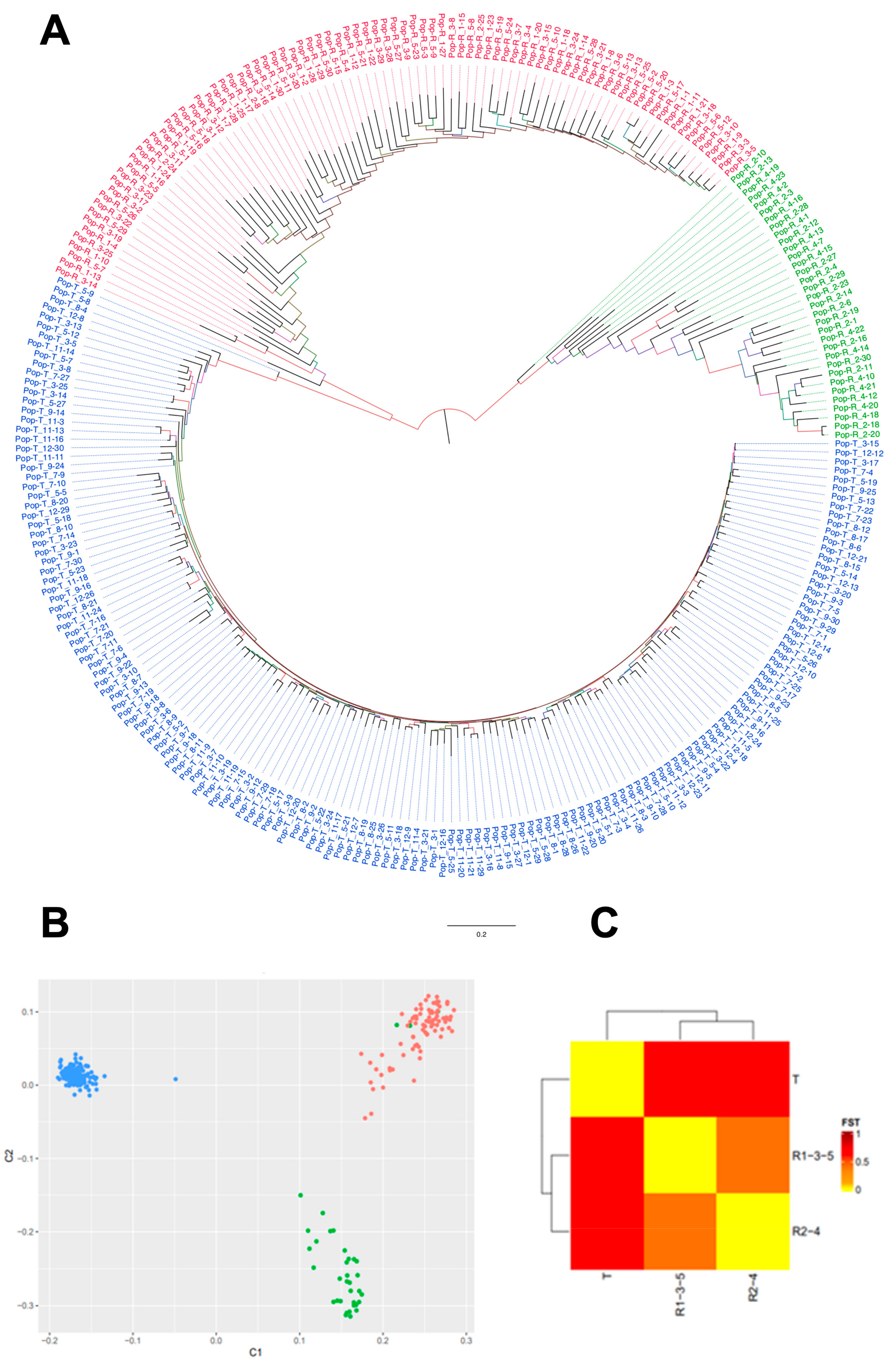
3.1. IBS Analysis
3.2. Genetic Relationships and Differentiation
3.3. Genetic Diversity Analysis
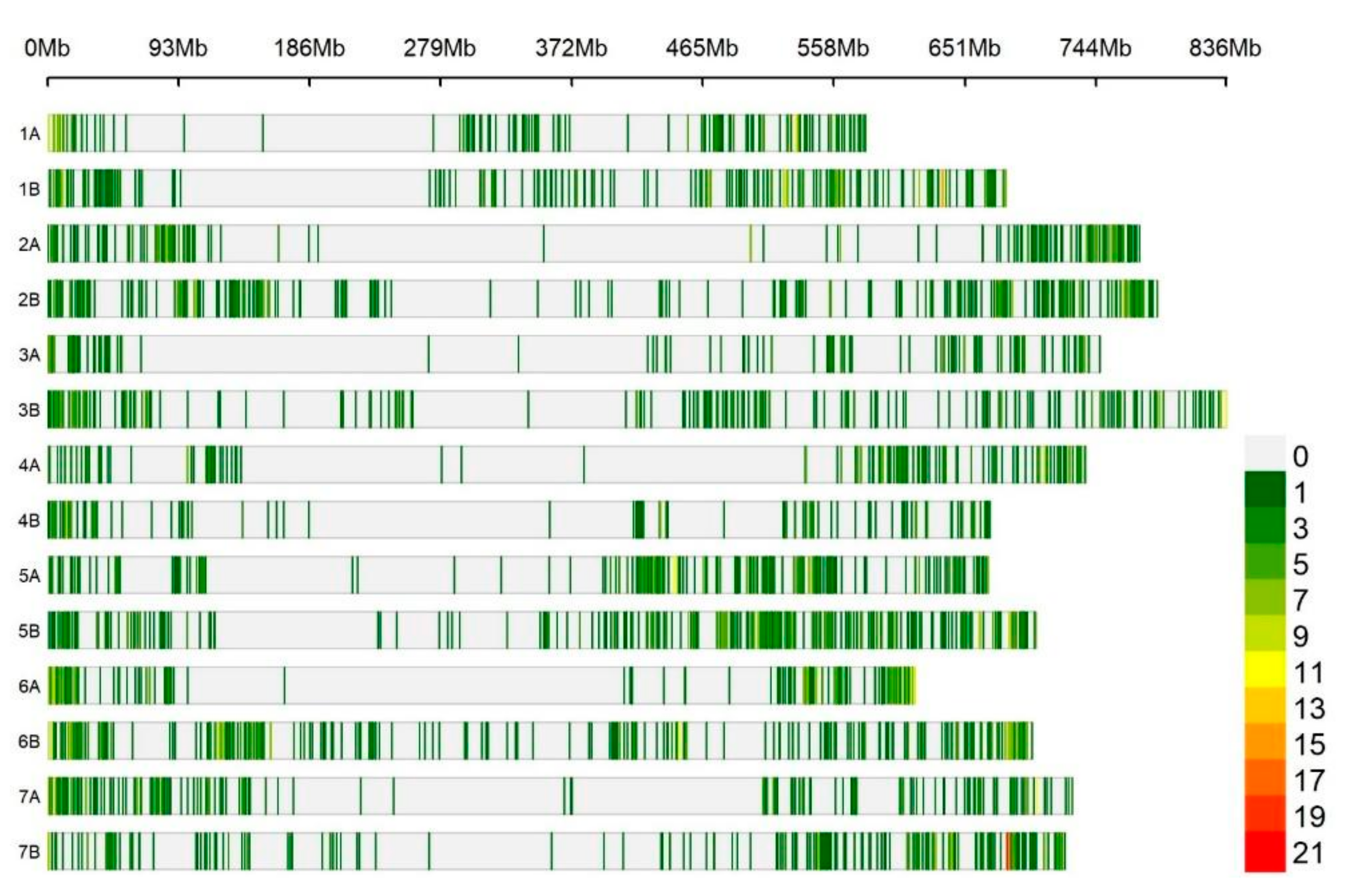
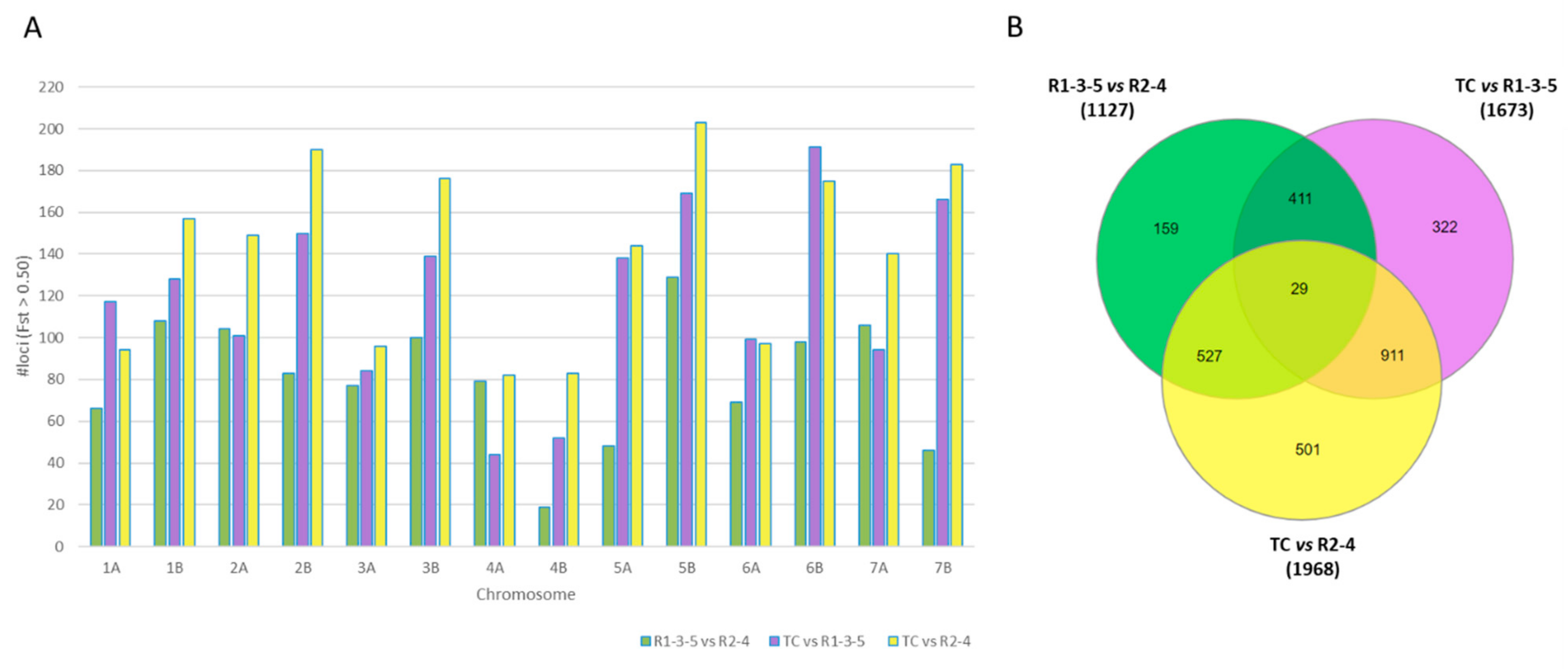
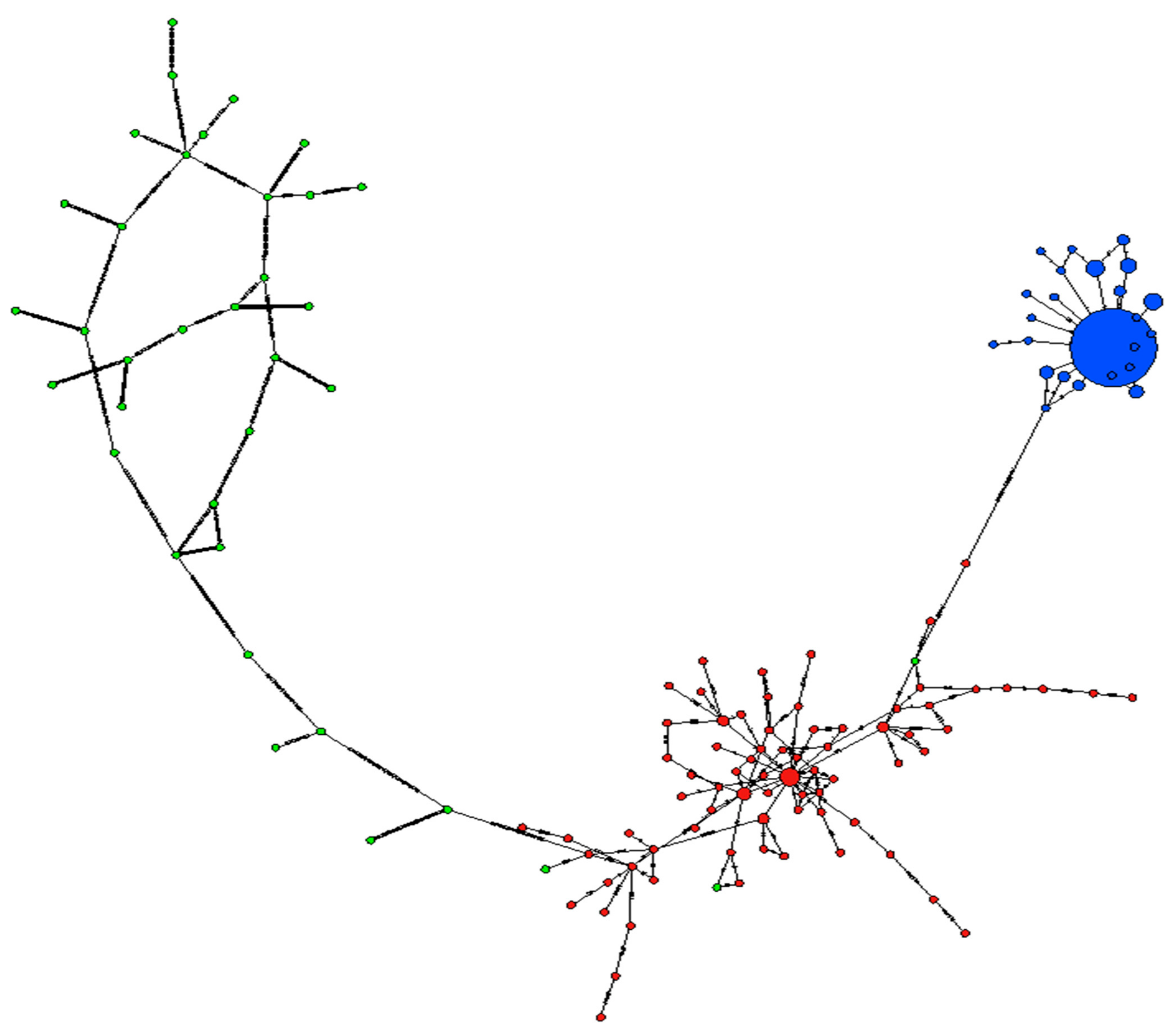
3.4. Divergent Loci and Haplotype Network
3.5. Patterns of Linkage Disequilibrium and Candidate Genes Associated with Divergent Loci
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bozzini, A.; Scarascia-Mugnozza, G.T. Relative frequency of chlorophyll to morphological and sterility mutations induced in durum wheat by radiations and chemicals. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 1970, 9, 589–597. [Google Scholar] [CrossRef]
- D’Amato, F. The progress of Italian wheat production in the first half of the 20th century: The contribution of breeders. Agric. Medit. 1989, 119, 157–174. [Google Scholar]
- De Vita, P.; Nicosia, O.L.D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding progress in morpho-physiological, agronomical and qualitative traits of durum wheat cultivars released in Italy during the 20th century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M.; Papa, R.; De Vita, P. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS ONE 2013, 8, e67280. [Google Scholar]
- Taranto, F.; D’Agostino, N.; Rodriguez, M.; Pavan, S.; Minervini, A.P.; Pecchioni, N.; Papa, R.; De Vita, P. Whole genome scan reveals molecular signatures of divergence and selection related to important traits in durum wheat germplasm. Front. Genet. 2020, 11, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vita, P.D.; Taranto, F. Durum wheat (Triticum turgidum ssp. durum) breeding to meet the challenge of climate change. In Advances in Plant Breeding Strategies: Cereals; Springer Pub.: Berlin/Heidelberg, Germany, 2019; pp. 471–524. [Google Scholar]
- Sharma, S.; Schulthess, A.W.; Bassi, F.M.; Badaeva, E.D.; Neumann, K.; Graner, A.; Özkan, H.; Werner, P.; Knüpffer, H.; Kilian, B. Introducing beneficial alleles from plant genetic resources into the wheat germplasm. Biology 2021, 10, 982. [Google Scholar] [CrossRef]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Migliorini, P.; Spagnolo, S.; Torri, L.; Arnoulet, M.; Lazzerini, G.; Ceccarelli, S. Agronomic and quality characteristics of old, modern and mixture wheat varieties and landraces for organic bread chain in diverse environments of northern Italy. Eur. J. Agron. 2016, 79, 131–141. [Google Scholar] [CrossRef]
- Giunta, F.; Bassu, S.; Mefleh, M.; Motzo, R. Is the technological quality of old durum wheat cultivars superior to that of modern ones when exposed to moderately high temperatures during grain filling? Foods 2020, 9, 778. [Google Scholar] [CrossRef]
- Salantur, A.; Karaoğlu, C. Macro-Microelements in Wheat Landraces and Their Use in Breeding. In Wheat Landraces; Springer Pub.: Berlin/Heidelberg, Germany, 2021; pp. 83–91. [Google Scholar]
- Mangini, G.; Nigro, D.; Margiotta, B.; De Vita, P.; Gadaleta, A.; Simeone, R.; Blanco, A. Exploring SNP diversity in wheat landraces germplasm and setting of a molecular barcode for fingerprinting. Cereal Res. Communic. 2018, 46, 377–387. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; et al. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Cillis, U. I Frumenti Siciliani; Tipografia Zuccarello and Izzi: Catania, Italy, 1942; ISBN 88-7751-229-6. [Google Scholar]
- Spina, A.; Blangiforti, S.; Cambrea, M.; Gallo, G.; Licciardello, S. Valorizzazione di germoplasma locale di frumento duro per la produzione di pani tipici. In Proceedings of the VIII Convegno Nazionale ‘‘La Biodiversità-una Risorsa per Sistemi Multifunzionali”, Lecce, Italy, 21–23 April 2008. [Google Scholar]
- Sciacca, F.; Cambrea, M.; Licciardello, S.; Pesce, A.; Romano, E.; Spina, A.; Virza, N.; Palumbo, M. Evolution of durum wheat: From Sicilian landraces to improved varieties. Options Méditerranéennes Ser. A 2014, 110, 139–145. [Google Scholar]
- Giancaspro, A.; Colasuonno, P.; Zito, A.; Blanco, A.; Pasqualone, A.; Gadaleta, A. Varietal traceability of bread ‘Pane Nero di Castelvetrano’ by denaturing high pressure liquid chromatography analysis of single nucleotide polymorphisms. Food Control 2016, 59, 809–817. [Google Scholar] [CrossRef]
- Spina, A.; Dinelli, G.; Palumbo, M.; Whittaker, A.; Cambrea, M.; Negri, L.; Bosi, S. Evaluation of standard physico-chemical and rheological parameters in predicting bread-making quality of durum wheat (Triticum turgidum L. ssp. durum [Desf.] Husn.). Intern. J. Food Sci. Techn. 2021, 56, 3278–3288. [Google Scholar] [CrossRef]
- Kornicke, F.; Werner, H. Handbuch des Getreidebaues: Die Arten and Varietaten des Getreides; Die sorten und der Anbau des Gertréîdes: Berlin, Germany, 1885. [Google Scholar]
- Sciacca, F.; Blanco, C.; Spina, A.; Virzì, N.; Palumbo, M. Biochemical and molecular characterization of durum wheat Sicilian landraces (Triticum durum Desf.). Italus Hortus 2006, 13, 503–507. [Google Scholar]
- Melini, V.; Melini, F.; Acquistucci, R. Nutritional characterization of an Italian traditional bread from ancient grains: The case study of the durum wheat bread “Pane di Monreale”. Eur. Food Res. Technol. 2020, 247, 193–200. [Google Scholar] [CrossRef]
- Visioli, G.; Bonas, U.; Dal Cortivo, C.; Pasini, G.; Marmiroli, N.; Mosca, G.; Vamerali, T. Variations in yield and gluten proteins in durum wheat varieties under late-season foliar vs. soil application of nitrogen fertilizer in a northern Mediterranean environment. J. Sci. Food Agric. 2018, 98, 2360–2369. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, G.M. La scelta della varietà ai fini della valorizzazione della qualità della granicoltura iblea. In Proceedings of the Conference “Il Futuro Della Granicoltura Iblea”, Modica, Italy, 11–17 December 1994. [Google Scholar]
- De Santis, M.A.; Giuliani, M.M.; Giuzio, L.; De Vita, P.; Lovegrove, A.; Shewry, P.R.; Flagella, Z. Differences in gluten protein composition between old and modern durum wheat genotypes in relation to 20th century breeding in Italy. Eur. J. Agron. 2017, 87, 19–29. [Google Scholar] [CrossRef]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Dataset of the metabolic and CM-like protein fractions in old and modern wheat Italian genotypes. Data Brief 2019, 27, 104730. [Google Scholar] [CrossRef]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Qualitative proteomic comparison of metabolic and CM-like protein fractions in old and modern wheat Italian genotypes by a shotgun approach. J. Proteom. 2020, 211, 103530. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Quantitative label-free comparison of the metabolic protein fraction in old and modern italian wheat genotypes by a shotgun approach. Molecules 2021, 26, 2596. [Google Scholar] [CrossRef] [PubMed]
- Pandolfo, A.; Messina, B.; Russo, G. Evaluation of Glycemic Index of Six Different Samples of Commercial and Experimental Pasta Differing in Wheat Varieties and Production Processes. Foods 2021, 10, 2221. [Google Scholar] [CrossRef] [PubMed]
- Blangiforti, S.; Gallo, G.; Porfiri, O. Morpho-agronomic and molecular assessment of a collection of Sicilian wheats [Triticum aestivum L.; Triticum durum Desf.]. Italus Hortus 2006, 13, 364–369. [Google Scholar]
- Figliuolo, G.; Mazzeo, M.; Greco, I. Temporal variation of diversity in Italian durum wheat germplasm. Gen. Res. Crop Evol. 2007, 54, 615–626. [Google Scholar] [CrossRef]
- Marzario, S.; Logozzo, G.; David, J.L.; Zeuli, P.S.; Gioia, T. Molecular genotyping (SSR) and agronomic phenotyping for utilization of durum wheat (Triticum durum Desf.) ex situ collection from Southern Italy: A combined approach including pedigreed varieties. Genes 2018, 9, 465. [Google Scholar] [CrossRef] [Green Version]
- Taranto, F.; Nicolia, A.; Pavan, S.; De Vita, P.; D’Agostino, N. Biotechnological and digital revolution for climate-smart plant breeding. Agronomy 2018, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Delvento, C.; Ricciardi, L.; Lotti, C.; Ciani, E.; D’Agostino, N. Recommendations for choosing the genotyping method and best practices for quality control in crop genome-wide association studies. Front. Genet. 2020, 11, 447. [Google Scholar] [CrossRef]
- Kabbaj, H.; Sall, A.T.; Al-Abdallat, A.; Geleta, M.; Amri, A.; Filali-Maltouf, A.; Belkadi, B.; Ortiz, R.; Bassi, F.M. Genetic diversity within a global panel of durum wheat (Triticum durum) landraces and modern germplasm reveals the history of alleles exchange. Front. Plant Sci. 2017, 8, 1277. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Alvaro, F.; Royo, C.; Soriano, J.M. From landraces to improved cultivars: Assessment of genetic diversity and population structure of Mediterranean wheat using SNP markers. PLoS ONE 2019, 14, e0219867. [Google Scholar] [CrossRef] [Green Version]
- Ganugi, P.; Palchetti, E.; Gori, M.; Calamai, A.; Burridge, A.; Biricolti, S.; Benedettelli, S.; Masoni, A. Molecular Diversity within a Mediterranean and European Panel of Tetraploid Wheat (T. turgidum subsp.) Landraces and Modern Germplasm Inferred Using a High-Density SNP Array. Agronomy 2021, 11, 414. [Google Scholar] [CrossRef]
- Soriano, J.M.; Colasuonno, P.; Marcotuli, I.; Gadaleta, A. Meta-QTL analysis and identification of candidate genes for quality, abiotic and biotic stress in durum wheat. Sci. Rep. 2021, 11, 11877. [Google Scholar] [CrossRef] [PubMed]
- Mangini, G.; Taranto, F.; Delvecchio, L.N.; Pasqualone, A.; Blanco, A. Development and validation of a new Ppo-A1 marker useful for marker-assisted selection in tetraploid wheats. Molec. Breed. 2014, 34, 385–392. [Google Scholar] [CrossRef]
- Wickham, H. Getting started with qplot. In Ggplot2; Springer Pub.: New York, NY, USA, 2009; pp. 9–26. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Molec. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Smouse, R.P.P.; Peakall, R. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Leviyang, S.; Hamilton, M.B. Properties of Weir and Cockerham’s Fst estimators and associated bootstrap confidence intervals. Theor. Popul. Biol. 2011, 79, 39–52. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Carranza-Gallego, G.; Guzmán, G.I.; Garcia-Ruíz, R.; Gonzalez de Molina, M.; Aguilera, E. Addressing the role of landraces in the sustainability of Mediterranean agroecosystems. Sustainability 2019, 11, 6029. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Schreiber, M.; Scholz, U.; Graner, A.; Reif, J.C.; Stein, N. Genebank genomics bridges the gap between the conservation of crop diversity and plant breeding. Nat. Genet. 2019, 51, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Raggi, L.; Pacicco, L.C.; Caproni, L.; Álvarez-Muñiz, C.; Annamaa, K.; Barata, A.M.; Negri, V. Analysis of landrace cultivation in Europe: A means to support in situ conservation of crop diversity. Biol. Conserv. 2022, 267, 109460. [Google Scholar] [CrossRef]
- Porceddu, E.; Vannella, S.; Perrino, P. Character analysis and numerical classification in a wheat collection from Sicily. Die Kult. 1981, 29, 251–266. [Google Scholar] [CrossRef]
- Laino, P.; Shelton, D.; Finnie, C.; Leonardis, A.M.D.; Mastrangelo, A.M.; Svensson, B.; Lafiandra, D.; Masci, S. Comparative proteome analysis of metabolic proteins from seeds of durum wheat (cv. Svevo) subjected to heat stress. Proteomics 2010, 10, 2359–2368. [Google Scholar] [CrossRef] [Green Version]
- Taranto, F.; Delvecchio, L.N.; Mangini, G.; Del Faro, L.; Blanco, A.; Pasqualone, A. Molecular and physico-chemical evaluation of enzymatic browning of whole meal and dough in a collection of tetraploid wheats. J. Cereal Sci. 2012, 55, 405–414. [Google Scholar] [CrossRef]
- Taranto, F.; Mangini, G.; Pasqualone, A.; Gadaleta, A.; Blanco, A. Mapping and allelic variations of Ppo-B1 and Ppo-B2 gene-related polyphenol oxidase activity in durum wheat. Mol. Breed. 2015, 35, 80. [Google Scholar] [CrossRef]
- Taranto, F.; Mangini, G.; Miazzi, M.M.; Stevanato, P.; De Vita, P. Polyphenol oxidase genes as integral part of the evolutionary history of domesticated tetraploid wheat. Genomics 2021, 113, 2989–3001. [Google Scholar] [CrossRef]
- Lo Bianco, M.; Siracusa, L.; Dattilo, S.; Venora, G.; Ruberto, G. Phenolic fingerprint of Sicilian modern varieties and old durum wheat landraces: A tool to assess with diversity. Cereal Chem. 2017, 94, 1045–1051. [Google Scholar]
- Morris, C.F. Determinants of wheat noodle color. J. Sci. Food Agric. 2018, 98, 5171–5180. [Google Scholar] [CrossRef]
- Siah, S.; Quail, K.J. Factors affecting Asian wheat noodle color and time-dependent discoloration—A review. Cereal Chem. 2018, 95, 189–205. [Google Scholar] [CrossRef]
- Liu, S.; Sun, Y.; Obadi, M.; Jiang, Y.; Chen, Z.; Jiang, S.; Xu, B. Effects of vacuum mixing and mixing time on the processing quality of noodle dough with high oat flour content. J. Cereal Sci. 2020, 91, 102885. [Google Scholar] [CrossRef]
- Zhai, S.; He, Z.; Wen, W.; Liu, J.; Jin, H.; Yan, J.; Zhang, P.; Zhang, Y.; Wan, Y.; Xia, X. Genetic architecture of polyphenol oxidase activity in wheat flour by genome-wide association study. Crop Sci. 2020, 60, 1281–1293. [Google Scholar] [CrossRef]
- Ali, I.; Iqbal, A.; Ullah, S.; Muhammad, I.; Yuan, P.; Zhao, Q.; Yang, M.; Zhang, H.; Huang, M.; Liang, H.; et al. Effects of Biochar Amendment and Nitrogen Fertilizer on RVA Profile and Rice Grain Quality Attributes. Foods 2022, 11, 625. [Google Scholar] [CrossRef]
- Taranto, F.; Pasqualone, A.; Mangini, G.; Tripodi, P.; Miazzi, M.M.; Pavan, S.; Montemurro, C. Polyphenol oxidases in crops: Biochemical, physiological and genetic aspects. Int. J. Mol. Sci. 2017, 18, 377. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, X. Recent advances in polyphenol oxidase-mediated plant stress responses. Phytochem 2021, 181, 112588. [Google Scholar] [CrossRef]
- Singh, S.; Kaur, I.; Kariyat, R. The multifunctional roles of polyphenols in plant-herbivore interactions. Int. J. Mol. Sci. 2021, 22, 1442. [Google Scholar] [CrossRef]
- Maucieri, C.; Caruso, C.; Bona, S.; Borin, M.; Barbera, A.C.; Cavallaro, V. Influence of salinity and osmotic stress on germination process in an old Sicilian landrace and a modern cultivar of Triticum durum Desf. Cereal Res. Commun. 2018, 46, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Ruisi, P.; Ingraffia, R.; Urso, V.; Giambalvo, D.; Alfonzo, A.; Corona, O.; Frenda, A.S. Influence of grain quality, semolinas and baker’s yeast on bread made from old landraces and modern genotypes of Sicilian durum wheat. Food Res. Int. 2021, 140, 110029. [Google Scholar] [CrossRef]
- Ficco, D.; Beleggia, R.; Pecorella, I.; Giovanniello, V.; Frenda, A.S.; Vita, P.D. Relationship between Seed Morphological Traits and Ash and Mineral Distribution along the Kernel Using Debranning in Durum Wheats from Different Geographic Sites. Foods 2020, 9, 1523. [Google Scholar] [CrossRef]
- Chai, S.; Yao, Q.; Zhang, X.; Xiao, X.; Fan, X.; Zeng, J.; Sha, L.; Kang, H.; Zhang, H.; Li, J. The semi-dwarfing gene Rht-dp from dwarf polish wheat (Triticum polonicum L.) is the “Green Revolution” gene Rht-B1b. BMC Genom. 2021, 22, 63. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | #Individuals | Na | PA | I | He |
|---|---|---|---|---|---|
| Russello | |||||
| R1 | 28 | 2.167 | 0.064 | 0.422 | 0.255 |
| R2 | 21 | 1.913 | 0.002 | 0.455 | 0.300 |
| R3 | 27 | 2.304 | 0.187 | 0.458 | 0.273 |
| R4 | 15 | 1.734 | 0.012 | 0.384 | 0.255 |
| R5 | 29 | 2.139 | 0.066 | 0.407 | 0.244 |
| Timilia | |||||
| T3 | 25 | 2.202 | 0.004 | 0.534 | 0.342 |
| T5 | 24 | 2.313 | 0.009 | 0.559 | 0.355 |
| T7 | 24 | 2.601 | 0.053 | 0.602 | 0.355 |
| T8 | 21 | 2.179 | 0.006 | 0.485 | 0.304 |
| T9 | 22 | 2.557 | 0.041 | 0.575 | 0.340 |
| T11 | 21 | 2.350 | 0.02 | 0.564 | 0.352 |
| T12 | 20 | 2.228 | 0.009 | 0.505 | 0.315 |
| RT Collection | |||||
| TC | 157 | 2.233 | 0.301 | 0.288 | 0.167 |
| RC | 120 | 2.597 | 0.665 | 0.515 | 0.319 |
| RC1-3-5 | 84 | 2.481 | 0.265 | 0.419 | 0.246 |
| RC2-4 | 36 | 1.871 | 0.048 | 0.396 | 0.258 |
| SNP Marker | Gene | Transcript ID | Distance (~Mb) | Comparison |
|---|---|---|---|---|
| tplb0025b13_1721 | MEI2-4 (OML4) | TRITD1Av1G001750 | 0 | R1-3-5 vs. R2-4 - TC vs. R2-4 |
| tplb0025b13_1721; TA001286-0611-w | Gamma-gliadin | TRITD1Av1G002070 | 0.50; 0 | R1-3-5 vs. R2-4; TC vs. R1-3-5 - TC vs. R2-4 |
| tplb0025b13_1721; Excalibur_c35919_107 | glutenin (LMW) | TRITD1Av1G002310 | 0.33; 0.01 | R1-3-5 vs. R2-4; TC vs. R1-3-5 - TC vs. R2-4 |
| Excalibur_c77035_156 | WCOR15 | TRITD1Av1G021590 | 0.55 | R1-3-5 vs. R2-4 |
| IAAV8147 | PP2A regulatory subunit TAP46 | TRITD1Av1G125250 | 0.18 | TC vs. R1-3-5 |
| RAC875_c6338_2719 | TELO2 | TRITD1Av1G127160 | 0.68 | TC vs. R1-3-5 |
| RAC875_c30138_595; IACX2941 | Gamma-gliadin | TRITD1Bv1G001870 | 0.27; 0.07 | TC vs. R1-3-5; TC vs. R2-4 |
| BS00004903_51 | Anthocyanin 5-aromatic acyltransferase | TRITD1Bv1G004660 | 0.38 | TC vs. R1-3-5 |
| BS00021975_51; CAP8_rep_c7343_88 | glutenin (HMW) | TRITD1Bv1G177800 | 0.18; 0 | TC vs. R1-3-5: R1-3-5 vs. R2-4 - TC vs. R2-4 |
| wsnp_Ex_c19556_28530231 | Ppd-A1 | TRITD2Av1G019250 | 0.27 | TC vs. R1-3-5; TC vs. R2-4 |
| wsnp_CAP11_rep_c8768_3788007 | FD-GOGAT | TRITD2Av1G036960 | 0 | TC vs. R1-3-5 - TC vs. R2-4 |
| wsnp_Ex_c12219_19526749 | SUS3 | TRITD2Av1G053920 | 3.9 | R1-3-5 vs. R2-4; TC vs. R1-3-5 |
| Kukri_c44442_274 | Lpx | TRITD2Av1G054540 | 0.62 | R1-3-5 vs. R2-4; TC vs. R1-3-5 |
| wsnp_be471201A_Ta_1_1 | Anthocyanin 3’-O-beta-glucosyltransferase | TRITD2Av1G252600 | 1.2 | TC vs. R2-4 |
| wsnp_be471201A_Ta_1_1 | Anthocyanin 5-aromatic acyltransferase | TRITD2Av1G253600 | 1.12 | R1-3-5 vs. R2-4; TC vs. R2-4 |
| RAC875_c110838_165; Ku_c13700_1196 | Ppo-A1 | TRITD2Av1G261300 | 0.62 | TC vs. R1-3-5; TC vs. R2-4 |
| RAC875_c110838_165; Ku_c13700_1196 | Ppo-A2 | TRITD2Av1G261390 | 0.4 | TC vs. R1-3-5; TC vs. R2-4 |
| RAC875_c110838_165; Ku_c13700_1196 | starch synthase | TRITD2Av1G261450 | 0.14 | TC vs. R1-3-5; TC vs. R2-4 |
| RAC875_c35438_474 | Beta-amylase | TRITD2Bv1G043100 | 1.2 | TC vs. R1-3-5; TC vs. R2-4 |
| BS00011630_51 | Ppo-B2 | TRITD2Bv1G224170 | 0.46 | TC vs. R1-3-5; TC vs. R2-4 |
| RAC875_c19534_68 | 9-cis-epoxycarotenoid dioxygenase | TRITD3Av1G020620 | 0.46 | R1-3-5 vs. R2-4 |
| BS00024548_51 | LEA | TRITD3Av1G260240 | 0 | R1-3-5 vs. R2-4 |
| BS00036352_51 | Carotenoid cleavage dioxygenase | TRITD3Bv1G162510 | 1.9 | TC vs. R2-4 |
| BobWhite_c1196_297 | Anthocyanin 3’-O-beta-glucosyltransferase | TRITD3Bv1G163770 | 0 | TC vs. R2-4 |
| Jagger_rep_c10288_53 | Rht-A1 | TRITD4Av1G194130 | 7.5 | TC vs. R2-4 |
| Jagger_rep_c10288_53 | NRT1/PTR | TRITD4Av1G198820 | 0.53 | TC vs. R2-4 |
| Tdurum_contig82378_264 | Rht-B1 | TRITD4Bv1G012280 | 4.5 | R1-3-5 vs. R2-4 - TC vs. R2-4 |
| Tdurum_contig82378_264 | Lpx1 | TRITD4Bv1G010710 | 7.8 | R1-3-5 vs. R2-4 - TC vs. R2-4 |
| Tdurum_contig82378_264 | Teosinte branched 1 protein | TRITD4Bv1G012050 | 4.5 | R1-3-5 vs. R2-4 - TC vs. R2-4 |
| wsnp_Ex_c6117_10704945 | Histone H3 | TRITD5Av1G041920 | 6 | TC vs. R1-3-5 |
| wsnp_Ra_c10053_16636851 | ASN | TRITD5Av1G043240 | 6 | TC vs. R1-3-5 |
| BS00076246_51 | Lpx | TRITD5Av1G200190 | 1.7 | TC vs. R1-3-5; TC vs. R2-4 |
| GENE-3601_145 | Vrn-A1 | TRITD5Av1G204680 | 10 | TC vs. R2-4 |
| BS00066456_51 | 15-cis-zeta-carotene isomerase | TRITD5Bv1G013720 | 5.7 | TC vs. R1-3-5; TC vs. R2-4 |
| GENE-3383_710 | 9-cis-epoxycarotenoid dioxygenase | TRITD5Bv1G192230 | 3.4 | R1-3-5 vs. R2-4 |
| wsnp_Ex_c5155_9140608 | Lpx | TRITD5Bv1G195300 | 1.7 | TC vs. R2-4 |
| Excalibur_c24051_502 | TdHMA3-B1 | TRITD5Bv1G197370 | 1 | TC vs. R1-3-5 |
| Excalibur_c24051_502 | Vrn-B1 | TRITD5Bv1G200510 | 10 | TC vs. R2-4 |
| BS00071573_51 | Terpene cyclase/mutase | TRITD6Av1G000890 | 0.08 | TC vs. R1-3-5; TC vs. R2-4 |
| BS00022660_51 | Terpene synthase | TRITD6Av1G008120 | 0.04 | TC vs. R2-4 |
| IACX5772 | Alpha-gliadin | TRITD6Av1G009340 | 1.6 | TC vs. R2-4 |
| IACX203 | Rht-14 | TRITD6Av1G140910 | 24 | TC vs. R2-4 |
| IACX203 | Nitrilase | TRITD6Av1G150150 | 0.17 | TC vs. R2-4 |
| BS00065082_51 | Nrt | TRITD6Av1G196930 | 0.56 | TC vs. R1-3-5 - TC vs. R2-4 |
| CAP7_c7415_267 | Nir | TRITD6Av1G199660 | 0 | TC vs. R2-4 |
| BS00109708_51 | Alpha-gliadin | TRITD6Bv1G014640 | 0.27 | TC vs. R1-3-5 |
| GENE-3945_245 | Anthocyanin 3’-O-beta-glucosyltransferase | TRITD6Bv1G159080 | 1.2 | R1-3-5 vs. R2-4; TC vs. R1-3-5 |
| BS00109912_51; RFL_Contig3621_1157 | Terpene cyclase/mutase | TRITD7Av1G000490 | 0.79 | R1-3-5 vs. R2-4; TC vs. R2-4 |
| BS00109912_51; RFL_Contig3621_1157 | Terpene synthase | TRITD7Av1G000650 | 0.84 | R1-3-5 vs. R2-4; TC vs. R2-4 |
| Excalibur_c25891_1402 | SUS1 | TRITD7Av1G009720 | 0.75 | TC vs. R1-3-5; TC vs. R2-4 |
| RAC875_c63822_185 | Anthocyanin 5-aromatic acyltransferase | TRITD7Av1G013740 | 0.94 | R1-3-5 vs. R2-4; TC vs. R2-4 |
| BS00062724_51 | GBSSI | TRITD7Av1G018110 | 0.82 | TC vs. R1-3-5 |
| Excalibur_c96483_102 | Beta-carotene isomerase | TRITD7Av1G226310 | 0.24 | TC vs. R1-3-5 |
| Jagger_c5275_99 | MADAGL17 | TRITD7Bv1G002790 | 0.56 | TC vs. R2-4 |
| Tdurum_contig85266_280 | GW2-B1 | TRITD7Bv1G002890 | 0 | R1-3-5 vs. R2-4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taranto, F.; Di Serio, E.; Miazzi, M.M.; Pavan, S.; Saia, S.; De Vita, P.; D’Agostino, N. Intra- and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat. Agronomy 2022, 12, 1326. https://doi.org/10.3390/agronomy12061326
Taranto F, Di Serio E, Miazzi MM, Pavan S, Saia S, De Vita P, D’Agostino N. Intra- and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat. Agronomy. 2022; 12(6):1326. https://doi.org/10.3390/agronomy12061326
Chicago/Turabian StyleTaranto, Francesca, Ermanno Di Serio, Monica Marilena Miazzi, Stefano Pavan, Sergio Saia, Pasquale De Vita, and Nunzio D’Agostino. 2022. "Intra- and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat" Agronomy 12, no. 6: 1326. https://doi.org/10.3390/agronomy12061326
APA StyleTaranto, F., Di Serio, E., Miazzi, M. M., Pavan, S., Saia, S., De Vita, P., & D’Agostino, N. (2022). Intra- and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat. Agronomy, 12(6), 1326. https://doi.org/10.3390/agronomy12061326