Effects of Low Temperature on the Amino Acid Composition of Wheat Grains

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
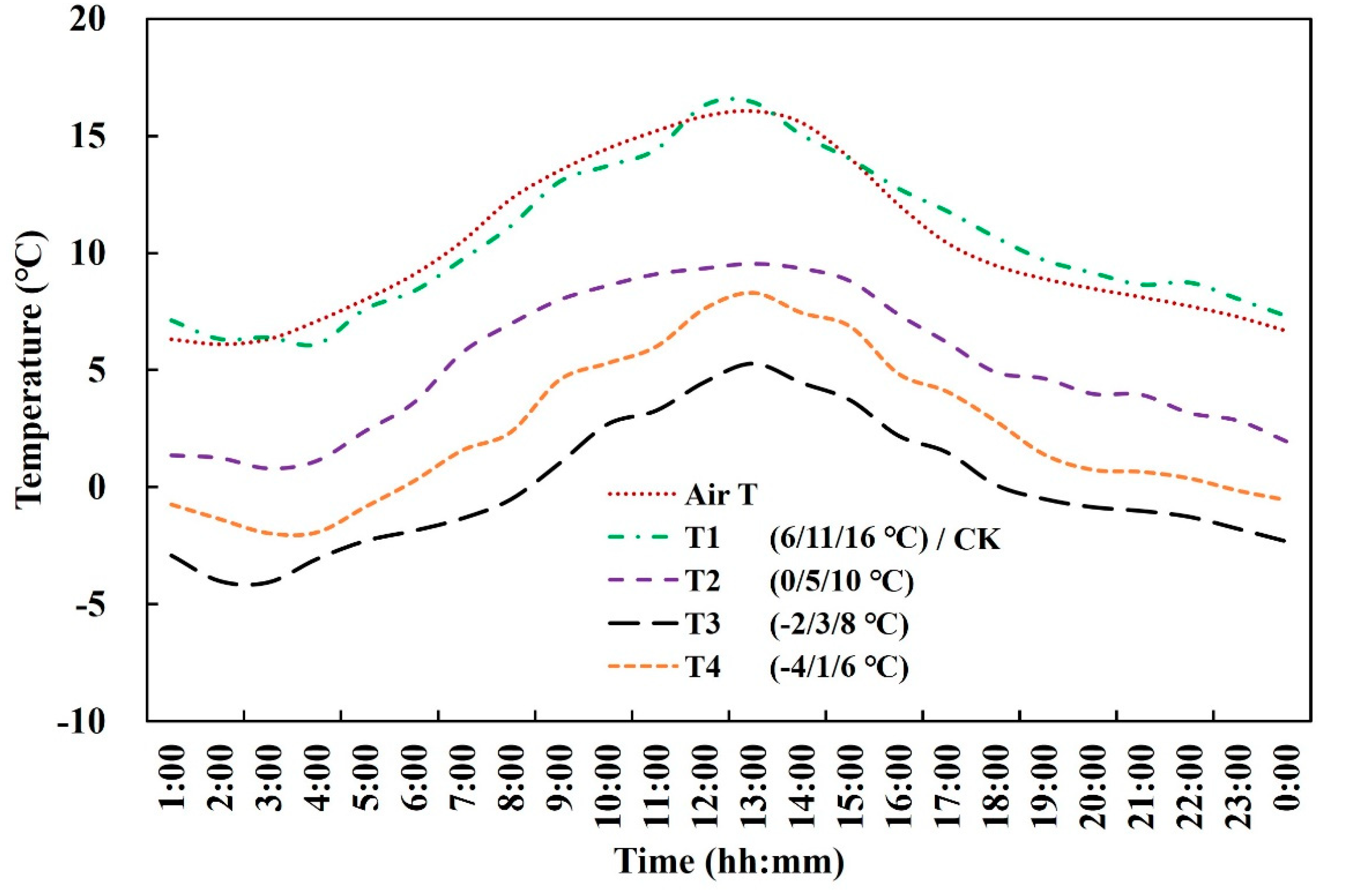
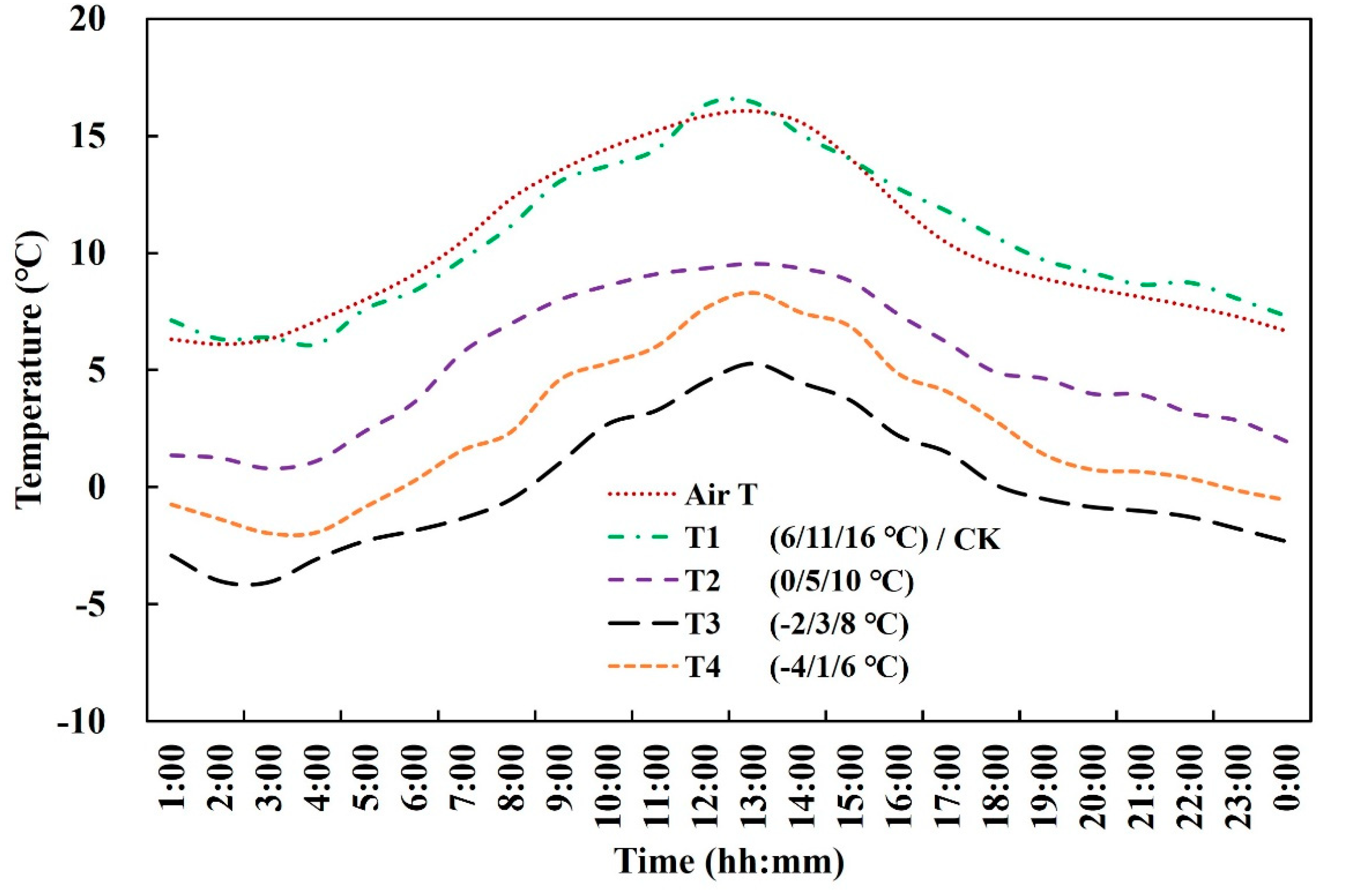
2.1. Experimental Design
2.2. Sample Preparation and Measurement
2.3. Amino Acid Scoring Method
2.3.1. Amino Acid Score
2.3.2. Ratio Coefficient of Amino Acids
2.3.3. Essential Amino Acid Index
2.4. Statistical Analysis
3. Results
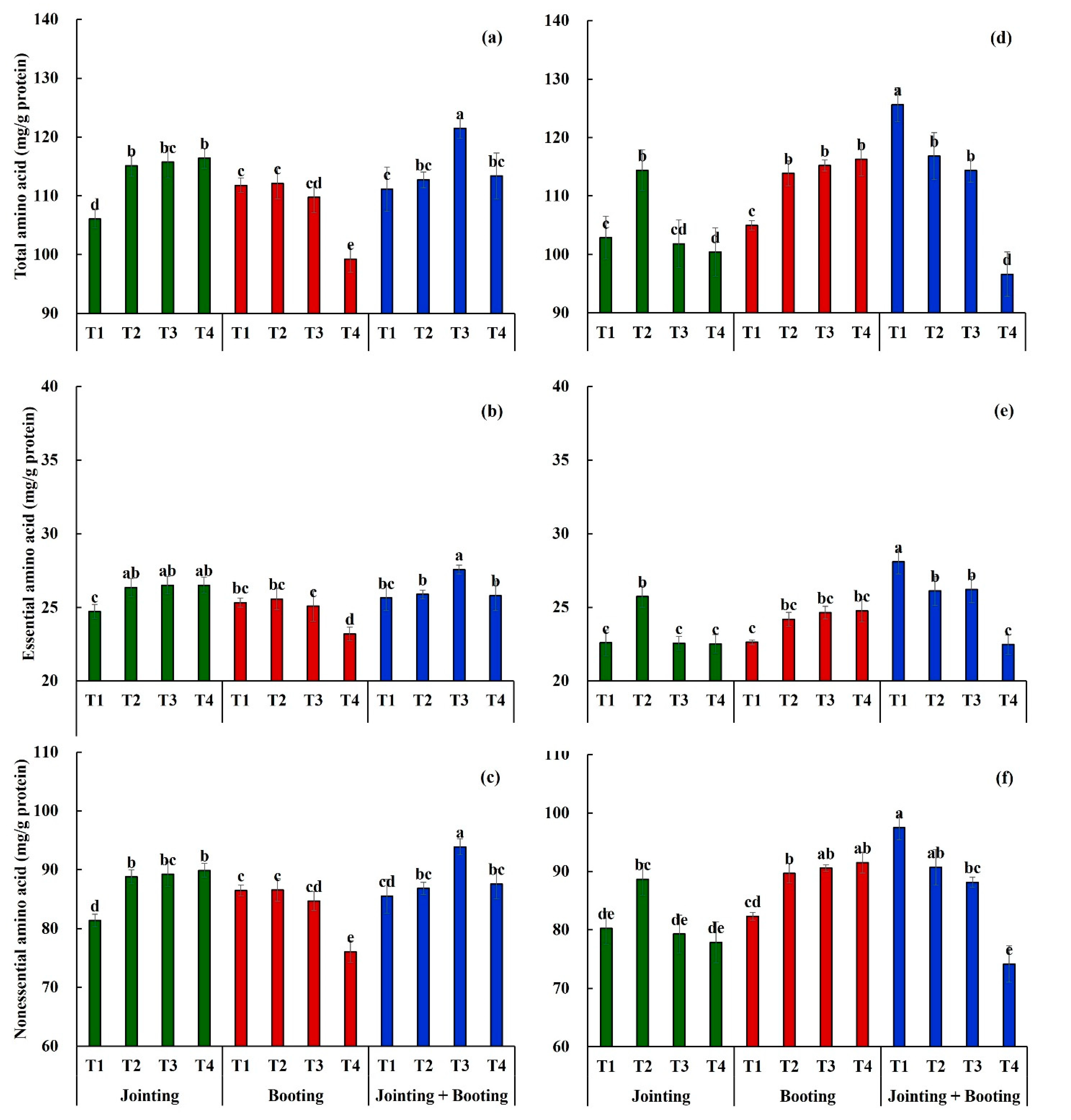
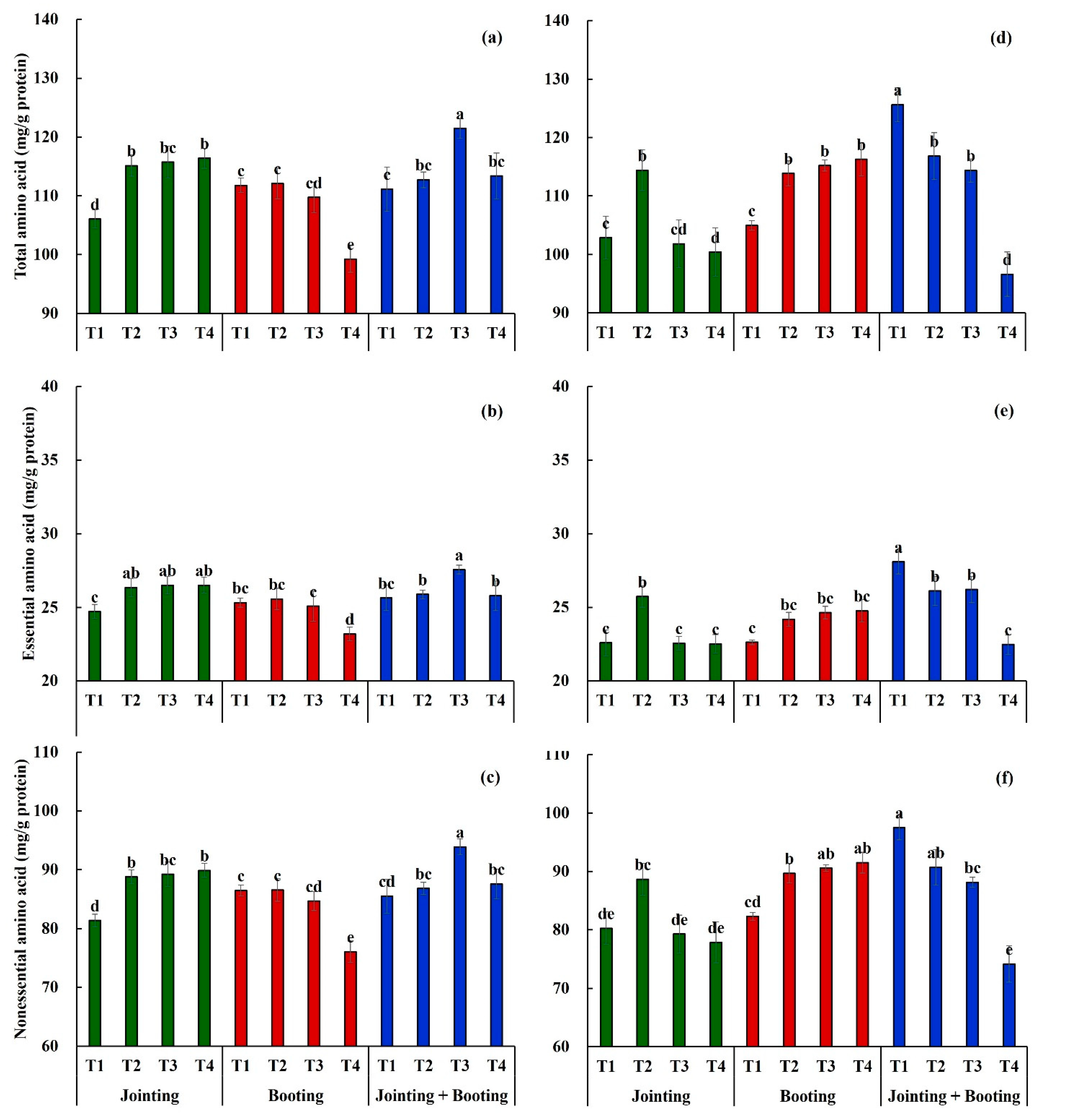
3.1. Effects of Low-Temperature on the Amino Acid Content in Wheat Grain
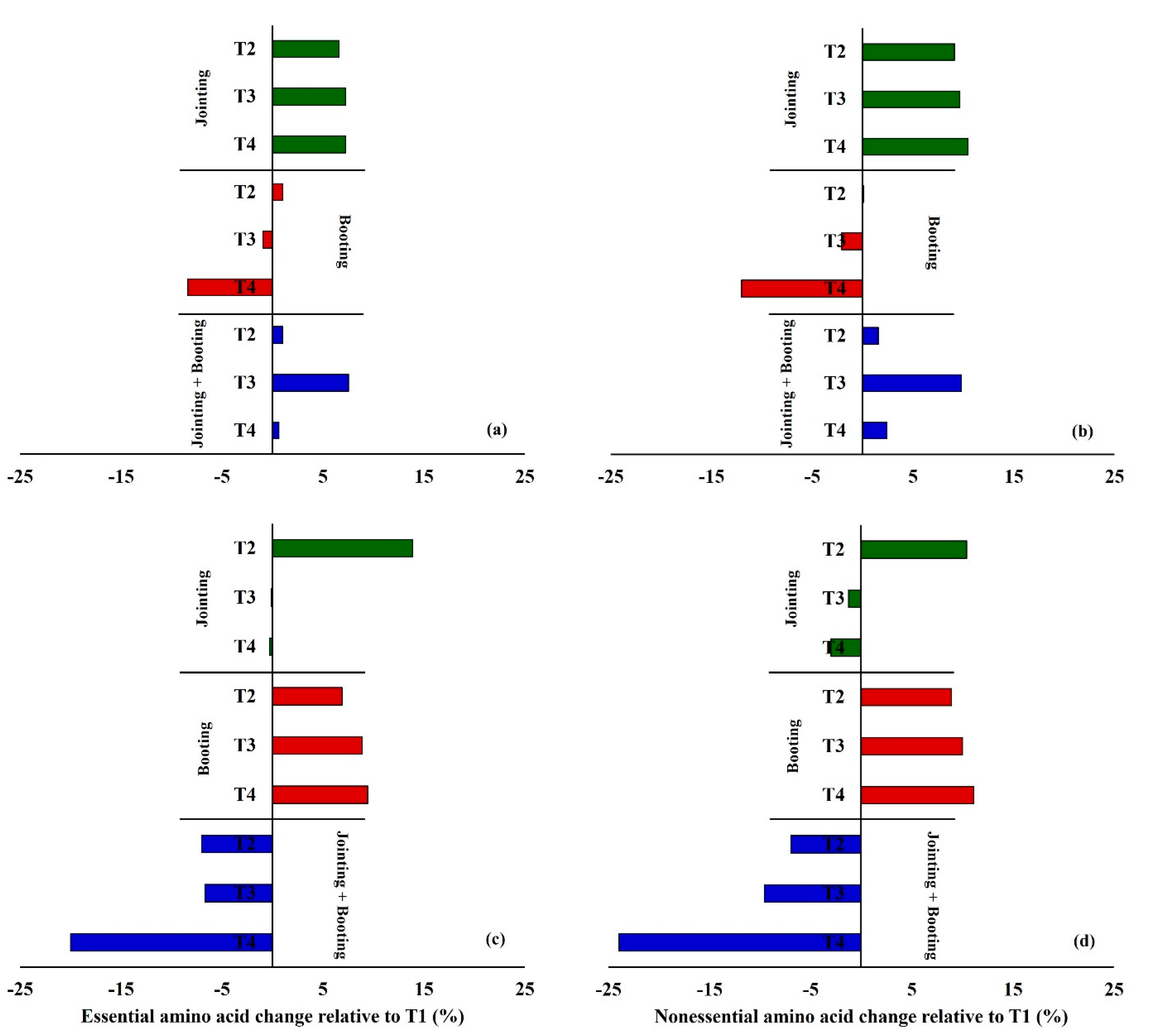
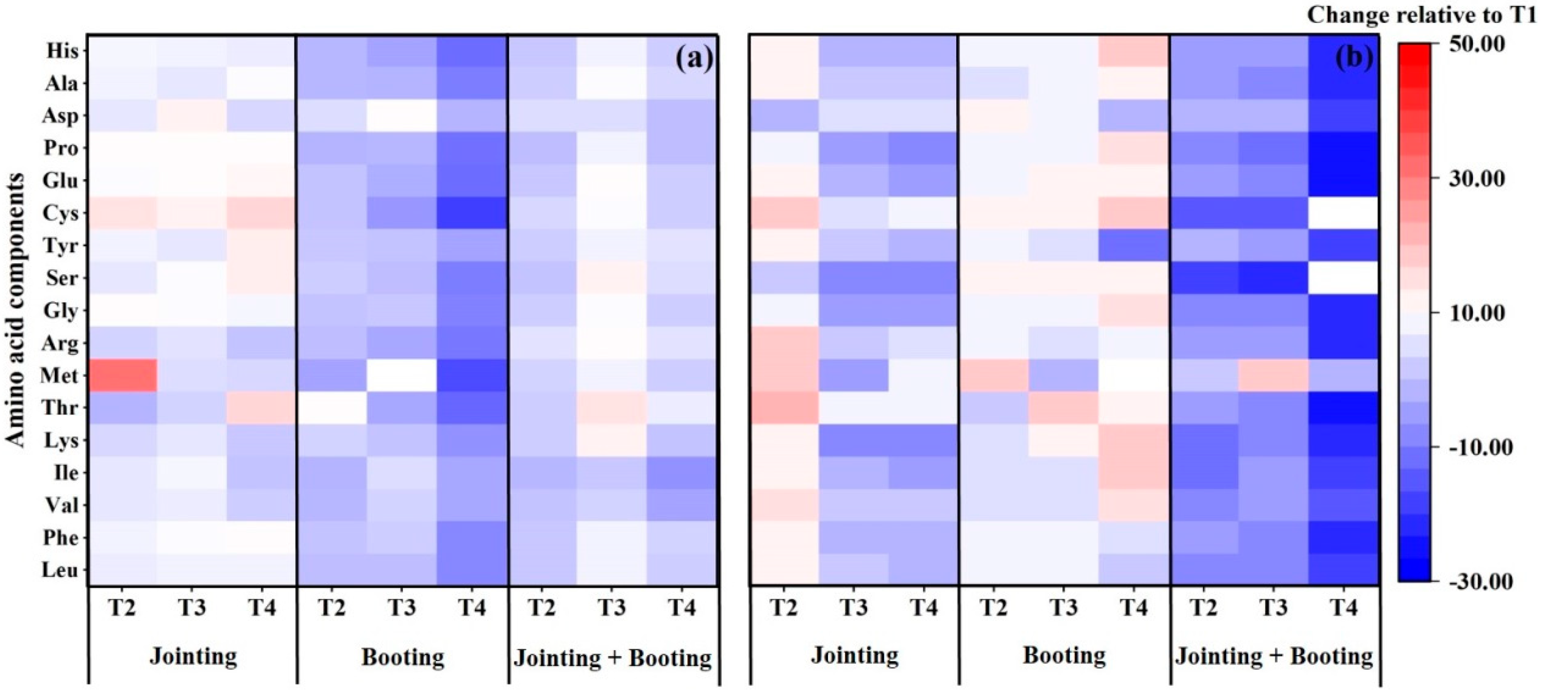
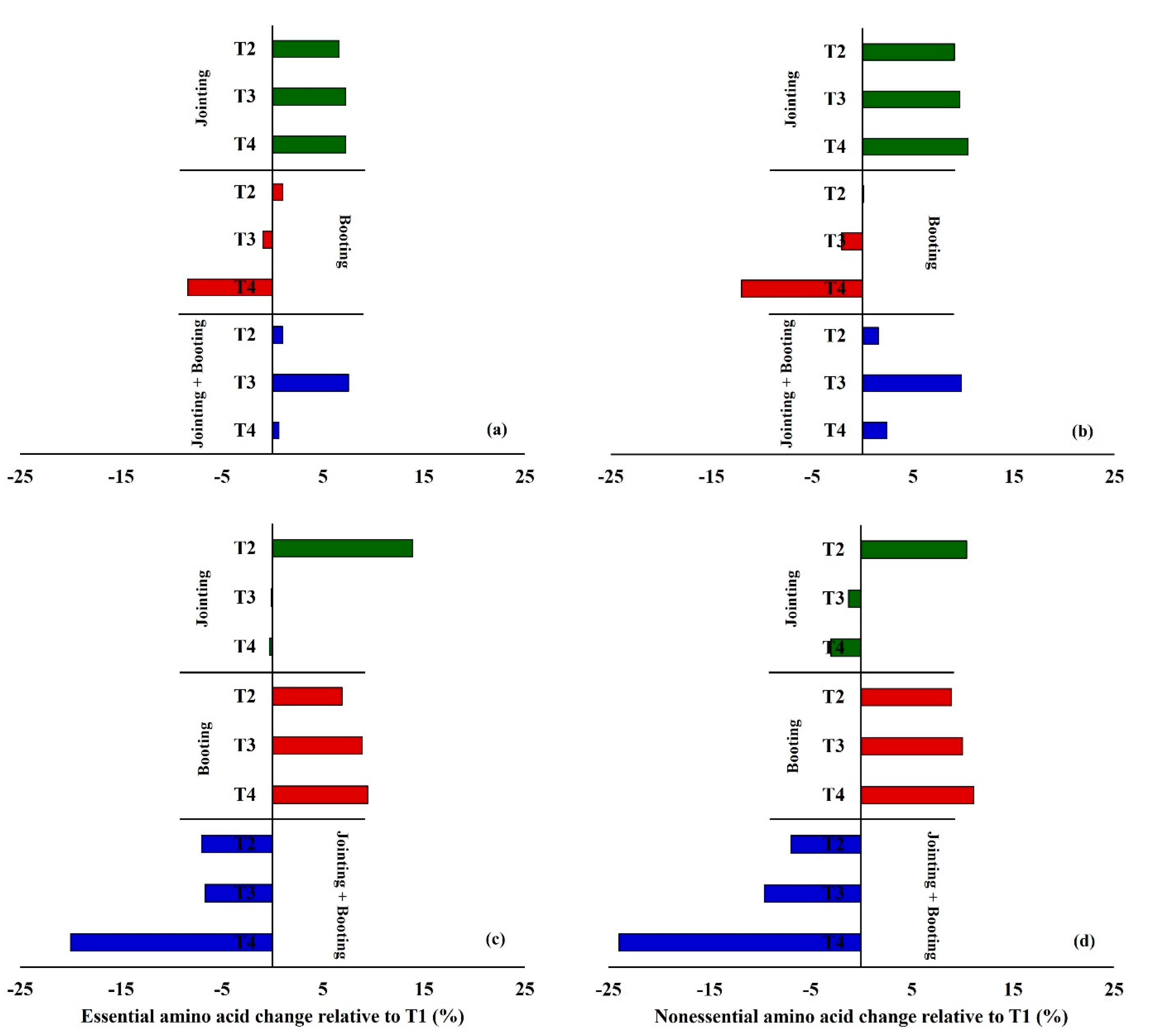
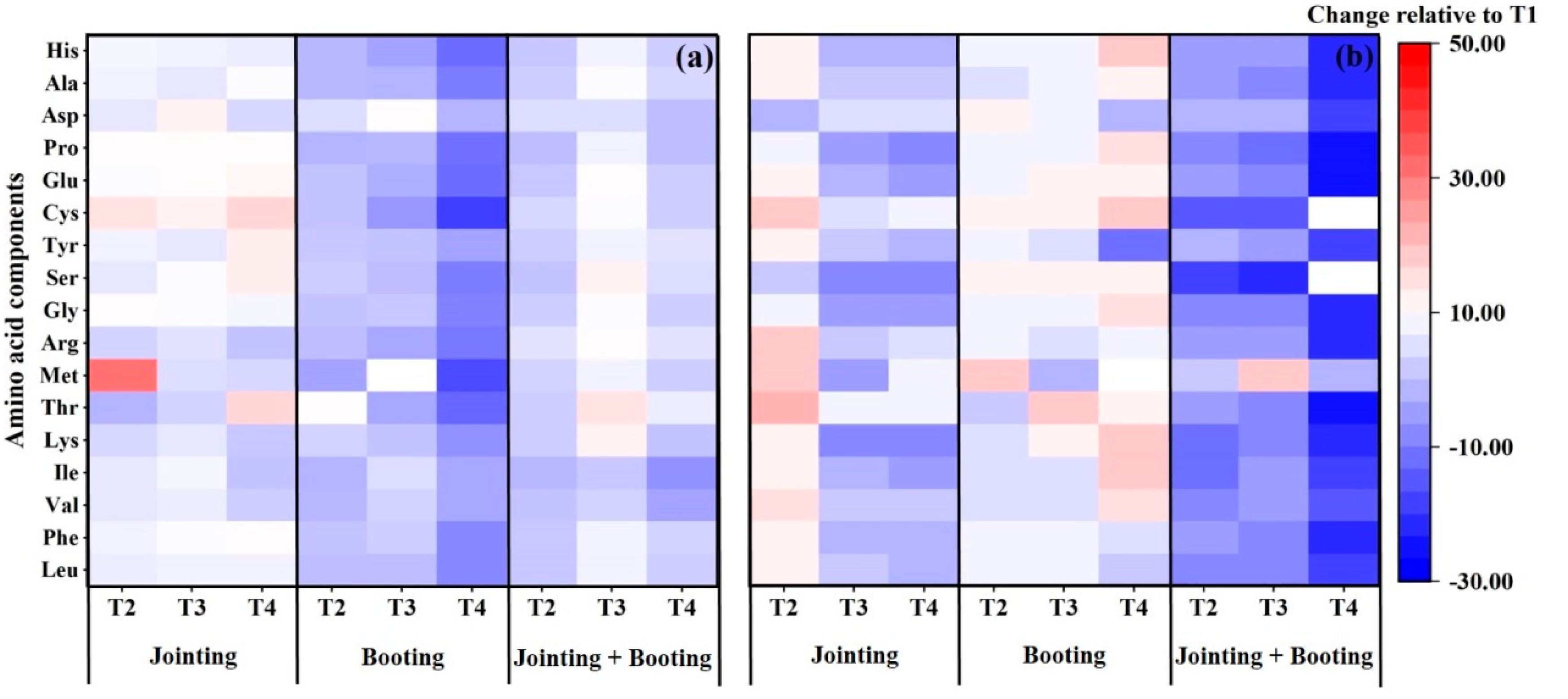
3.2. Effects of Low Temperature on the Amino Acid Composition in Wheat Grain
3.3. Evaluation of the Effect of Low Temperature on the Nutritional Value of Amino Acids in Wheat Grain
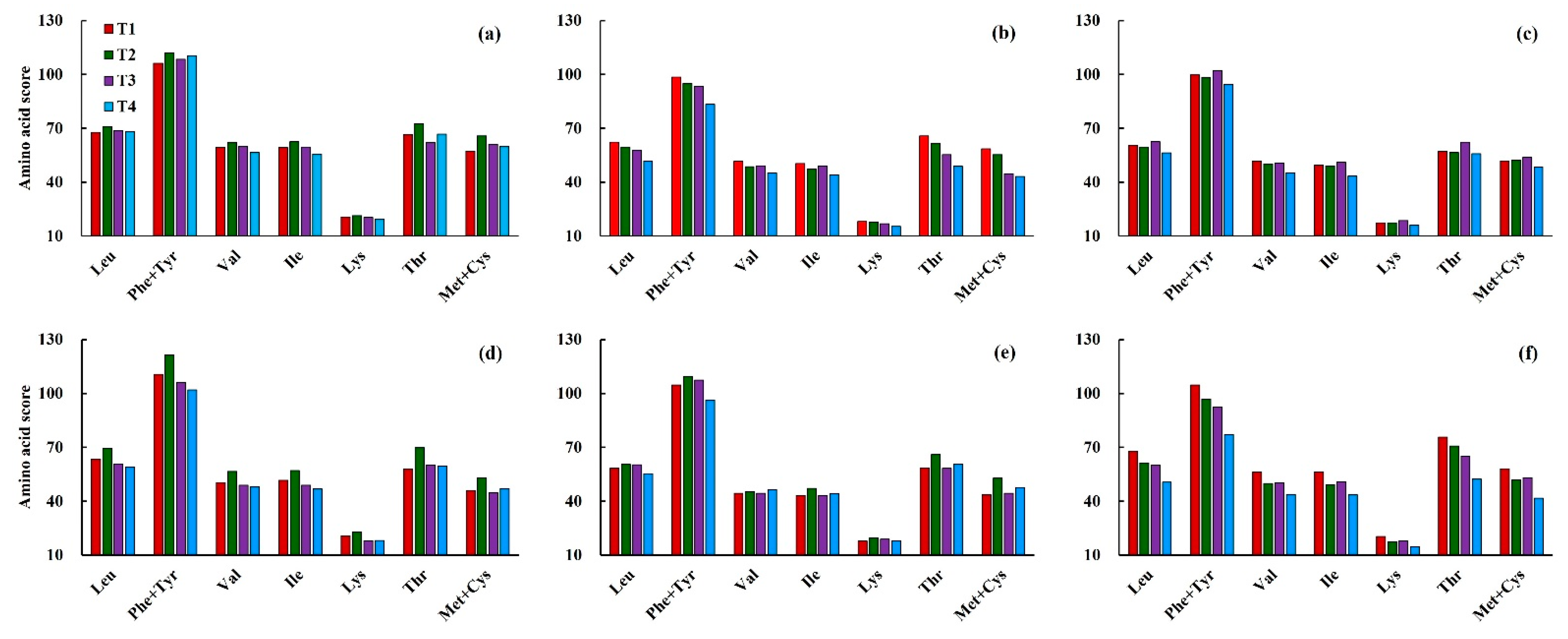
3.3.1. Effects of Low Temperature on Amino Acid Score (AAS) in Wheat Grain
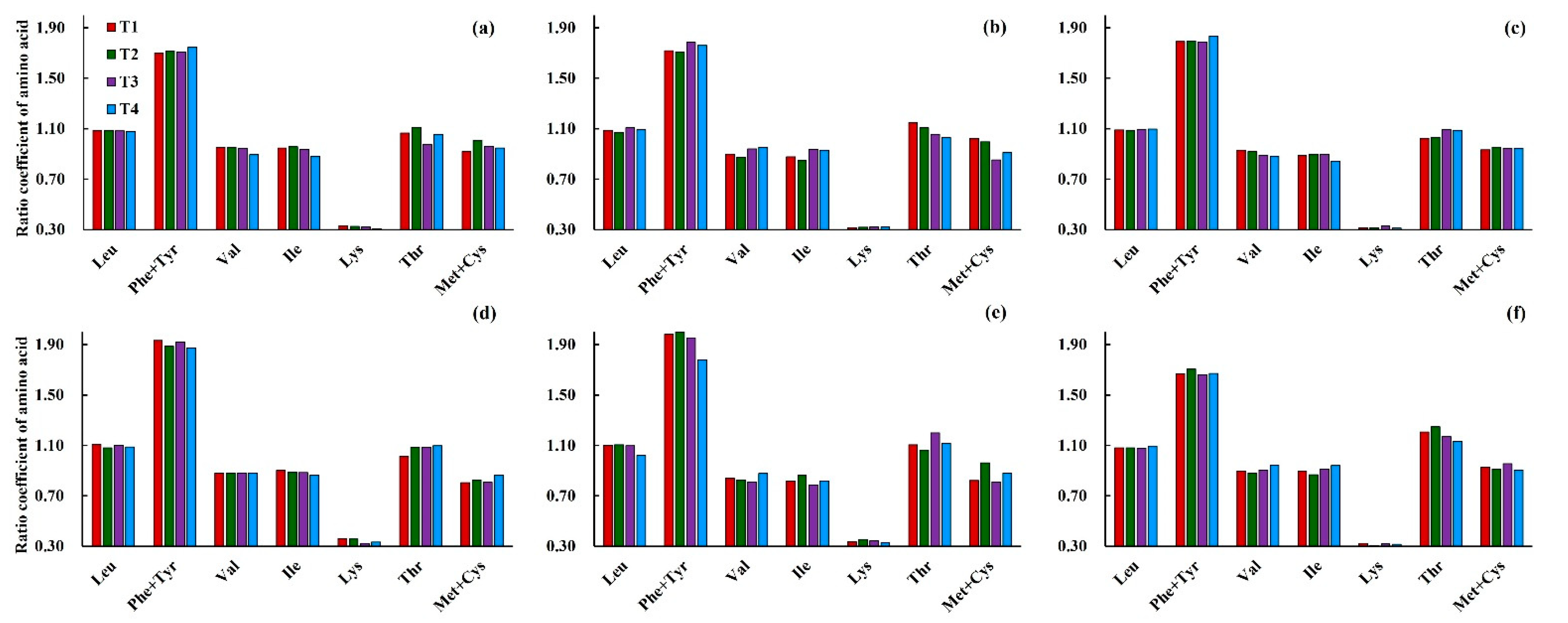
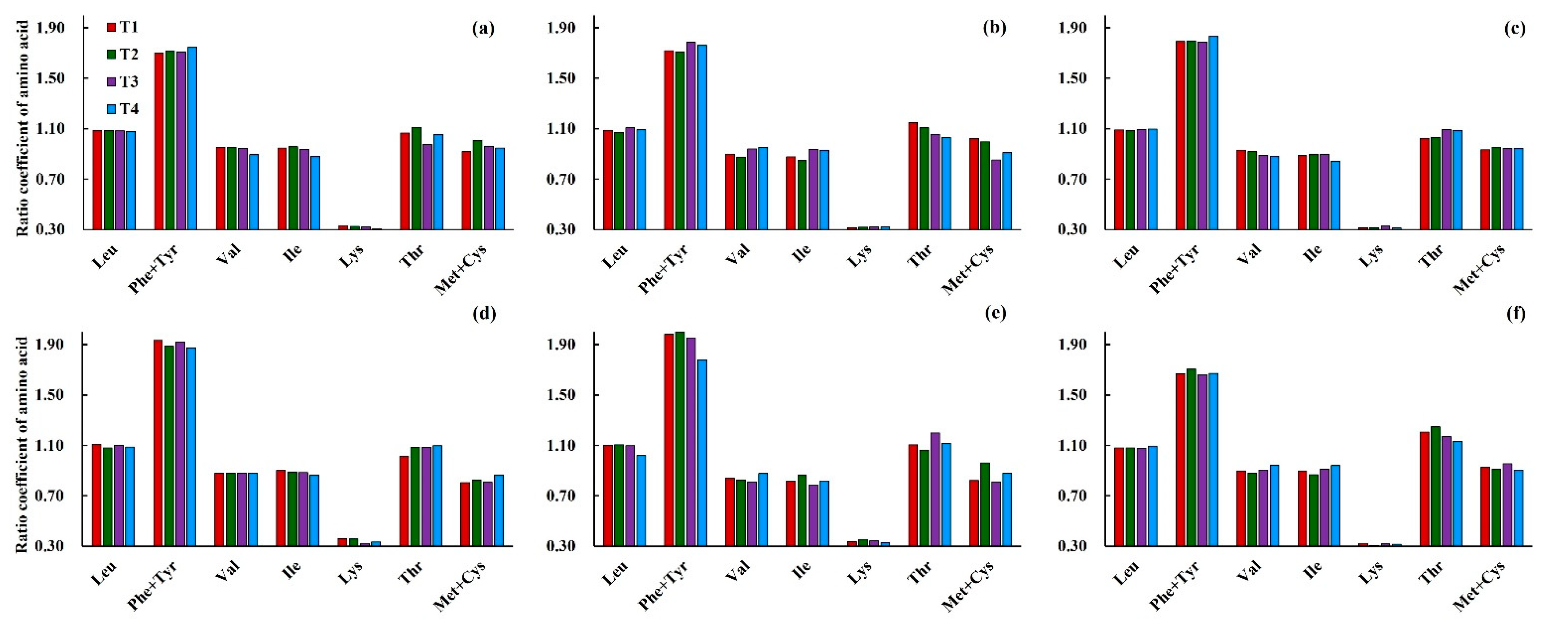
3.3.2. Effects of Low Temperature on the Ratio Coefficient of Amino Acids (RC) in Wheat Grain
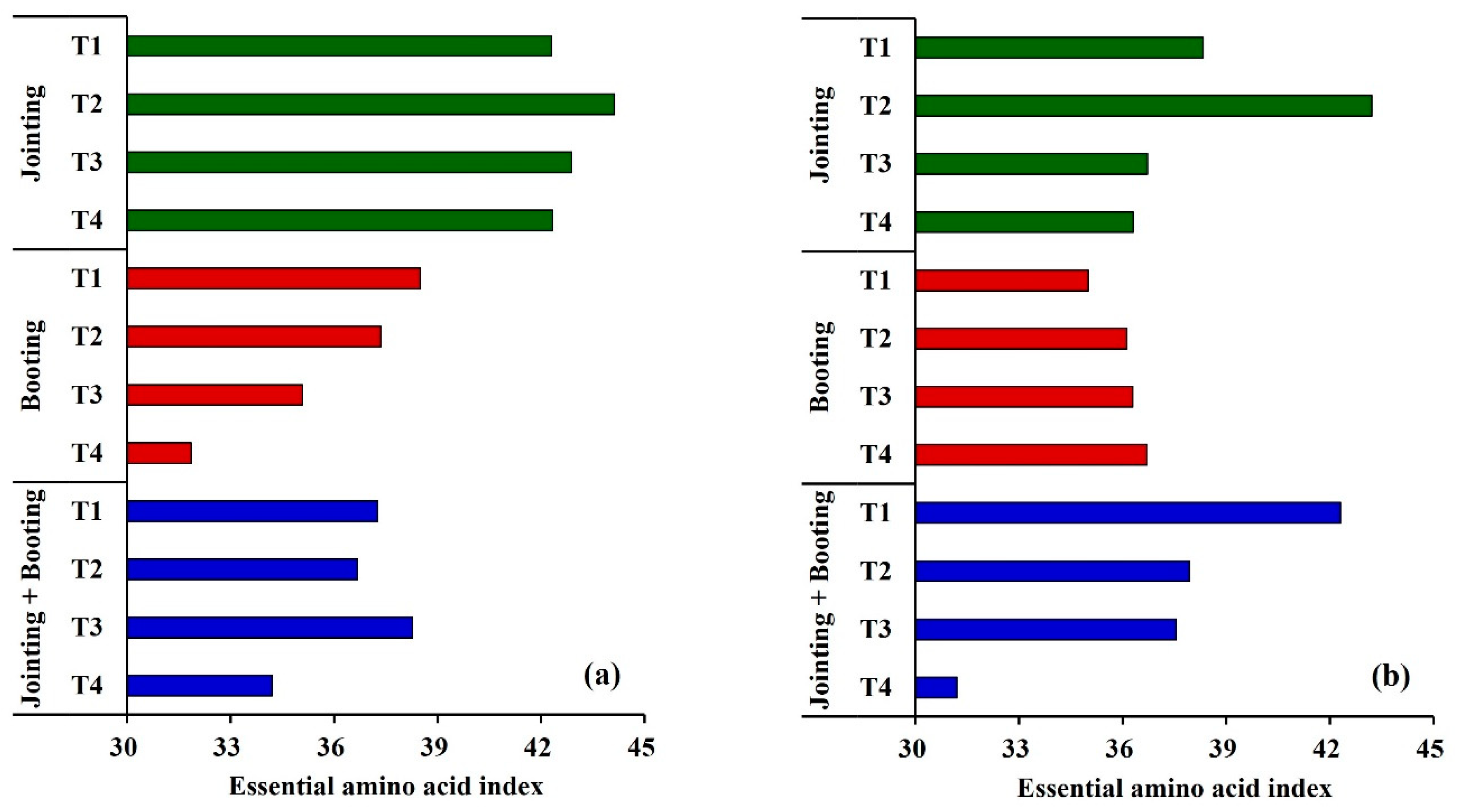
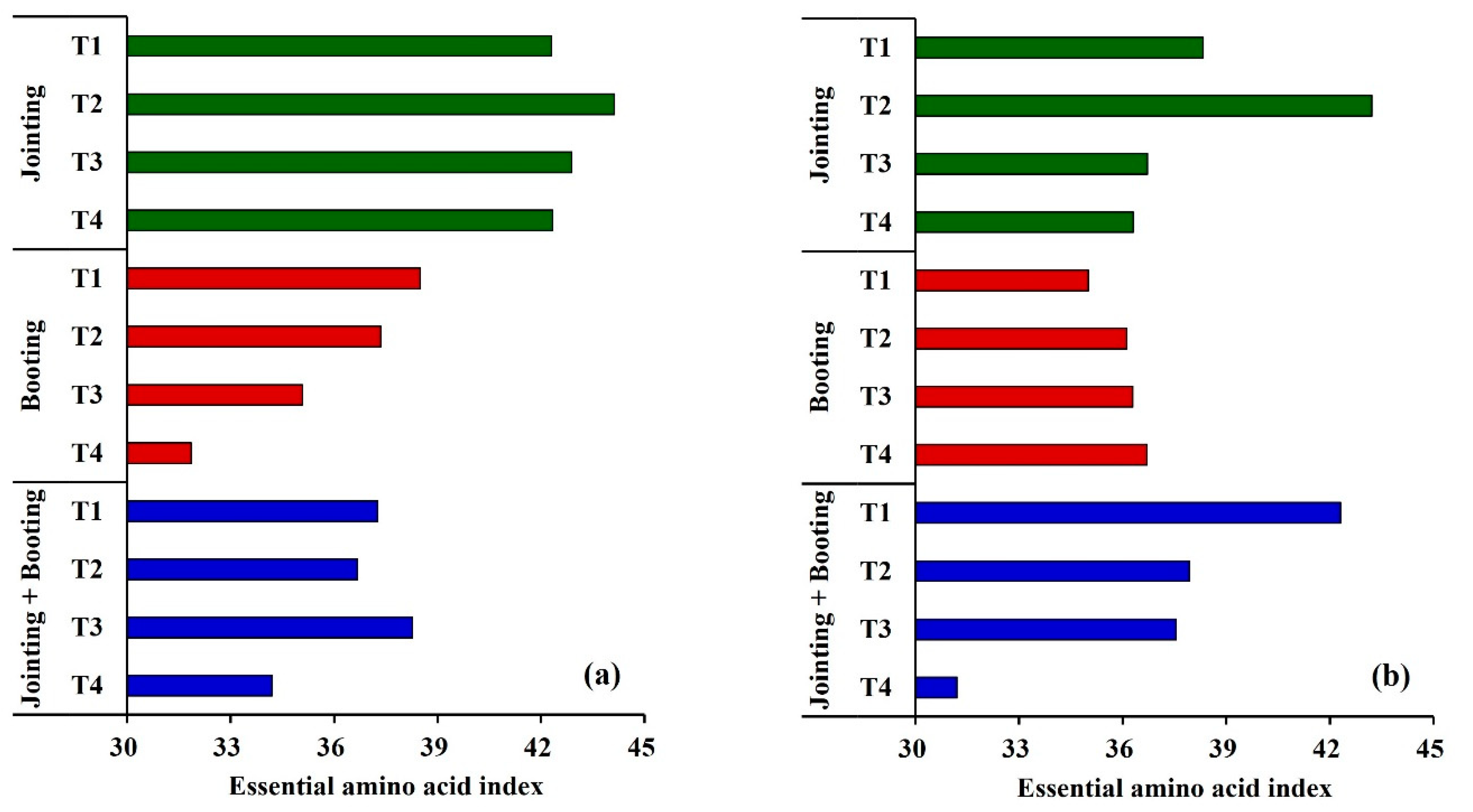
3.3.3. Effects of Low Temperature on the Essential Amino Acid Index (EAAI) of Wheat Grain
4. Discussion and Conclusions
4.1. Responses of Amino Acids in Wheat Grains to Low Temperature
4.2. Comprehensive Evaluation of the Impact of Low Temperature on Nutritional Value of Amino Acids in Wheat
Author Contributions
Funding
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Jood, S.; Kapoor, A.C.; Singh, R. Amino acid composition and chemical ecaluation of protein qualtiy of cereals as affected by insect indestation. Plant Foods Hum. Nutr. 1995, 48, 159–167. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, M.; Cai, J.; Wang, X.; Zhou, Q.; Cao, W.; Dai, T.; Jiang, D. Nitrogen topdressing timing influences the spatial distribution patterns of protein components and quality traits of flours from different pearling fractions of wheat (Triticum aestivum L.) grains. Field Crops Res. 2018, 216, 120–128. [Google Scholar] [CrossRef]
- Boila, R.J.; Stothers, S.C.; Campbell, L.D. The relationship between the concentrations of individual amino acids and protein in wheat and barley grown at selected locations throughout Manitoba. Can. J. Anim. Sci. 1996, 76, 163–169. [Google Scholar] [CrossRef]
- Anjum, F.M.; Ahmad, I.; Butt, M.S.; Sheikh, M.A.; Pasha, I. Amino acid composition of spring wheats and losses of lysine during chapati baking. J. Food Compos. Anal. 2005, 18, 523–532. [Google Scholar] [CrossRef]
- Graciela, C.J.; Francisco, A.V.O.; Maria, I.G.H. Amino acid composition, score and in vitro protein digestibility of foods community consumed in Norhwest Mexico. Nutr. Hosp. 2013, 28, 365–371. [Google Scholar]
- Mladenov, N.; Przulj, N.; Hristov, N.; Djuric, V.; Milovanovic, M. Cultivar-by-environment interactions for wheat quality traits in semiarid conditions. Cereal Chem. 2011, 78, 363–367. [Google Scholar] [CrossRef]
- Peterson, C.J.; Graybosch, R.A.; Baenziger, P.S.; Grombacher, A.W. Genotype and environment effects on quality characteristics of hard red winter wheat. Crop Sci. 1992, 32, 98–103. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined effect of genetic and environmental factors on the accumulation of proteins in the wheat grain and their relationship to bread-making quality. J. Cereal Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- Liu, P.; Guo, W.; Jiang, Z.; Pu, H.; Feng, C.; Zhu, X.; Peng, Y.; Kuang, A.; Little, C.R. Effects of high temprature after anthesis on starch granules in grains of wheat (Triticum aestivum L.). J. Agric. Sci. 2011, 149, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.H.; Lv, Z.Y.; Zhang, Y.; Ma, L.L.; Qin, B.Y.; Liu, Q.X.; Zhang, W.J.; Ma, S.Y.; Ma, C.X.; Huang, Z.L. Pre-anthesis night warming improves post-anthesis physiological activity and plant productivity to post-anthesis heat stress in winter wheat (Triticum aestivum L.). Environ. Exp. Bot. 2022, 197, 104819. [Google Scholar] [CrossRef]
- Xin, Z.H.; Guo, J.P.; Tan, K.Y.; Liu, K.W.; Yang, R.G.; Zhang, L.H.; Sun, Y. Relationship between amino acid quality of winter wheat and meteorological ecological factors. J. Arid Meteoro. 2020, 38, 148–156. [Google Scholar]
- García Del Moral, L.F.; Rharrabti, Y.; Martos, V.; Royo, C. Environmentally induced changes in amino acid composition in the grain of durum wheat grown under different water and temperature regimes in a mediterranean environment. J. Agr. Food Chem. 2007, 55, 8144–8151. [Google Scholar] [CrossRef] [PubMed]
- Labuschagne, M.T.; Elago, O.; Koen, E. The influence of temperature extremes on some quality and starch characteristics in bread, biscuit and durum wheat. J. Cereal Sci. 2009, 49, 184–189. [Google Scholar] [CrossRef]
- Xia, L.J.; Liu, L.L.; Asseng, S.; Xia, Y.M.; Tang, L.; Liu, B.; Cao, W.X.; Zhu, Y. Estimating spring frost and its impact on yield across winter wheat in China. Agr. Forest Meteorol. 2018, 260, 154–164. [Google Scholar] [CrossRef]
- Liu, L.L.; Song, H.; Shi, K.J.; Liu, B.; Zhang, Y.; Tang, L.; Cao, W.X.; Zhu, Y. Response of wheat grain quality to low temperature during jointing and booting stages—On the importance of considering canopy temperature. Agr. Forest Meteorol. 2019, 278, 107658. [Google Scholar] [CrossRef]
- Kodra, E.; Steinhaeuser, K.; Ganguly, A.R. Persisting cold extremes under 21st-century warming scenarios. Geophys. Res. Lett. 2011, 38, L08705. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Q.; Ren, G.Y. Change in extreme temperature event frequency over mainland China, 1961–2008. Clim. Res. 2011, 50, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.Y.; Chapman, S.C.; Christopher, J.T.; Frederiks, T.M.; Chenu, K. Frost trends and their estimated impact on yield in the Australian wheatbelt. J. Exp. Bot. 2015, 66, 3611–3623. [Google Scholar] [CrossRef] [Green Version]
- Asseng, S.; Foster, I.A.N.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Change Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Liu, B.; Liu, L.L.; Tian, L.Y.; Cao, W.X.; Zhu, Y.; Asseng, S. Post-heading heat stress and yield impact in winter wheat of China. Glob. Change Biol. 2014, 20, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Qiu, X.L.; Chen, J.; Cammarano, D.; Ge, Z.L.; Ruane, A.C.; Liu, L.L.; Tang, L.; Cao, W.X.; Liu, B.; et al. Impacts of 1.5 °C and 2.0 °C global warming above pre-industrial on potential winter wheat production of China. Eur. J. Agron. 2020, 120, 126149. [Google Scholar] [CrossRef]
- Triboï, E.; Martre, P.; Triboï-Blondel, A.M. Environmentally-induced changes in protein composition in developing grains of wheat are related to changes in total protein content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Gu, K.J.; Gu, D.X.; Zhang, S.M.; Wu, J.J. Quantifying the effect of low-temperature events on the grain quality formation of wheat. J. Cereal Sci. 2021, 100, 103257. [Google Scholar] [CrossRef]
- Rubin, M.; Schoonouer, D.R.; Bossard, E.H. Amino acid profiles of corn and soybean meal by a modified analysis technique. Poult Sci. J. 1975, 54, 1811–1817. [Google Scholar]
- World Health Organization. Energy and Protein Requirements; World Health Organization: Rome, Italy, 1973. [Google Scholar]
- Björck, I.; Asp, N.G.; Dahlqvist, A. Protein nutritional value of extrusion-cooked wheat flours. Food Chem. 1984, 15, 203–214. [Google Scholar] [CrossRef]
- Bailey, R.L.; West, K.P.; Black, R.E. The epidemiology of global micronutrient deficiencies. Ann. Nutr. Metab. 2015, 66, 22–33. [Google Scholar] [CrossRef]
- Altenbach, S.B. New insights into the effects of high temperature, drought and post-anthesis fertilizer on wheat grain development. J. Cereal Sci. 2012, 56, 39–50. [Google Scholar] [CrossRef]
- Khoury, C.K.; Bjorkman, A.D.; Dempewolf, H.; Ramirez-Villegas, J.; Guarino, L.; Jarvis, A.; Rieseberg, L.H.; Struik, P.C. Increasing homogeneity in global food supplies and the implications for food security. Proc. Natl. Acad. Sci. USA 2014, 111, 4001–4006. [Google Scholar] [CrossRef] [Green Version]
- Lobell, D.B.; Hammer, G.L.; Chenu, K.; Zheng, B.; McLean, G.; Chapman, S.C. The shifting influence of drought and heat stress for crops in northeast Australia. Glob. Change Biol. 2015, 21, 4115–4127. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, D.Z.; Zhang, H.X.; Asseng, S.; Yin, T.W.; Qiu, X.L.; Ye, Z.; Liu, L.L.; Tang, L.; Cao, W.X.; et al. Seperating the impacts of heat stress events from rising mean temperature on winter wheat yield of China. Environ. Res. Lett. 2021, 16, 124035. [Google Scholar] [CrossRef]
- Zheng, D.X.; Yang, X.G.; Mínguez, M.I.; Mu, C.Y.; He, Q.; Wu, X. Effect of freezing temperature and duration on winter survival and grain yield of winter wheat. Agr. Forest Meteorol. 2018, 260, 1–8. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Global Warming of 1.5 °C: An IPCC Special Report on the Impacts Of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context Of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pötner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018. [Google Scholar]
- Muhammar, A.H.; Chen, X.; Muhammad, F.; Muhammad, N.; Zhang, Y.; Xu, H.; Ke, Y.Y.; Attiogbe, K.B.; Zhang, L.L.; Li, J.C. Cold stress in wheat: Plant acclimation responses and management strateies. Front. Plant. Sci. 2021, 12, 676884. [Google Scholar]
- Frederiks, T.M.; Christopher, J.T.; Sutherland, M.W.; Borrell, A.K. Post-head emergence frost in wheat and barley: Defining the problem, assessing the damage, and identifying resistance. J. Exp. Bot. 2015, 66, 3487–3498. [Google Scholar] [CrossRef] [Green Version]
- Kovács, Z.; Simon-Sarkade, L.; Sovány, C.; Kirsch, K.; Galiba, G.; Kocsy, G. Differential effects of cold acclimation and abscisic acid on free amino acid composition in wheat. Plant Sci. 2011, 180, 61–68. [Google Scholar] [CrossRef]
- Xu, Q.; Fan, N.L.; Zhuang, L.L.; Yu, J.J. Enhanced stolon growth and metabolic adjustment in creeping bentgrass with elevated CO2 concentration. Environ. Exp. Bot. 2018, 155, 87–97. [Google Scholar] [CrossRef]
- Serpil, T.; Yasemin, E. Variation of total soluble seminal root proteins of tetraploid wild and cultivated wheat induced at cold acclimation and freezing. Acta Physiol. Plant. 2004, 26, 443–450. [Google Scholar]
- Gomes, F.P.; Oliva, M.A.; Mielke, M.S.; Almeida, A.F.; Aquino, L.A. Osmotic adjustment, proline accumulation and cell membrane stability in leaves of cocos nucifera submitted to drought stress. Sci. Hortic. 2010, 126, 379–384. [Google Scholar] [CrossRef]
- Lu, T.B.; Lv, J.Y.; Lu, H.P.; Zhang, L.S.; Wang, C.F.; Zhang, J.Q.; Zhang, X.T. The relationship of contents of protein and amino acid in winter wheat leaves with cold resistance at the turngreen stage. Acta Agric. Boreali-Occident. Sin. 2009, 18, 56–59. [Google Scholar]
- Zhao, C.X.; Zhang, R.; Niu, K.J.; Zhu, R.T.; Wang, Y.; Ma, X.; Ma, H.L. Metabolomics study of Qinghai wild Poa pratensis in response to low temperature stress. Acta Agrestia Sin. 2020, 28, 904–914. [Google Scholar]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Parida, A.K. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of the halophyte Salvadora persica. Environ. Exp. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Ito, H.; Takaichi, S.; Tsuji, H.; Tanaka, A. Properties of synthesis of chlorophyll a from chlorophyll b in cucumber etioplasts. J. Biol. Chem. 1994, 35, 22034–22038. [Google Scholar] [CrossRef]
- Mokochinski, J.B.; Mazzafera, P.; Sawaya, A.C.H.F.; Mumm, R.; De Vos, R.C.H.; Hall, R.D. Metabolic responses of eucalyptus species to different temperature regimes. J. Integr. Plant Biol. 2018, 60, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, C.H.; Li, F.U.; Li, K.Y.; Yang, N.; Yang, Y.E. Contents of protein and amino acid of wheat grain in different wheat production regions and their evaluation. Acta Agron. Sin. 2016, 42, 768–777. [Google Scholar] [CrossRef]
- Young, V.R.; Pellett, P.L. Plant protein in relation to human protein and amino acid nutrition. Am. J. Clin. Nutr. 1994, 59, 1203S–1212S. [Google Scholar] [CrossRef]
- Millward, D.J. The nutritional value of plant-based diets in relation to human amino acid and protein requirements. Proc. Nutr. Soc. 1999, 58, 249–260. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Cultivar | Sowing Date | Treatment Stage | Start and End Dates of Low-Temperature Treatment (m/dd) | Duration (days) |
|---|---|---|---|---|---|
| 2017–2018 | Yangmai16 | 26 October | Jointing | 3/06–3/08 | 3 |
| Booting | 3/27–3/29 | 3 | |||
| Jointing and Booting | 3/06–3/08 and 3/27–3/29 | 3 and 3 | |||
| Xumai30 | Jointing | 3/18–3/20 | 3 | ||
| Booting | 4/08–4/10 | 3 | |||
| Jointing and Booting | 3/18–3/20 and 4/08–4/10 | 3 and 3 | |||
| 2018–2019 | Yangmai16 | 1 November | Jointing | 3/06–3/08 | 3 |
| Booting | 3/29–3/31 | 3 | |||
| Jointing and Booting | 3/06–3/08 and 3/29–3/31 | 3 and 3 | |||
| Xumai30 | Jointing | 3/18–3/20 | 3 | ||
| Booting | 4/08–4/10 | 3 | |||
| Jointing and Booting | 3/18–3/20 and 4/08–4/10 | 3 and 3 |
| Cultivar | Source of Variation | Essential Amino Acid | Nonessential Amino Acid | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leu | Phe | Val | Ile | Lys | Thr | Met | Arg | Gly | Ser | Tyr | Cys | Glu | Pro | Asp | Ala | His | ||
| Yangmai16 | T | ns | ns | ns | ns | * | ns | ns | ns | ** | ns | ns | ns | * | * | ns | ns | ns |
| S | * | ** | * | * | ns | ns | ns | ** | * | * | ** | ns | ** | * | ns | * | ns | |
| S × T | ns | ns | ns | ns | ns | ** | ns | ns | * | ** | ns | ** | ** | * | ns | ** | * | |
| Xumai30 | T | ns | ns | ns | * | ns | ns | ns | ns | ns | * | ** | ns | ns | ns | ns | ns | ns |
| S | ns | ns | ** | ** | ns | ** | ** | ns | ns | ** | ** | ns | * | ns | ** | ns | ** | |
| S × T | ns | ns | * | * | ns | ns | ns | ns | * | ** | ns | * | ns | * | ns | ns | * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, X.; Ma, J.; Qian, W.; Cao, Y.; Zhang, Y.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Effects of Low Temperature on the Amino Acid Composition of Wheat Grains. Agronomy 2022, 12, 1171. https://doi.org/10.3390/agronomy12051171
Hu X, Ma J, Qian W, Cao Y, Zhang Y, Liu B, Tang L, Cao W, Zhu Y, Liu L. Effects of Low Temperature on the Amino Acid Composition of Wheat Grains. Agronomy. 2022; 12(5):1171. https://doi.org/10.3390/agronomy12051171
Chicago/Turabian StyleHu, Xinyi, Jifeng Ma, Weihao Qian, Yuan Cao, Yu Zhang, Bing Liu, Liang Tang, Weixing Cao, Yan Zhu, and Leilei Liu. 2022. "Effects of Low Temperature on the Amino Acid Composition of Wheat Grains" Agronomy 12, no. 5: 1171. https://doi.org/10.3390/agronomy12051171
APA StyleHu, X., Ma, J., Qian, W., Cao, Y., Zhang, Y., Liu, B., Tang, L., Cao, W., Zhu, Y., & Liu, L. (2022). Effects of Low Temperature on the Amino Acid Composition of Wheat Grains. Agronomy, 12(5), 1171. https://doi.org/10.3390/agronomy12051171