
Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance



Abstract
1. Introduction
2. Material and Methods
2.1. Collection and Identification
2.2. Light Microscopy
2.3. Scanning Electron Microscopy
2.4. Quantitative Analysis
3. Results and Discussion
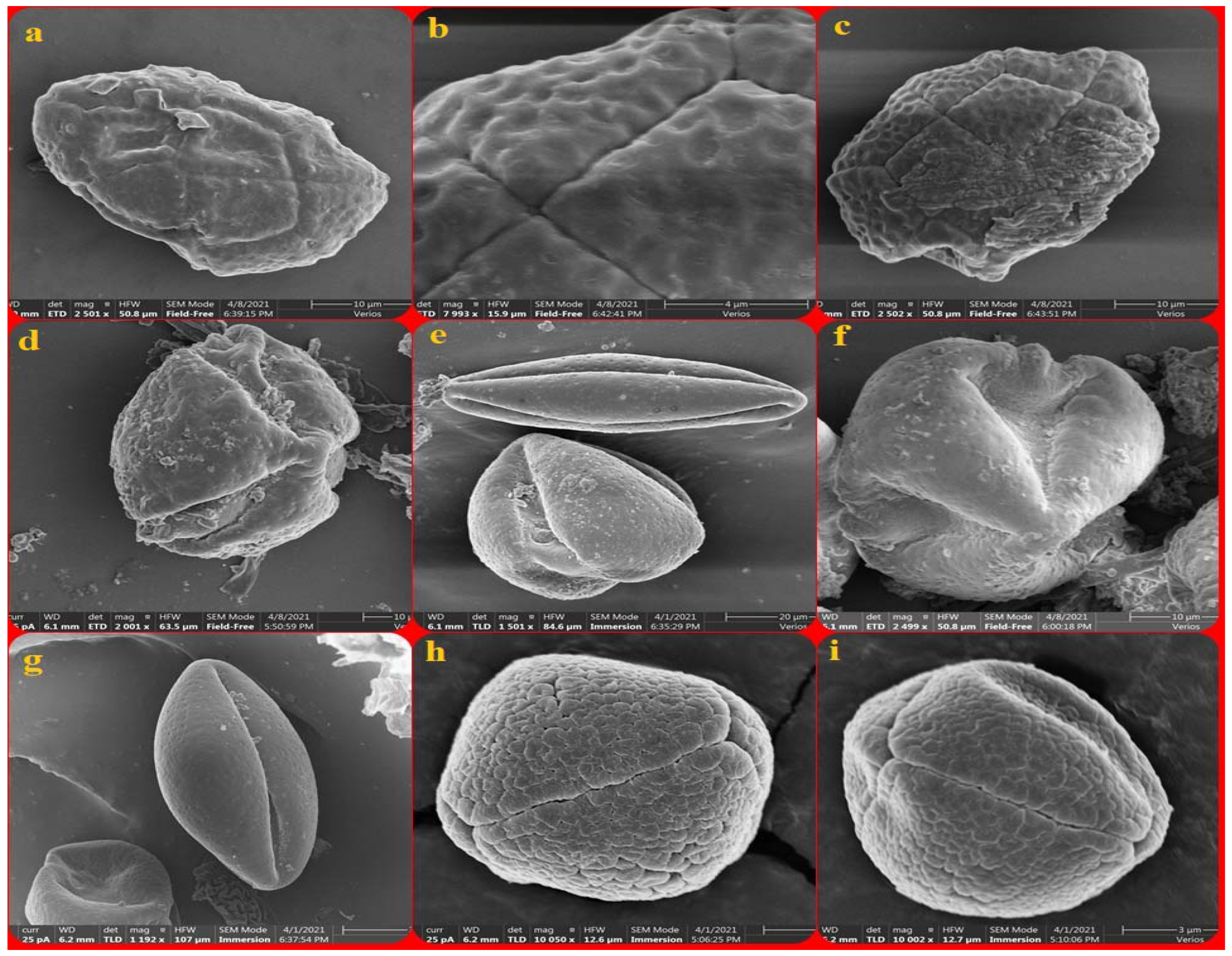
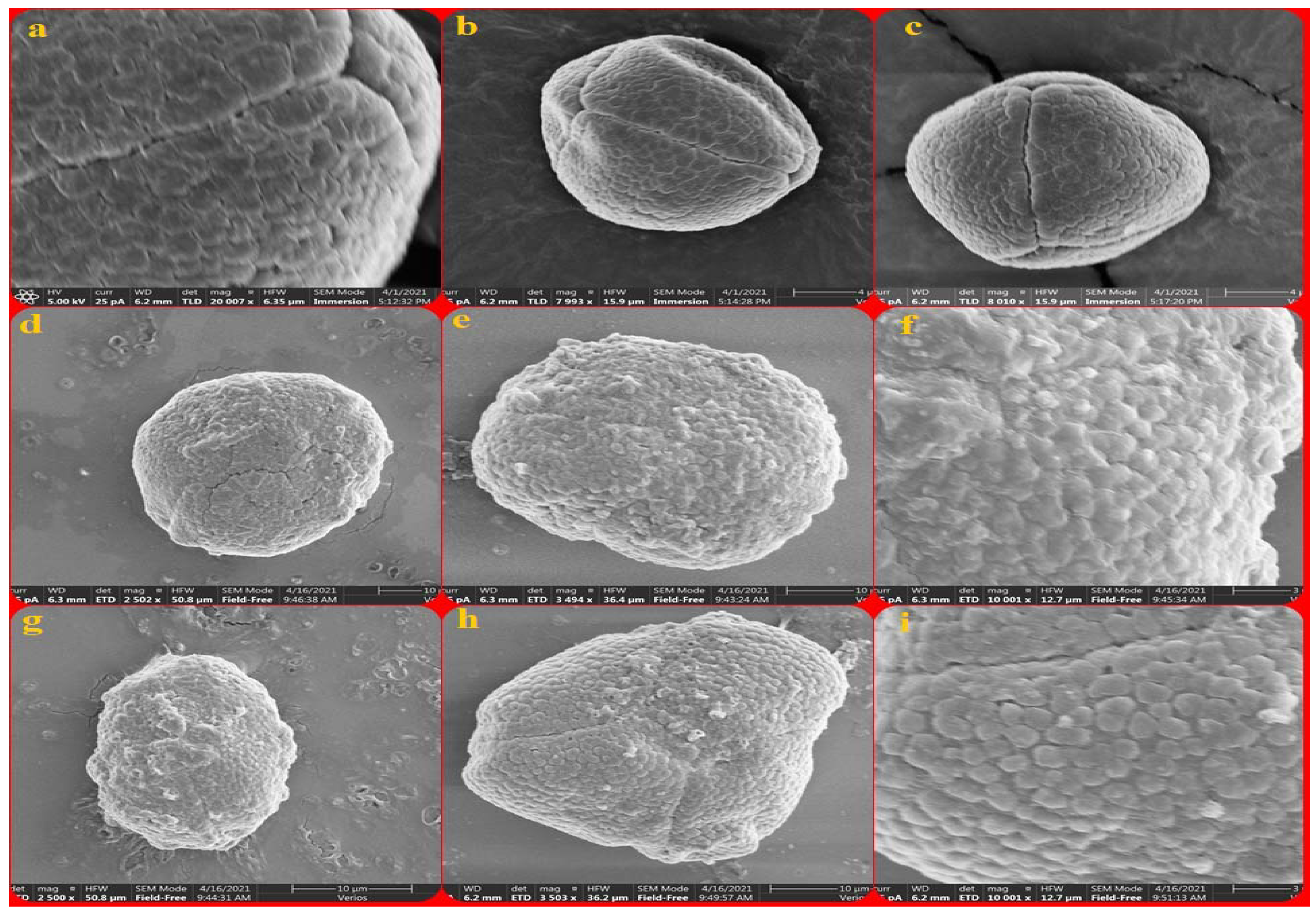
3.1. Acacia auricifoliformis
3.2. Acacia confusa
3.3. Acacia mangium
3.4. Albizia julibrissin
3.5. Albizia lebback
3.6. Calliandra haematocephala
3.7. Entada phaseoloides
3.8. Leucaena leucocephala
3.9. Mimosa bimucronata
3.10. Mimosa diplotricha
3.11. Mimosa pudica
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, G.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World; The Bath Press (CPI Group): The Royal Botanic Gardens, Kew, UK, 2005; p. 577. [Google Scholar]
- Legume Phylogeny Working Group. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny. Taxon 2017, 66, 44–47. [Google Scholar] [CrossRef]
- Shinwari, Z.K.; Jamil, K.; Zahra, N.B. Molecular systematics of selected genera of subfamily Mimosoideae-Fabaceae. Pak. J. Bot. 2014, 46, 591–598. [Google Scholar]
- Polhill, R.M. Evolution and systematics of the Leguminosae. Advences Legume Syst. 1981, 1, 1–26. [Google Scholar]
- Vural, C.; Ekici, M.; Akan, H.; Aytaç, Z. Seed morphology and its systematic implications for genus Astragalus L. sections Onobrychoidei DC., Uliginosi Gray and Ornithopodium Bunge (Fabaceae). Plant Syst. Evol. 2008, 274, 255–263. [Google Scholar] [CrossRef]
- Fawzi, N.M. Macro-and micromorphological seed characteristics of some selected species of Caesalpinioideae-Leguminosae. Res. J. Bot. 2011, 6, 68–77. [Google Scholar] [CrossRef]
- Mabberley, D.J. The Plant-Book: A Portable Dictionary of the Vascular Plants; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Murphy, D.J.; Brown, G.K.; Miller, J.T.; Ladiges, P.Y. Molecular phylogeny of Acacia Mill. (Mimosoideae: Leguminosae): Evidence for major clades and informal classification. Taxon 2010, 59, 7–19. [Google Scholar] [CrossRef]
- Caccavari, M.; Dome, E. An Account of Morphology and Structural Characterization of American Mimosoideae Pollen. Part I: Tribe Acaceae. Palynology 2000, 24, 231–248. [Google Scholar] [CrossRef]
- Hutchinson, J. The Genera of Flowering Plants. Dicotylédones; Oxford University Press: London, UK, 1964; Volume 1. [Google Scholar]
- Panicker, K.T.C.; Sreedevi, P. Studies in the pollen morphology of Mimosaceae monad, tetrad and octad taxa. J. Palynol. 2004, 40, 9–21. [Google Scholar]
- Costa, J.A.S.; Nunes, T.S.; Ferreira, A.P.L.; Stradmann, M.T.S.; Queiroz, L.P. Leguminosasforrageiras da Caatinga: Espéciesimportantespara as Comunidadesrurais do Sertão da Bahia; UniversidadeEstadual de Feira de Santana, SASOP: Feira de Santana, Brazil, 2002; pp. 1–8. [Google Scholar]
- Souza, R.S.O. Jurema-preta (Mimosa tenuiflora (Willd.)Poir.): A review of its traditional use, phytochymistry and pharmacology. Braz. Arch. Biol. Technol. 2008, 51, 937–947. [Google Scholar] [CrossRef]
- Ahmad, F.; Anwar, F.; Hira, S. Review on medicinal importance of Fabaceae family. Pharmacologyonline 2016, 3, 151–157. [Google Scholar]
- Rahman, A.H.M.M.; Parvin, M.I.A. Study of medicinal uses on Fabaceae family at Rajshahi, Bangladesh. Res. Plant Sci. 2014, 2, 6–8. [Google Scholar]
- Molares, S.; Ladio, A. The usefulness of edible and medicinal Fabaceae in Argentine and Chilean Patagonia: Environmental availability and other sources of supply. Evid. Based Complement Alternat. Med. 2012, 2012, 901918. [Google Scholar] [CrossRef]
- Turnbull, J.W. (Ed.) Australian vegetation. In Multipurpose Australian Trees and Shrubs: Lesser-Known Species for Fuelwood and Agroforestry; Australian Centre for International Agricultural Research: Canberra, Australia, 1986; Volume 14, pp. 29–44. [Google Scholar]
- Sornsathapornkul, P.; Owens, J.N. Pollination Biology in a Tropical Acacia Hybrid (A. mangium Willd. × A. auriculiformis A. Cunn. ex Benth.). Ann Bot. 1998, 81, 631–645. [Google Scholar] [CrossRef][Green Version]
- Joseph, B.; George, J.; Mohan, J. Pharmacology and traditional uses of Mimosa pudica. Int. J. Pharm. Sci. Drug Res. 2013, 5, 41–44. [Google Scholar]
- Mambou, A.H.M.Y.; Bila, R.B.; Wanyu, B.Y.; Nkwemeh, C.N.; Jugha, V.T.; Toukam, L.L.; Bopda, O.S.M.; Taiwe, G.S.; Bum, E.N. Antifatigue properties of an aqueous extract of Mimosa pudica Linn. (Fabaceae) in mice subjected to weight loaded force swimming test. GSC Biol. Pharma. Sci. 2022, 18, 224–233. [Google Scholar] [CrossRef]
- Arasu, M.V.; Al-Dhabi, N.A.; Choi, K.C.; Bensy, A.D.V.; Rajaselvam, J. Bioactive potential of Albizia lebbeck extract against phytopathogens and protective properties on tomato plant against speck disease in greenhouse. Phy. Mol. Plant Patho. 2022, 117, 101750. [Google Scholar] [CrossRef]
- Bahadur, S.; Ahmad, M.; Mir, S.; Zafar, M.; Sultana, S.; Ashfaq, S.; Arfan, M. Identification of monocot flora using pollen features through scanning electron microscopy. Microsc. Res. Tech. 2018, 81, 599–613. [Google Scholar] [CrossRef]
- Ali, M.; Liu, Y.J.; Xia, Q.P.; Bahadur, S.; Hussain, A.; Shao, J.W.; Shuaib, M. Pollen micromorphology of eastern Chinese Polygonatum and its role in taxonomy by using scanning electron microscopy. Microsc. Res. Tech. 2021, 84, 1451–1461. [Google Scholar] [CrossRef]
- Ashfaq, S.; Zafar, M.; Ahmad, M.; Sultana, S.; Bahadur, S.; Khan, A.; Shah, A. Microscopic investigations of palynological features of convolvulaceous species from arid zone of Pakistan. Microsc. Res. Tech. 2018, 81, 228–239. [Google Scholar] [CrossRef]
- Ashfaq, S.; Ahmad, M.; Zafar, M.; Sultana, S.; Bahadur, S.; Ahmed, S.N.; Gul, S.; Nazish, M. Pollen morphology of family Solanaceae and its taxonomic significance. An. Acad. Bras. Cienc. 2020, 92, e20181221. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Ahmad, M.; Zafar, M.; Gul, S.; Shuaib, M.; Butt, M.A.; Hanif, U.; Nizamani, M.M.; Hussain, F.; et al. Authentication of the therapeutic Lamiaceae taxa by using pollen traits observed under scanning electron microscopy. Microsc. Res. Tech. 2022, 84, 1–19. [Google Scholar] [CrossRef]
- Bahadur, S.; Taj, S.; Long, W.; Ahmad, M. Pollen morphology and its implication in the taxonomy of some selected taxa of the bi and tri-ovulate Euphorbiaceae of the Hainan Island by using multiple microscopic techniques. Microsc. Res. Tech. 2022, 26, 1–16. [Google Scholar] [CrossRef]
- Butt, M.A.; Zafar, M.; Ahmad, M.; Kayani, S.; Bahadur, S.; Ullah, F.; Khatoon, S. The use of taxonomic studies to the identification of wetlands weeds. Adv. Weed Sci. 2021, 39, e222645. [Google Scholar] [CrossRef]
- Hameed, A.; Zafar, M.; Ahmad, M.; Sultana, S.; Bahadur, S.; Anjum, F.; Shuaib, M.; Taj, S.; Irm, M.; Altaf, M.A. Chemo-taxonomic and biological potential of highly therapeutic plant Pedicularis groenlandica Retz. using multiple microscopic techniques. Microsc. Res. Tech. 2021, 84, 2890–2905. [Google Scholar] [CrossRef]
- Javed, S.; Javaid, A.; Hanif, U.; Bahadur, S.; Sultana, S.; Shuaib, M.; Ali, S. Effect of necrotrophic fungus and PGPR on the comparative histochemistry of Vigna radiata by using multiple microscopic techniques. Microsc. Res. Tech. 2021, 84, 2737–2748. [Google Scholar] [CrossRef]
- Nabila; Ahmad, M.; Zafar, M.; Bahadur, S.; Sultana, S.; Taj, S.; Celep, F.; Majeed, S. Palynomorphological diversity among the Asteraceous honeybee flora: An aid to the correct taxonomic identification using multiple microscopic techniques. Microsc. Res. Tech. 2022, 85, 570–590. [Google Scholar] [CrossRef]
- Ullah, F.; Ahmad, M.; Zafar, M.; Parveen, B.; Ashfaq, S.; Bahadur, S.; Safdar, Q.; Bin Safdar, L.; Alam, F.; Luqman, M. Pollen morphology and its taxonomic potential in some selected taxa of Caesalpiniaceae observed under light microscopy and scanning electron microscopy. Microsc. Res. Tech. 2021, 85, 1410–1420. [Google Scholar] [CrossRef]
- Zaman, W.; Ahmad, M.; Zafar, M.; Amina, H.; Lubna; Ullah, F.; Bahadur, S.; Ayaz, A.; Saqib, S.; Begum, N.; et al. The quest for some novel antifertility herbals used as male contraceptives in district Shangla, Pakistan. Acta Ecol. Sin. 2020, 40, 102–112. [Google Scholar] [CrossRef]
- Rasool, S.; Faheem, M.; Hanif, U.; Bahadur, S.; Taj, S.; Liaqat, F.; Pereira, L.; Liaqat, I.; Shaheen, S.; Shuaib, M.; et al. Toxicological effects of the chemical and green ZnO NPs on Cyprinus carpio L. observed under light and scanning electron microscopy. Microsc. Res. Tech. 2021, 85, 848–860. [Google Scholar] [CrossRef]
- Rubab, S.; Rizwani, G.H.; Bahadur, S.; Shah, M.; Alsamadany, H.; Alzahrani, Y.; Shuaib, M.; Hershan, A.; Hobani, Y.H.; Shah, A.A. Determination of the GC–MS analysis of seed oil and assessment of pharmacokinetics of leaf extract of Camellia sinensis L. J. King Saud Univ. Sci. 2020, 32, 3138–3144. [Google Scholar] [CrossRef]
- Park, J.M.; Song, U.S. Pollen morphology of the genus Rhododendron (Ericaceae) in Korea. J. Korean Soc. Forest Sci. 2010, 99, 663–672. [Google Scholar]
- Sorsa, P. Pollen morphological studies on the Mimosaceae. Ann. Bot. Fenn. 1969, 6, 1–34. [Google Scholar]
- Guinet, P.H. Les Mimosacees, Etude de palynologiefundamentale, correlations, evolution. Inst. Fr. Pondichery Trav. Sci. Tech. 1969, 9, 1–293. [Google Scholar]
- Guinet, P. Mimosoideae: The characters of their pollen grains. In Advances in Legume Systematics; Polhill, R.R., Raven, P.H., Eds.; The Bath Press (CPI Group): The Royal Botanic Gardens, Kew, UK, 1981; Volume 2, pp. 835–855. [Google Scholar]
- Khan, S.U.; Zafar, M.; Ahmad, M.; Anjum, F.; Sultana, S.; Kilic, O.; Ozdemir, F.A.; Nazir, A.; Yaseen, G.; Aabidin, S.Z.U. Pollen micromorphological analysis of tribe Acacieae (Mimosaceae) with LM and SEM techniques. Microsc. Res. Tech. 2019, 82, 1610–1620. [Google Scholar] [CrossRef]
- Khan, S.U.; Zafar, M.; Ullah, R.; Shahat, A.A.; Ahmad, M.; Sultana, S.; Malik, K. Pollen diversity and its implications to the systematics of mimosaceous species by LM and SEM. Microsc. Res. Tech. 2021, 84, 42–55. [Google Scholar] [CrossRef]
- El Ghazali, G.E.; Satti, A.M.; Tsuji, S.I. Intra-specific pollen polymorphism in Mimosa pigra (Mimosaceae). Grana 1997, 36, 279–283. [Google Scholar] [CrossRef]
- Kordofani, M.; Ingrouille, M. Geographical variation in the pollen of Acacia (Mimosaceae) in Sudan. Grana 1992, 31, 113–118. [Google Scholar] [CrossRef]
- Perveen, A.; Qaiser, M. Pollen Flora of Pakistan-XI. Leguminosae (Subfamily: Mimosoideae). Turk J Botany. 1998, 22, 151–156. [Google Scholar]
- Aftab, R.; Perveen, A. A palynological study of some cultivated trees from Karachi. Pak. J. Bot. 2006, 38, 15–28. [Google Scholar]
- Hughes, C.E. Variation in anther and pollen morphology in Leucaena Benth. (Leguminosae-Mimosoideae). Bot. J. Linn. Soc. 1997, 123, 177–196. [Google Scholar] [CrossRef]
- De Assis Ribeiro dos Santos, F.; de Oliveira Romão, C. Pollen morphology of some species of Calliandra Benth. (Leguminosae-Mimosoideae) from Bahia, Brazil. Grana 2008, 47, 101–116. [Google Scholar] [CrossRef]
- Zhi-Min, L.I. Studies on the Pollen Morphology of Some Genera in Mimosaceae from China. Plant Divers. 1994, 16, 1–3. [Google Scholar]
- Simon, M.F.; Grether, R.; Queiroz, L.P.; Särkinen, T.E.; Dutra VF Hughes, C.E. The evolutionary history of Mimosa (Leguminosae): Toward a phylogeny of the sensitive plants. Am. J. Bot. 2011, 98, 1201–1221. [Google Scholar] [CrossRef]
- Long, W.-X.; Ding, Y.; Zang, R.-G.; Yang, M.; Chen, S.-W. Environmental characteristics of tropical cloud forests in the rainy season in Bawangling National Nature Reserve on Hainan Island, South China. Chin. J. Plant Ecol. 2011, 35, 137. [Google Scholar] [CrossRef]
- Long, W.; Yang, X.; Li, D. Patterns of species diversity and soil nutrients along a chronosequence of vegetation recovery in Hainan Island, South China. Ecol. Res. 2012, 27, 561–568. [Google Scholar] [CrossRef]
- Wang, X.X.; Long, W.X.; Yang, X.B.; Xiong, M.H.; Kang, Y.; Huang, J.; Wang, X.; Hong, X.J.; Zhou, Z.L.; Lu, Y.Q.; et al. Patterns of plant diversity within and among three tropical cloud forest communities in Hainan Island. Chin. J. Plant Ecol. 2016, 40, 469. [Google Scholar]
- Punt, W.; Hoen, P.P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Erdtman, G. Pollen Morph Ology and Plant Taxonomy. Angiosperms; Chronica Botanica Co.: Waltham, MA, USA, 1952; Volume 539, p. 261. [Google Scholar]
- Hyde, H.A.; Adams, K.F. An Atlas of Airborn Pollen Grains; Macmillan & Co.: London, UK, 1958; 112p. [Google Scholar]
- Brown, G.K.; Murphy, D.J.; Miller, J.T.; Ladiges, P.Y. Acacia ss and its relationship among tropical legumes, tribe Ingeae (Leguminosae: Mimosoideae). Syst Bot. 2008, 33, 739–751. [Google Scholar] [CrossRef]
- Wodehouse, R.P. Preparation of pollen for microscopic examination. Bull. Torrey Bot. Club. 1933, 60, 417–421. [Google Scholar] [CrossRef]
- Wang, X.J.; Cao, X.L.; Hong, Y. Isolation and characterization of flower-specific transcripts in Acacia mangium. Tree Physiol. 2005, 25, 167–178. [Google Scholar] [CrossRef]
- Agashe, S.N.; Caulton, E. Pollen and Spores: Applications with Special Emphasis on Aerobiology and Allergy, 1st ed.; CRC Press: Boca Raton, FL, USA, 2009; p. 412. [Google Scholar] [CrossRef]
- Van Campo, M.; Guinet, P. Les pollen compose’s. L’exempledeˆsMimosace´es. Pollen Spores 1961, 8, 201–218. [Google Scholar]
- Chen, Y.-Y. Studies on the development of polyad grains of Calliandra haematocephala Hassk. with electron microscopic technique. Taiwania 1973, 17, 18–28. [Google Scholar]
- Rao, A.N.; Lee, Y.K. Studies on Singapore pollen. Pac. Sci. 1970, 24, 255–268. [Google Scholar]
- Santos-Silva, J.; Simon, M.; Tozzi, A.M.G.D.A. Pollen diversity and its phylogenetic implications in Mimosa ser. Leiocarpae Benth. (Leguminosae, Mimosoideae). Grana 2013, 52, 15–25. [Google Scholar] [CrossRef]
- Caccavari, M.A. Granos de polen de las leguminosas de la Argentina IV. Genero Mimosa. Bol. Soc. Argent. Bot. 1985, 24, 151–167. [Google Scholar]
- Caccavari, M.A. Estudio de los caracteres del polen en las MimosaLepidotae. Pollen Spores 1986, 28, 29–42. [Google Scholar]
- Caccavari, M.A. 1987. Study of pollen grains in Mimosa Glanduliferae. In Proceedings of the VII Argentine Symposium on Paleobotany and Palynology, Buenos Aires, Argentina, 30 August 2007; pp. 141–145. [Google Scholar]
- Caccavari, M.A. Ultraestructura del polen de Mimosa (Mimosoideae-Leguminosae). Pollen Spores 1988, 30, 275–296. [Google Scholar]
- Caccavari, M.A. Pollen morphology and structure of tropical and subtropical American genera of the Piptadenia-group (Leguminosae: Mimosoideae). Grana 2022, 41, 130–141. [Google Scholar] [CrossRef]
- Medina-Acosta, M.; Grether, R.; Martínez-Bernal, A.; Ramírez-Arriaga, E. Comparative study of pollen morphology and exine ultrastructure in tetrads, octads and polyads of the genus Mimosa (Leguminosae). Palynology 2019, 43, 188–212. [Google Scholar] [CrossRef]
- Jamwal, R. Palynological investigations on some selected bee forage plants of family fabaceae using light and scanning electron microscopy from himachal pradesh, India. Plants Arch. 2021, 21, 1047–1053. [Google Scholar] [CrossRef]
- Kenrick, J.; Knox, R.B. Function of the polyad in reproduction of Acacia. Ann. Bot. 1982, 50, 721–727. [Google Scholar] [CrossRef]
- Rajurkar, A.V.; Tidke, J.A.; Jadhav, S.S. Palynomorphological Studies on Family Mimosaceae. Int. J. Pharma. Bio. Sci. 2013, 4, 273–279. [Google Scholar]
- Guinet, P.; Hernández, H.M. Pollen characters in the genera Zapoteca and Calliandra (Leguminosae, Mimosoideae), their systematic and phylogenetic relevance. Pollen Spores 1989, 31, 5–22. [Google Scholar]
- Thulin, M.; Guinet PHunde, A. Calliandra (Leguminosae) in continental Africa. Nord. J. Bot. 1981, 1, 27–34. [Google Scholar] [CrossRef]
- Niezgoda, C.J.; Feuer, S.M.; Nevling, L.I. Pollen ultrastructure of the tribe Ingeae (Mimosoideae-Leguminosae). Am. J. Bot. 1983, 70, 650–667. [Google Scholar] [CrossRef]
- Hernández, H.M.; Sousa, S.M. Two new species of Calliandra (Leguminosae: Mimosoideae) from Southern Mexico. Syst. Bot. 1988, 13, 519–524. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Altitude | Sea Level |
|---|---|---|---|
| Acacia auriculiformis A.Cunn. ex Benth. | Sanya | N: 18.26415 E: 109. 52084 | 119 m |
| Acacia confusa Merr. | Wanning, Botanical Garden, Haikou | N: 18.69396 E:110.23170 | 118 m |
| Acacia mangium Willd. | Sanya, Chang Jiang | N: 19.33905 E: 108.210 | 33 m |
| Albizia julibrissin Durazz. | Wanning | N: 18.69396 E:110.23170 | 118 m |
| Albizia lebbeck (L.) | Haikou | N: 20.062363 E: 110.3186713 | 3 m |
| Calliandra haematocephala Hassk. | Sanya | N: 18.26415 E: 109.52084 | 119 m |
| Entada phaseoloides (Linn.) Merr. | Wanning, Botanical Garden | N: 18.69396 E:110.23170 | 118 m |
| Leucaena leucocephala (Lam.) | Haikou, Sanya | N: 20.062363 E: 110.3186713 | 3 m |
| Mimosa bimucronata (DC.) Kuntze | Haikou, Wanning | N: 18.69396 E:110.23170 | 118 m |
| Mimosa diplotricha C. Wright | Beihualing, Haikou | N:19.00262 E: 109.81523 | 471 m |
| Mimosa pudica Linn. | Haikou, Wanning, Sanya, Chang Jiang | N: 19.33905 E: 108.210 | 33 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahadur, S.; Taj, S.; Long, W.; Hanif, U. Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance. Agronomy 2022, 12, 1122. https://doi.org/10.3390/agronomy12051122
Bahadur S, Taj S, Long W, Hanif U. Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance. Agronomy. 2022; 12(5):1122. https://doi.org/10.3390/agronomy12051122
Chicago/Turabian StyleBahadur, Saraj, Sehrish Taj, Wenxing Long, and Uzma Hanif. 2022. "Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance" Agronomy 12, no. 5: 1122. https://doi.org/10.3390/agronomy12051122
APA StyleBahadur, S., Taj, S., Long, W., & Hanif, U. (2022). Pollen Morphological Peculiarities of Selected Mimosoideae Taxa of Hainan Island and Their Taxonomic Relevance. Agronomy, 12(5), 1122. https://doi.org/10.3390/agronomy12051122