1. Introduction
Italian ryegrass (
Lolium multiflorum Lam.) is one of the most productive and valuable grass species in field forage cropping. It is usually divided into short-lived annual or biennial (Westerwolds) cultivars and cultivars able to persist for several years, with the latter being widely used by Croatian farmers. In the humid regions of the USA (central Pennsylvania), a study conducted on two highly productive commercial dairy farms in [
1] found that tetraploid Italian ryegrass persisted well for 4 years in a mixture with lucerne (
Medicago sativa L.). Consequently, the authors pointed out that “annual” nomenclature commonly used for Italian ryegrass in the USA could be misleading. Under favourable growing conditions of the Willamette Valley in Oregon,
Lolium perenne L. seed production fields can maintain economically viable production for 3 or 4 years [
2,
3]. Multiyear seed production of Italian ryegrass crop is also of interest for growers, but previous research focused on the first-production year only (e.g., [
4,
5,
6]). Thus, research on best management practices and operations in multiyear cropping systems for seed production of Italian ryegrass is warranted.
Rainfall coupled with winds favour lodging and seed shattering of Italian ryegrass crop, which is commonly observed in farmer fields in north-western Croatia. Lodging is ubiquitous for ryegrasses and [
7] reported that the severity of lodging in perennial ryegrass (
Lolium perenne L.) was remarkably similar among nine environments in Oregon’s Willamette Valley, characterised by conditions that ranged from drought to excessive precipitation. Previous research [
8] found that the seed yield of a
Lolium perenne crop, supported mechanically to prevent lodging, was higher than that of a lodged crop. An abundant supply of water and N following lodging facilitate light penetration to lower buds in
Lolium perenne seed grass, and consequently, a flush of new tillers during this period [
9]. They are generally unwanted in seed production because they compete with developing seed for assimilate. Furthermore, the growth of new tillers (secondary regrowth) may lead to difficulties in harvesting such as seed yield losses due to no-cutting of lodged stems, and increased moisture of threshed material. Various management strategies such as the application of plant growth regulator (PGR) and early spring grazing or forage cut have demonstrated great potential in reducing the incidence of lodging in grass seed yields [
6,
10]. However, spring cutting for forage removes the first-formed tillers and leaves seed crop to be produced by late-formed tillers of lower yield and quality. The tolerance of perennial ryegrass to management systems incorporating grazing and/or cutting in early spring and N application is well-documented. For example, Hill and Watkin [
11] showed that the application of N to grazed plots fully overcame the depression in ear length found in unfertilized plots. However, limited information is available about the effect of spring cutting dates for forage on seed yield from secondary growth in Italian ryegrass crop. Inclement weather is usually the main reason for Croatian farmers to delay the date of the first forage cut in spring.
In addition to lodging, a second undesirable feature of Italian ryegrass is its inability to retain seed until the time of harvest. Thus, Italian ryegrass is commonly direct combined at higher seed moisture content compared to perennial ryegrass [
12]. The main characteristic of grass seed production is brought about by prolonged periods of tillering and by a considerable range in the size and position of the tillers at harvest. Adverse weather conditions just before harvest and differences in maturity (moisture content) between seeds of the same ear or between plants are the major factors accounting for large seed losses due to shattering. In
Lolium perenne [
13], seed shedding is unlikely to be the major factor in seed yield losses through shattering because the terminal floret in each spikelet (those which have the lowest actual percentage of florets which set seed) are the first to shed their seeds. However, in glasshouse conditions, Lombardy populations of
Lolium multiflorum had lost approximately 30% of their seed at seed moisture concentration of 43%, which increases to 40% at a moisture concentration of just under 40% [
14]. In a companion paper [
15], it was found that the first-year crop of Italian ryegrass may shed large amounts of seeds (up to 800 kg ha
−1) before harvest by direct combining. The main objective of this study was to assess the impact of various management systems (single- and dual-purpose crops) and N fertilization rates on the performance of second-year seed crop of Italian ryegrass.
2. Materials and Methods
The management systems consisted of the single-purpose (pure-seed) and the dual-purpose (combined forage-seed) crops. The single-purpose crops consisted of system management without (Seed-crop) and with (SeedPGR-crop) PGR (trinexapac-ethyl) application. The dual-purpose crops had early (EF-seed-crop) and late (LF-seed-crop) spring forage cut followed by seed harvest. Nitrogen fertilization was applied at the total rates of 0, 60, 120, and 180 kg ha
−1. A second-year crop of Italian ryegrass was grown during the growing season of 2012/2013 with weather data presented in
Table 1 at the Maksimir experimental field (45°49′ N, 16°2′ E, 123 m above sea level) on silt loam soil (Eutric Cambisol) with trial design, management practices, and operations for the first-production year crop described earlier [
15]. There was almost no herbage regrowth following the seed harvest of the first-crop year, and, therefore, no data are presented. In October of 2012, plots were defoliated to a height of 7 cm to remove early autumn vegetative growth (cleaning cut). Plots were freed of weeds, diseases, and pests throughout experimentation.
Granular N fertilization, as calcium ammonium nitrate (27% N), was top broadcast in split applications with dates of application and corresponding crop growth stages shown in
Table 2. Plant growth regulator Moddus EC 250 (trinexapac-ethyl) was applied with backsprayer at a dose of 1.0 L ha
−1 (250 g a.i. L
−1) at ZCK 32–33 [
16] in the SeedPGR-crop (
Table 2). The spray volume used in the PGR application was 200 L ha
−1. For plots with forage cut management, the early spring forage cut date was during stem elongation stage (ZCK 32–33) on 3 May 2013, while the late forage cut was on 14 May 2013 when plants were at the beginning of heading (ZCK 52–53). Plants were cut at a height of 7 cm and forage and dry matter yields were determined by sampling the central 6 rows of the plot on each day of cutting. Growing degree days among various crop growth stages were calculated from the data of a weather station located 300 m from the experimental site using a base temperature of 0 °C.
The total number of ears per unit area was based on the sample that was taken 10 days after full anthesis (ZCK 65) from each plot at a 30 cm length of two adjacent rows where all tillers were cut off at ground level. The total number of ears in each sample was recorded, and, afterward, the sample was divided into the vegetative (unproductive) tillers (no ears), early-formed reproductive (fertile) tillers, and late-formed reproductive tillers (greenheads). The difference between the late-formed reproductive tillers (greenheads) and the early-formed reproductive tillers was that the former had ears that have not yet begun anthesis (ZCK 61), while the latter ears were in a more developed stage. In the early- and late-formed reproductive tillers, the following analyses were carried out: measurement of the stem length to the ear; counting the number of visible nodes on the stem, number of spikelets per ear; and the number of florets per spikelet in the lower, middle, and upper section of ear. Paired spikelets from the lower, middle, and upper sections of each ear were selected for the determination of the number of florets per spikelet at these positions within the ear.
One week after anthesis, two tin containers (15.0 cm in width, 60.0 cm in length, and 5.0 cm in depth) with an overall surface area of 1800 cm2 were placed on the ground between the adjacent central rows of each plot to determine the weight of shed seed. Containers were removed at harvest and the weight of shed seed and the seed moisture content were determined. Crop lodging severity was assessed visually and scored as a percentage of plot lodged where 0 is not lodged (plants are fully upright) and 100% is the most severe lodging (plants are lying flat on the ground). Lodging was scored daily until harvest. The six 30 cm sections in the row of each plot were clipped at ground level on harvest days to determine the above-ground biomass and harvest index. The plants were clipped during morning hours with minimal disturbance to maximally prevent the shattering of seed from ears. The harvest index was calculated as the ratio of clean seed yield to above-ground biomass.
The crop was harvested by direct combining when seed moisture content reached approximately 40–45%. Mature grain was harvested with a small plot combine (Wintersteiger, Ried im Innkreis, Austria) with drum speed and concave settings to simulate commercial farm practice. Direct combining is not usually advisable at moisture concentrations above 400 g kg
−1, but seed crop of tetraploid Italian ryegrass can be successfully combined directly, starting at around 45%, as long as combine drum speeds are adjusted correctly [
17]. At the harvest, only 6 central rows were harvested from the middle of each plot, and plants at the 0.3 m length were cut and removed from both ends of plots to avoid the border effect. Harvest dates are given in
Table 2. After harvest, samples were taken for determination of seed moisture content for harvested and shed seed. Natural seed yield was dried at room temperature for several weeks and then cleaned of impurities to determine clean seed weight (yield). In this paper, seed yield and seed shattering are expressed with 14% moisture content. Seed moisture contents were determined after drying at 60 °C for 48 h. No measurable seed production was achieved in the third-crop year so that data are not presented, and the field experiment was discontinued.
A thousand seeds’ weight was determined by twice counting 200 seeds and then weighed. Germination was determined approximately 90 days after harvest in accordance with the International Seed Testing Association (ISTA) rules [
18] by placing 100 seeds to imbibe on a moist germination paper. After a chilling treatment for five days at 5 °C, the seeds were germinated at 20 °C. The final seedling count was made after 14 days.
A two-factorial experiment (management systems and N fertilization) was conducted on biannual Italian ryegrass crops for seed production. The field trial was arranged in a strip-plot design with four replicates. The crop management treatments were randomized to the main plots within replicates. Nitrogen fertilization was assigned to the sub-plots. The data were analysed using Mixed Model procedures in SAS/STAT Software [
19]. The analysis of variance was computed with crop management and N fertilization was considered fixed. The means separation was calculated using the Fisher’s Least Significant Difference (LSD) test at
p ≤ 0.05.
4. Discussion
A companion paper [
15] showed that the first-year crop of Italian ryegrass shattered large amounts of seeds before the directly combined harvest during June and July of 2012, depending on the management system. In no-till cropping systems, Maia et al. [
20] found that Italian ryegrass seed banks had a low persistence rate, and its viability in the soil was variable. However, under favourable conditions, shattered seed may germinate, and voluntary seedlings can develop into a very dense stand in the next season (i.e., second-year crop). In our research, seeds shattered on the soil surface failed to germinate before October 2012, and voluntary seedlings with one or two developed leaves (ZCK 11–12) were observed in the early November of 2012. This delayed germination of shed seed might have been due to seed dormancy. Previous research [
21] reported a high level of post-harvest dormancy for Italian ryegrass as germination counts taken 3 months after harvest were significantly higher than those taken earlier and there were no differences among various cultivars or harvesting dates. In addition, it is known that the establishment of cool-season grasses is the least effective using the oversowing method (e.g., [
22]), which is comparable to natural seed shattering (self-seeding) in a multiyear crop of Italian ryegrass for seed production. In pastures of the southern Great Plains of the United States, between 885 and 5650 ears m
−2 are required for annual ryegrass to achieve a minimum rate of 500 established seedlings m
−2 through self-seeding [
23]. In our companion paper [
15], the density of reproductive tillers of the first-year crop averaged 664 per square meter only. In addition, delayed germination of shattered seed could have been associated with unfavourable weather conditions following the harvest of first-year crop. Low precipitation coupled with warm temperatures during August and early- and mid-September (
Table 1) might delay the germination of shattered seed until the first autumn rain in late September. However, voluntary seedlings from shed seed were killed by low temperatures (up to −16 °C with no snow cover) during December of 2012. Although ryegrasses are not as winter-hardy as many other grasses, no visible freezing was observed on old plants established from standard sowing practice in the previous year. Thus, it appears that post-harvest seed dormancy coupled with the lack of soil moisture delayed germination and these late-emerging voluntary seedlings did not have sufficient time to harden for winter survival. Winter kill of voluntary seedlings was probably associated with their growing point above the soil surface. In Atlantic Canada [
22], it was found that the poor persistence of Italian ryegrass due to winter kill may limit its commercial production after the establishment year. Using an artificial freezing test (glycol freezing tank), Eagles et al. [
24] reported that the lethal temperatures for the hardened whole plant of
Lolium multiflorum at ZCK 12–13 were from −9.2 °C to −9.8 °C, depending on tested cultivars. As expected, forage yields were higher in the LF-seed-crop when compared to the EF-seed-crop due to the later spring cutting date for the former (
Table 2). Forage yield increments associated with higher N fertilization rates were consistent in the LF-seed-crop (
Figure 1). However, forage yields did not differ between the two highest N rates in the EF-seed-crop; clearly indicating that factor other than N fertilization was more limiting for vegetative crop growth.
In contrast to the first-year crop responses [
15], a significant management system × N fertilization interaction existed for seed yield and most measured traits so that discussion is focused on interaction effects. Italian ryegrass grown under various system managements had to be directly combined at various harvest dates, but seed moisture content was within the optimum window (
Table 3) and averaged 44.6%. The large seeds are more susceptible to mechanical damage at harvest and therefore require lower drum speed. In the LF-seed-crop, seed moisture content was similar at all N levels probably because of the hottest weather conditions around harvest (
Table 1), which facilitate high daily moisture loss. Moisture content can be expected to fall by 1 to 3% per day in good conditions [
14]. The SeedPGR-crop had higher seed moisture content than the Seed-crop at all fertilization levels (
Table 3), which we could not compare to the findings of other researchers. In most field experiments with Italian ryegrass for seed production, the crop is either hand-harvested [
10,
22] or cut with a forage plot harvester [
6,
7,
21,
25], put in jute bags, and dried. A seed moisture concentration of approximately 40% is quoted by [
26] as suitable for cutting Italian ryegrass for later picking up by combine when seed moisture drops to about 12%.
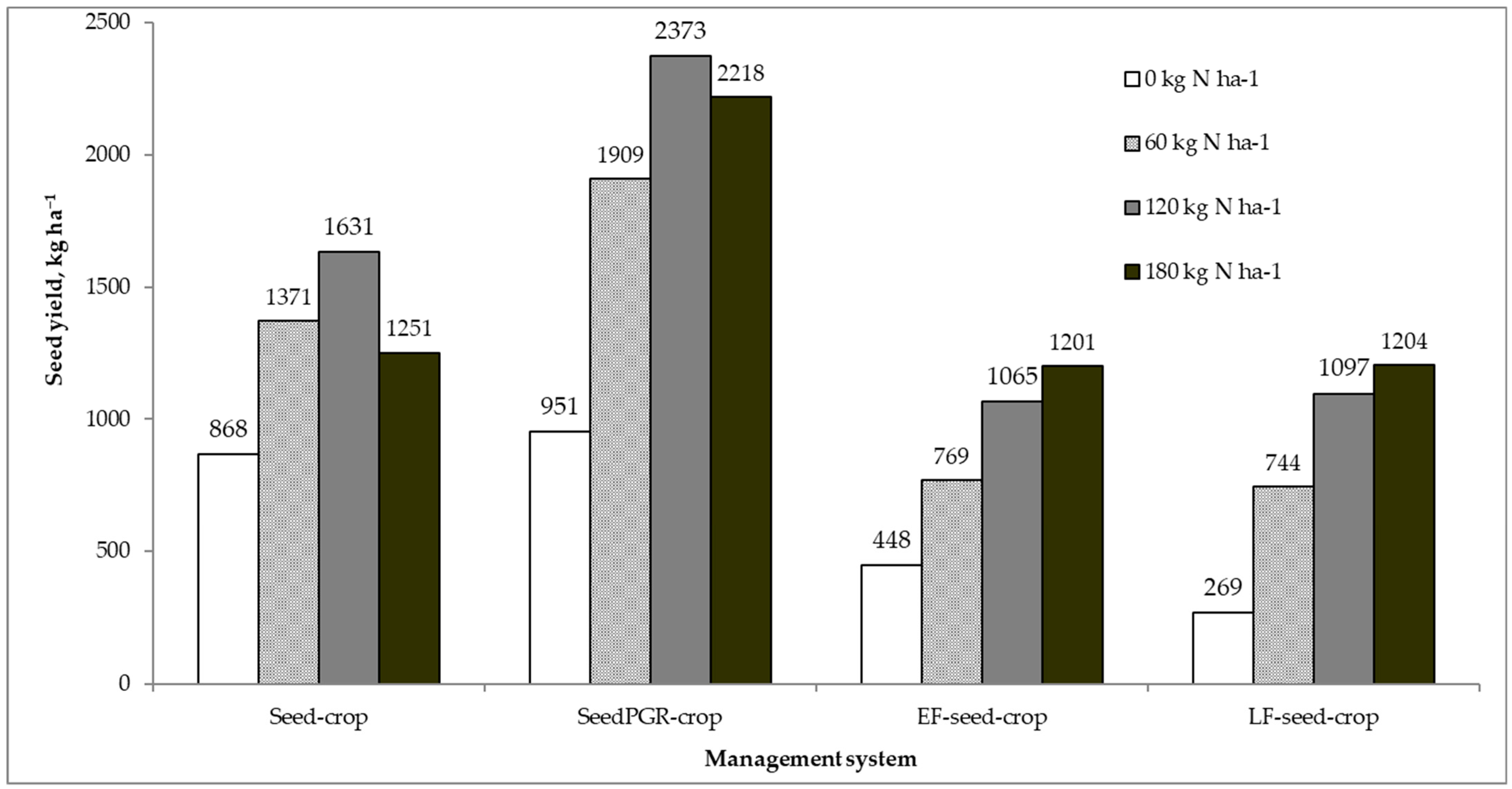
Unfertilized plots yielded 868 kg ha
−1 in the Seed-crop (
Figure 2) that had no lodging at harvest (
Table 4) and seed shattering was relatively low (64 kg ha
−1). In Atlantic Canada, Westerwolds ryegrass without applied N did not produce seed crop [
22]. Despite an associate increase in lodging and seed shattering, the application of N fertilization rate of 60 kg ha
−1 significantly increased yield in the Seed-crop (
Figure 2). This yield increment was primarily due to more early- and late-formed fertile tillers (ears) per unit area, which had more spikelets per ear and florets per spikelet in various ear sections (
Table 3), as well as increased seed weight (
Table 4). The improved number of fertile tillers following spring application of N fertilizer is well-documented. For example, it is stated that the seed yield of forage grasses depend strongly on the number of ears and that early-formed tillers are largely responsible for producing fertile ears [
27]. In our research, early-formed inflorescences consistently had more spikelets per ear and flowers per spikelet when compared to late-formed ones (
Table 3). Seed weight increases in lodged perennial ryegrass crop were previously reported [
4], which is similar to our findings.
Seed-crop yielded maximally (1631 kg ha
−1) at 120 kg N ha
−1, which was primarily associated with the highest density of reproductive tillers (702 per square meter) and the largest above-ground biomass (11,526 kg ha
−1). In contrast, Elgersma [
28] reported that the seed yield of nine cultivars of perennial ryegrass was not associated with the ear number or total matter yield of the seed crop. However, Koeritz et al. [
29] found that vegetative biomass production was positively correlated with seed yield in perennial ryegrass except for situations where lodging was severe. In Serbia, the maximum seed yield (1095 kg ha
−1) of the first-year Italian ryegrass crop was produced at a relatively low N fertilization of 50 kg ha
−1 [
5]. In Belgium, seed yields increased between 0, 60, and 90 kg N ha
−1, but no differences were found between 90, 120, and 150 kg N ha
−1 [
6]. In comparison to 120 kg N ha
−1, the highest N fertilization brought about a significant yield reduction by 26% (380 kg ha
−1) in the Seed-crop (
Figure 2). There were no differences in seed shedding at the two highest N fertilization (
Table 4), and, consequently, yield reduction in the Seed-crop at the highest N rate was attributed to severe and early occurring lodging (
Table 2). Seed yield responses to lodging in grass crops are sometimes difficult to interpret because lodging may often occur on several occasions with varying degrees of severity during the growing season. Moreover, the effects of lodging depend on the growth stage of the crop at the time of lodging, and on subsequent weather conditions.
Lolium perenne crops receiving recommended amounts of N for optimum seed yield usually begin to lodge at about the first ear emergence, and lodging is usually severe by anthesis [
30]. Following a couple of rainy days with strong winds, the onset of lodging in our experiment was also just before heading (
Table 2), when plots fertilized with 180 kg N ha
−1 completely lodged (97.3%). This early-season lodging resulted in shading and thus, the above-ground biomass at harvest was significantly smaller in the Seed-crop grown at 180 kg N ha
−1 compared to Seed-crop at 120 kg N ha
−1 (
Table 4). It was suggested that the physiological explanation of lodging effects on yield lies in its effects on crop growth rate because the less-favourable distribution of light reduces post-lodging crop growth rate. For example, the decrease in final total dry weight resulting from lodging was found on wheat (
Triticum aestivum L.) by [
31] with these reductions being equal to or more than the reductions in grain yield. In our study, Italian ryegrass crop lodged before heading failed to right itself, though some small degree of re-erection of the upper part of culms with such early lodging was observed (data not shown). Interestingly, the reduction in above-ground biomass in the Seed-crop at the highest N fertilization was found despite the abundant growth of new vegetative tillers (
Table 4), and, consequently, the Seed-crop had the largest number of vegetative tillers at harvest. Other authors such as [
32] found that the growth of new tillers after the onset of anthesis had no effect on seed yield in perennial ryegrass. However, in our study, high number of vegetative tillers in the Seed-crop grown at 180 kg N ha
−1 resulted in the smallest proportion of ear-bearing tillers in the total tiller number (46.5%) and low harvest index (12.5%). Thus, vegetative growth most likely competed with reproductive growth for the supply of assimilates, resulting in decreased seed yields. In perennial ryegrass, Koeritz et al. [
29] found that lodging reduced harvest index. In addition, the loss of seed yield due to lodging is likely to be greater when normal farmer field-harvesting procedures are used, because our small plot combine-harvester gathered most lodged ears in our field experiment. Compared with N rate of 120 kg ha
−1, earlier and more severe lodging in the Seed-crop occurring at the highest N fertilization had no effect on the number of reproductive tillers and the 1000-seed weight (
Table 4), which indicated that differences in seed yields were associated with seed number per unit area. In wheat crop, Fisher and Stapper [
31] found that kernel-number reductions were greater with lodging commencing at or before anthesis, while kernel-weight reductions were greater with later lodging. This pattern supports the idea that the reductions in seed number with early-occurring lodging were mainly due to reductions in photoassimilate supply. Previous research [
33] reported that partitioning to seed in perennial ryegrass and consequent seed filling is not dependent on the mobilization of water-soluble carbohydrates from the internodes to developing seed, and in fact, storage in the spike is sufficient to fill all available seeds. This is in accordance with our findings because higher lodging severity brought about by more intensive N fertilization had no negative effect on seed weights (
Table 4).
For unfertilized plots of the single-purpose crops, the application of PGR largely decreased above-ground biomass but produced slightly higher seed yields (
Figure 2) compared to unsprayed plots of the Seed-crop primarily due to differences in harvest index (
Table 4). The greatest yield increment with increasing N fertilization in the SeedPGR-crop was with the N rate of 60 kg ha
−1 (
Figure 2). Plants treated with PGR consistently had shorter stems and smaller ears on both early- and late-formed reproductive tillers (
Table 3), and ear length was reduced much in the same way as stem length. Ear rachis was shortened causing a reduction in the distance between spikelets along the rachis, but the number and size of the spikelets were not affected by PGR application. Thus, the beneficial modification of the crop canopy thru reduced stem and ear length following PGR application most likely allowed more efficient partitioning of dry matter to seed yield. In perennial ryegrass, neither [
7] nor [
34] found an effect of PGR on dry matter production regardless of application rate, but similarly to our findings, the latter research work reported that seed yield increases following PGR application were due to its positive impact on harvest index.
The SeedPGR-crop produced the highest seed yield of 2373 kg ha
−1 at 120 kg N ha
−1 (
Figure 2), which was 45% larger than the Seed-crop at the same N fertilization, and despite the fact that the latter had larger aboveground biomass at harvest (
Table 4). This highest yielding SeedPGR-crop had relatively high lodging severity at harvest, but the onset of lodging occurred almost a month later (on 7 June) in comparison to the Seed-crop at the same fertilization level. These results suggest that delayed lodging was the main reason for enhanced yield in the SeedPGR-crop. In addition, the number of vegetative tillers at 120 kg N ha
−1 was much smaller in the SeedPGR-crop compared to the unsprayed plants of the Seed-crop (
Table 4). Plants in the SeedPGR-crop developed less vegetative tillers most probably because of lower lodging intensity coupled with the suppressive effect of PGR on forming and growth of new tillers. Consequently, the maximum harvest index of 22.9% was measured in the SeedPGR-crop at 120 kg N ha
−1 and achieved in stand density of 581 fertile tillers per square meter, of which 300 were early-formed and 281 were late-formed ones (greenheads). There is a wide range over which numbers of fertile tillers, spikelets per ear, and fertile florets per spikelet can compensate to attain maximum seed yields, and our results indicate that large tiller populations may not be a necessary prerequisite for large seed yields of Italian ryegrass crop. For annual ryegrass, total tillers ranged from 1790 to 4830 per square meter, while fertile tillers ranged from 110 to 2020 per square meter in the research reported by [
10]. In
Lolium perenne, the number of fertile tillers per square meter may range from 1765 to 3100 in crops for seed production [
7].
Cutting of grasses in the field, with proper timing and height of defoliation, stimulates tillering from buds previously suppressed and in perennial ryegrass, secondary tillers arising from the decapitated tillers produced a greater proportion of mature fertile tillers than secondary tillers arising simultaneously from intact tillers [
35]. However, the dual-purpose crops consistently produced a lower tiller number than the single-purpose crops (
Table 4). Compared to the single-purpose crops, plants from the dual-purpose crops also had smaller and less productive early- and late-formed fertile ears (
Table 3). Almost half a century ago, Herron [
36] reported a decrease in spike length, spikelets pre-spike, and florets per spikelet when Italian ryegrass crop grazing was extended from mid-April to early May in Oregon. The decrease in size and productivity of the ears in the dual-purpose crops (
Table 3) was most likely related to less favourable growing conditions (longer day length and higher temperatures) during their development and growth (
Table 1 and
Table 2), as found in perennial ryegrass by [
37]. Seed weight also significantly decreased in the dual-purpose crops compared to the single-purpose ones (
Table 4), which we associated with the reduced number of days for grain filling (
Table 2). In contrast, the seed weight of annual ryegrass was not affected by grazing treatments in a study [
10]. However, spring cutting for forage removes the first-formed tillers and leaves seed crop to be produced from tillers of lower yield and quality. Consequently, the dual-purpose crops produced smaller seed yields than the single-purpose crops at all N fertilization levels (
Figure 2) even though they had lower seed shattering and lodging intensity at harvest (
Table 4). The smallest seed yield of 269 kg ha
−1 was produced in unfertilized plots of the LF-seed-crop, which was associated with the smallest above-ground production (2004 kg ha
−1), and consequently, the lowest harvest index (9.5%). In contrast to the single-purpose crop responses, the dual-purpose crops consistently increased seed yields with higher N fertilization primarily by means of an improved number of reproductive tillers and heavier seed weight. Ear length consistently increased with increasing N fertilization for early- and late-formed reproductive tillers in the dual-purpose crops (
Table 3). Higher N rates failed to increase spikelet number on early-formed ears but tended to increase it on late-formed ones. The smallest number of vegetative tillers was counted in both the EF-seed-crop and LF-seed-crop (
Table 4), which had limited and late-occurring (
Table 2) lodging at the highest N rate only. The development of new vegetative organs (tillers) occurs at the same time as flowering and seed formation processes in grass crops. It was suggested that the development of tillers from axillary buds in Westerwolds ryegrass is inhibited by auxin translocated from adjacent reproductive tillers and, therefore, the development of late-formed tillers is suppressed during the flowering of non-lodged crop [
38]. In addition to smaller lodging occurrence, the reduction in the number of vegetative tillers in the dual-purpose crops in our research could be also associated with a shorter post-anthesis period (
Table 2) and less favourable (hotter) weather conditions (
Table 1). At the highest N fertilization, the dual-purpose crops succeeded to yield similarly to the Seed-crop grown at the same N level (
Figure 2). In a companion paper [
15], it was shown that under the conditions of late-season and limited lodging, seed-yields of the first-year Italian ryegrass crop consistently increased with higher N fertilization rates regardless of management systems. Therefore, maximum seed yields in the dual-purpose crops most likely have not been achieved so total N fertilization rates higher than 180 kg ha
−1 could be recommended for combined forage-seed crop management.
Across management systems, the single-purpose crops had the heaviest seed weight and seed germination (
Table 4). The dual-purpose crops had significantly lighter seeds than the single-purpose crops, which was primarily associated with the reduced number of days for seed filling (
Table 2). Anthesis to harvest period lasted about 3 weeks in the dual-purpose crops, which was one week shorter compared to the single-purpose crops. In Wales, Hides et al. [
21] reported that the seed weight of Italian ryegrass populations increased up to 32 days after anthesis. In tetraploid Italian ryegrass [
39], the research results showed that increasing seed weight, both between and within commercial seed lots is important because of increased seedling growth, emergence at 5 and 10 °C, and vigour. The highest germination rates were found in the single-purpose crops grown at the N fertilization of 120 kg ha
−1, which averaged 94.0% in the Seed-crop and 95.5% in the SeedPGR-crop. Seeds originating from the LF-seed-crop had the lowest germination (
Table 4). Seed germination was unaffected by N fertilization in the single-purpose crops as well as in the EF-seed-crop. However, unfertilized plots in the LF-seed-crop produced a 1000-seed weight of 3.45 g and a germination rate of 79.8% only. Unfertilized plots in the EF-seed-crop had seeds of similar weight but with significantly higher germination (90.8%). In Oregon [
10], it was reported that seed germination of annual ryegrass ranged from 86 to 97%. Previous research [
40] showed that it seemed likely that some viable but immature and low-weight seeds of perennial ryegrass contained insufficient food reserves to maintain their viability after three months’ storage. However, increasing N fertilization resulted in heavier seeds as well as in higher germination rate (
Table 4), and the LF-seed-crop grown at the highest N fertilization produced seeds that germinated similarly to those from other management systems.


 ,
, 


{kind=link}
{kind=link}