
Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing

 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site and Irrigation Treatments
2.2. Growing Degree Days, Flowering and Pit Hardening
2.3. Fruit Set, Plant Morphology and Physiological Observations
2.4. Yield, Irrigation Water Productivity and Canopy Leaf Area
2.5. Statistical Analysis
3. Results and Discussion
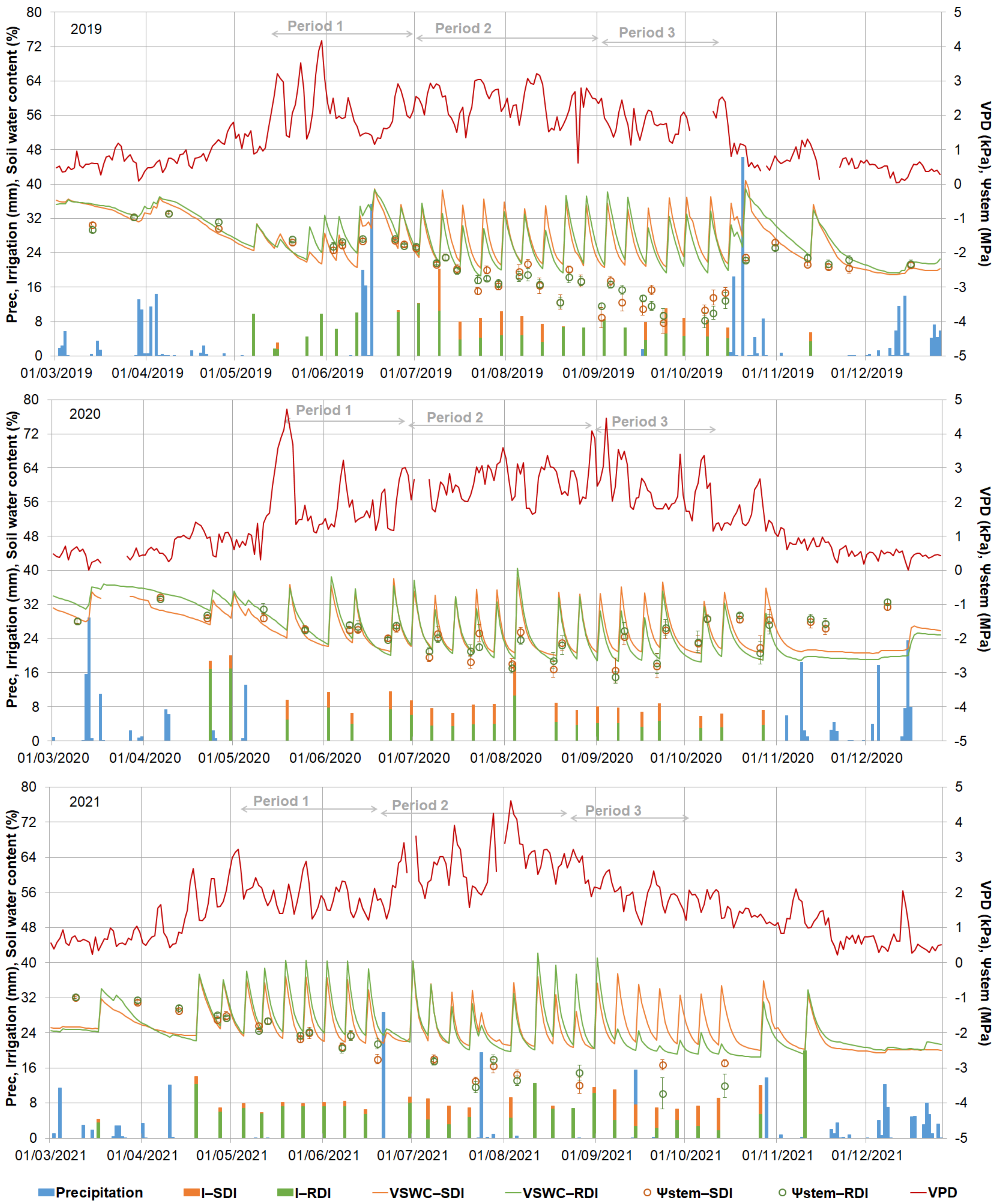
3.1. Environmental Conditions and Irrigation Monitoring
3.2. Growing Degree Days, Flowering and Fruit Set
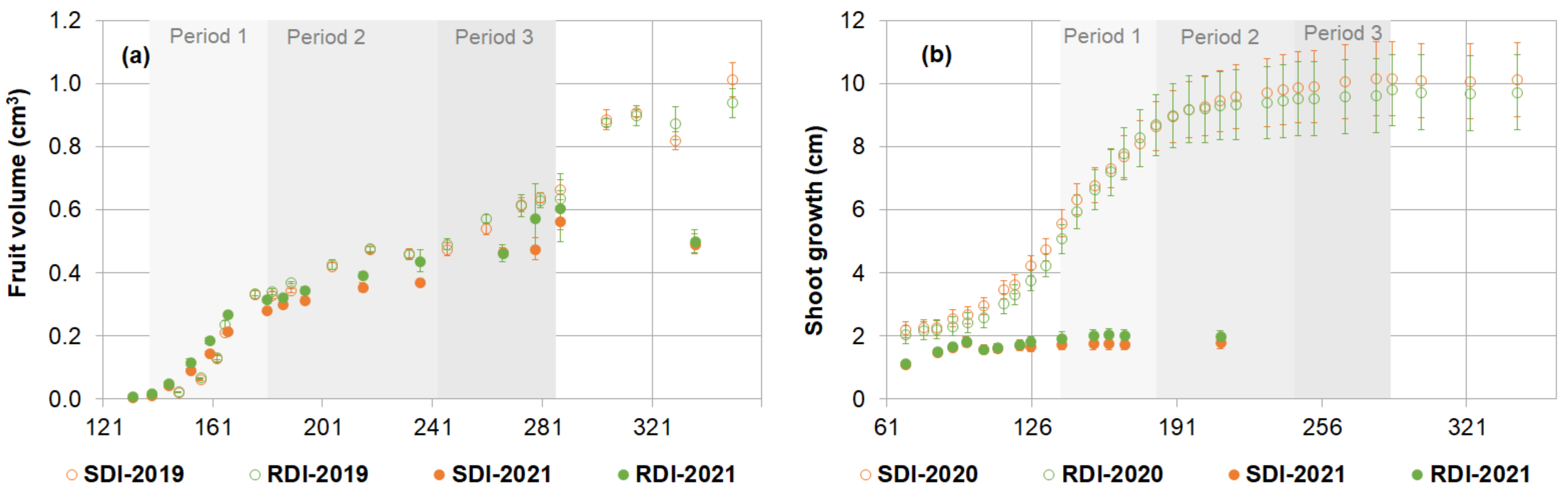
3.3. Plant Morphology
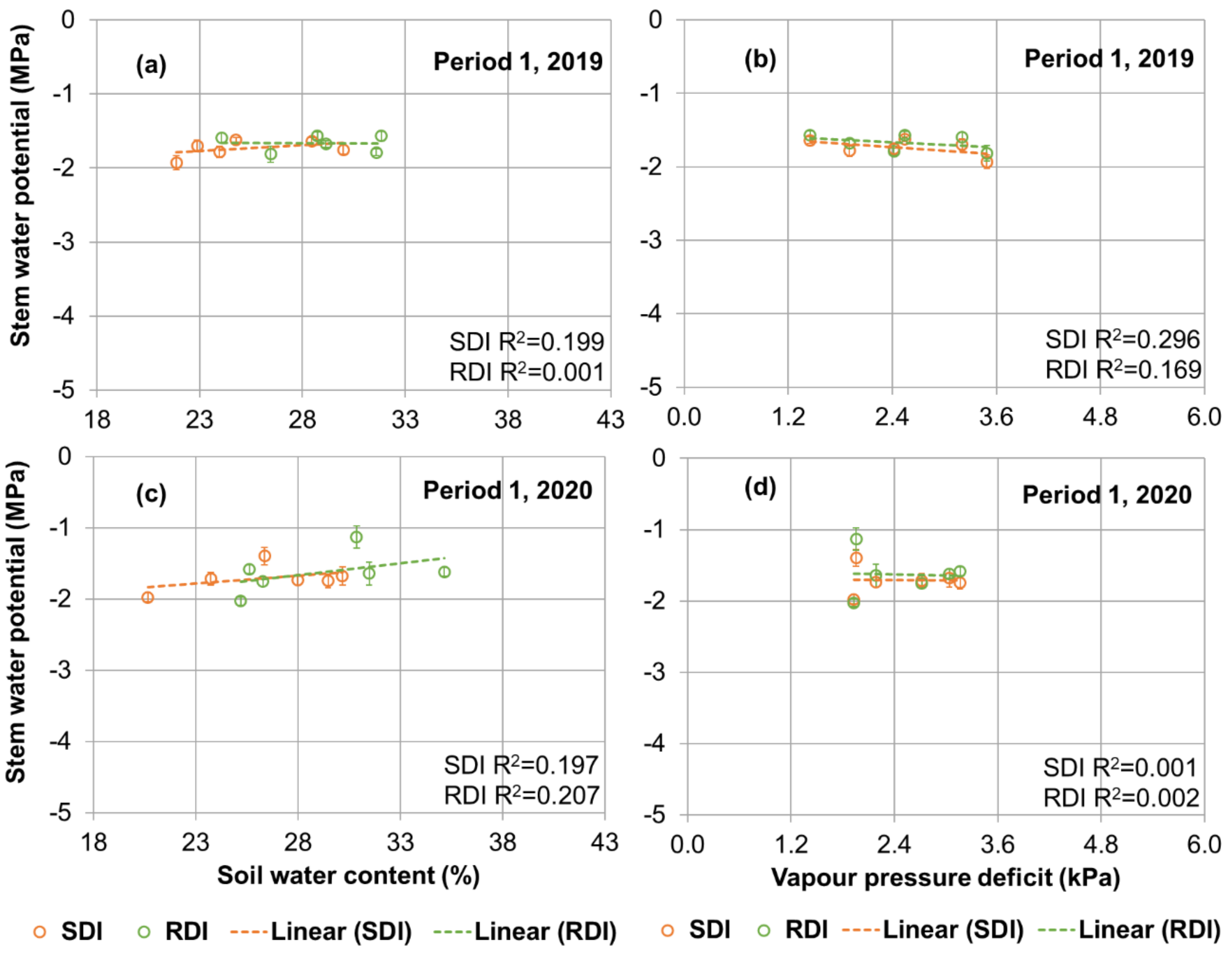
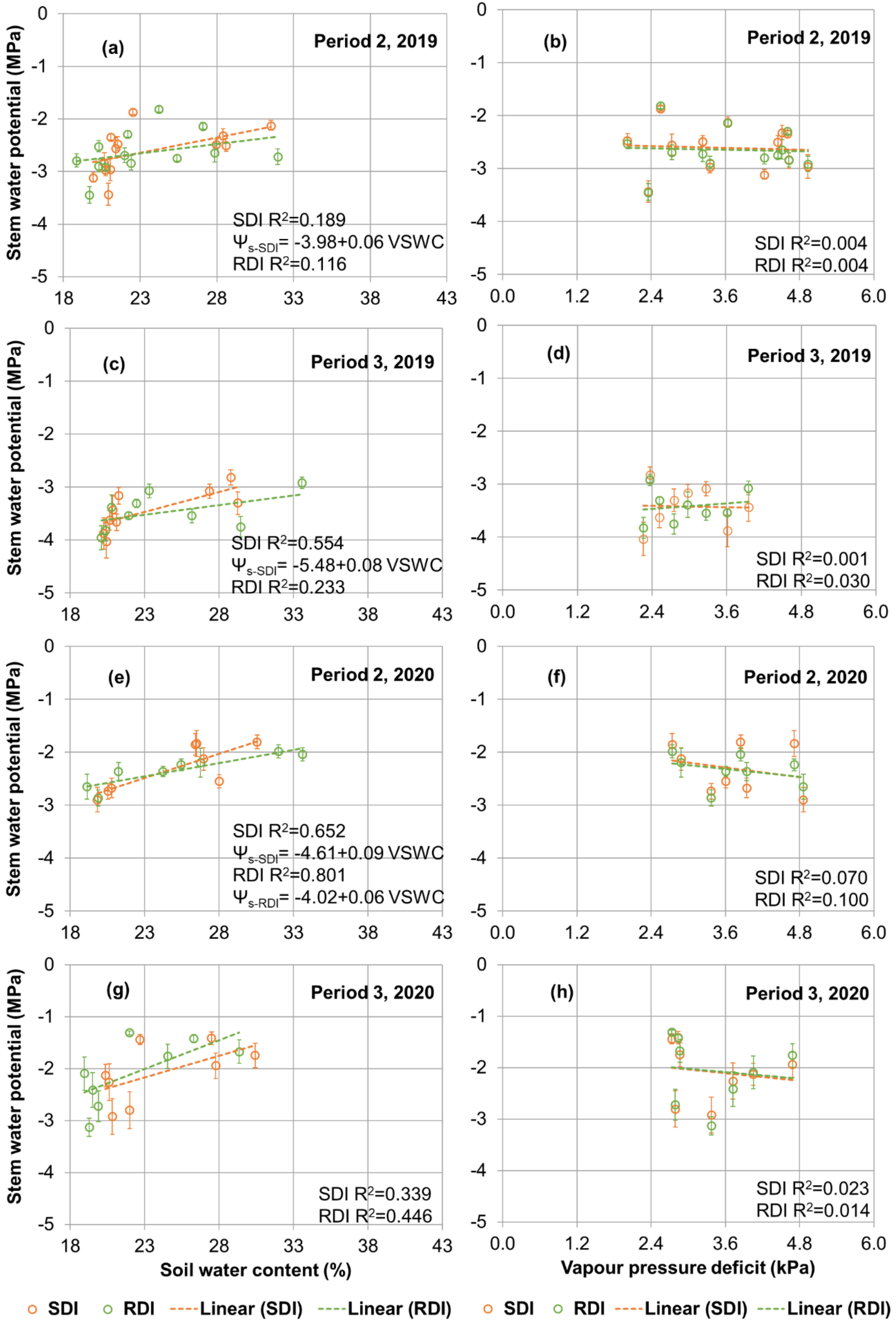
3.4. Plant Physiology
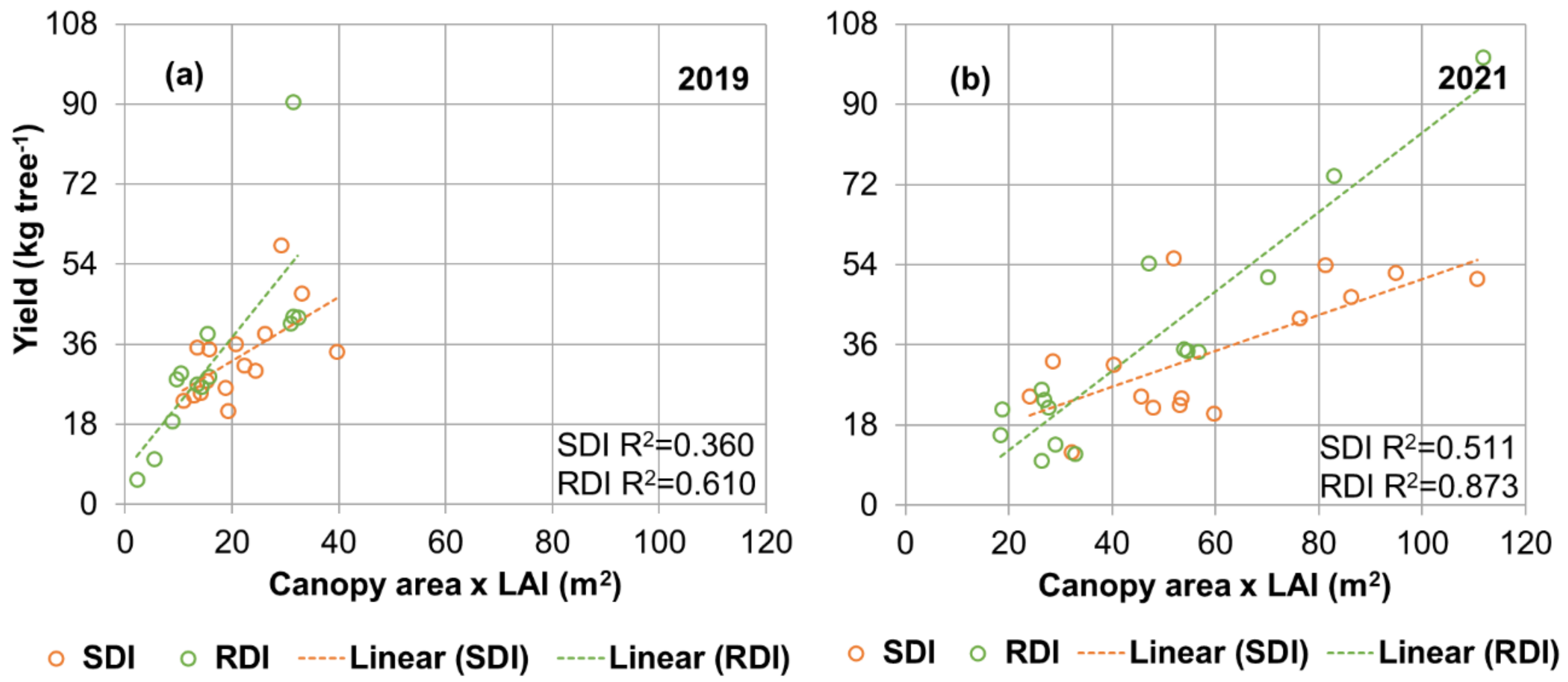
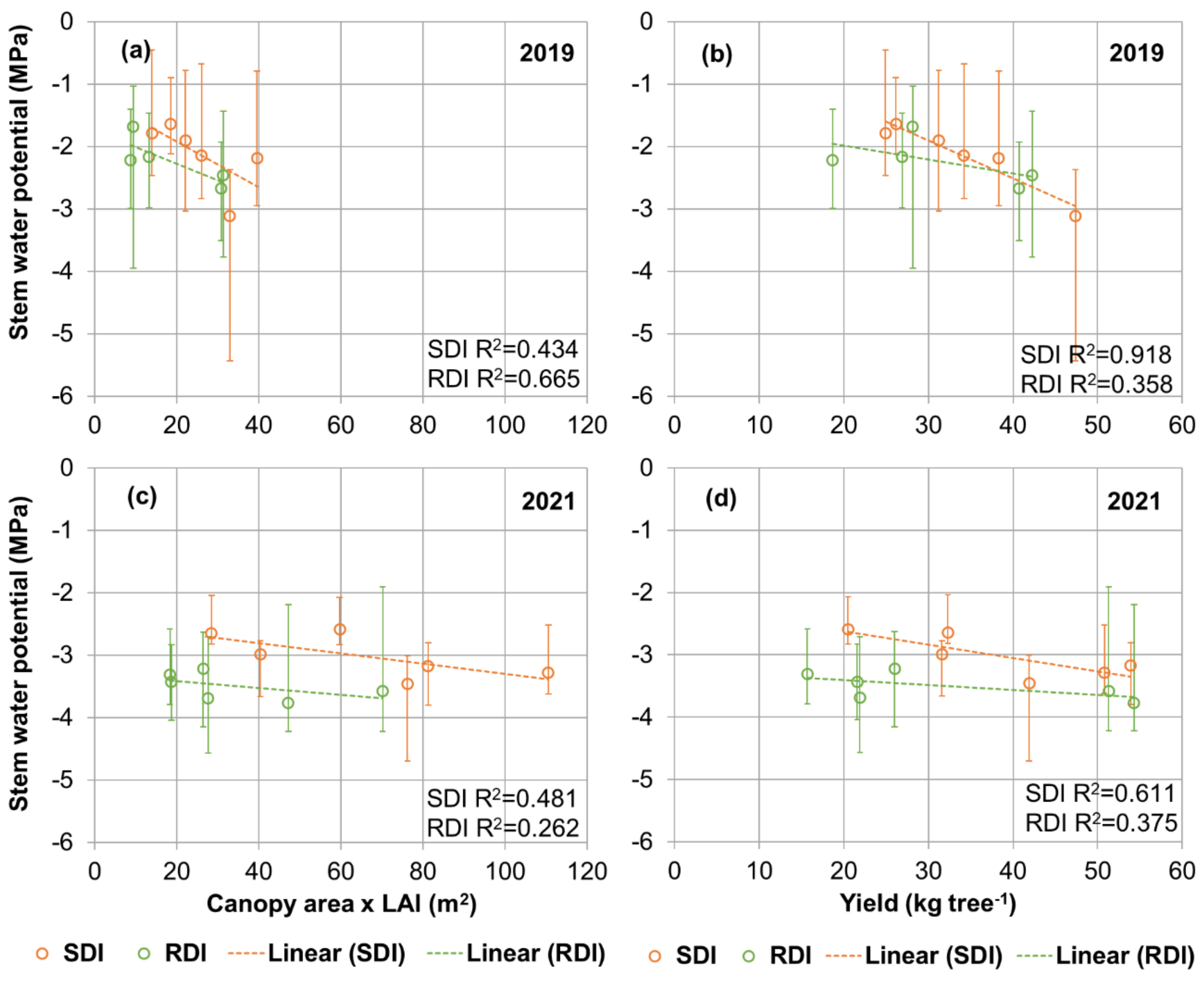
3.5. Relations between Ψs, Canopy Leaf Area and Yield
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Monthly Meteorological Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Month | P mm | Tmin °C | Tmax °C | RHmin % | RHmax % | VPD kPa | WS m s−1 | ETo mm |
|---|---|---|---|---|---|---|---|---|---|
| 2019 | Jan | 128.6 | 3.5 | 15.2 | 57.5 | 98.6 | 0.4 | 0.3 | 28.4 |
| 2019 | Feb | 132.2 | 4.6 | 15.9 | 58.7 | 98.6 | 0.4 | 0.3 | 38.6 |
| 2019 | Mar | 40.3 | 5.4 | 19.1 | 48.9 | 96.9 | 0.6 | 0.3 | 68.9 |
| 2019 | Apr | 33.2 | 8.0 | 22.4 | 42.5 | 95.4 | 0.9 | 0.3 | 97.5 |
| 2019 | May | 0.1 | 13.0 | 31.7 | 23.7 | 84.0 | 2.1 | 0.5 | 150.9 |
| 2019 | Jun | 72.0 | 17.5 | 33.6 | 30.6 | 93.5 | 1.9 | 0.4 | 154.8 |
| 2019 | Jul | 0.0 | 19.0 | 35.9 | 24.9 | 86.1 | 2.4 | 0.5 | 167.8 |
| 2019 | Aug | 0.0 | 19.7 | 36.7 | 27.4 | 93.5 | 2.3 | 0.5 | 149.3 |
| 2019 | Sep | 1.6 | 17.5 | 33.7 | 32.9 | 93.4 | 1.8 | 0.4 | 110.5 |
| 2019 | Oct | 67.0 | 15.9 | 29.7 | 39.0 | 89.0 | 1.2 | 0.3 | 50.0 |
| 2019 | Nov | 0.3 | 10.3 | 24.2 | 53.7 | 96.7 | 0.8 | 0.3 | 32.9 |
| 2019 | Dec | 89.5 | 5.9 | 18.0 | 64.8 | 99.6 | 0.4 | 0.2 | 25.7 |
| 2020 | Jan | 101.3 | 3.7 | 14.5 | 70.6 | 99.5 | 0.3 | 0.3 | 27.4 |
| 2020 | Feb | 22.8 | 3.9 | 15.8 | 65.6 | 99.4 | 0.3 | 0.4 | 40.8 |
| 2020 | Mar | 62.1 | 7.7 | 19.8 | 63.4 | 100.0 | 0.4 | 0.4 | 48.6 |
| 2020 | Apr | 17.6 | 8.2 | 24.1 | 48.8 | 99.0 | 0.8 | 0.4 | 104.0 |
| 2020 | May | 13.9 | 13.3 | 30.2 | 29.1 | 83.1 | 1.8 | 0.5 | 145.1 |
| 2020 | Jun | 0.0 | 15.4 | 32.2 | 29.9 | 89.8 | 1.9 | 0.5 | 156.3 |
| 2020 | Jul | 0.0 | 19.6 | 37.9 | 24.8 | 95.5 | 2.6 | 0.5 | 176.0 |
| 2020 | Aug | 0.0 | 19.6 | 37.9 | 24.0 | 91.7 | 2.6 | 0.5 | 156.4 |
| 2020 | Sep | 0.0 | 19.2 | 36.9 | 25.9 | 91.2 | 2.5 | 0.4 | 118.3 |
| 2020 | Oct | 0.1 | 15.6 | 31.8 | 30.1 | 88.7 | 1.8 | 0.3 | 75.7 |
| 2020 | Nov | 37.9 | 11.0 | 22.3 | 50.9 | 97.5 | 0.7 | 0.2 | 36.8 |
| 2020 | Dec | 62.1 | 6.9 | 19.1 | 59.7 | 100.0 | 0.5 | 0.2 | 26.1 |
| 2021 | Jan | 25.7 | 4.9 | 17.6 | 58.2 | 99.8 | 0.4 | 0.2 | 30.1 |
| 2021 | Feb | 10.7 | 4.4 | 18.6 | 52.0 | 97.6 | 0.5 | 0.3 | 42.8 |
| 2021 | Mar | 26.0 | 4.8 | 19.5 | 45.3 | 96.2 | 0.6 | 0.4 | 72.4 |
| 2021 | Apr | 15.7 | 8.3 | 25.9 | 30.5 | 91.5 | 1.4 | 0.4 | 110.2 |
| 2021 | May | 0.2 | 13.7 | 33.1 | 24.1 | 88.1 | 2.1 | 0.4 | 155.9 |
| 2021 | Jun | 28.7 | 16.0 | 32.6 | 29.3 | 86.3 | 1.9 | 0.5 | 147.8 |
| 2021 | Jul | 0.0 | 19.8 | 34.8 | 24.9 | 85.8 | 2.5 | 0.4 | 161.6 |
| 2021 | Aug | 1.2 | 20.8 | 39.1 | 20.3 | 86.2 | 3.0 | 0.5 1 | 161.5 |
| 2021 | Sep | 15.9 | 16.8 | 33.4 | 29.1 | 91.8 | 1.9 | 0.5 1 | 114.1 |
| 2021 | Oct | 13.6 | 13.0 | 29.2 | 30.7 | 89.1 | 1.5 | 0.5 1 | 78.6 |
| 2021 | Nov | 7.5 | 10.1 | 24.9 | 41.4 | 91.3 | 1.0 | 0.5 1 | 46.3 |
| 2021 | Dec | 204.7 | 6.0 | 17.0 | 54.8 | 97.0 | 0.6 | 0.5 1 | 29.5 |
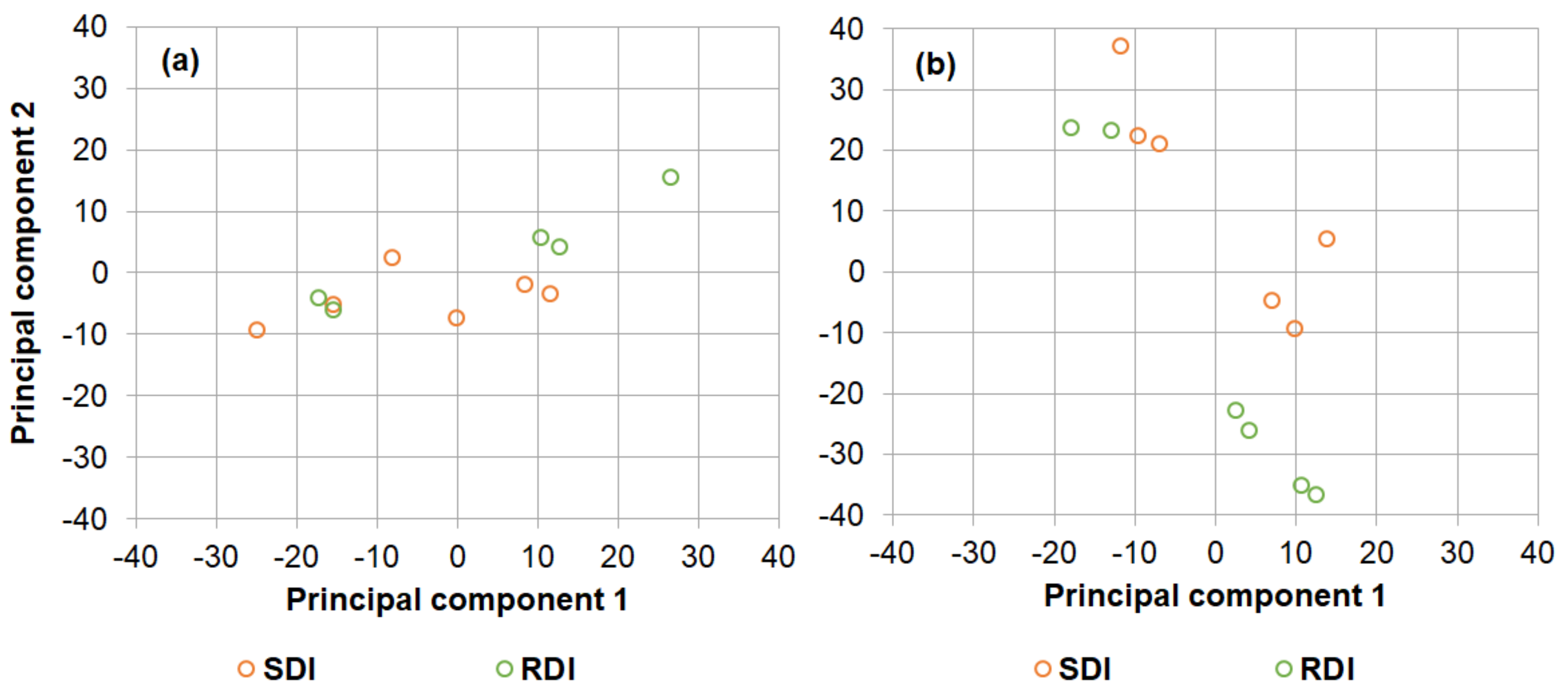
Appendix B. Principal Component Analysis
| Component Matrix 2019 | |||
|---|---|---|---|
| Component | |||
| 1 | 2 | 3 | |
| Variance explained (%) | 42 | 24 | 15 |
| Yield | −0.961 | −0.103 | −0.053 |
| Canopy area × LAI | −0.786 | −0.306 | 0.390 |
| Ψs_Period 3 | 0.705 | −0.493 | 0.447 |
| Ψs_Period 2 | 0.696 | −0.570 | 0.307 |
| Ψs_Period 1 | 0.678 | 0.211 | −0.412 |
| VSWC_Period 1 | −0.021 | 0.873 | 0.278 |
| VSWC_Period 3 | 0.526 | 0.637 | 0.395 |
| VSWC_Period 2 | −0.327 | 0.168 | 0.600 |
| Component Matrix 2021 | |||
|---|---|---|---|
| Component | |||
| 1 | 2 | 3 | |
| Variance explained (%) | 46 | 23 | 16 |
| Ψs_Period 3 | 0.916 | 0.161 | −0.048 |
| Ψs_Period 2 | 0.912 | 0.059 | −0.205 |
| Ψs_Period 1 | 0.827 | −0.089 | −0.321 |
| VSWC_Period 3 | 0.783 | 0.274 | 0.356 |
| Canopy area × LAI | −0.166 | 0.899 | 0.113 |
| Yield | −0.501 | 0.778 | 0.128 |
| VSWC_Period 1 | −0.192 | −0.565 | 0.717 |
| VSWC_Period 2 | 0.637 | 0.140 | 0.689 |

References
- Zittis, G.; Bruggeman, A.; Lelieveld, J. Revisiting future extreme precipitation trends in the Mediterranean. Weather Clim. Extrem. 2021, 34, 100380. [Google Scholar] [CrossRef] [PubMed]
- Moriana, A.; Orgaz, F.; Pastor, M.; Fereres, E. Yield responses of a mature olive orchard to water deficits. J. Am. Soc. Hortic. Sci. 2003, 128, 425–431. [Google Scholar] [CrossRef]
- Ennajeh, M.; Tounekti, T.; Vadel, A.M.; Khemira, H.; Cochard, H. Water relations and drought-induced embolism in olive (Olea europaea) varieties ‘Meski’ and ‘Chemlali’ during severe drought. Tree Physiol. 2008, 28, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Rossi, L.; Sebastiani, L.; Tognetti, R.; d’Andria, R.; Morelli, G.; Cherubini, P. Tree-ring wood anatomy and stable isotopes show structural and functional adjustments in olive trees under different water availability. Plant Soil 2013, 372, 567–579. [Google Scholar] [CrossRef]
- Diaz-Espejo, A.; Fernández, J.E.; Torres-Ruiz, J.M.; Rodriguez-Dominguez, C.M.; Perez-Martin, A.; Hernandez-Santana, V. The olive tree under water stress: Fitting the pieces of response mechanisms in the crop performance puzzle. In Water Scarcity and Sustainable Agriculture in Semiarid Environment, 1st ed.; García Tejero, I.F., Durán Zuazo, V.H., Eds.; Elsevier: London, UK, 2018; pp. 439–479. [Google Scholar] [CrossRef]
- Tognetti, R.; d’Andria, R.; Lavini, A.; Morelli, G. The effect of deficit irrigation on crop yield and vegetative development of Olea europaea L. (cvs. Frantoio and Leccino). Eur. J. Agron. 2006, 25, 356–364. [Google Scholar] [CrossRef]
- Fernández, J.E. Understanding olive adaptation to abiotic stresses as a tool to increase crop performance. Environ. Exp. Bot. 2014, 103, 158–179. [Google Scholar] [CrossRef]
- Moriana, A.; Pérez-López, D.; Prieto, M.H.; Ramírez-Santa-Pau, M.; Pérez-Rodrigues, J.M. Midday stem water potential as a useful tool for estimating irrigation requirements in olive trees. Agric. Water Manag. 2012, 112, 43–54. [Google Scholar] [CrossRef]
- Bianco, R.L.; Scalisi, A. Water relations and carbohydrate partitioning of four greenhouse-grown olive genotypes under long-term drought. Trees 2017, 31, 717–727. [Google Scholar] [CrossRef]
- Naor, A.; Schneider, D.; Ben-Gal, A.; Zipori, I.; Dag, A.; Kerem, Z.; Birger, R.; Peres, M.; Gal, Y. The effects of crop load and irrigation rate in the oil accumulation stage on oil yield and water relations of ‘Koroneiki’ olives. Irrig. Sci. 2013, 31, 781–791. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Goncalves, B.C.; Ferreira, H.F.; Correia, C.M. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2006, 60, 183–192. [Google Scholar] [CrossRef]
- Grattan, S.R.; Berenguer, M.J.; Connell, J.H.; Polito, V.S.; Vossen, P.M. Olive oil production as influenced by different quantities of applied water. Agric. Water Manag. 2006, 85, 133–140. [Google Scholar] [CrossRef]
- Rapoport, H.F.; Costagli, G.; Gucci, R. The effect of water deficit during early fruit development on olive fruit morphogenesis. J. Amer. Soc. Hort. Sci. 2004, 129, 121–127. [Google Scholar] [CrossRef]
- Hueso, A.; Camacho, G.; Gómez-del-Campo, M. Spring deficit irrigation promotes significant reduction on vegetative growth, flowering, fruit growth and production in hedgerow olive orchards (cv. Arbequina). Agric. Water Manag. 2021, 248, 106695. [Google Scholar] [CrossRef]
- Gucci, R.; Caruso, G.; Gennai, C.; Esposto, S.; Urbani, S.; Servili, M. Fruit growth, yield and oil quality changes induced by deficit irrigation at different stages of olive fruit development. Agric. Water Manag. 2019, 212, 88–98. [Google Scholar] [CrossRef]
- Monson, R.; Baldocchi, D. Boundary layer and stomatal control over leaf fluxes. In Terrestrial Biosphere-Atmosphere Fluxes, 1st ed.; Cambridge University Press: Cambridge, UK, 2014; pp. 136–172. [Google Scholar] [CrossRef]
- Bonan, G. Plant hydraulics. In Climate Change and Terrestrial Ecosystem Modelling, 1st ed.; Cambridge University Press: Cambridge, UK, 2019; pp. 213–227. [Google Scholar] [CrossRef]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. Lond. 1976, 273, 593–610. [Google Scholar] [CrossRef]
- Ortuño, M.F.; García-Orellana, Y.; Conejero, W.; Ruiz-Sánchez, M.C.; Mounzer, O.; Alarcón, J.J.; Torrecillas, A. Relationships between climatic variables and sap flow, stem water potential and maximum daily trunk shrinkage in lemon trees. Plant Soil 2006, 279, 229–242. [Google Scholar] [CrossRef]
- Corell, M.; Pérez-López, D.; Martín-Palomo, M.J.; Centeno, A.; Girón, I.; Galindo, A.; Moreno, M.M.; Moreno, C.; Memmi, H.; Torrecillas, A.; et al. Comparison of the water potential baseline in different locations. Usefulness for irrigation scheduling of olive orchards. Agric. Water Manag. 2016, 177, 308–316. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Sadras, V.O.; Puertas, C.M. Effects of the source:sink ratio of the phenotypic plasticity of stem water potential in olive (Olea europaea L.). J. Exp. Bot. 2011, 62, 3535–3543. [Google Scholar] [CrossRef]
- Martín-Vertedor, A.I.; Rodríguez, J.M.P.; Losada, H.P.; Castiel, E.F. Interactive responses to water deficits and crop load in olive (olea europaea L., cv. Morisca) I.—Growth and water relations. Agric. Water Manag. 2011, 98, 941–949. [Google Scholar] [CrossRef][Green Version]
- Dahl, A.; Galán, C.; Hajkova, L.; Pauling, A.; Sikoparija, B.; Smith, M.; Vokou, D. The onset, course and intensity of the pollen season. In Allergenic Pollen: A Review of the Production, Release, Distribution and Health Impacts, 1st ed.; Sofiev, M., Bergmann, K.C., Eds.; Springer: Dordrecht, The Nehterlands; Heidelberg/Berlin, Germany; New York, NY, USA; London, UK, 2013; pp. 29–70. [Google Scholar] [CrossRef]
- Aguilera, F.; Ruiz, L.; Fornaciari, M.; Romano, B.; Galán, C.; Oteros, J.; Dhiab, A.B.; Msallem, M.; Orlandi, F. Heat accumulation period in the Mediterranean region: Phenological response of the olive in different climate areas (Spain, Italy and Tunisia). Int. J. Biometeorol. 2014, 58, 867–876. [Google Scholar] [CrossRef]
- Orlandi, F.; Sgromo, C.; Bonofiglio, T.; Ruga, L.; Romano, B.; Fornaciari, M. Spring influences on olive flowering and threshold temperatures related to reproductive structure formation. HortScience 2010, 45, 1052–1057. [Google Scholar] [CrossRef]
- Garrido, A.; Fernández-González, M.; Álvarez-López, S.; González-Fernández, E.; Rodríguez-Rajo, F.J. First phenological and aerobiological assessment of olive orchards at the Northern limit of the Mediterranean bioclimatic area. Aerobiologia 2020, 36, 641–656. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration. Guideline for computing crop water requirements. In FAO Irrigation and Drainage Paper No. 56; FAO: Rome, Italy, 1998; pp. 1–326. [Google Scholar]
- Connor, D.J.; Fereres, E. The physiology of adaptation and yield expression in olive. In Horticultural Reviews, 1st ed.; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2005; Volume 31, pp. 155–229. [Google Scholar] [CrossRef]
- Bustan, A.; Dag, A.; Yermiyahu, U.; Erel, R.; Presnov, E.; Agam, N.; Kool, D.; Iwema, J.; Zipori, I.; Ben-Gal, A. Fruit load governs transpiration of olive trees. Tree Physiol. 2016, 36, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, F.J.; Orgaz, F.; Testi, L.; Fereres, E. Measurement and modelling of evapotranspiration of olive (Olea europaea L.) orchards. Eur. J. Agron. 2000, 13, 155–163. [Google Scholar] [CrossRef]
- Testi, L.; Villalobos, F.J.; Orgaz, F.; Fereres, E. Water requirements of olive orchards: I simulation of daily evapotranspiration for scenario analysis. Irrig. Sci. 2006, 24, 69–76. [Google Scholar] [CrossRef]
- Siakou, M.; Bruggeman, A.; Eliades, M.; Zoumides, C.; Djuma, H.; Kyriacou, M.C.; Emmanouilidou, M.; Spyros, A.; Manolopoulou, E.; Moriana, A. Effects of deficit irrigation on ‘Koroneiki’ olive tree growth, physiology and olive oil quality at different harvest dates. Agric. Water Manag. 2021, 258, 107200. [Google Scholar] [CrossRef]
- Fernández, J. Plant-based methods for irrigation scheduling of woody crops. Horticulturae 2017, 3, 35. [Google Scholar] [CrossRef]
- Sanz-Cortés, F.; Martinez-Calvo, J.; Badenes, M.L.; Bleiholder, H.; Hack, H.; Llacer, G.; Meier, U. Phenological growth stages of olive trees (Olea europaea). Ann. Appl. Biol. 2002, 140, 151–157. [Google Scholar] [CrossRef]
- Mcmaster, G.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Pérez-López, D.; Ribas, F.; Moriana, A.; Rapoport, H.F.; De Juan, A. Influence of temperature on the growth and development of olive (Olea europaea L.) trees. J. Hortic. Sci. Biotech. 2008, 83, 171–176. [Google Scholar] [CrossRef]
- Oteros, J.; García-Mozo, H.; Vázquez, L.; Mestre, A.; Domínguez-Vilches, E.; Galán, C. Modelling olive phenological response to weather and topography. Agric. Ecosyst. Environ. 2013, 179, 62–68. [Google Scholar] [CrossRef]
- Rapoport, H.F.; Pérez-López, D.; Hammami, S.B.M.; Agüera, J.; Moriana, A. Fruit pit hardening: Physical measurement during olive fruit growth. Ann. Appl. Biol. 2013, 163, 200–208. [Google Scholar] [CrossRef]
- Fernández, J.E.; Alcon, F.; Diaz-Espejo, A.; Hernandez-Santana, V.; Cuevas, M.V. Water use indicators and economic analysis for on-farm irrigation decision: A case study of a super high density olive tree orchard. Agric. Water Manag. 2020, 237, 106074. [Google Scholar] [CrossRef]
- LI-COR. LAI-2200C Plant Canopy Analyser: Instruction Manual. Available online: https://www.licor.com/env/support/LAI-2200C/manuals.html (accessed on 4 November 2019).
- Camera, C.; Bruggeman, A.; Hadjinicolaou, P.; Pashiardis, S.; Lange, M.A. Evaluation of interpolation techniques for the creation of gridded daily precipitation (1 × 1 km2); Cyprus, 1980–2010. J. Geophys. Res. 2013, 119, 693–712. [Google Scholar] [CrossRef]
- Cammalleri, C.; Rallo, G.; Agnese, C.; Ciraolo, G.; Minacapilli, M.; Provenzano, G. Combined use of eddy covariance and sap flow techniques for partition of ET fluxes and water stress assessment in an irrigated olive orchard. Agric. Water Manag. 2013, 120, 89–97. [Google Scholar] [CrossRef]
- Girón, I.F.; Corell, M.; Galindo, A.; Torrecillas, E.; Morales, D.; Dell’Amico, J.; Torrecillas, A.; Moreno, F.; Moriana, A. Changes in the physiological response between leaves and fruits during a moderate water stress in table olive trees. Agric. Water Manag. 2015, 148, 280–286. [Google Scholar] [CrossRef]
- Beyá-Marshall, V.; Herrera, J.; Fichet, T.; Trentacoste, E.R.; Kremer, C. The effect of water status on productive and flowering variables in young ‘Arbequina’ olive trees under limited irrigation water availability in a semiarid region of Chile. Hortic. Environ. Biotech. 2018, 59, 815–826. [Google Scholar] [CrossRef]
- Dag, A.; Bustan, A.; Avni, A.; Tzipori, I.; Lavee, S.; Riov, J. Timing of fruit removal affects concurrent vegetative growth and subsequent return bloom and yield in olive (Olea europaea L.). Sci. Hortic. 2010, 123, 469–472. [Google Scholar] [CrossRef]
- Fernández, F.J.; Ladux, J.L.; Searles, P.S. Dynamics of shoot and fruit growth following fruit thinning in olive trees: Same season and subsequent season responses. Sci. Hortic. 2015, 192, 320–330. [Google Scholar] [CrossRef]
- Ben-Gal, A.; Yermiyahu, U.; Zipori, I.; Presnov, E.; Hanoch, E.; Dag, A. The influence of bearing cycles on olive oil production response to irrigation. Irrig. Sci. 2011, 29, 253–263. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Correia, C.M.; Moutinho-Pereira, J.M.; Goncalves, B.C.; Lopes, J.I.; Torres-Pereira, J.M.G. Sclerophylly and leaf anatomical traits of five field-grown olive cultivars growing under drought conditions. Tree Physiol. 2004, 24, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Pierantozzi, P.; Torres, M.; Tivani, M.; Contreras, C.; Gentili, L.; Parera, C.; Maestri, D. Spring deficit irrigation in olive (cv. Genovesa) growing under arid continental climate: Effects on vegetative growth and productive parameters. Agric. Water Manag. 2020, 238, 106212. [Google Scholar] [CrossRef]






| Year | Period | Dates | Phenological Growth Stage | ETc-SDI (%) | ETc-RDI (%) | |
|---|---|---|---|---|---|---|
| 2019 | 1 | 13 May–30 Jun | 69–71 | End of flowering–early fruit growth | 70 | 70 |
| 2 | 01 Jul–01 Sep | 71–79 | Maximum rate of pit hardening | 70 | 35.70 1 | |
| 3 | 02 Sep–12 Oct | 80–81 | Oil accumulation–start of rainfall | 70 | 35.70 2 | |
| 2020 | 1 | 18 May–28 Jun | 31–37 | Fast shoot elongation | 70 | 35 |
| 2 | 29 Jun–30 Aug | 38 | Small rate of shoot elongation | 70 | 35 | |
| 3 | 31 Aug–11 Oct | 39 | End of shoot elongation-start of rainfall | 70 | 35 | |
| 2021 | 1 | 04 May–19 Jun | 69–71 | End of flowering–early fruit growth | 70 | 70 |
| 2 | 20 Jun–22 Aug | 71–79 | Maximum rate of pit hardening | 70 | 35.70 1 | |
| 3 | 23 Aug–02 Oct | 80–81 | Oil accumulation–start of rainfall | 70 | 35.70 2 | |
| Year | Period | P | ETo | ETc | ISDI | IRDI | DRSDI | DRRDI | ΔVSWCSDI | ΔVSWCRDI | Kc | Ψs-SDI | Ψs-RDI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mm | mm | mm | mm | mm | mm | mm | mm | mm | MPa | MPa | |||
| 2019 “On” | 1 | 72 | 256 | 102 | 46 | 46 | −5 | −6 | −13 | −13 | 0.32 | −1.9 | −1.8 |
| 2 | 0 | 325 | 130 | 102 | 52 | −12 | −1 | −20 | −18 | 0.39 | −3.4 | −3.4 | |
| 3 | 2 | 143 | 57 | 57 | 37 | −10 | −5 | 16 | 2 | 0.41 | −4.0 | −4.0 | |
| 2020 “Off” | 1 | 0 | 213 | 85 | 42 | 24 | −7 | −3 | −11 | −23 | 0.29 | −2.0 | −2.0 |
| 2 | 0 | 339 | 136 | 82 | 41 | −10 | −5 | −12 | −29 | 0.41 | −2.9 | −2.9 | |
| 3 | 0 | 155 | 62 | 41 | 20 | −5 | 0 | 6 | 5 | 0.38 | −2.9 | −3.1 | |
| 2021 “On” | 1 | 0 | 236 | 94 | 53 | 47 | −4 | −7 | −13 | −1 | 0.30 | −2.8 | −2.4 |
| 2 | 51 | 345 | 138 | 61 | 44 | −6 | −9 | −8 | −14 | 0.39 | −3.5 | −3.5 | |
| 3 | 16 | 168 | 67 | 49 | 30 | −3 | −4 | 7 | −21 | 0.52 | −2.9 | −3.7 |
| Growth Stages | Threshold Temperatures (°C) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 6 | 7 | 8 | 9 | 11 | 13 | 14 | 15 | ||
| Full flowering (°C days) | 11 May 2019 | 947 | 817 | 690 | 565 | 445 | 252 | 139 | 103 | 74 |
| 27 Apr 2021 | 902 | 787 | 674 | 564 | 462 | 280 | 144 | 106 | 81 | |
| Maximum rate of pit hardening (°C days) | 25 Jun 2019 | 1844 | 1669 | 1496 | 1327 | 1162 | 879 | 676 | 595 | 521 |
| 20 Jun 2021 | 1876 | 1708 | 1541 | 1377 | 1221 | 931 | 687 | 595 | 516 | |
| Treatment | Flowers/Branch | Fruits/Branch | Fruit Set (%) | ||
|---|---|---|---|---|---|
| 30 Apr 2019 | 27 May 2019 | 27 May 2019 | 11 Jun 2019 | 09 Jul 2019 | |
| SDI | 234 ± 10 | 18 ± 2 | 7.8 | 2.5 | 2.2 |
| RDI | 210 ± 12 | 19 ± 2 | 9.0 | 3.0 | 2.6 |
| 15 Apr 2021 | 18 May 2021 | 18 May 2021 | 27 May 2021 | 29 Jun 2021 | |
| SDI | 214 ± 5 | 21 ± 3 | 10.2 | 5.7 | 3.9 |
| RDI | 238 ± 29 | 18 ± 2 | 7.9 | 5.0 | 3.9 |
| Year | Period | N | Model (p-Value) Ψs × VSWC × VPD | Significance (p-Value) | ||||
|---|---|---|---|---|---|---|---|---|
| Ψs × VSWC | Ψs × VPD | |||||||
| SDI | RDI | SDI | RDI | SDI | RDI | |||
| 2019 | 1 | 6 | 0.646 | 0.274 | 0.984 | 0.164 | 0.502 | 0.140 |
| 2 | 13 | 0.131 | 0.412 | 0.050 | 0.199 | 0.681 | 0.739 | |
| 3 | 8 | 0.069 | 0.172 | 0.029 | 0.228 | 0.527 | 0.188 | |
| 2020 | 1 | 6 | 0.323 | 0.300 | 0.428 | 0.324 | 0.686 | 0.294 |
| 2 | 8 | 0.074 | 0.011 | 0.035 | 0.005 | 0.984 | 0.555 | |
| 3 | 7 | 0.326 | 0.240 | 0.167 | 0.115 | 0.762 | 0.999 | |
| 2021 | 1 | 7 | 0.478 | 0.356 | 0.263 | 0.239 | 0.639 | 0.761 |
| 2 | 4 | 0.993 | 0.977 | 0.969 | 0.881 | 0.998 | 0.971 | |
| 3 | 2 | – | – | – | – | – | – | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siakou, M.; Bruggeman, A.; Eliades, M.; Djuma, H.; Kyriacou, M.C.; Moriana, A. Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing. Agronomy 2022, 12, 879. https://doi.org/10.3390/agronomy12040879
Siakou M, Bruggeman A, Eliades M, Djuma H, Kyriacou MC, Moriana A. Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing. Agronomy. 2022; 12(4):879. https://doi.org/10.3390/agronomy12040879
Chicago/Turabian StyleSiakou, Melpomeni, Adriana Bruggeman, Marinos Eliades, Hakan Djuma, Marios C. Kyriacou, and Alfonso Moriana. 2022. "Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing" Agronomy 12, no. 4: 879. https://doi.org/10.3390/agronomy12040879
APA StyleSiakou, M., Bruggeman, A., Eliades, M., Djuma, H., Kyriacou, M. C., & Moriana, A. (2022). Phenology, Morphology and Physiology Responses of Deficit Irrigated ‘Koroneiki’ Olive Trees as Affected by Environmental Conditions and Alternate Bearing. Agronomy, 12(4), 879. https://doi.org/10.3390/agronomy12040879