
Chemical Composition of Plant Residues Regulates Soil Organic Carbon Turnover in Typical Soils with Contrasting Textures in Northeast China Plain

Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied Soil and Crop Residues
2.2. Aerobic Soil Incubation and Measurement of C Mineralization
2.3. Analyses of Soil and Crop Residue Samples
2.4. Statistical Analysis
3. Results
3.1. Chemical Characteristics of Different Parts of Maize and Soybean Residues
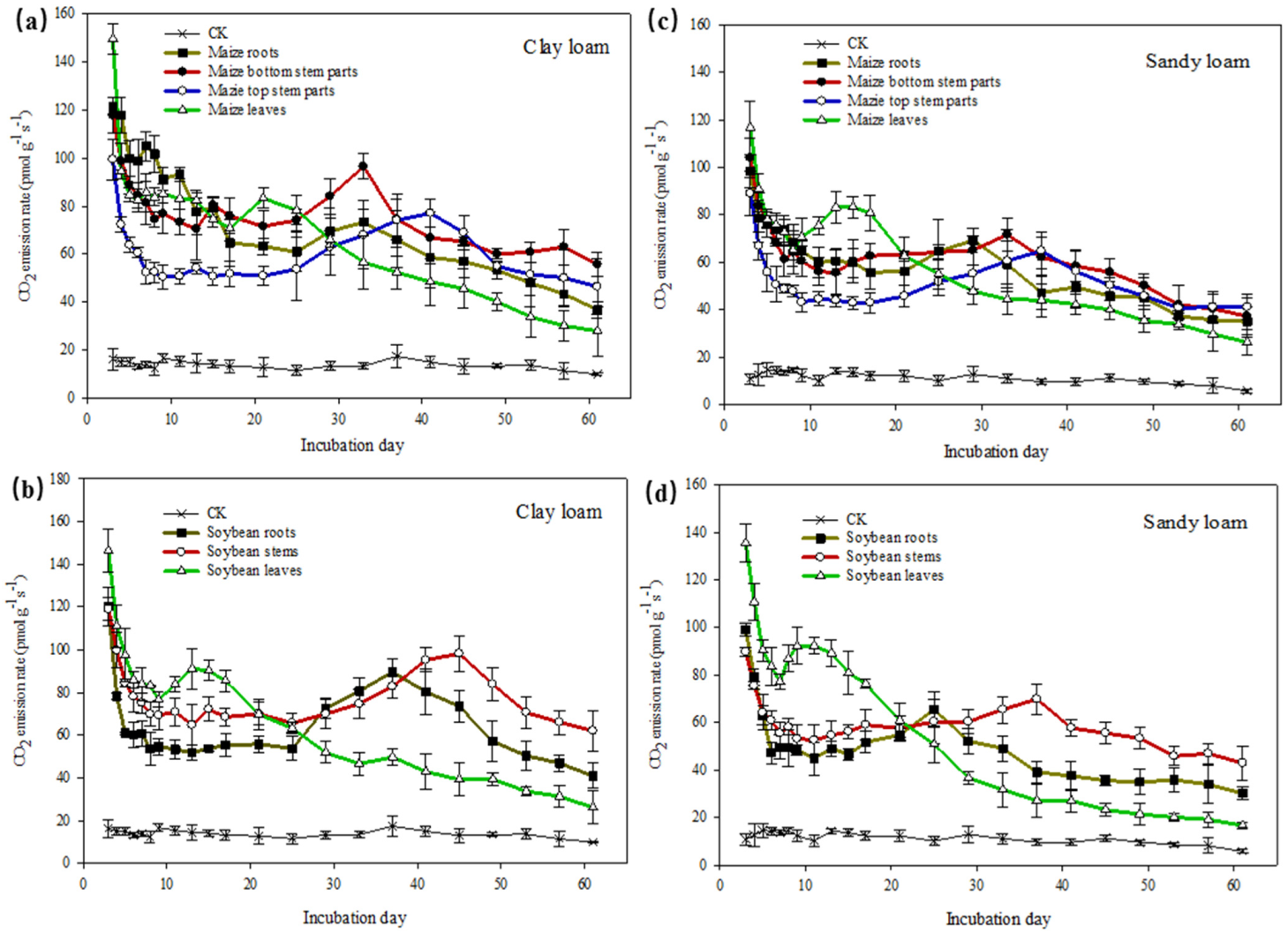
3.2. Soil CO2 Emission and Carbon Mineralization Model Fit in the Two Soils
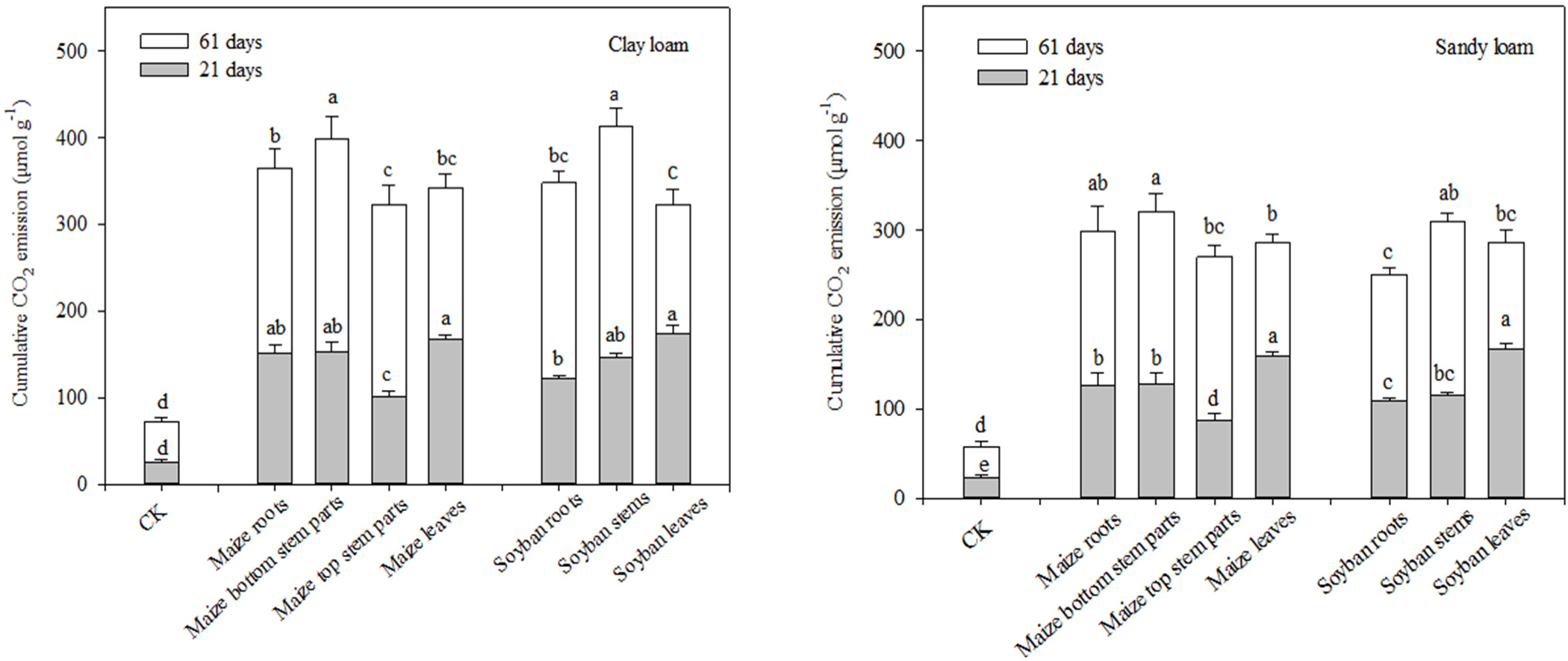
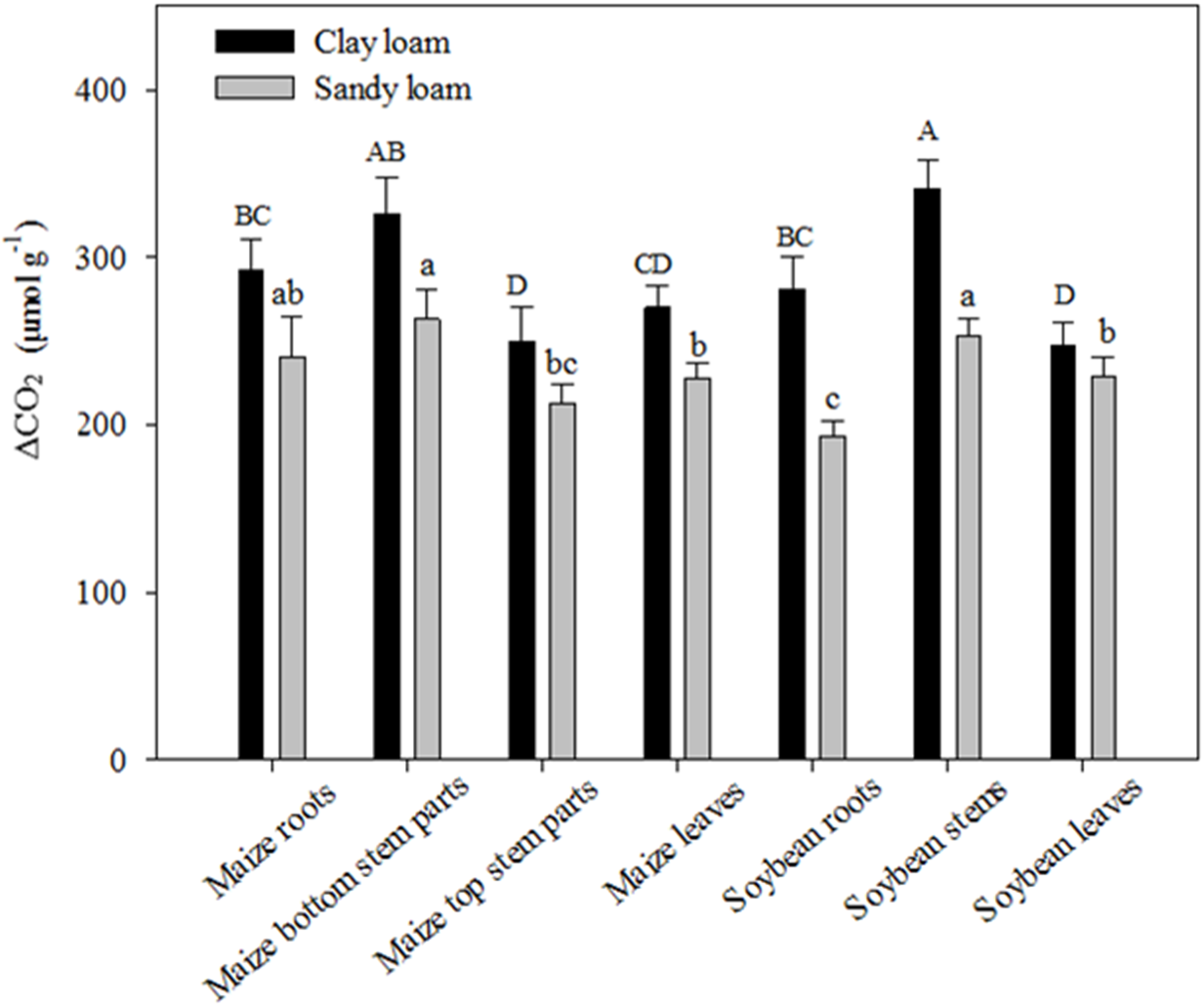
3.3. Effects of Different Residue Types on C Sequestration and Biochemical Properties of the Two Soils
3.4. C Mineralization in Relation to Chemical Composition of Crop Residues
4. Discussion
4.1. Residue Biochemistry and C Mineralization in the Two Soils
4.2. Contribution of Various Residue Types to C Emission and SOC Accumulation in the Two Soils
4.3. Soil Biochemical Properties and N Availability as Influenced by Various Residue Types in the Two Soils
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zech, W.; Senesi, N.; Guggenberger, G.; Kaiser, K.; Lehmann, J.; Miano, T.M.; Miltnera, A.; Schroth, G. Factors controlling humification and mineralization of soil organic matter in the tropics. Geoderma 1997, 79, 117–161. [Google Scholar] [CrossRef]
- Khasanah, N.; Noordwijk, M.V.; Ningsih, H.; Rahayu, S. Carbon neutral? No change in mineral soil carbon stock under oil palm plantations derived from forest or non-forest in indonesia. Agric. Ecosyst. Environ. 2015, 211, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Shar, A.G.; Li, S.; Chen, Y.; Shi, J.; Zhang, X.; Tian, X. Effect of straw return mode on soil aggregation and aggregate carbon content in an annual maize-wheat double cropping system. Soil Till. Res. 2018, 175, 178–186. [Google Scholar] [CrossRef]
- Davidson, E.A. Soil carbon in a beer can. Nat. Geosci. 2015, 8, 748–749. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Janzen, H.; Ellert, B.H.; Helgason, B.L.; Qian, B.D.; Zebarth, B.J.; Anger, D.A.; Beyaert, R.P.; Drury, C.F.; Duguid, S.D. Litter decay controlled by temperature, not soil properties, affecting future soil carbon. Glob. Change Biol. 2017, 23, 1725–1734. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Ghimire, R.; Machado, S.; Bista, P. Decline in soil organic carbon and nitrogen limits yield in wheat-fallow systems. Plant Soil 2018, 422, 423–435. [Google Scholar] [CrossRef]
- Fan, R.Q.; Du, J.J.; Liang, A.Z.; Lou, J.; Li, J.Y. Carbon sequestration in aggregates from native and cultivated soils as affected by soil stoichiometry. Biol. Fertil. Soils 2020, 56, 1109–1120. [Google Scholar] [CrossRef]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; Passioura, J.B.; Batten, G.D.; Blanchard, C.; Kirkegaard, J.A. Nutrient availability limits carbon sequestration in arable soils. Soil Biol. Biochem. 2014, 68, 402–409. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kuzyakov, Y.; Heitkamp, F. Decrease of soil organic matter stabilization with increasing inputs: Mechanisms and controls. Geoderma 2017, 304, 76–82. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2004, 7, 314–320. [Google Scholar] [CrossRef]
- Aye, N.S.; Butterly, C.R.; Sale, P.W.G.; Tang, C. Interactive effects of initial pH and nitrogen status on soil organic carbon priming by glucose and lignocellulose. Soil Biol. Biochem. 2018, 123, 33–44. [Google Scholar] [CrossRef]
- Almagro, M.; Ruiz, N.A.; Diaz, P.E.; Albaladejo, J.; Martínez, M.M. Plant residue chemical quality modulates the soil microbial response related to decomposition and soil organic carbon and nitrogen stabilization in a rainfed Mediterranean agroecosystem. Soil Biol. Biochem. 2021, 156, 108198. [Google Scholar] [CrossRef]
- Ge, N.N.; Wei, X.R.; Wang, X.; Liu, X.T.; Shao, M.G.; Jia, X.X.; Li, X.Z.; Zhang, Q.Y. Soil texture determines the distribution of aggregate-associated carbon, nitrogen and phosphorous under two contrasting land use types in the Loess Plateau. Catena 2019, 172, 148–157. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil. Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Chivenge, P.; Vanlauwe, B.; Gentile, R.; Six, J. Comparison of organic versus mineral resource effects on short-term aggregate carbon and nitrogen dynamics in a sandy soil versus a fine textured soil. Agric. Ecosyst. Environ. 2011, 140, 361–371. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Urbanski, L.; Hobley, E.; Lang, B.; von Lützow, M.; Marin-Spiotta, E.; van Wesemael, B.; Rabot, E.; Ließ, M.; Garcia-Franco, N.; et al. Soil organic carbon storage as a key function of soils—A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Fang, M.; Motavalli, P.P.; Kremer, R.J.; Nelson, K.A. Assessing changes in soil microbial communities and carbon mineralization in Bt and non-Bt corn residue-amended soils. Appl. Soil Ecol. 2007, 37, 150–160. [Google Scholar] [CrossRef]
- Fontaine, S.; Henault, C.; Aamor, A.; Bdioui, N.; Bloor, J.M.G.; Maire, V.; Mary, B.; Revaillot, S.; Maron, P.A. Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect. Soil Biol. Biochem. 2011, 43, 86–96. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Ellis, B.E.; Grayston, S.J. Plant residue chemistry impacts soil processes and microbial community structure: A study with Arabidopsis thaliana cell wall mutants. Appl. Soil Ecol. 2012, 60, 84–91. [Google Scholar] [CrossRef]
- Xu, Y.D.; Ding, F.; Gao, X.D.; Wang, Y.; Li, M.; Wang, J.K. Mineralization of plant residues and native soil carbon as affected by soil fertility and residue type. J. Soil Sediment 2018, 19, 1407–1415. [Google Scholar] [CrossRef]
- Guenet, B.; Neill, C.; Bardoux, G.; Abbadie, L. Is there a linear relationship between priming effect intensity and the amount of organic matter input? Appl. Soil Ecol. 2010, 46, 436–442. [Google Scholar] [CrossRef]
- Gärdenäs, A.I.; Ågren, G.I.; Bird, J.A.; Clarholm, M.; Hallin, S.; Ineson, P.; Kätterer, T.; Knicker, H.; Nilsson, S.I.; Näsholm, T.; et al. Knowledge gaps in soil carbon and nitrogen interactions–From molecular to global scale. Soil Biol. Biochem. 2011, 43, 702–717. [Google Scholar] [CrossRef]
- Jensen, L.S.; Salo, T.; Palmason, F.; Breland, T.A.; Henriksen, T.M.; Stenberg, B.; Pedersen, A.; Lundstr€om, C.; Esala, M. Influence of biochemical quality on C and N mineralisation from a broad variety of plant materials in soil. Plant Soil 2005, 273, 307–326. [Google Scholar] [CrossRef]
- Bonanomi, G.; Incerti, G.; Antignani, V.; Capodilupo, M.; Mazzoleni, S. Decomposition and nutrient dynamics in mixed litter of Mediterranean species. Plant Soil 2010, 331, 481–496. [Google Scholar] [CrossRef]
- Thippayarugs, S.; Toomsan, B.; Vityakon, P.; Limpinuntana, V.; Patanothai, A.; Cadisch, G. Interactions in decomposition and N mineralization between tropical legume residue components. Agrofor. Syst. 2007, 72, 137–148. [Google Scholar] [CrossRef]
- Redin, M.; Recous, S.; Aita, C.; Dietrich, G.; Skolaude, A.C.; Ludke, W.H.; Schmatz, R.; Giacomini, S.J. How the chemical composition and heterogeneity of crop residue mixtures decomposing at the soil surface affects C and N mineralization. Soil Biol. Biochem. 2014, 78, 65–75. [Google Scholar] [CrossRef]
- Heim, A.; Schmidt, M.W.I. Lignin turnover in arable soil and grassland analysed with two different labelling approaches. Eur. J. Soil Sci. 2007, 58, 599–608. [Google Scholar] [CrossRef]
- Abiven, S.; Heim, A.; Schmidt, M.W.I. Lignin content and chemical characteristics in maize and wheat vary between plant organs and growth stages: Consequences for assessing lignin dynamics in soil. Plant Soil 2011, 343, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Puget, P.; Drinkwater, L.E. Short-term dynamics of root- and shoot-derived carbon from a leguminous green manure. Soil Sci. Soc. Am. J. 2001, 65, 771–779. [Google Scholar] [CrossRef]
- Lu, Y.; Watanabe, A.; Kimura, M. Carbon dynamics of rhizodeposits, root- and shoot-residues in a rice soil. Soil Biol. Biochem. 2003, 35, 1223–1230. [Google Scholar] [CrossRef]
- Bertrand, I.; Chabbert, B.; Kurek, B.; Recous, S. Can the biochemical features and histology of wheat residues explain their decomposition in soil? Plant Soil 2006, 281, 291–307. [Google Scholar] [CrossRef]
- Abiven, S.; Recous, S.; Reyes, V.; Oliver, R. Mineralisation of C and N from root, stem and leaf residues in soil and role of their biochemical quality. Biol. Fert. Soils 2005, 42, 119–128. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Coq, S.; Barantal, S.; Tanya, I. Leaf traits and decomposition in tropical rainforests: Revisiting some commonly held views and towards a new hypothesis. New Phytol. 2011, 189, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Xu, Y.H.; Chen, Z.M.; Fontaine, S.; Wang, W.J.; Luo, J.F.; Fan, J.L.; Ding, W.X. Dominant effects of organic carbon chemistry on decomposition dynamics of crop residues in a Mollisol. Soil Biol. Biochem. 2017, 115, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.L.; Han, X.; Wang, Y.Q.; Gu, R.S.; Li, Y.C.; Wang, D.Y.; Yun, A.P.; Guo, L.P. Changes in soil organic carbon concentration, chemical composition, and aggregate stability as influenced by tillage systems in the semi-arid and semi-humid area of North China. Can. J. Soil Sci. 2018, 98, 91–102. [Google Scholar] [CrossRef]
- Adnan, M.; Xu, H.; Atizaz, S.; Syed, A.A.S.; Abrar, M.M.; Maitlo, A.A.; Kubar, K.A.; Saeed, Q.; Kamran, M.; Naveed, M.; et al. Long-term fertilization alters chemical composition and stability of aggregate-associated organic carbon in a Chinese red soil: Evidence from aggregate fractionation, C mineralization, and 13C NMR analyses. J. Soil Sediments 2021, 21, 2483–2496. [Google Scholar]
- Bonanomi, G.; Incerti, G.; Barile, E.; Capodilupo, M.; Antignani, V.; Mingo, A.; Lanzotti, V.; Scala, F.; Mazzoleni, S. Phytotoxicity, not nitrogen immobilization, explains plant litter inhibitory effects: Evidence from solid-state 13C-NMR spectroscopy. New Phytol. 2011, 191, 1018–1030. [Google Scholar] [CrossRef] [Green Version]
- Mathers, N.J.; Jolota, R.K.; Dalal, R.C.; Boyd, S.E. 13 C-NMR analysis of decomposing litter and fine roots in the semi-arid Mulga Lands of southern Queensland. Soil Biol. Biochem. 2007, 39, 993–1006. [Google Scholar] [CrossRef]
- Preston, C.M.; Nault, J.R.; Trofymow, J.A. Chemical changes during 6 years of decomposition of 11 litters in some Canadian forest sites. Part 2. 13 C abundance, solid-state 13 C NMR spectroscopy and the meaning of “lignin”. Ecosystems 2009, 12, 1078–1102. [Google Scholar] [CrossRef]
- Ono, K.; Hiradate, S.; Morita, S.; Ohse, K.; Hirai, K. Humification processes of needle litters on forest floors in Japanese cedar (Cryptomeria japonica) and Hinoki cypress (Chamaecyparis obtusa) plantations in Japan. Plant Soil 2011, 338, 171–181. [Google Scholar] [CrossRef]
- Liang, A.Z.; Zhang, X.P.; Yang, X.M.; McLaughlin, N.B.; Shen, Y.; Li, W.F. Estimation of total erosion in cultivated Black soils in northeast China from vertical profiles of soil organic carbon. Eur. J. Soil Sci. 2009, 60, 223–229. [Google Scholar] [CrossRef]
- Shi, X.H.; Zhang, X.P.; Yang, X.M.; Drury, C.F.; McLaughlin, N.B.; Liang, A.Z.; Fan, R.Q.; Jia, S.X. Contribution of winter soil respiration to annual soil CO2 emission in a Mollisol under different tillage practices in northeast China. Global Biogeochem. Cycles 2012, 26, GB2007. [Google Scholar] [CrossRef]
- Li, L.; Han, X.Z.; You, M.Y.; Yuan, Y.R.; Ding, X.L.; Qiao, Y.F. Carbon and nitrogen mineralization patterns of two contrasting crop residues in a Mollisol: Effects of residue type and placement in soils. Eur. J. Soil Sci. 2013, 54, 1–6. [Google Scholar] [CrossRef]
- Fan, R.Q.; Yang, X.M.; Drury, C.F.; Guo, X.B.; Zhang, X.P. Distribution and stability of organic carbon in soil aggregate external and internal layers under three different land-use systems. Soil Sci. Soc. Am. J. 2013, 77, 1625–1635. [Google Scholar] [CrossRef]
- Zhang, Z.D.; Yang, X.M.; Drury, C.F.; Reynolds, W.D.; Zhao, L.P. Mineralization of active soil organic carbon in particle size fractions of a Brookston clay soil under no-tillage and mouldboard plough tillage. Can. J. Soil Sci. 2010, 90, 551–557. [Google Scholar] [CrossRef]
- Fang, Y.; Singh, B.; Singh, B.P. Effect of temperature on biochar priming effects and its stability in soils. Soil Biol. Biochem. 2015, 80, 136–145. [Google Scholar] [CrossRef]
- Molina, J.A.E.; Clapp, C.E.; Larson, W.E. Potentially mineralizable nitrogen in soil: The simple exponential model does not apply to the first 12 weeks of incubation. Soil Sci. Soc. Am. J. 1980, 44, 442–443. [Google Scholar] [CrossRef] [Green Version]
- Gregorich, E.G.; Kachanoski, R.G.; Voroney, R.P. Carbon mineralization in soil size fractions after various amounts of aggregate disruption. Eur. J. Soil Sci. 1989, 40, 649–659. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 7, 3584–3597. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of microbial biomass carbon and nitrogen in soil. In Advances in Nitrogen Cycling in Agricultural Ecosystems; CAB International: Wallingford, UK, 1988; pp. 368–386. [Google Scholar]
- Kjeldahl, J. A new method for the determination of nitrogen in organic matter. Z. Anal. Chem. 1883, 22, 366. [Google Scholar] [CrossRef] [Green Version]
- Baumann, K.; Marschner, P.; Smernik, R.J.; Baldock, J.A. Residue chemistry and microbial community structure during decomposition of eucalypt, wheat and vetch residues. Soil Biol. Biochem. 2009, 41, 1966–1975. [Google Scholar] [CrossRef]
- Liu, S.Y.; Fan, R.Q.; Yang, X.M.; Zhang, Z.H.; Zhang, X.P.; Liang, A.Z. Decomposition of maize stover varies with maize type and stover management strategies: A microcosm study on a Black soil (Mollisol) in northeast China. J. Environ. Manag. 2019, 234, 226–236. [Google Scholar] [CrossRef]
- Dias, T.; Oakley, S.; Alarcón-Gutiérrez, E.; Ziarelli, F.; Trindade, H.; Martins-Loução, M.A.; Sheppard, L.; Ostle, N.; Cruz, C. N-driven changes in a plant community affect leaf-litter traits and may delay organic matter decomposition in a Mediterranean maquis. Soil Biol. Biochem. 2013, 58, 163–171. [Google Scholar] [CrossRef]
- Ma, Q.; Watanabe, T.; Zheng, J.S.; Funakawa, S. Interactive effects of crop residue quality and nitrogen fertilization on soil organic carbon priming in agricultural soils. J. Soils Sediments 2020, 21, 83–95. [Google Scholar] [CrossRef]
- Li, J.Y.; Zhang, Q.C.; Yong, L.; Liu, Y.M.; Xu, J.M.; Di, H.J. Effects of long-term mowing on the fractions and chemical composition of soil organic matter in a semiarid grassland. Biogeosciences 2017, 14, 2685–2696. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Baldock, J.A.; Dalal, R.C.; Moody, P.W. Decomposition dynamics of plant materials in relation to nitrogen availability and biochemistry determined by NMR and wet-chemical analysis. Soil Biol. Biochem. 2004, 36, 2045–2058. [Google Scholar] [CrossRef]
- Xu, Y.; Ge, Y.; Song, J.; Rensing, C. Assembly of root-associated microbial community of typical rice cultivars in different soil types. Biol. Fert. Soils 2020, 56, 249–260. [Google Scholar] [CrossRef]
- Xu, Q.; Jin, J.; Wang, X.J.; Armstrong, R.; Tang, C.X. Susceptibility of soil organic carbon to priming after long-term CO2 fumigation is mediated by soil texture. Sci. Total Environ. 2019, 657, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Schmatz, R.; Recous, S.; Aita, C.; Tahir, M.M.; Schu, A.L.; Chaves, B.; Giacomini, S.J. Crop residue quality and soil type influence the priming effect but not the fate of crop residue C. Plant Soil 2017, 414, 229–245. [Google Scholar] [CrossRef]
- Yang, Y.; Mohammat, A.; Feng, J.; Zhou, R.; Fang, J. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry 2007, 84, 131–141. [Google Scholar] [CrossRef]
- Rui, J.; Peng, J.; Lu, Y. Succession of bacterial populations during plant residue decomposition in rice field soil. Appl. Environ. Microbiol. 2009, 75, 4879–4886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gul, S.; Whalen, J. Plant life history and residue chemistry influences emissions of CO2 and N2O from soil perspectives for genetically modified cell wall mutants. Plant Sci. 2013, 32, 344–368. [Google Scholar]
- Kleber, M.; Sollins, P.; Sutton, R. A conceptual model of organo-mineral interactions in soils: Self-assembly of organic molecular fragments into zonal structures on mineral surfaces. Biogeochemistry 2007, 85, 9–24. [Google Scholar] [CrossRef]
- Fan, R.Q.; Yang, X.M.; Drury, C.F.; Zhang, Z.H. Curve-fitting techniques improve the mid-infrared analysis of soil organic carbon: A case study for Brookston clay loam particle-size fractions. Sci. Rep. 2018, 8, 12174. [Google Scholar] [CrossRef] [PubMed]
- Rasse, D.P.; Rumpel, C.; Dignac, M.F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 2005, 269, 341–356. [Google Scholar] [CrossRef]
- Heitkamp, F.; Wendland, M.; Offenberger, K.; Gerold, G. Implications of input estimation, residue quality and carbon saturation on the predictive power of the Rothamsted Carbon Model. Geoderma 2012, 170, 168–175. [Google Scholar] [CrossRef]
- Freschet, G.T.; Cornwell, W.K.; Wardle, D.A.; Elumeeva, T.G.; Liu, W.; Jackson, B.G.; Onipchenko, V.G.; Soudzilovskaia, N.A.; Tao, J.; Cornelissen, J.H.C. Linking litter decomposition of above- and below-ground organs to plant–soil feedbacks worldwide. J. Ecol. 2013, 101, 943–952. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Bol, R. Sources and mechanisms of priming effect induced in two grassland soils amended with slurry and sugar. Soil Biol. Biochem. 2006, 38, 747–758. [Google Scholar] [CrossRef]
- Yadav, R.K.; Purakayastha, T.J.; Khan, M.A.; Kaushik, S.C. Long-term impact of manuring and fertilization on enrichment, stability and quality of organic carbon in Inceptisol under two potato-based cropping systems. Sci. Total Environ. 2017, 609, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.; Scheer, C.; Rowlings, D.; Conant, R.T.; Cotrufo, M.F.; Grace, P. Amount and incorporation of plant residue inputs modify residue stabilisation dynamics in soil organic matter fractions. Agric. Ecosyst. Environ. 2018, 256, 82–91. [Google Scholar] [CrossRef]
- Li, L.; Xia, Z.B.; Ye, R.Z.; Doane, T.A.; Horwath, W.R. Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability. Soil Biol. Biochem. 2018, 119, 41–49. [Google Scholar] [CrossRef]
- Begum, N.; Guppy, C.; Herridge, D.; Schwenke, G. Influence of source and quality of plant residues on emissions of N2O and CO2 from a fertile, acidic Black Vertisol. Biol. Fertil. Soils 2014, 50, 499–506. [Google Scholar] [CrossRef]
- Köbke, S.; Senbayram, M.; Pfeiffer, B.; Nacke, H.; Dittert, K. Post-harvest N2O and CO2 emissions related to plant residue incorporation of oilseed rape and barley straw depend on soil NO3-content. Soil Till. Res. 2018, 179, 105–113. [Google Scholar] [CrossRef]
- Huang, J.S.; Liu, W.X.; Yang, S.; Yang, L.; Peng, Z.Y.; Deng, M.F.; Xu, S.; Zhang, B.B.; Ahirwal, J.; Li, L.L. Plant carbon inputs through shoot, root, and mycorrhizal pathways affect soil organic carbon turnover differently. Soil Biol. Biochem. 2021, 160, 108322. [Google Scholar] [CrossRef]
- Ros, G.H.; Hoffland, E.; Temminghoff, E.J.M. Dynamics of dissolved and extractable organic nitrogen upon soil amendment with crop residues. Soil Biol. Biochem. 2010, 42, 2094–2101. [Google Scholar] [CrossRef]
- Li, J.; Zhao, B.Q.; Li, X.Y.; Hwat, B.S. Effects of long-term combined application of organic and mineral fertilizers on soil microbiological properties and soil fertility. Sci. Agric. Sin. 2008, 41, 144–152. (In Chinese) [Google Scholar]
- Yu, Z.; Chen, L.; Pan, S.; Li, Y.; Kuzyakov, Y.; Xu, J.; Brookes, P.; Luo, Y. Feedstock determines biochar-induced soil priming effects by stimulating the activity of specific microorganisms. Soil Use Manag. 2018, 69, 521–534. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Type | Soil Texture (%) | pH (Water) | Organic C (g kg−1) | Total N (g kg−1) | C/N | Available P (mg kg−1) | Available K (mg kg−1) | ||
|---|---|---|---|---|---|---|---|---|---|
| Sand | Silt | Clay | |||||||
| Clay loam | 31.7 | 26.4 | 41.9 | 5.4 | 17.6 | 1.7 | 10.4 | 15.5 | 110.3 |
| Sandy loam | 67.0 | 7.7 | 25.3 | 5.5 | 8.0 | 1.2 | 7.2 | 11.2 | 87.3 |
| Residue type | Residue Part | Carbonyl C | Phenolic C | Aromatic C | di-O-alkyl C | O-alkyl C | Methoxyl C | Alkyl C | Lignin | Total C | Total N | Aromaticity | Lignin/N | C/N | HB/HI | A/OA |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (g kg−1) | (%) | |||||||||||||||
| Maize | Roots | 19.9 c † | 22.9 b | 46.8 b | 55.6 c | 186.1 d | 25.3 b | 35.6 c | 110.3 b | 392 b | 9.6 b | 17.8 a | 12 b | 41 d | 0.45 bc | 0.19 c |
| Bottom stem parts | 10.7 d | 17.9 c | 40.4 c | 75.7 a | 239.0 b | 18.3 cd | 25.6 d | 89.8 c | 438 a | 6.7 c | 13.3 b | 13 b | 65 c | 0.31 d | 0.11 d | |
| Top stem parts | 11.5 d | 17.8 c | 39.0 c | 77.3 a | 264.4 a | 16.8 d | 11.0 e | 88.0 c | 438 a | 4.0 d | 13.0 b | 22 a | 108 a | 0.24 e | 0.04 e | |
| Leaves | 32.7 a | 17.6 c | 31.8 d | 56.5 c | 194.1 c | 29.6 b | 61.5 a | 76.4 d | 424 a | 10.2 b | 13.6 b | 8 c | 41 d | 0.50 b | 0.32 b | |
| Soybean | Roots | 21.1 bc | 27.9 a | 54.2 a | 59.5 c | 251.1 ab | 34.7 a | 23.1 d | 127.9 a | 450 a | 6.5 c | 18.3 a | 20 a | 69 c | 0.42 c | 0.09 d |
| Stems | 19.9 c | 21.1 b | 41.9 c | 67.5 b | 238.0 b | 26.9 b | 32.4 c | 97.4 bc | 448 a | 4.9 d | 14.1 b | 20 a | 92 b | 0.38 c | 0.14 cd | |
| Leaves | 24.9 b | 13.2 d | 27.5 e | 36.0 d | 123.8 d | 20.3 c | 50.4 b | 63.5 e | 307 b | 13.8 a | 13.8 b | 5 d | 21 e | 0.60 a | 0.41 a | |
| Model Parameters † | Maize | Soybean | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Roots | Bottom Stem Parts | Top Stem Parts | Leaves | Roots | Stems | Leaves | ||||||||
| Clay | Sandy | Clay | Sandy | Clay | Sandy | Clay | Sandy | Clay | Sandy | Clay | Sandy | Clay | Sandy | |
| Ca (µmol g−1) | 30.41 b ‡ | 35.23 a | 36.90 b | 45.49 a | 16.01 b | 20.18 a | 40.02 b | 45.19 a | 24.79 b | 30.55 a | 35.59 b | 40.01 a | 41.13 b | 45.58 a |
| ka (µmol g−1 d−1) | 0.387 b | 0.718 a | 0.901 b | 1.074 a | 0.575 b | 0.912 a | 0.322 b | 0.797 a | 0.029 b | 0.062 a | 0. 045 b | 0.058 a | 0.058 b | 0.089 a |
| Cs (µmol g−1) | 37.32 a | 11.73 b | 29.94 a | 8.910 b | 57.18 a | 40.15 b | 15.07 a | 7.786 b | 39.37 b | 18.79 a | 29.36 b | 14.74 b | 11.16 a | 6.032 b |
| ks (µmol g−1 d−1) | 0.010 a | 0.008 a | 0.006 a | 0.005 a | 0.0018 a | 0.0017 a | 0.022 a | 0.019 a | 0.007 a | 0.005 b | 0.008 a | 0.007 b | 0.021 a | 0.012 b |
| T(1/2)a (d) | 5.655 b | 9.791 a | 4.645 b | 8.769 a | 6.760 b | 10.25 a | 3.87 b | 6.153 a | 6.118 b | 8.390 a | 5.395 b | 6.540 a | 3.788 b | 4.935 a |
| T(1/2)s (d) | 134.6 a | 119.3 b | 88.6 a | 75.50 b | 207.7 a | 185.1 b | 36.48 a | 31.01 a | 138.6 a | 99.02 b | 99.02 a | 80.64 b | 57.76 a | 33.01 b |
| R2 | 0.999 | 0.999 | 0.999 | 0.999 | 0.995 | 0.998 | 0.999 | 0.999 | 0.992 | 0.998 | 0.994 | 0.999 | 0.999 | 0.999 |
| Residue Type | Residue Part | SOC (g·kg−1) | MBC (mg·kg−1) | MBN (mg·kg−1) | MBC/MBN | Available N (mg·kg−1) |
|---|---|---|---|---|---|---|
| For clay loam | ||||||
| Maize | Blank control | 17.61 ab † | 91.01 f | 7.64 d | 11.93 a | 45.41 a |
| Roots | 17.15 ab | 186.91 cd | 25.4 bc | 7.36 cd | 14.61 bc | |
| Bottom stem parts | 17.21 ab | 190.12 d | 28.81 b | 6.61 de | 11.77 c | |
| Top stem parts | 18.12 a | 173.78 e | 21.79 c | 7.97 cd | 8.31 d | |
| Leaves | 17.60 ab | 221.74 b | 34.33 a | 6.46 e | 15.33 b | |
| Soybean | Roots | 17.93 ab | 200.34 c | 20.14 c | 9.96 b | 8.44 d |
| Stems | 17.13 ab | 214.31 bc | 24.60 bc | 8.71 c | 8.40 d | |
| Leaves | 16.98 b | 252.30 a | 32.62 a | 7.74 d | 19.91 b | |
| For sandy loam | ||||||
| Maize | Blank control | 8.03 cd | 56.30 e | 5.23 e | 10.8 a | 39.53 a |
| Roots | 8.37 c | 165.79 c | 24.24 bc | 6.91 cd | 12.5 b | |
| Bottom stem parts | 8.52 c | 191.63 b | 27.53 b | 6.96 cd | 7.82 c | |
| Top stem parts | 9.37 ab | 180.93 b | 23.23 c | 7.80 c | 5.34 d | |
| Leaves | 8.63 c | 222.08 a | 31.91 a | 6.53 d | 13.9 b | |
| Soybean | Roots | 9.46 a | 119.04 d | 15.76 d | 7.53 c | 7.17 c |
| Stems | 8.72 bc | 179.07 b | 20.11 c | 8.91 b | 7.02 c | |
| Leaves | 7.74 d | 180.82 b | 30.50 a | 7.80 c | 15.12 b | |
| DF | Cumulative CO2 | Ca | Cs | T(1/2)a | T(1/2)s | SOC | MBC | MBN | Available N | ΔCO2 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | F | P | F | P | F | P | F | P | F | P | F | P | ||
| Residue type | 6 | 5.46 | * | 10.12 | * | 4.34 | * | 39.81 | *** | 13.87 | ** | 9.39 | * | 7.52 | * | 33.31 | ** | 34.36 | *** | 4.44 | * |
| Soil type | 1 | 45.42 | *** | 32.04 | ** | 16.86 | ** | 50.02 | *** | 19.29 | ** | 5463 | *** | 16.39 | ** | 11.10 | * | 23.24 | ** | 21.47 | ** |
| Residue type × Soil type | 6 | 3.25 | * | 7.06 | * | 3.99 | * | 19.36 | ** | 5.240 | * | 15.76 | ** | 2.52 | ns | 3.59 | * | 14.37 | ** | 3.11 | * |
| Residue Chemistry | Cumulative CO2 Emissionday21 | ΔCO2 | Ca | Cs | T(1/2)a | T(1/2)s | SOC | MBC | MBN | Available N |
|---|---|---|---|---|---|---|---|---|---|---|
| For clay loam | ||||||||||
| C | −0. 597 | 0.516 | −0.457 | 0.568 | 0.560 | 0.392 | 0.550 | −0.692 * | −0.551 | −0.785 * |
| N | 0.807 *† | −0.440 | 0.669 | −0.784 * | −0.773 * | −0.675 | −0.554 | 0.749 * | 0.749 * | 0.933 ** |
| Lignin | −0.540 | 0.357 | −0.519 | 0.563 | 0.686 | 0.508 | 0.336 | −0.568 | −0.800 * | −0.664 |
| Alkyl C | 0.879 * | 0.165 | 0.825 * | −0.903 ** | −0.873 * | −0.809 * | −0.490 | 0.774 * | 0.868 * | 0.781 * |
| Aromatic C | −0.585 | −0.386 | −0.554 | 0.616 | 0.772 | 0.558 | 0.341 | −0.623 | −0.834 * | −0.652 |
| Aromaticity | −0.154 | −0.049 | −0.239 | 0.197 | 0.356 | 0.225 | 0.079 | −0.182 | −0.482 | −0.112 |
| Lignin/N | −0.812 * | 0.313 | −0.776 * | 0.834 * | 0.884 * | 0.779 * | 0.587 | −0.681 * | −0.898 * | −0.954 *** |
| C/N | −0.835 * | −0.635 | −0.704 | 0.779 * | 0.766 * | 0.710 | 0.574 | −0.651 | −0.714 | −0.920 ** |
| Alkyl/O-alkyl | 0.871 ** | 0.799 ** | 0.772 * | −0.888 ** | −0.862 * | −0.794 * | −0.718 * | 0.879 * | 0.849 * | 0.925 ** |
| For sandy loam | ||||||||||
| C | −0.597 | −0.038 | −0.387 | 0.427 | 0.519 | 0.435 | 0.819 * | −0.129 | −0.555 | −0.870 * |
| N | 0.893 ** | 0.037 | 0.617 | −0.704 | −0.612 | −0.692 | −0.814 * | 0.233 | 0.670 | 0.962 *** |
| Lignin | −0.540 | −0.319 | −0.472 | 0.235 | 0.574 | 0.443 | 0.632 | −0.792 * | −0.865 * | −0.510 |
| Alkyl C | 0.897 ** | 0.178 | 0.760 * | −0.866 * | −0.776 * | −0.854 * | −0.687 * | 0.536 | 0.696 * | 0.895 ** |
| Aromatic C | −0.585 | −0.280 | −0.496 | 0.279 | 0.627 | 0.495 | 0.647 | −0.793 * | −0.877 ** | −0.581 |
| Aromaticity | −0.154 | −0.406 | −0.235 | −0.104 | 0.291 | 0.139 | 0.217 | −0.580 | −0.587 | 0.041 |
| Lignin/N | −0.982 *** | −0.749 * | −0.759 * | 0.876 * | 0.699 * | 0.810 * | 0.872 ** | −0.741 * | −0.839 * | −0.955 *** |
| C/N | −0.849 * | −0.163 | −0.679 | 0.811 * | 0.525 | 0.732 | 0.775 * | −0.169 | −0.604 | −0.940 ** |
| Alkyl/O-alkyl | 0.860 * | 0.110 | 0.694 * | −0.689 * | −0.807 * | −0.793 * | −0.813 * | 0.429 | 0.729 * | 0.916 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Li, J.; Liang, A.; Duan, Y.; Chen, H.; Yu, Z.; Fan, R.; Liu, H.; Pan, H. Chemical Composition of Plant Residues Regulates Soil Organic Carbon Turnover in Typical Soils with Contrasting Textures in Northeast China Plain. Agronomy 2022, 12, 747. https://doi.org/10.3390/agronomy12030747
Liu S, Li J, Liang A, Duan Y, Chen H, Yu Z, Fan R, Liu H, Pan H. Chemical Composition of Plant Residues Regulates Soil Organic Carbon Turnover in Typical Soils with Contrasting Textures in Northeast China Plain. Agronomy. 2022; 12(3):747. https://doi.org/10.3390/agronomy12030747
Chicago/Turabian StyleLiu, Siyi, Jiangye Li, Aizhen Liang, Yan Duan, Haibin Chen, Zhuyun Yu, Ruqin Fan, Haiyang Liu, and Hong Pan. 2022. "Chemical Composition of Plant Residues Regulates Soil Organic Carbon Turnover in Typical Soils with Contrasting Textures in Northeast China Plain" Agronomy 12, no. 3: 747. https://doi.org/10.3390/agronomy12030747
APA StyleLiu, S., Li, J., Liang, A., Duan, Y., Chen, H., Yu, Z., Fan, R., Liu, H., & Pan, H. (2022). Chemical Composition of Plant Residues Regulates Soil Organic Carbon Turnover in Typical Soils with Contrasting Textures in Northeast China Plain. Agronomy, 12(3), 747. https://doi.org/10.3390/agronomy12030747